

Targeting Lactate: An Emerging Strategy for Macrophage Regulation in Chronic Inflammation and Cancer



Abstract
:1. Introduction
2. Lactate as a Biomarker in Chronic Inflammation and Cancer
2.1. Chronic Inflammation
2.1.1. Rheumatoid Arthritis (RA)
2.1.2. Atherosclerosis
2.1.3. Obesity
2.1.4. Inflammatory Bowel Disease (IBD)
2.1.5. Cancer
Non-Small-Cell Lung Cancer (NSCLC)
Melanoma
Cerebral Glioma
3. Phenotypes and Functions of Macrophage in Chronic Inflammation and Cancer
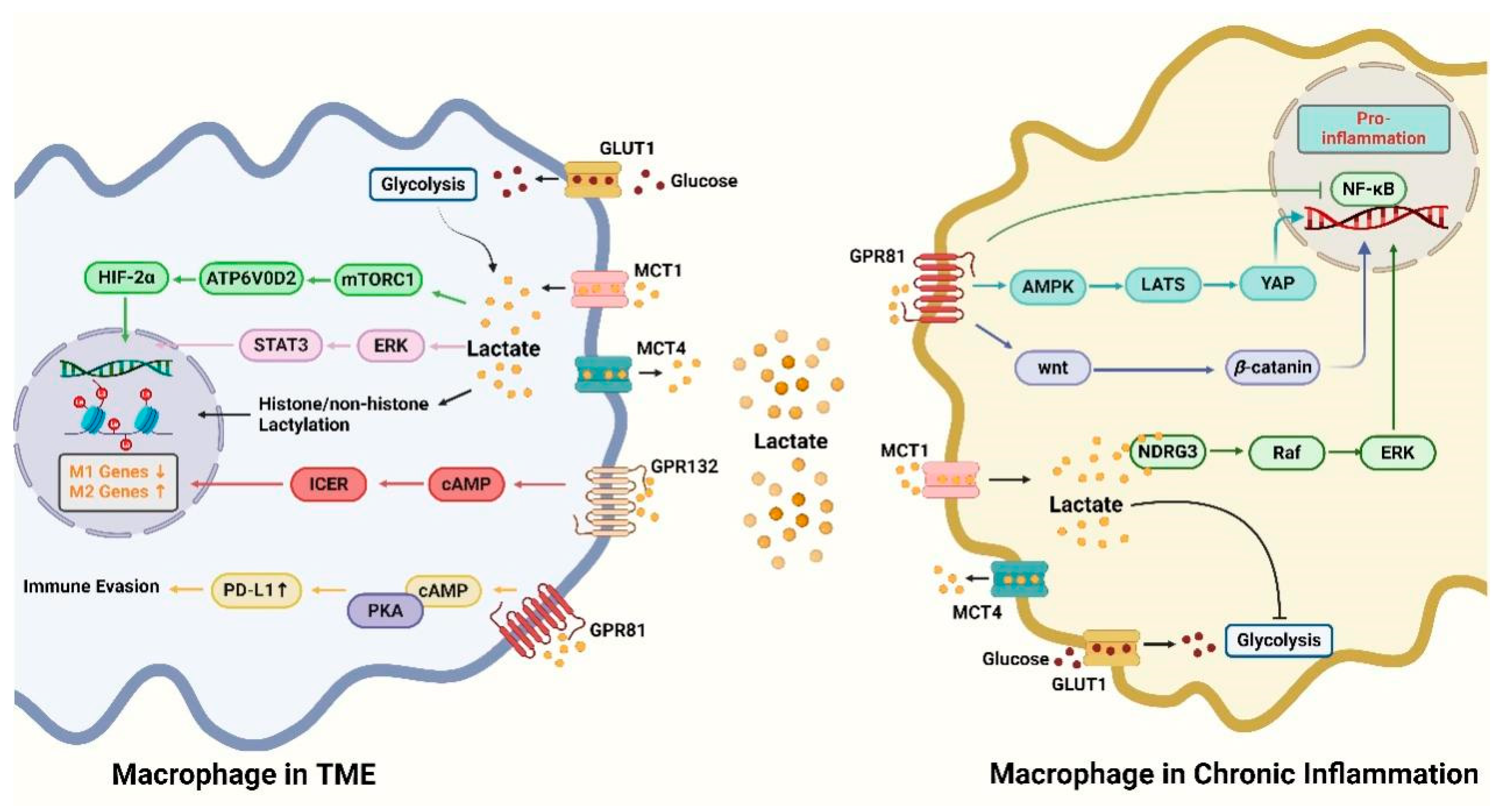
4. Pathways of Lactate Regulation of Macrophages
4.1. GPR81 Signaling Pathway
4.2. GPR132 Signaling Pathway
4.3. mTORC1 Signaling Pathway
4.4. Histone Lactylation Modification
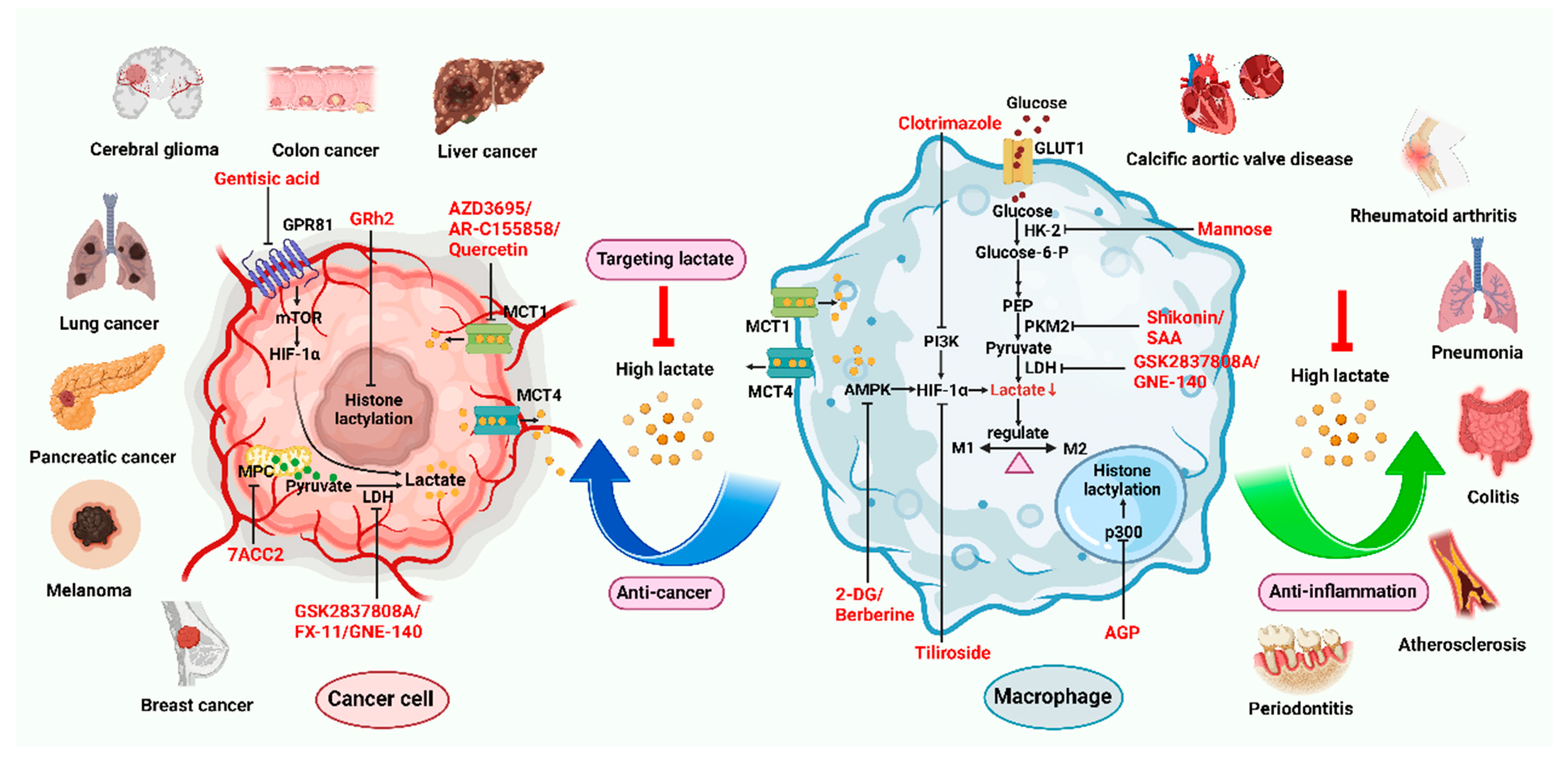
5. Small Molecules Targeting Lactate
5.1. Targeting Lactate to Modulate Macrophages
5.2. Lactate Transporter Inhibitors
5.3. Lactate Metabolism Inhibitors
5.4. Lactate-Related Signaling Pathways Inhibitors
6. Conclusions and Perspective
Author Contributions
Funding
Conflicts of Interest
References
- Coates, A.M.; Joyner, M.J.; Little, J.P.; Jones, A.M.; Gibala, M.J. A Perspective on High-Intensity Interval Training for Performance and Health. Sports Med. 2023, 53, 85–96. [Google Scholar] [CrossRef]
- Certo, M.; Tsai, C.H.; Pucino, V.; Ho, P.C.; Mauro, C. Lactate modulation of immune responses in inflammatory versus tumour microenvironments. Nat. Rev. Immunol. 2021, 21, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef] [PubMed]
- Rabinowitz, J.D.; Enerbäck, S. Lactate: The ugly duckling of energy metabolism. Nat. Metab. 2020, 2, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Muri, J.; Fitzgerald, G.; Gorski, T.; Gianni-Barrera, R.; Masschelein, E.; D’Hulst, G.; Gilardoni, P.; Turiel, G.; Fan, Z.; et al. Endothelial Lactate Controls Muscle Regeneration from Ischemia by Inducing M2-like Macrophage Polarization. Cell Metab. 2020, 31, 1136–1153.E7. [Google Scholar] [CrossRef] [PubMed]
- De-Brito, N.M.; Duncan-Moretti, J.; Da-Costa, H.C.; Saldanha-Gama, R.; Paula-Neto, H.A.; Dorighello, G.G.; Simões, R.L.; Barja-Fidalgo, C. Aerobic glycolysis is a metabolic requirement to maintain the M2-like polarization of tumor-associated macrophages. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118604. [Google Scholar] [CrossRef]
- Li, F.; Sami, A.; Noristani, H.N.; Slattery, K.; Qiu, J.; Groves, T.; Wang, S.; Veerasammy, K.; Chen, Y.X.; Morales, J.; et al. Glial Metabolic Rewiring Promotes Axon Regeneration and Functional Recovery in the Central Nervous System. Cell Metab. 2020, 32, 767–785.e767. [Google Scholar] [CrossRef]
- Roumes, H.; Dumont, U.; Sanchez, S.; Mazuel, L.; Blanc, J.; Raffard, G.; Chateil, J.F.; Pellerin, L.; Bouzier-Sore, A.K. Neuroprotective role of lactate in rat neonatal hypoxia-ischemia. J. Cereb. Blood Flow Metab. 2021, 41, 342–358. [Google Scholar] [CrossRef]
- Veys, K.; Fan, Z.; Ghobrial, M.; Bouché, A.; García-Caballero, M.; Vriens, K.; Conchinha, N.V.; Seuwen, A.; Schlegel, F.; Gorski, T.; et al. Role of the GLUT1 Glucose Transporter in Postnatal CNS Angiogenesis and Blood-Brain Barrier Integrity. Circ. Res. 2020, 127, 466–482. [Google Scholar] [CrossRef]
- Yin, J.; Forn-Cuní, G.; Surendran, A.M.; Lopes-Bastos, B.; Pouliopoulou, N.; Jager, M.J.; Le Dévédec, S.E.; Chen, Q.; Snaar-Jagalska, B.E. Lactate secreted by glycolytic conjunctival melanoma cells attracts and polarizes macrophages to drive angiogenesis in zebrafish xenografts. Angiogenesis 2024. [Google Scholar] [CrossRef]
- Yao, S.; Chai, H.; Tao, T.; Zhang, L.; Yang, X.; Li, X.; Yi, Z.; Wang, Y.; An, J.; Wen, G.; et al. Role of lactate and lactate metabolism in liver diseases (Review). Int. J. Mol. Med. 2024, 54, 59. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, J.; Wang, H.; Huang, J. The role of lactate in cardiovascular diseases. Cell Commun. Signal 2023, 21, 317. [Google Scholar] [CrossRef]
- Cai, Y.; Guo, H.; Han, T.; Wang, H. Lactate: A prospective target for therapeutic intervention in psychiatric disease. Neural Regen. Res. 2024, 19, 1473–1479. [Google Scholar] [CrossRef]
- Bader, J.E.; Voss, K.; Rathmell, J.C. Targeting Metabolism to Improve the Tumor Microenvironment for Cancer Immunotherapy. Mol. Cell 2020, 78, 1019–1033. [Google Scholar] [CrossRef]
- Zhao, H.; Wu, L.; Yan, G.; Chen, Y.; Zhou, M.; Wu, Y.; Li, Y. Inflammation and tumor progression: Signaling pathways and targeted intervention. Signal Transduct. Target. Ther. 2021, 6, 263. [Google Scholar] [CrossRef]
- Nakagawa, T.; Sanchez-Lozada, L.G.; Andres-Hernando, A.; Kojima, H.; Kasahara, M.; Rodriguez-Iturbe, B.; Bjornstad, P.; Lanaspa, M.A.; Johnson, R.J. Endogenous Fructose Metabolism Could Explain the Warburg Effect and the Protection of SGLT2 Inhibitors in Chronic Kidney Disease. Front. Immunol. 2021, 12, 694457. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Shao, F.; Cai, M.; Liu, Z.; Peng, Y.; Huang, Q.; Meng, F. Activated Pancreatic Stellate Cells Enhance the Warburg Effect to Cause the Malignant Development in Chronic Pancreatitis. Front. Oncol. 2021, 11, 714598. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yin, H.; Wang, Z.; Yuan, Q.; Xu, F.; Chen, Y.; Li, C. Rho A/ROCK1 signaling-mediated metabolic reprogramming of valvular interstitial cells toward Warburg effect accelerates aortic valve calcification via AMPK/RUNX2 axis. Cell Death Dis. 2023, 14, 108. [Google Scholar] [CrossRef]
- Ravi, A.K.; Muthukrishnan, S.K. Combination of Probiotics and Natural Compounds to Treat Multiple Sclerosis via Warburg Effect. Adv. Pharm. Bull. 2022, 12, 515–523. [Google Scholar] [CrossRef]
- Pucino, V.; Certo, M.; Bulusu, V.; Cucchi, D.; Goldmann, K.; Pontarini, E.; Haas, R.; Smith, J.; Headland, S.E.; Blighe, K.; et al. Lactate Buildup at the Site of Chronic Inflammation Promotes Disease by Inducing CD4(+) T Cell Metabolic Rewiring. Cell Metab. 2019, 30, 1055–1074.E8. [Google Scholar] [CrossRef]
- Austermann, J.; Roth, J.; Barczyk-Kahlert, K. The Good and the Bad: Monocytes’ and Macrophages’ Diverse Functions in Inflammation. Cells 2022, 11, 1979. [Google Scholar] [CrossRef] [PubMed]
- Yunna, C.; Mengru, H.; Lei, W.; Weidong, C. Macrophage M1/M2 polarization. Eur. J. Pharmacol. 2020, 877, 173090. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Yang, L.; Cai, J.; Hu, X.; Li, H.; Zhang, X.; Zhang, X.; Chen, X.; Dong, H.; Nie, H.; et al. A sweet spot for macrophages: Focusing on polarization. Pharmacol. Res. 2021, 167, 105576. [Google Scholar] [CrossRef] [PubMed]
- Caslin, H.L.; Abebayehu, D.; Pinette, J.A.; Ryan, J.J. Lactate Is a Metabolic Mediator That Shapes Immune Cell Fate and Function. Front. Physiol. 2021, 12, 688485. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhai, Z.; Duan, J.; Wang, X.; Zhong, J.; Wu, L.; Li, A.; Cao, M.; Wu, Y.; Shi, H.; et al. Lactate: The Mediator of Metabolism and Immunosuppression. Front. Endocrinol. 2022, 13, 901495. [Google Scholar] [CrossRef]
- de la Cruz-López, K.G.; Castro-Muñoz, L.J.; Reyes-Hernández, D.O.; García-Carrancá, A.; Manzo-Merino, J. Lactate in the Regulation of Tumor Microenvironment and Therapeutic Approaches. Front. Oncol. 2019, 9, 1143. [Google Scholar] [CrossRef]
- Chen, L.; Huang, L.; Gu, Y.; Cang, W.; Sun, P.; Xiang, Y. Lactate-Lactylation Hands between Metabolic Reprogramming and Immunosuppression. Int. J. Mol. Sci. 2022, 23, 11943. [Google Scholar] [CrossRef]
- Gauthier, T.; Chen, W. Modulation of Macrophage Immunometabolism: A New Approach to Fight Infections. Front. Immunol. 2022, 13, 780839. [Google Scholar] [CrossRef]
- Lin, Y.; Wang, Y.; Li, P.F. Mutual regulation of lactate dehydrogenase and redox robustness. Front. Physiol. 2022, 13, 1038421. [Google Scholar] [CrossRef]
- Wu, H.; Wang, Y.; Ying, M.; Jin, C.; Li, J.; Hu, X. Lactate dehydrogenases amplify reactive oxygen species in cancer cells in response to oxidative stimuli. Signal Transduct. Target. Ther. 2021, 6, 242. [Google Scholar] [CrossRef]
- Ye, L.; Jiang, Y.; Zhang, M. Crosstalk between glucose metabolism, lactate production and immune response modulation. Cytokine Growth Factor. Rev. 2022, 68, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.C.; Sohn, H.A.; Park, Z.Y.; Oh, S.; Kang, Y.K.; Lee, K.M.; Kang, M.; Jang, Y.J.; Yang, S.J.; Hong, Y.K.; et al. A lactate-induced response to hypoxia. Cell 2015, 161, 595–609. [Google Scholar] [CrossRef] [PubMed]
- Hayes, C.; Donohoe, C.L.; Davern, M.; Donlon, N.E. The oncogenic and clinical implications of lactate induced immunosuppression in the tumour microenvironment. Cancer Lett. 2021, 500, 75–86. [Google Scholar] [CrossRef]
- Na, Y.R.; Jung, D.; Song, J.; Park, J.W.; Hong, J.J.; Seok, S.H. Pyruvate dehydrogenase kinase is a negative regulator of interleukin-10 production in macrophages. J. Mol. Cell Biol. 2020, 12, 543–555. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xue, L.; Zhu, W.; Liu, L.; Zhang, S.; Luo, D. Lactate from glycolysis regulates inflammatory macrophage polarization in breast cancer. Cancer Immunol. Immunother. 2023, 72, 1917–1932. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Kwon, E.J.; Lee, J.J. Rheumatoid Arthritis: Pathogenic Roles of Diverse Immune Cells. Int. J. Mol. Sci. 2022, 23, 905. [Google Scholar] [CrossRef]
- Pucino, V.; Nefla, M.; Gauthier, V.; Alsaleh, G.; Clayton, S.A.; Marshall, J.; Filer, A.; Clark, A.R.; Raza, K.; Buckley, C.D. Differential effect of lactate on synovial fibroblast and macrophage effector functions. Front. Immunol. 2023, 14, 1183825. [Google Scholar] [CrossRef]
- Hanlon, M.M.; Canavan, M.; Barker, B.E.; Fearon, U. Metabolites as drivers and targets in rheumatoid arthritis. Clin. Exp. Immunol. 2022, 208, 167–180. [Google Scholar] [CrossRef]
- Zhang, D.; Tang, Z.; Huang, H.; Zhou, G.; Cui, C.; Weng, Y.; Liu, W.; Kim, S.; Lee, S.; Perez-Neut, M.; et al. Metabolic regulation of gene expression by histone lactylation. Nature 2019, 574, 575–580. [Google Scholar] [CrossRef]
- del Rey, M.J.; Izquierdo, E.; Caja, S.; Usategui, A.; Santiago, B.; Galindo, M.; Pablos, J.L. Human inflammatory synovial fibroblasts induce enhanced myeloid cell recruitment and angiogenesis through a hypoxia-inducible transcription factor 1alpha/vascular endothelial growth factor-mediated pathway in immunodeficient mice. Arthritis Rheum. 2009, 60, 2926–2934. [Google Scholar] [CrossRef]
- Sun, W.; Ma, J.; Zhao, H.; Xiao, C.; Zhong, H.; Ling, H.; Xie, Z.; Tian, Q.; Chen, H.; Zhang, T.; et al. Resolvin D1 suppresses pannus formation via decreasing connective tissue growth factor caused by upregulation of miRNA-146a-5p in rheumatoid arthritis. Arthritis Res. Ther. 2020, 22, 61. [Google Scholar] [CrossRef] [PubMed]
- Keeter, W.C.; Ma, S.; Stahr, N.; Moriarty, A.K.; Galkina, E.V. Atherosclerosis and multi-organ-associated pathologies. Semin. Immunopathol. 2022, 44, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Pålsson-McDermott, E.M.; O’Neill, L.A.J. Targeting immunometabolism as an anti-inflammatory strategy. Cell Res. 2020, 30, 300–314. [Google Scholar] [CrossRef]
- Feng, T.; Zhao, X.; Gu, P.; Yang, W.; Wang, C.; Guo, Q.; Long, Q.; Liu, Q.; Cheng, Y.; Li, J.; et al. Adipocyte-derived lactate is a signalling metabolite that potentiates adipose macrophage inflammation via targeting PHD2. Nat. Commun. 2022, 13, 5208. [Google Scholar] [CrossRef] [PubMed]
- Koltai, T.; Fliegel, L. Exploring monocarboxylate transporter inhibition for cancer treatment. Explor. Target. Antitumor Ther. 2024, 5, 135–169. [Google Scholar] [CrossRef]
- Elia, I.; Haigis, M.C. Metabolites and the tumour microenvironment: From cellular mechanisms to systemic metabolism. Nat. Metab. 2021, 3, 21–32. [Google Scholar] [CrossRef]
- Faubert, B.; Li, K.Y.; Cai, L.; Hensley, C.T.; Kim, J.; Zacharias, L.G.; Yang, C.; Do, Q.N.; Doucette, S.; Burguete, D.; et al. Lactate Metabolism in Human Lung Tumors. Cell 2017, 171, 358–371.e359. [Google Scholar] [CrossRef]
- Vanhove, K.; Graulus, G.J.; Mesotten, L.; Thomeer, M.; Derveaux, E.; Noben, J.P.; Guedens, W.; Adriaensens, P. The Metabolic Landscape of Lung Cancer: New Insights in a Disturbed Glucose Metabolism. Front. Oncol. 2019, 9, 1215. [Google Scholar] [CrossRef]
- Guo, Z.; Hu, L.; Wang, Q.; Wang, Y.; Liu, X.P.; Chen, C.; Li, S.; Hu, W. Molecular Characterization and Prognosis of Lactate-Related Genes in Lung Adenocarcinoma. Curr. Oncol. 2023, 30, 2845–2861. [Google Scholar] [CrossRef]
- Zhang, Y.; Peng, Q.; Zheng, J.; Yang, Y.; Zhang, X.; Ma, A.; Qin, Y.; Qin, Z.; Zheng, X. The function and mechanism of lactate and lactylation in tumor metabolism and microenvironment. Genes Dis. 2023, 10, 2029–2037. [Google Scholar] [CrossRef]
- Chasseuil, E.; Saint-Jean, M.; Chasseuil, H.; Peuvrel, L.; Quéreux, G.; Nguyen, J.M.; Gaultier, A.; Varey, E.; Khammari, A.; Dréno, B. Blood Predictive Biomarkers for Nivolumab in Advanced Melanoma. Acta Derm. Venereol. 2018, 98, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhao, J.; Wang, J.; Sun, C.; Zhu, X. Prognostic value of lactate dehydrogenase for melanoma patients receiving anti-PD-1/PD-L1 therapy: A meta-analysis. Medicine 2021, 100, e25318. [Google Scholar] [CrossRef] [PubMed]
- Komici, K.; Perna, A.; Rocca, A.; Bencivenga, L.; Rengo, G.; Guerra, G. Endothelial Progenitor Cells and Rheumatoid Arthritis: Response to Endothelial Dysfunction and Clinical Evidences. Int. J. Mol. Sci. 2021, 22, 13675. [Google Scholar] [CrossRef] [PubMed]
- Gan, P.R.; Wu, H.; Zhu, Y.L.; Shu, Y.; Wei, Y. Glycolysis, a driving force of rheumatoid arthritis. Int. Immunopharmacol. 2024, 132, 111913. [Google Scholar] [CrossRef]
- de Oliveira, P.G.; Farinon, M.; Sanchez-Lopez, E.; Miyamoto, S.; Guma, M. Fibroblast-Like Synoviocytes Glucose Metabolism as a Therapeutic Target in Rheumatoid Arthritis. Front. Immunol. 2019, 10, 1743. [Google Scholar] [CrossRef]
- Pradenas, C.; Luque-Campos, N.; Oyarce, K.; Contreras-Lopez, R.; Bustamante-Barrientos, F.A.; Bustos, A.; Galvez-Jiron, F.; Araya, M.J.; Asencio, C.; Lagos, R.; et al. Lactate: An alternative pathway for the immunosuppressive properties of mesenchymal stem/stromal cells. Stem Cell Res. Ther. 2023, 14, 335. [Google Scholar] [CrossRef]
- Yamashita, A.; Zhao, Y.; Matsuura, Y.; Yamasaki, K.; Moriguchi-Goto, S.; Sugita, C.; Iwakiri, T.; Okuyama, N.; Koshimoto, C.; Kawai, K.; et al. Increased metabolite levels of glycolysis and pentose phosphate pathway in rabbit atherosclerotic arteries and hypoxic macrophage. PLoS ONE 2014, 9, e86426. [Google Scholar] [CrossRef]
- Hardy, O.T.; Perugini, R.A.; Nicoloro, S.M.; Gallagher-Dorval, K.; Puri, V.; Straubhaar, J.; Czech, M.P. Body mass index-independent inflammation in omental adipose tissue associated with insulin resistance in morbid obesity. Surg. Obes. Relat. Dis. 2011, 7, 60–67. [Google Scholar] [CrossRef]
- Wang, S.P.; Rubio, L.A.; Duncan, S.H.; Donachie, G.E.; Holtrop, G.; Lo, G.; Farquharson, F.M.; Wagner, J.; Parkhill, J.; Louis, P.; et al. Pivotal Roles for pH, Lactate, and Lactate-Utilizing Bacteria in the Stability of a Human Colonic Microbial Ecosystem. mSystems 2020, 5. [Google Scholar] [CrossRef]
- Kaczmarczyk, O.; Dąbek-Drobny, A.; Woźniakiewicz, M.; Paśko, P.; Dobrowolska-Iwanek, J.; Woźniakiewicz, A.; Piątek-Guziewicz, A.; Zagrodzki, P.; Mach, T.; Zwolińska-Wcisło, M. Fecal Levels of Lactic, Succinic and Short-Chain Fatty Acids in Patients with Ulcerative Colitis and Crohn Disease: A Pilot Study. J. Clin. Med. 2021, 10, 4701. [Google Scholar] [CrossRef]
- Jiang, G.; Hong, J.; Sun, L.; Wei, H.; Gong, W.; Wang, S.; Zhu, J. Glycolysis regulation in tumor-associated macrophages: Its role in tumor development and cancer treatment. Int. J. Cancer 2024, 154, 412–424. [Google Scholar] [CrossRef] [PubMed]
- Lugano, R.; Ramachandran, M.; Dimberg, A. Tumor angiogenesis: Causes, consequences, challenges and opportunities. Cell Mol. Life Sci. 2020, 77, 1745–1770. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Huang, D.; Jiang, Y.; Hou, J.; Tian, M.; Li, J.; Sun, L.; Zhang, Y.; Zhang, T.; Li, Z.; et al. Lactate Modulates Cellular Metabolism Through Histone Lactylation-Mediated Gene Expression in Non-Small Cell Lung Cancer. Front. Oncol. 2021, 11, 647559. [Google Scholar] [CrossRef] [PubMed]
- Van Wilpe, S.; Koornstra, R.; Den Brok, M.; De Groot, J.W.; Blank, C.; De Vries, J.; Gerritsen, W.; Mehra, N. Lactate dehydrogenase: A marker of diminished antitumor immunity. Oncoimmunology 2020, 9, 1731942. [Google Scholar] [CrossRef]
- Sahni, S.; Valecha, G.; Sahni, A. Role of Anti-PD-1 Antibodies in Advanced Melanoma: The Era of Immunotherapy. Cureus 2018, 10, e3700. [Google Scholar] [CrossRef]
- Daneshmandi, S.; Wegiel, B.; Seth, P. Blockade of Lactate Dehydrogenase-A (LDH-A) Improves Efficacy of Anti-Programmed Cell Death-1 (PD-1) Therapy in Melanoma. Cancers 2019, 11, 450. [Google Scholar] [CrossRef] [PubMed]
- Faria-Pereira, A.; Morais, V.A. Synapses: The Brain’s Energy-Demanding Sites. Int. J. Mol. Sci. 2022, 23, 3627. [Google Scholar] [CrossRef]
- Park, I.; Larson, P.E.; Zierhut, M.L.; Hu, S.; Bok, R.; Ozawa, T.; Kurhanewicz, J.; Vigneron, D.B.; Vandenberg, S.R.; James, C.D.; et al. Hyperpolarized 13C magnetic resonance metabolic imaging: Application to brain tumors. Neuro Oncol. 2010, 12, 133–144. [Google Scholar] [CrossRef]
- Sitter, B.; Forsmark, A.; Solheim, O. Elevated Serum Lactate in Glioma Patients: Associated Factors. Front. Oncol. 2022, 12, 831079. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, S.; Li, J.; Yuan, Y.; Chen, S.; Zuo, M.; Li, W.; Feng, W.; Chen, M.; Liu, Y. Prognostic value of lactate metabolism-related gene expression signature in adult primary gliomas and its impact on the tumor immune microenvironment. Front. Oncol. 2022, 12, 1008219. [Google Scholar] [CrossRef]
- Yan, L.; Wang, J.; Cai, X.; Liou, Y.C.; Shen, H.M.; Hao, J.; Huang, C.; Luo, G.; He, W. Macrophage plasticity: Signaling pathways, tissue repair, and regeneration. MedComm 2024, 5, e658. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Saeed, A.; Liu, Q.; Jiang, Q.; Xu, H.; Xiao, G.G.; Rao, L.; Duo, Y. Macrophages in immunoregulation and therapeutics. Signal Transduct. Target. Ther. 2023, 8, 207. [Google Scholar] [CrossRef] [PubMed]
- Locati, M.; Curtale, G.; Mantovani, A. Diversity, Mechanisms, and Significance of Macrophage Plasticity. Annu. Rev. Pathol. 2020, 15, 123–147. [Google Scholar] [CrossRef]
- Smith, R.L.; Soeters, M.R.; Wüst, R.C.I.; Houtkooper, R.H. Metabolic Flexibility as an Adaptation to Energy Resources and Requirements in Health and Disease. Endocr. Rev. 2018, 39, 489–517. [Google Scholar] [CrossRef]
- de Souza, C.R.T.; Almeida, M.C.A.; Khayat, A.S.; da Silva, E.L.; Soares, P.C.; Chaves, L.C.; Burbano, R.M.R. Association between Helicobacter pylori, Epstein-Barr virus, human papillomavirus and gastric adenocarcinomas. World J. Gastroenterol. 2018, 24, 4928–4938. [Google Scholar] [CrossRef]
- Chu, D.; Liu, T.; Yao, Y. Implications of viral infections and oncogenesis in uterine cervical carcinoma etiology and pathogenesis. Front. Microbiol. 2023, 14, 1194431. [Google Scholar] [CrossRef]
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular carcinoma. Nat. Rev. Dis. Prim. 2021, 7, 6. [Google Scholar] [CrossRef]
- Sato, Y.; Tsujinaka, S.; Miura, T.; Kitamura, Y.; Suzuki, H.; Shibata, C. Inflammatory Bowel Disease and Colorectal Cancer: Epidemiology, Etiology, Surveillance, and Management. Cancers 2023, 15, 4154. [Google Scholar] [CrossRef] [PubMed]
- Durham, A.L.; Adcock, I.M. The relationship between COPD and lung cancer. Lung Cancer 2015, 90, 121–127. [Google Scholar] [CrossRef]
- Costache, D.O.; Bejan, H.; Poenaru, M.; Costache, R.S. Skin Cancer Correlations in Psoriatic Patients. Cancers 2023, 15, 2451. [Google Scholar] [CrossRef]
- Ramos-Nino, M.E. The role of chronic inflammation in obesity-associated cancers. ISRN Oncol. 2013, 2013, 697521. [Google Scholar] [CrossRef] [PubMed]
- Kung, C.C.; Dai, S.P.; Chiang, H.; Huang, H.S.; Sun, W.H. Temporal expression patterns of distinct cytokines and M1/M2 macrophage polarization regulate rheumatoid arthritis progression. Mol. Biol. Rep. 2020, 47, 3423–3437. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Lin, H.; Li, M.; Lin, B. Tumor-associated macrophage polarization in the inflammatory tumor microenvironment. Front. Oncol. 2023, 13, 1103149. [Google Scholar] [CrossRef] [PubMed]
- Vezzani, B.; Carinci, M.; Previati, M.; Giacovazzi, S.; Della Sala, M.; Gafà, R.; Lanza, G.; Wieckowski, M.R.; Pinton, P.; Giorgi, C. Epigenetic Regulation: A Link between Inflammation and Carcinogenesis. Cancers 2022, 14, 1221. [Google Scholar] [CrossRef]
- Huang, R.; Kang, T.; Chen, S. The role of tumor-associated macrophages in tumor immune evasion. J. Cancer Res. Clin. Oncol. 2024, 150, 238. [Google Scholar] [CrossRef]
- Liu, N.; Luo, J.; Kuang, D.; Xu, S.; Duan, Y.; Xia, Y.; Wei, Z.; Xie, X.; Yin, B.; Chen, F.; et al. Lactate inhibits ATP6V0d2 expression in tumor-associated macrophages to promote HIF-2α-mediated tumor progression. J. Clin. Investig. 2019, 129, 631–646. [Google Scholar] [CrossRef]
- Mu, X.; Shi, W.; Xu, Y.; Xu, C.; Zhao, T.; Geng, B.; Yang, J.; Pan, J.; Hu, S.; Zhang, C.; et al. Tumor-derived lactate induces M2 macrophage polarization via the activation of the ERK/STAT3 signaling pathway in breast cancer. Cell Cycle 2018, 17, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Bohn, T.; Rapp, S.; Luther, N.; Klein, M.; Bruehl, T.J.; Kojima, N.; Aranda Lopez, P.; Hahlbrock, J.; Muth, S.; Endo, S.; et al. Tumor immunoevasion via acidosis-dependent induction of regulatory tumor-associated macrophages. Nat. Immunol. 2018, 19, 1319–1329. [Google Scholar] [CrossRef]
- Yang, K.; Xu, J.; Fan, M.; Tu, F.; Wang, X.; Ha, T.; Williams, D.L.; Li, C. Lactate Suppresses Macrophage Pro-Inflammatory Response to LPS Stimulation by Inhibition of YAP and NF-κB Activation via GPR81-Mediated Signaling. Front. Immunol. 2020, 11, 587913. [Google Scholar] [CrossRef]
- Vallée, A.; Vallée, J.-N.; Lecarpentier, Y. Metabolic reprogramming in atherosclerosis: Opposed interplay between the canonical WNT/β-catenin pathway and PPARγ. J. Mol. Cell. Cardiol. 2019, 133, 36–46. [Google Scholar] [CrossRef]
- Brown, T.P.; Ganapathy, V. Lactate/GPR81 signaling and proton motive force in cancer: Role in angiogenesis, immune escape, nutrition, and Warburg phenomenon. Pharmacol. Ther. 2020, 206, 107451. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yan, Y.; Du, X.; Zhang, H.; Li, H.; Chen, W. Yogurt Prevents Colitis-Associated Colorectal Cancer in Mice. Mol. Nutr. Food Res. 2023, 67, e2300444. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yao, Z.; Qian, J.; Li, H.; Li, H. Lactate Protects Intestinal Epithelial Barrier Function from Dextran Sulfate Sodium-Induced Damage by GPR81 Signaling. Nutrients 2024, 16, 582. [Google Scholar] [CrossRef]
- Sun, Z.; Han, Y.; Song, S.; Chen, T.; Han, Y.; Liu, Y. Activation of GPR81 by lactate inhibits oscillatory shear stress-induced endothelial inflammation by activating the expression of KLF2. IUBMB Life 2019, 71, 2010–2019. [Google Scholar] [CrossRef]
- Cha, J.H.; Chan, L.C.; Li, C.W.; Hsu, J.L.; Hung, M.C. Mechanisms Controlling PD-L1 Expression in Cancer. Mol. Cell 2019, 76, 359–370. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, X.; Yang, X.; Li, W.; Li, S.; Hu, Z.; Ling, C.; Shi, R.; Liu, J.; Chen, G.; et al. Dual Blockade of Lactate/GPR81 and PD-1/PD-L1 Pathways Enhances the Anti-Tumor Effects of Metformin. Biomolecules 2021, 11, 1373. [Google Scholar] [CrossRef]
- Insel, P.A.; Sriram, K.; Salmerón, C.; Wiley, S.Z. Proton-sensing G protein-coupled receptors: Detectors of tumor acidosis and candidate drug targets. Future Med. Chem. 2020, 12, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Wang, X.; Zhang, Z.; Chen, J.; Wang, F.; Wang, L.; Liu, J. Moderate l-lactate administration suppresses adipose tissue macrophage M1 polarization to alleviate obesity-associated insulin resistance. J. Biol. Chem. 2022, 298, 101768. [Google Scholar] [CrossRef]
- Chen, P.; Zuo, H.; Xiong, H.; Kolar, M.J.; Chu, Q.; Saghatelian, A.; Siegwart, D.J.; Wan, Y. Gpr132 sensing of lactate mediates tumor-macrophage interplay to promote breast cancer metastasis. Proc. Natl. Acad. Sci. USA 2017, 114, 580–585. [Google Scholar] [CrossRef]
- Kern, K.; Schäfer, S.M.G.; Cohnen, J.; Pierre, S.; Osthues, T.; Tarighi, N.; Hohmann, S.; Ferreiros, N.; Brüne, B.; Weigert, A.; et al. The G2A Receptor Controls Polarization of Macrophage by Determining Their Localization Within the Inflamed Tissue. Front. Immunol. 2018, 9, 2261. [Google Scholar] [CrossRef]
- Frasch, S.C.; McNamee, E.N.; Kominsky, D.; Jedlicka, P.; Jakubzick, C.; Zemski Berry, K.; Mack, M.; Furuta, G.T.; Lee, J.J.; Henson, P.M.; et al. G2A Signaling Dampens Colitic Inflammation via Production of IFN-γ. J. Immunol. 2016, 197, 1425–1434. [Google Scholar] [CrossRef] [PubMed]
- Vadevoo, S.M.P.; Gunassekaran, G.R.; Lee, C.; Lee, N.; Lee, J.; Chae, S.; Park, J.Y.; Koo, J.; Lee, B. The macrophage odorant receptor Olfr78 mediates the lactate-induced M2 phenotype of tumor-associated macrophages. Proc. Natl. Acad. Sci. USA 2021, 118, e2102434118. [Google Scholar] [CrossRef]
- Liu, Y.; Jiang, C.; Xu, C.; Gu, L. Systematic analysis of integrated bioinformatics to identify upregulated THBS2 expression in colorectal cancer cells inhibiting tumour immunity through the HIF1A/Lactic Acid/GPR132 pathway. Cancer Cell Int. 2023, 23, 253. [Google Scholar] [CrossRef] [PubMed]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef]
- Zhang, A.; Xu, Y.; Xu, H.; Ren, J.; Meng, T.; Ni, Y.; Zhu, Q.; Zhang, W.B.; Pan, Y.B.; Jin, J.; et al. Lactate-induced M2 polarization of tumor-associated macrophages promotes the invasion of pituitary adenoma by secreting CCL17. Theranostics 2021, 11, 3839–3852. [Google Scholar] [CrossRef]
- Tao, H.; Zhong, X.; Zeng, A.; Song, L. Unveiling the veil of lactate in tumor-associated macrophages: A successful strategy for immunometabolic therapy. Front. Immunol. 2023, 14, 1208870. [Google Scholar] [CrossRef]
- Zhou, H.; Yao, J.; Zhong, Z.; Wei, H.; He, Y.; Li, W.; Hu, K. Lactate-Induced CCL8 in Tumor-Associated Macrophages Accelerates the Progression of Colorectal Cancer through the CCL8/CCR5/mTORC1 Axis. Cancers 2023, 15, 5795. [Google Scholar] [CrossRef]
- Yang, K.; Fan, M.; Wang, X.; Xu, J.; Wang, Y.; Tu, F.; Gill, P.S.; Ha, T.; Liu, L.; Williams, D.L.; et al. Lactate promotes macrophage HMGB1 lactylation, acetylation, and exosomal release in polymicrobial sepsis. Cell Death Differ. 2022, 29, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Chai, P.; Xie, M.; Ge, S.; Ruan, J.; Fan, X.; Jia, R. Histone lactylation drives oncogenesis by facilitating m(6)A reader protein YTHDF2 expression in ocular melanoma. Genome Biol. 2021, 22, 85. [Google Scholar] [CrossRef]
- Park, J.; Lee, K.; Kim, K.; Yi, S.J. The role of histone modifications: From neurodevelopment to neurodiseases. Signal Transduct. Target. Ther. 2022, 7, 217. [Google Scholar] [CrossRef]
- Gu, X.; Zhuang, A.; Yu, J.; Yang, L.; Ge, S.; Ruan, J.; Jia, R.; Fan, X.; Chai, P. Histone lactylation-boosted ALKBH3 potentiates tumor progression and diminished promyelocytic leukemia protein nuclear condensates by m1A demethylation of SP100A. Nucleic Acids Res. 2024, 52, 2273–2289. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Jiang, H.; Dong, M.; Min, J.; He, X.; Tan, Y.; Liu, F.; Chen, M.; Chen, X.; Yin, Q.; et al. Macrophage MCT4 inhibition activates reparative genes and protects from atherosclerosis by histone H3 lysine 18 lactylation. Cell Rep. 2024, 43, 114180. [Google Scholar] [CrossRef]
- Cui, Z.; Li, Y.; Lin, Y.; Zheng, C.; Luo, L.; Hu, D.; Chen, Y.; Xiao, Z.; Sun, Y. Lactylproteome analysis indicates histone H4K12 lactylation as a novel biomarker in triple-negative breast cancer. Front. Endocrinol. 2024, 15, 1328679. [Google Scholar] [CrossRef]
- Zhao, Y.; Jiang, J.; Zhou, P.; Deng, K.; Liu, Z.; Yang, M.; Yang, X.; Li, J.; Li, R.; Xia, J. H3K18 lactylation-mediated VCAM1 expression promotes gastric cancer progression and metastasis via AKT-mTOR-CXCL1 axis. Biochem. Pharmacol. 2024, 222, 116120. [Google Scholar] [CrossRef]
- Wang, J.; Yang, P.; Yu, T.; Gao, M.; Liu, D.; Zhang, J.; Lu, C.; Chen, X.; Zhang, X.; Liu, Y. Lactylation of PKM2 Suppresses Inflammatory Metabolic Adaptation in Pro-inflammatory Macrophages. Int. J. Biol. Sci. 2022, 18, 6210–6225. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yu, Y.; Yang, X.; Wang, Y.; Song, Y.; Wang, Q.; Chen, Z.; Zong, S.; Fan, M.; Meng, X.; et al. Berberine attenuates arthritis in adjuvant-induced arthritic rats associated with regulating polarization of macrophages through AMPK/NF-кB pathway. Eur. J. Pharmacol. 2019, 852, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Cheng, J.; Zong, S.; Yu, Y.; Wang, Y.; Song, Y.; He, R.; Yuan, S.; Chen, T.; Hu, M.; et al. The glycolysis inhibitor 2-deoxyglucose ameliorates adjuvant-induced arthritis by regulating macrophage polarization in an AMPK-dependent manner. Mol. Immunol. 2021, 140, 186–195. [Google Scholar] [CrossRef]
- Li, J.; Zeng, G.; Zhang, Z.; Wang, Y.; Shao, M.; Li, C.; Lu, Z.; Zhao, Y.; Zhang, F.; Ding, W. Urban airborne PM(2.5) induces pulmonary fibrosis through triggering glycolysis and subsequent modification of histone lactylation in macrophages. Ecotoxicol. Environ. Saf. 2024, 273, 116162. [Google Scholar] [CrossRef]
- Richiardone, E.; Al Roumi, R.; Lardinois, F.; Giolito, M.V.; Ambroise, J.; Boidot, R.; Drotleff, B.; Ghesquière, B.; Bellahcène, A.; Bardelli, A.; et al. MCT1-dependent lactate recycling is a metabolic vulnerability in colorectal cancer cells upon acquired resistance to anti-EGFR targeted therapy. Cancer Lett. 2024, 598, 217091. [Google Scholar] [CrossRef]
- Saulle, E.; Spinello, I.; Quaranta, M.T.; Pasquini, L.; Pelosi, E.; Iorio, E.; Castelli, G.; Chirico, M.; Pisanu, M.E.; Ottone, T.; et al. Targeting Lactate Metabolism by Inhibiting MCT1 or MCT4 Impairs Leukemic Cell Proliferation, Induces Two Different Related Death-Pathways and Increases Chemotherapeutic Sensitivity of Acute Myeloid Leukemia Cells. Front. Oncol. 2020, 10, 621458. [Google Scholar] [CrossRef]
- Kvacskay, P.; Yao, N.; Schnotz, J.H.; Scarpone, R.; Carvalho, R.A.; Klika, K.D.; Merkt, W.; Tretter, T.; Lorenz, H.M.; Tykocinski, L.O. Increase of aerobic glycolysis mediated by activated T helper cells drives synovial fibroblasts towards an inflammatory phenotype: New targets for therapy? Arthritis Res. Ther. 2021, 23, 56. [Google Scholar] [CrossRef] [PubMed]
- Purkey, H.E.; Robarge, K.; Chen, J.; Chen, Z.; Corson, L.B.; Ding, C.Z.; DiPasquale, A.G.; Dragovich, P.S.; Eigenbrot, C.; Evangelista, M.; et al. Cell Active Hydroxylactam Inhibitors of Human Lactate Dehydrogenase with Oral Bioavailability in Mice. ACS Med. Chem. Lett. 2016, 7, 896–901. [Google Scholar] [CrossRef] [PubMed]
- Giri, P.; Camarillo, I.G.; Sundararajan, R. Enhancement of reactive oxygen species production in triple negative breast cancer cells treated with electric pulses and resveratrol. Explor. Target. Antitumor Ther. 2023, 4, 42–56. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, X.; Liu, W.; Chen, G.; Liu, J.; Ma, Q.; Hou, P.; Liang, L.; Liu, C. Polyphenol nanocomplex modulates lactate metabolic reprogramming and elicits immune responses to enhance cancer therapeutic effect. Drug Resist. Updat. 2024, 73, 101060. [Google Scholar] [CrossRef] [PubMed]
- Wendt, E.H.U.; Schoenrogge, M.; Vollmar, B.; Zechner, D. Galloflavin Plus Metformin Treatment Impairs Pancreatic Cancer Cells. Anticancer Res. 2020, 40, 153–160. [Google Scholar] [CrossRef]
- Souto-Carneiro, M.M.; Klika, K.D.; Abreu, M.T.; Meyer, A.P.; Saffrich, R.; Sandhoff, R.; Jennemann, R.; Kraus, F.V.; Tykocinski, L.; Eckstein, V.; et al. Effect of Increased Lactate Dehydrogenase A Activity and Aerobic Glycolysis on the Proinflammatory Profile of Autoimmune CD8+ T Cells in Rheumatoid Arthritis. Arthritis Rheumatol. 2020, 72, 2050–2064. [Google Scholar] [CrossRef] [PubMed]
- Tavoulari, S.; Sichrovsky, M.; Kunji, E.R.S. Fifty years of the mitochondrial pyruvate carrier: New insights into its structure, function, and inhibition. Acta Physiol. 2023, 238, e14016. [Google Scholar] [CrossRef]
- Umar, S.M.; Dev, A.J.R.; Kashyap, A.; Rathee, M.; Chauhan, S.S.; Sharma, A.; Prasad, C.P. 7-amino carboxycoumarin 2 inhibits lactate induced epithelial-to-mesenchymal transition via MPC1 in oral and breast cancer cells. Cell Biol. Int. 2024, 48, 1185–1197. [Google Scholar] [CrossRef]
- Izumi, H.; Takahashi, M.; Uramoto, H.; Nakayama, Y.; Oyama, T.; Wang, K.Y.; Sasaguri, Y.; Nishizawa, S.; Kohno, K. Monocarboxylate transporters 1 and 4 are involved in the invasion activity of human lung cancer cells. Cancer Sci. 2011, 102, 1007–1013. [Google Scholar] [CrossRef]
- Liu, Q.; Li, X.; Zhang, H.; Li, H. Mannose Attenuates Colitis-Associated Colorectal Tumorigenesis by Targeting Tumor-Associated Macrophages. J. Cancer Prev. 2022, 27, 31–41. [Google Scholar] [CrossRef]
- Wang, Z.H.; Peng, W.B.; Zhang, P.; Yang, X.P.; Zhou, Q. Lactate in the tumour microenvironment: From immune modulation to therapy. EBioMedicine 2021, 73, 103627. [Google Scholar] [CrossRef] [PubMed]
- Anand David, A.V.; Arulmoli, R.; Parasuraman, S. Overviews of Biological Importance of Quercetin: A Bioactive Flavonoid. Pharmacogn. Rev. 2016, 10, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Chen, H.; Le, Y.; Guo, J.; Liu, Z.; Dou, X.; Lu, D. Salvianolic acid A regulates pyroptosis of endothelial cells via directly targeting PKM2 and ameliorates diabetic atherosclerosis. Front. Pharmacol. 2022, 13, 1009229. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Wang, Q.; Jiang, L.; Lu, S.; Li, C.; Xu, C.; Wang, C.; Zhang, E.; Zhang, X. Shikonin ameliorated mice colitis by inhibiting dimerization and tetramerization of PKM2 in macrophages. Front. Pharmacol. 2022, 13, 926945. [Google Scholar] [CrossRef]
- Li, H.M.; Guo, H.L.; Xu, C.; Liu, L.; Hu, S.Y.; Hu, Z.H.; Jiang, H.H.; He, Y.M.; Li, Y.J.; Ke, J.; et al. Inhibition of glycolysis by targeting lactate dehydrogenase A facilitates hyaluronan synthase 2 synthesis in synovial fibroblasts of temporomandibular joint osteoarthritis. Bone 2020, 141, 115584. [Google Scholar] [CrossRef]
- Gupta, V.K.; Sharma, N.S.; Durden, B.; Garrido, V.T.; Kesh, K.; Edwards, D.; Wang, D.; Myer, C.; Mateo-Victoriano, B.; Kollala, S.S.; et al. Hypoxia-Driven Oncometabolite L-2HG Maintains Stemness-Differentiation Balance and Facilitates Immune Evasion in Pancreatic Cancer. Cancer Res. 2021, 81, 4001–4013. [Google Scholar] [CrossRef]
- Zhuang, H.; Lv, Q.; Zhong, C.; Cui, Y.; He, L.; Zhang, C.; Yu, J. Tiliroside Ameliorates Ulcerative Colitis by Restoring the M1/M2 Macrophage Balance via the HIF-1α/glycolysis Pathway. Front. Immunol. 2021, 12, 649463. [Google Scholar] [CrossRef]
- Ochioni, A.C.; Imbroisi Filho, R.; Esteves, A.M.; Leandro, J.G.B.; Demaria, T.M.; do Nascimento Júnior, J.X.; Pereira-Dutra, F.S.; Bozza, P.T.; Sola-Penna, M.; Zancan, P. Clotrimazole presents anticancer properties against a mouse melanoma model acting as a PI3K inhibitor and inducing repolarization of tumor-associated macrophages. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166263. [Google Scholar] [CrossRef]
- Zhao, P.; Wang, S.; Jiang, J.; Gao, Y.; Wang, Y.; Zhao, Y.; Zhang, J.; Zhang, M.; Huang, Y. Targeting lactate metabolism and immune interaction in breast tumor via protease-triggered delivery. J. Control. Release 2023, 358, 706–717. [Google Scholar] [CrossRef]
- Terana, G.T.; Abd-Alhaseeb, M.M.; Omran, G.A.; Okda, T.M. Quercetin potentiates 5-fluorouracil effects in human colon cancer cells through targeting the Wnt/β-catenin signalling pathway: The role of miR-27a. Contemp Oncol. 2022, 26, 229–238. [Google Scholar] [CrossRef]
- Sandforth, L.; Ammar, N.; Dinges, L.A.; Röcken, C.; Arlt, A.; Sebens, S.; Schäfer, H. Impact of the Monocarboxylate Transporter-1 (MCT1)-Mediated Cellular Import of Lactate on Stemness Properties of Human Pancreatic Adenocarcinoma Cells. Cancers 2020, 12, 581. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.; Antunes, B.; Batista, A.; Pinto-Ribeiro, F.; Baltazar, F.; Afonso, J. In Vivo Anticancer Activity of AZD3965: A Systematic Review. Molecules 2021, 27, 181. [Google Scholar] [CrossRef] [PubMed]
- Halford, S.; Veal, G.J.; Wedge, S.R.; Payne, G.S.; Bacon, C.M.; Sloan, P.; Dragoni, I.; Heinzmann, K.; Potter, S.; Salisbury, B.M.; et al. A Phase I Dose-escalation Study of AZD3965, an Oral Monocarboxylate Transporter 1 Inhibitor, in Patients with Advanced Cancer. Clin. Cancer Res. 2023, 29, 1429–1439. [Google Scholar] [CrossRef] [PubMed]
- Manerba, M.; Vettraino, M.; Fiume, L.; Di Stefano, G.; Sartini, A.; Giacomini, E.; Buonfiglio, R.; Roberti, M.; Recanatini, M. Galloflavin (CAS 568-80-9): A novel inhibitor of lactate dehydrogenase. ChemMedChem 2012, 7, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Yang, Y.; Sheng, Y.; Wang, J.; Li, W.; Zhou, X.; Ruan, S.; Han, C. Galloflavin Relieves the Malignant Behavior of Colorectal Cancer Cells in the Inflammatory Tumor Microenvironment. Front. Pharmacol. 2021, 12, 752118. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.; Zhang, L.; Bao, W.; Ni, J.; Wang, Y.; Huang, Y.; Lyv, J.; Cao, X.; Chen, T.; You, K.; et al. Gentisic acid prevents colorectal cancer metastasis via blocking GPR81-mediated DEPDC5 degradation. Phytomedicine 2024, 129, 155615. [Google Scholar] [CrossRef]
- Cheng, S.; Chen, L.; Ying, J.; Wang, Y.; Jiang, W.; Zhang, Q.; Zhang, H.; Wang, J.; Wang, C.; Wu, H.; et al. 20(S)-ginsenoside Rh2 ameliorates ATRA resistance in APL by modulating lactylation-driven METTL3. J. Ginseng Res. 2024, 48, 298–309. [Google Scholar] [CrossRef]
- Wang, C.; Wang, S.; Wang, Z.; Han, J.; Jiang, N.; Qu, L.; Xu, K. Andrographolide regulates H3 histone lactylation by interfering with p300 to alleviate aortic valve calcification. Br. J. Pharmacol. 2024, 181, 1843–1856. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Disease Type | Source of Lactate | Biological Effects of High-Level Lactate | Ref. |
|---|---|---|---|
| Rheumatoid arthritis | Rheumatoid arthritis synovial fibroblasts | Modulate T cells, macrophages, dendritic cells, and osteoclasts differentiation, activation, and function | [36] |
| Amplifying the inflammatory cascade and propelling the pathogenesis | [37,38] | ||
| Atherosclerosis | Smooth muscle cells and macrophages | Activate macrophages and maintain inflammation | [39,40] |
| Obesity | Adipocyte | Intensifies the polarization of adipose tissue macrophages towards a pro-inflammatory stance | [41] |
| Linked to increased fat inflammation and insulin resistance | [42] | ||
| Inflammatory bowel disease | Intestinal anaerobic bacteria | Excessive accumulation leads to macrophage infiltration and increased inflammation | [43,44] |
| Non-small-cell lung cancer | Tumor cells glycolysis | An important potential source of energy | [45] |
| Promote metabolic reprogramming by histone lactylation | [46,47] | ||
| Melanoma | Tumor cells glycolysis | Counteract the efficacy of anti-PD-1/PD-L1 antibodies | [48,49,50] |
| Cerebral glioma | Tumor cells glycolysis | Promote tumor invasion | [51,52] |
| Target | Small Molecules | Structures | Pharmacological Functions | Research Phase | Ref. |
|---|---|---|---|---|---|
| GPR81 | Gentisic acid | 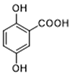 | Inhibits GPR81; alleviates lactate-induced EMT; attenuates the lactate-induced immunosuppression environment | Preclinical studies | [118] |
| MPC | 7ACC2 | 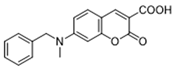 | By impeding pyruvate metabolism, significantly reduced the high lactate-induced protein expression of mesenchymal markers, negating the protective effect of the lactate treatment of tumor cells | Preclinical studies | [119,120] |
| LDH | GSK2837808A | 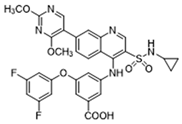 | Increased the production of hyaluronic acid by upregulating the expression of hyaluronan synthase 2 and reduces inflammation; increases sensitivity to anti-PD-1 treatment; reduces tumor volume and induce apoptosis | Preclinical studies | [121,122] |
| FX-11 |  | Mitigates local inflammation by specifically targeting the enzyme LDHA in CD8+ T cells associated with RA | Preclinical studies | [123,124] | |
| GNE-140 | 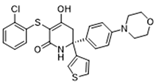 | Ameliorated PM2.5-induced pulmonary inflammation and fibrosis in mice; augmented binding affinity between PGC-1α and LDHA proteins | Preclinical studies | [125,126] | |
| MCT1 | AZD3965 | 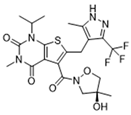 | Inhibits MCT1, suppressing tumor growth | Phase 1 | [127,128] |
| AR-C155858 | 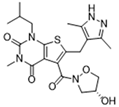 | Inhibits MCT1 and MCT2; enhances the susceptibility of leukemia cells to the chemotherapeutic agent cytarabine | Preclinical studies | [129] | |
| Quercetin | 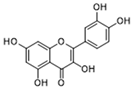 | Inhibits MCT1; enhances the antitumor immune response; enhances the cytotoxic effects of 5-FU | Phase 1/2 | [130,131,132] | |
| AMPK | 2-deoxyglucose | 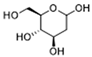 | Glycolysis inhibitor; underwent a metabolic shift from a glycolytic to an oxidative phosphorylation state in LPS-stimulated macrophages | Phase 1/2 | [114] |
| Berberine | 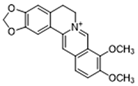 | Suppresses glycolysis in M1 macrophages by diminishing lactate production, reducing glucose uptake and enhancing intracellular ATP levels; significantly elevates the ratio of M2 macrophages | Preclinical studies | [113] | |
| PI3K | Clotrimazole |  | Negatively regulates PI3K; induced macrophage M1 polarization by inhibiting glycolytic metabolism | FDA-approved | [133] |
| HIF-1α | Tiliroside | 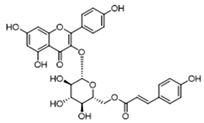 | Downregulated HIF-1α-regulated glycolytic enzymes, preventing the classic M1 macrophage polarization in macrophages | Preclinical studies | [117] |
| PKM2 | Shikonin | 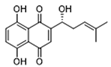 | Inhibits PKM2; improved the symptoms of colitis in DSS mice; inhibits the polarization of M1 macrophages | Preclinical studies | [116] |
| Salvianolic acid A | 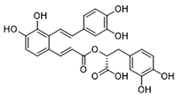 | Inhibits Y105 phosphorylation and blocks the nuclear translocation of PKM2; suppresses the downstream NLR family pyrin domain-containing 3 inflammasome activation in macrophages | Preclinical studies | [115] | |
| HK2 | Mannose | 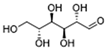 | Targets HK2 in colorectal cancer cells and downregulates markers of the M2-like phenotype of macrophages; inhibits LPS-induced macrophage activation and IL-1β production by raising intracellular mannose-6-phosphate levels and impairing glucose metabolism | Preclinical studies | [134] |
| Histone lactylation | 20 (S)-Ginsenoside Rh2 | 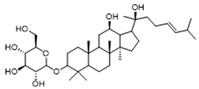 | Histone deacetylase inhibitor; enhances the sensitivity of ATRA differentiation therapy | Preclinical studies | [135] |
| Andrographolide | 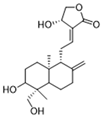 | Downregulates high lactate-induced H3Kla and H3K9la by targeting p300 transferase | Preclinical studies | [136] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, R.; Ren, W.-J.; Wang, L.-Y.; Zhang, W.; Jiang, Z.-H.; Zhu, G.-Y. Targeting Lactate: An Emerging Strategy for Macrophage Regulation in Chronic Inflammation and Cancer. Biomolecules 2024, 14, 1202. https://doi.org/10.3390/biom14101202
Jiang R, Ren W-J, Wang L-Y, Zhang W, Jiang Z-H, Zhu G-Y. Targeting Lactate: An Emerging Strategy for Macrophage Regulation in Chronic Inflammation and Cancer. Biomolecules. 2024; 14(10):1202. https://doi.org/10.3390/biom14101202
Chicago/Turabian StyleJiang, Rong, Wen-Jing Ren, Li-Ying Wang, Wei Zhang, Zhi-Hong Jiang, and Guo-Yuan Zhu. 2024. "Targeting Lactate: An Emerging Strategy for Macrophage Regulation in Chronic Inflammation and Cancer" Biomolecules 14, no. 10: 1202. https://doi.org/10.3390/biom14101202