


Design of a Robust Flow Cytometric Approach for Phenotypical and Functional Analysis of Human Monocyte Subsets in Health and Disease
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Blood Samples and Study Population
2.2. Preparation and Characterisation of Fluorescent Lipid Nanoparticle
2.3. Flow Cytometry
2.3.1. Sample Preparation and Staining
2.3.2. Fixation and Permeabilisation for Intracellular Staining
2.3.3. Cellular Uptake of Perfluorocarbon Nanoemulsions (PFCs)
2.3.4. Cellular Uptake of 2-NBD-Glucose
2.3.5. Analysis of Flow Cytometric Data
2.4. Statistical Analysis of the Data
3. Results
3.1. Identification of Monocytes and Monocyte Subsets
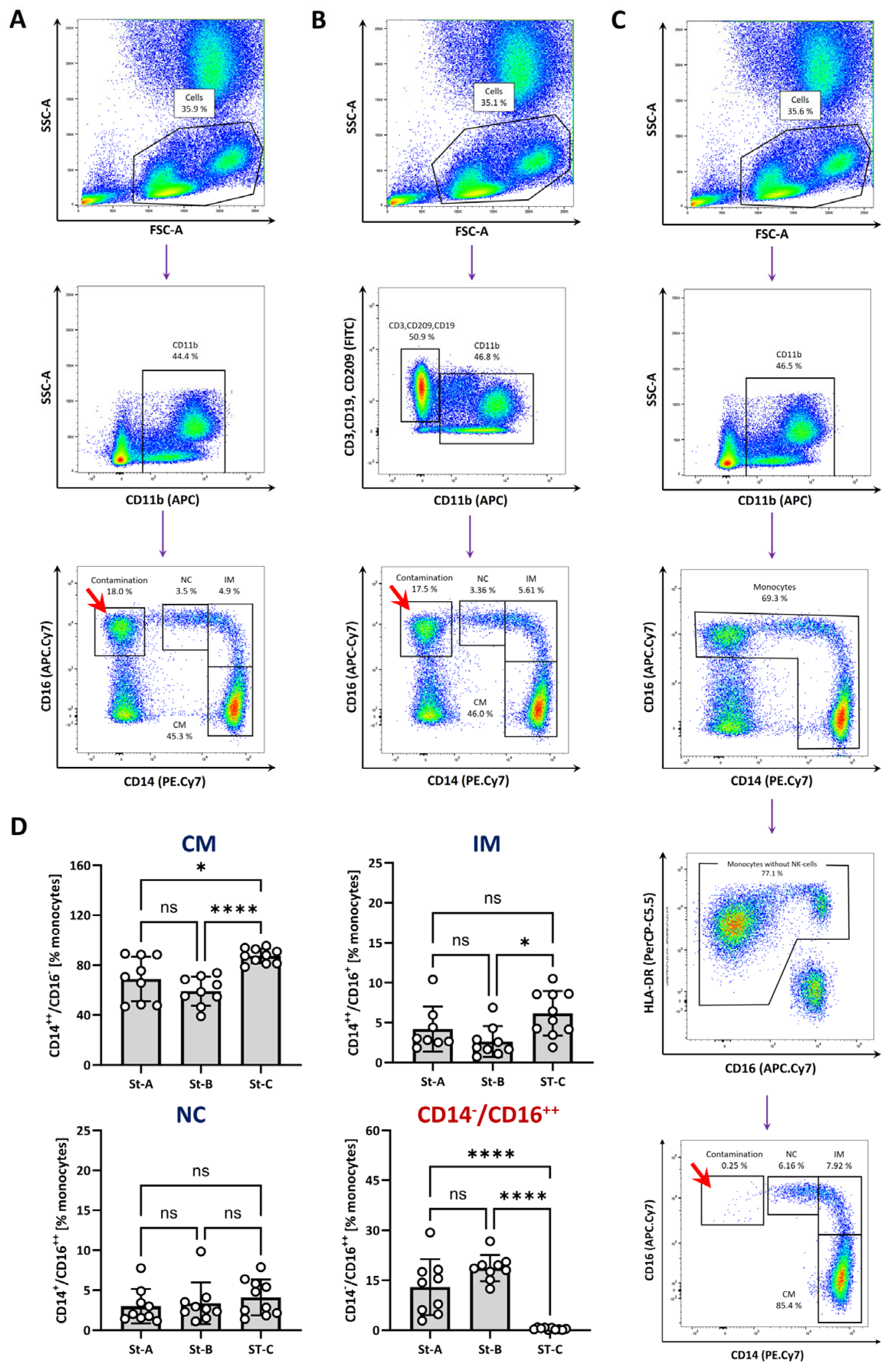
3.2. Analysis of Monocyte Subpopulations in Patients with Coronary Heart Disease (CHD) and with ST-Elevation Myocardial Infarction (STEMI)
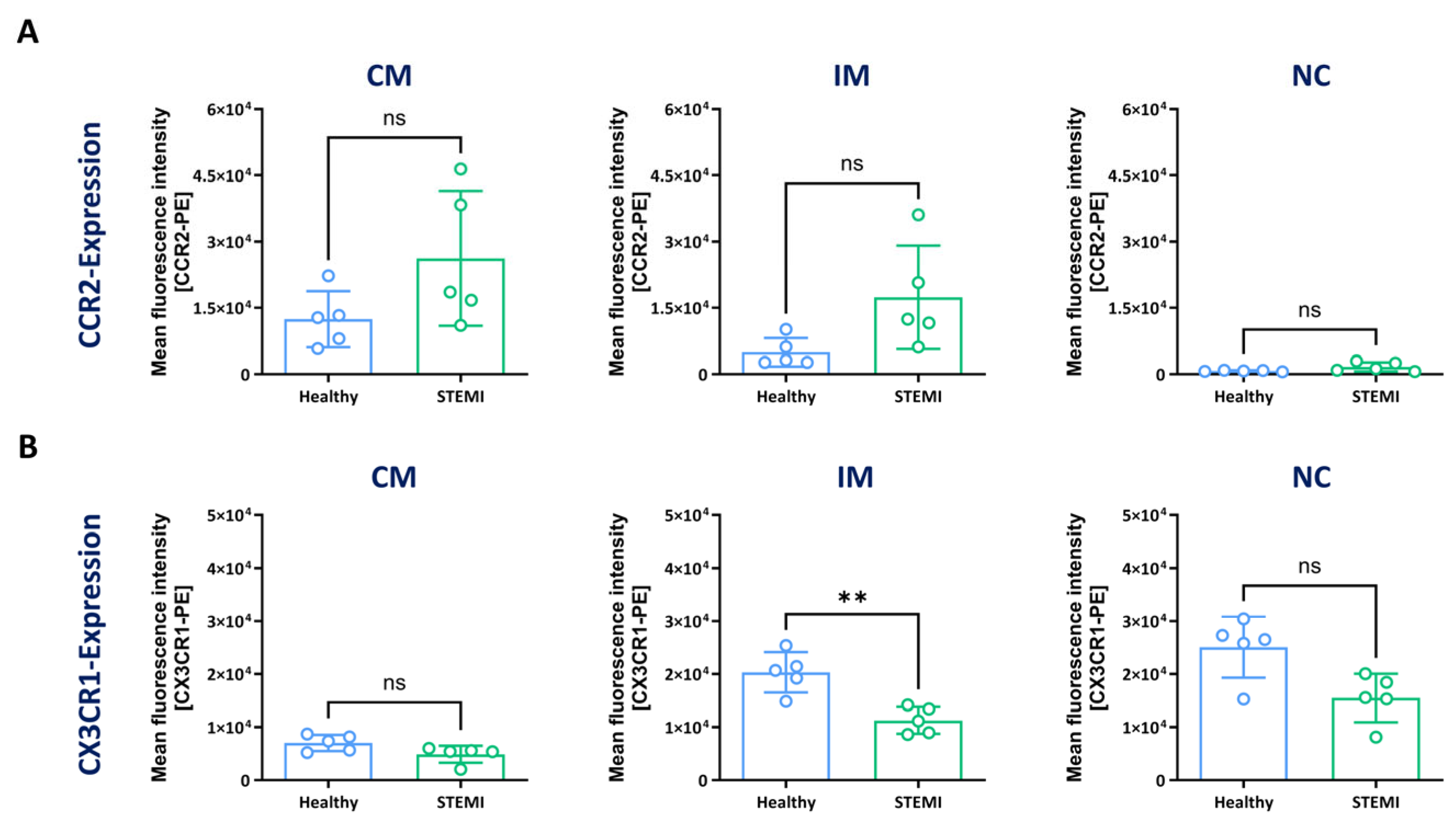
3.3. Expression of CCR2 and CX3CR1 in Monocyte Subsets
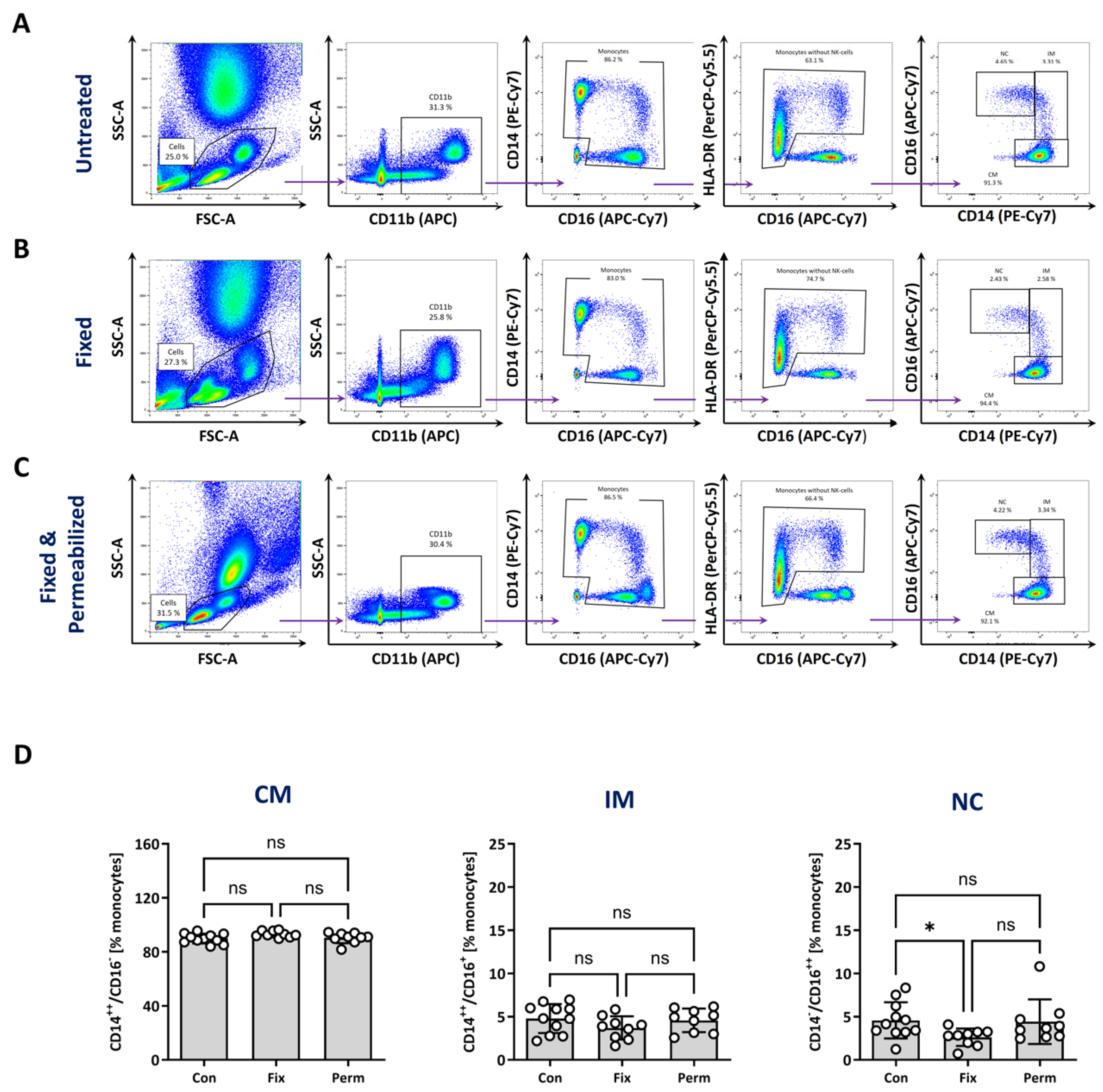
3.4. Identification of Monocyte Subtypes after Fixation and Permeabilisation
3.5. Assessment of Phagocytosis and Glucose Uptake in Monocyte Subsets
3.6. Intracellular Analysis of Components of the MHC-Class II Pathway
4. Discussion
4.1. Flow Cytometry of Monocyte Subtypes
4.2. Phenotype and Function of Monocyte Subtypes in Health and Disease
4.3. Drawbacks of the CD14/CD16 Classification of Monocyte Subtypes
4.4. Limitations of this Study
4.5. Conclusions and Outlook
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Albegger, K.W. Zur Morphologie und Bedeutung des Mononukleären-Phagozyten-Systems (MPS) bei der chronischen Rhinosinusitis. Arch. Otorhinolaryngol. 1976, 214, 27–48. [Google Scholar] [CrossRef] [PubMed]
- Spahn, J.H.; Kreisel, D. Monocytes in Sterile Inflammation: Recruitment and Functional Consequences. Arch. Immunol. Ther. Exp. 2014, 62, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Ożańska, A.; Szymczak, D.; Rybka, J. Pattern of Human Monocyte Subpopulations in Health and Disease. Scand. J. Immunol. 2020, 92, e12883. [Google Scholar] [CrossRef] [PubMed]
- Kapellos, T.S.; Bonaguro, L.; Gemünd, I.; Reusch, N.; Saglam, A.; Hinkley, E.R.; Schultze, J.L. Human Monocyte Subsets and Phenotypes in Major Chronic Inflammatory Diseases. Front. Immunol. 2019, 10, 2035. [Google Scholar] [CrossRef]
- Wong, K.L.; Yeap, W.H.; Tai, J.J.Y.; Ong, S.M.; Dang, T.M.; Wong, S.C. The Three Human Monocyte Subsets: Implications for Health and Disease. Immunol. Res. 2012, 53, 41–57. [Google Scholar] [CrossRef]
- Sampath, P.; Moideen, K.; Ranganathan, U.D.; Bethunaickan, R. Monocyte Subsets: Phenotypes and Function in Tuberculosis Infection. Front. Immunol. 2018, 9, 1726. [Google Scholar] [CrossRef]
- Patel, A.A.; Zhang, Y.; Fullerton, J.N.; Boelen, L.; Rongvaux, A.; Maini, A.A.; Bigley, V.; Flavell, R.A.; Gilroy, D.W.; Asquith, B.; et al. The Fate and Lifespan of Human Monocyte Subsets in Steady State and Systemic Inflammation. J. Exp. Med. 2017, 214, 1913–1923. [Google Scholar] [CrossRef]
- Shi, C.; Pamer, E.G. Monocyte Recruitment during Infection and Inflammation. Nat. Rev. Immunol. 2011, 11, 762–774. [Google Scholar] [CrossRef]
- Angelidou, A.; Diray-Arce, J.; Conti, M.-G.; Netea, M.G.; Blok, B.A.; Liu, M.; Sanchez-Schmitz, G.; Ozonoff, A.; van Haren, S.D.; Levy, O. Human Newborn Monocytes Demonstrate Distinct BCG-Induced Primary and Trained Innate Cytokine Production and Metabolic Activation In Vitro. Front. Immunol. 2021, 12, 674334. [Google Scholar] [CrossRef]
- Guilliams, M.; Mildner, A.; Yona, S. Developmental and Functional Heterogeneity of Monocytes. Immunity 2018, 49, 595–613. [Google Scholar] [CrossRef]
- Serbina, N.V.; Jia, T.; Hohl, T.M.; Pamer, E.G. Monocyte-Mediated Defense Against Microbial Pathogens. Annu. Rev. Immunol. 2008, 26, 421–452. [Google Scholar] [CrossRef] [PubMed]
- Medrano-Bosch, M.; Simón-Codina, B.; Jiménez, W.; Edelman, E.R.; Melgar-Lesmes, P. Monocyte-Endothelial Cell Interactions in Vascular and Tissue Remodeling. Front. Immunol. 2023, 14, 1196033. [Google Scholar] [CrossRef] [PubMed]
- Cros, J.; Cagnard, N.; Woollard, K.; Patey, N.; Zhang, S.-Y.; Senechal, B.; Puel, A.; Biswas, S.K.; Moshous, D.; Picard, C.; et al. Human CD14dim Monocytes Patrol and Sense Nucleic Acids and Viruses via TLR7 and TLR8 Receptors. Immunity 2010, 33, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Buscher, K.; Marcovecchio, P.; Hedrick, C.C.; Ley, K. Patrolling Mechanics of Non-Classical Monocytes in Vascular Inflammation. Front. Cardiovasc. Med. 2017, 4, 80. [Google Scholar] [CrossRef] [PubMed]
- Auffray, C.; Fogg, D.; Garfa, M.; Elain, G.; Join-Lambert, O.; Kayal, S.; Sarnacki, S.; Cumano, A.; Lauvau, G.; Geissmann, F. Monitoring of Blood Vessels and Tissues by a Population of Monocytes with Patrolling Behavior. Science 2007, 317, 666–670. [Google Scholar] [CrossRef]
- Chimen, M.; Yates, C.M.; McGettrick, H.M.; Ward, L.S.C.; Harrison, M.J.; Apta, B.; Dib, L.H.; Imhof, B.A.; Harrison, P.; Nash, G.B.; et al. Monocyte Subsets Coregulate Inflammatory Responses by Integrated Signaling through TNF and IL-6 at the Endothelial Cell Interface. J. Immunol. Author Choice 2017, 198, 2834–2843. [Google Scholar] [CrossRef]
- Boyette, L.B.; Macedo, C.; Hadi, K.; Elinoff, B.D.; Walters, J.T.; Ramaswami, B.; Chalasani, G.; Taboas, J.M.; Lakkis, F.G.; Metes, D.M. Phenotype, Function, and Differentiation Potential of Human Monocyte Subsets. PLoS ONE 2017, 12, e0176460. [Google Scholar] [CrossRef]
- Rogacev, K.S.; Zawada, A.M.; Emrich, I.; Seiler, S.; Böhm, M.; Fliser, D.; Woollard, K.J.; Heine, G.H. Lower Apo A-I and Lower HDL-C Levels Are Associated with Higher Intermediate CD14++CD16+ Monocyte Counts That Predict Cardiovascular Events in Chronic Kidney Disease. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2120–2127. [Google Scholar] [CrossRef]
- Zawada, A.M.; Rogacev, K.S.; Rotter, B.; Winter, P.; Marell, R.-R.; Fliser, D.; Heine, G.H. SuperSAGE Evidence for CD14++CD16+ Monocytes as a Third Monocyte Subset. Blood 2011, 118, e50–e61. [Google Scholar] [CrossRef]
- Bocsi, J.; Melzer, S.; Dähnert, I.; Tárnok, A. OMIP-023: 10-Color, 13 Antibody Panel for in-Depth Phenotyping of Human Peripheral Blood Leukocytes. Cytom. A 2014, 85, 781–784. [Google Scholar] [CrossRef]
- Hristov, M.; Schmitz, S.; Nauwelaers, F.; Weber, C. A Flow Cytometric Protocol for Enumeration of Endothelial Progenitor Cells and Monocyte Subsets in Human Blood. J. Immunol. Methods 2012, 381, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Hally, K.E.; Ferrer-Font, L.; Pilkington, K.R.; Larsen, P.D. OMIP 083: A 21-Marker 18-Color Flow Cytometry Panel for in-Depth Phenotyping of Human Peripheral Monocytes. Cytom. A 2022, 101, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Zawada, A.M.; Rogacev, K.S.; Schirmer, S.H.; Sester, M.; Böhm, M.; Fliser, D.; Heine, G.H. Monocyte Heterogeneity in Human Cardiovascular Disease. Immunobiology 2012, 217, 1273–1284. [Google Scholar] [CrossRef] [PubMed]
- Bönner, F.; Jung, C.; Polzin, A.; Erkens, R.; Dannenberg, L.; Ipek, R.; Kaldirim, M.; Cramer, M.; Wischmann, P.; Zaharia, O.-P.; et al. SYSTEMI—Systemic Organ Communication in STEMI: Design and Rationale of a Cohort Study of Patients with ST-Segment Elevation Myocardial Infarction. BMC Cardiovasc. Disord. 2023, 23, 232. [Google Scholar] [CrossRef] [PubMed]
- Flögel, U.; Burghoff, S.; van Lent, P.L.E.M.; Temme, S.; Galbarz, L.; Ding, Z.; El-Tayeb, A.; Huels, S.; Bönner, F.; Borg, N.; et al. Selective Activation of Adenosine A2A Receptors on Immune Cells by a CD73-Dependent Prodrug Suppresses Joint Inflammation in Experimental Rheumatoid Arthritis. Sci. Transl. Med. 2012, 4, 146ra108. [Google Scholar] [CrossRef]
- Flögel, U.; Ding, Z.; Hardung, H.; Jander, S.; Reichmann, G.; Jacoby, C.; Schubert, R.; Schrader, J. In Vivo Monitoring of Inflammation after Cardiac and Cerebral Ischemia by 19F Magnetic Resonance Imaging. Circulation 2008, 118, 140–148. [Google Scholar] [CrossRef]
- Berger, M.; Murdoch, D.M.; Staats, J.S.; Chan, C.; Thomas, J.P.; Garrigues, G.; Browndyke, J.N.; Cooter, M.; Quinones, Q.J.; Mathew, J.P.; et al. Flow Cytometry Characterization of Cerebrospinal Fluid Monocytes in Patients with Postoperative Cognitive Dysfunction: A Pilot Study. Anesth. Analg. 2019, 129, e150–e154. [Google Scholar] [CrossRef]
- Marimuthu, R.; Francis, H.; Dervish, S.; Li, S.C.H.; Medbury, H.; Williams, H. Characterization of Human Monocyte Subsets by Whole Blood Flow Cytometry Analysis. J. Vis. Exp. JoVE 2018, 140, e57941. [Google Scholar] [CrossRef]
- Mukherjee, R.; Kanti Barman, P.; Kumar Thatoi, P.; Tripathy, R.; Kumar Das, B.; Ravindran, B. Non-Classical Monocytes Display Inflammatory Features: Validation in Sepsis and Systemic Lupus Erythematous. Sci. Rep. 2015, 5, 13886. [Google Scholar] [CrossRef]
- Nienhaus, F.; Colley, D.; Jahn, A.; Pfeiler, S.; Flocke, V.; Temme, S.; Kelm, M.; Gerdes, N.; Flögel, U.; Bönner, F. Phagocytosis of a PFOB-Nanoemulsion for 19F Magnetic Resonance Imaging: First Results in Monocytes of Patients with Stable Coronary Artery Disease and ST-Elevation Myocardial Infarction. Molecules 2019, 24, 2058. [Google Scholar] [CrossRef]
- Kovalenko, E.I.; Streltsova, M.A.; Kanevskiy, L.M.; Erokhina, S.A.; Telford, W.G. Identification of Human Memory-Like NK Cells. Curr. Protoc. Cytom. 2017, 79, 9–50. [Google Scholar] [CrossRef] [PubMed]
- Krasselt, M.; Baerwald, C.; Wagner, U.; Rossol, M. CD56+ Monocytes Have a Dysregulated Cytokine Response to Lipopolysaccharide and Accumulate in Rheumatoid Arthritis and Immunosenescence. Arthritis Res. Ther. 2013, 15, R139. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.; Wang, F.; Sun, R.; Ling, B.; Tian, Z.; Wei, H. CD11b and CD27 Reflect Distinct Population and Functional Specialization in Human Natural Killer Cells. Immunology 2011, 133, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.L.; Tai, J.J.-Y.; Wong, W.-C.; Han, H.; Sem, X.; Yeap, W.-H.; Kourilsky, P.; Wong, S.-C. Gene Expression Profiling Reveals the Defining Features of the Classical, Intermediate, and Nonclassical Human Monocyte Subsets. Blood 2011, 118, e16–e31. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.S.; Anzinger, J.J.; Butterfield, T.R.; McCune, J.M.; Crowe, S.M. A Simple Flow Cytometric Method to Measure Glucose Uptake and Glucose Transporter Expression for Monocyte Subpopulations in Whole Blood. J. Vis. Exp. JoVE 2016, 114, 54255. [Google Scholar] [CrossRef]
- Jensen, P.E.; Weber, D.A.; Thayer, W.P.; Chen, X.; Dao, C.T. HLA-DM and the MHC Class II Antigen Presentation Pathway. Immunol. Res. 1999, 20, 195–205. [Google Scholar] [CrossRef]
- Schröder, B. The Multifaceted Roles of the Invariant Chain CD74—More than Just a Chaperone. Biochim. Biophys. Acta BBA—Mol. Cell Res. 2016, 1863, 1269–1281. [Google Scholar] [CrossRef]
- Ziegler-Heitbrock, L.; Ancuta, P.; Crowe, S.; Dalod, M.; Grau, V.; Hart, D.N.; Leenen, P.J.M.; Liu, Y.-J.; MacPherson, G.; Randolph, G.J.; et al. Nomenclature of Monocytes and Dendritic Cells in Blood. Blood 2010, 116, e74–e80. [Google Scholar] [CrossRef]
- Duan, M.; Steinfort, D.P.; Smallwood, D.; Hew, M.; Chen, W.; Ernst, M.; Irving, L.B.; Anderson, G.P.; Hibbs, M.L. CD11b Immunophenotyping Identifies Inflammatory Profiles in the Mouse and Human Lungs. Mucosal Immunol. 2016, 9, 550–563. [Google Scholar] [CrossRef]
- Lasitschka, F.; Giese, T.; Paparella, M.; Kurzhals, S.R.; Wabnitz, G.; Jacob, K.; Gras, J.; Bode, K.A.; Heninger, A.-K.; Sziskzai, T.; et al. Human Monocytes Downregulate Innate Response Receptors Following Exposure to the Microbial Metabolite N-Butyrate. Immun. Inflamm. Dis. 2017, 5, 480–492. [Google Scholar] [CrossRef]
- Heimbeck, I.; Hofer, T.P.J.; Eder, C.; Wright, A.K.; Frankenberger, M.; Marei, A.; Boghdadi, G.; Scherberich, J.; Ziegler-Heitbrock, L. Standardized Single-Platform Assay for Human Monocyte Subpopulations: Lower CD14+CD16++ Monocytes in Females. Cytom. A 2010, 77A, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Autissier, P.; Soulas, C.; Burdo, T.H.; Williams, K.C. Immunophenotyping of Lymphocyte, Monocyte and Dendritic Cell Subsets in Normal Rhesus Macaques by 12-Color Flow Cytometry: Clarification on DC Heterogeneity. J. Immunol. Methods 2010, 360, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Grip, O.; Bredberg, A.; Lindgren, S.; Henriksson, G. Increased Subpopulations of CD16+ and CD56+ Blood Monocytes in Patients with Active Crohn’s Disease. Inflamm. Bowel Dis. 2007, 13, 566–572. [Google Scholar] [CrossRef] [PubMed]
- Papewalis, C.; Jacobs, B.; Baran, A.M.; Ehlers, M.; Stoecklein, N.H.; Willenberg, H.S.; Schinner, S.; Anlauf, M.; Raffel, A.; Cupisti, K.; et al. Increased Numbers of Tumor-Lysing Monocytes in Cancer Patients. Mol. Cell. Endocrinol. 2011, 337, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Mangold, A.; Hofbauer, T.M.; Ondracek, A.S.; Artner, T.; Scherz, T.; Speidl, W.S.; Krychtiuk, K.A.; Sadushi-Kolici, R.; Jakowitsch, J.; Lang, I.M. Neutrophil Extracellular Traps and Monocyte Subsets at the Culprit Lesion Site of Myocardial Infarction Patients. Sci. Rep. 2019, 9, 16304. [Google Scholar] [CrossRef]
- Pfluecke, C.; Wydra, S.; Berndt, K.; Tarnowski, D.; Cybularz, M.; Barthel, P.; Linke, A.; Ibrahim, K.; Poitz, D.M. CD11b Expression on Monocytes and Data of Inflammatory Parameters after Transcatheter Aortic Valve Implantation in Dependence of Early Mortality. Data Brief 2020, 31, 105798. [Google Scholar] [CrossRef]
- Ong, S.-M.; Teng, K.; Newell, E.; Chen, H.; Chen, J.; Loy, T.; Yeo, T.-W.; Fink, K.; Wong, S.-C. A Novel, Five-Marker Alternative to CD16–CD14 Gating to Identify the Three Human Monocyte Subsets. Front. Immunol. 2019, 10, 1761. [Google Scholar] [CrossRef]
- Jurado, R.; Huguet, M.; Xicoy, B.; Cabezon, M.; Jimenez-Ponce, A.; Quintela, D.; De La Fuente, C.; Raya, M.; Vinets, E.; Junca, J.; et al. Optimization of Monocyte Gating to Quantify Monocyte Subsets for the Diagnosis of Chronic Myelomonocytic Leukemia. Cytom. B Clin. Cytom. 2022, 104, 319–330. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, X.-L.; Ji, W.-J.; Liu, J.-X.; Guo, Z.-Z.; Ren, D.; Ma, Y.-Q.; Zeng, S.; Xu, Z.-W.; Li, H.-X.; et al. The Kinetics of Circulating Monocyte Subsets and Monocyte-Platelet Aggregates in the Acute Phase of ST-Elevation Myocardial Infarction: Associations with 2-Year Cardiovascular Events. Medicine 2016, 95, e3466. [Google Scholar] [CrossRef]
- Tapp, L.D.; Shantsila, E.; Wrigley, B.J.; Pamukcu, B.; Lip, G.Y.H. The CD14++CD16+ Monocyte Subset and Monocyte-Platelet Interactions in Patients with ST-Elevation Myocardial Infarction. J. Thromb. Haemost. 2012, 10, 1231–1241. [Google Scholar] [CrossRef]
- Wrigley, B.J.; Shantsila, E.; Tapp, L.D.; Lip, G.Y.H. CD14++CD16+ Monocytes in Patients with Acute Ischaemic Heart Failure. Eur. J. Clin. Investig. 2013, 43, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Kawanaka, N.; Yamamura, M.; Aita, T.; Morita, Y.; Okamoto, A.; Kawashima, M.; Iwahashi, M.; Ueno, A.; Ohmoto, Y.; Makino, H. CD14+,CD16+ Blood Monocytes and Joint Inflammation in Rheumatoid Arthritis. Arthritis Rheum. 2002, 46, 2578–2586. [Google Scholar] [CrossRef] [PubMed]
- Nockher, W.A.; Scherberich, J.E. Expanded CD14+ CD16+ Monocyte Subpopulation in Patients with Acute and Chronic Infections Undergoing Hemodialysis. Infect. Immun. 1998, 66, 2782–2790. [Google Scholar] [CrossRef] [PubMed]
- Silvis, M.J.M.; Kaffka genaamd Dengler, S.E.; Odille, C.A.; Mishra, M.; van der Kaaij, N.P.; Doevendans, P.A.; Sluijter, J.P.G.; de Kleijn, D.P.V.; de Jager, S.C.A.; Bosch, L.; et al. Damage-Associated Molecular Patterns in Myocardial Infarction and Heart Transplantation: The Road to Translational Success. Front. Immunol. 2020, 11, 599511. [Google Scholar] [CrossRef] [PubMed]
- Frangogiannis, N.G. The Inflammatory Response in Myocardial Injury, Repair and Remodeling. Nat. Rev. Cardiol. 2014, 11, 255–265. [Google Scholar] [CrossRef]
- Dietl, K.; Renner, K.; Dettmer, K.; Timischl, B.; Eberhart, K.; Dorn, C.; Hellerbrand, C.; Kastenberger, M.; Kunz-Schughart, L.A.; Oefner, P.J.; et al. Lactic Acid and Acidification Inhibit TNF Secretion and Glycolysis of Human Monocytes. J. Immunol. 2009, 184, 1200–1209. [Google Scholar] [CrossRef]
- Kirichenko, T.; Blokhina, T.; Markina, Y.; Bogatyreva, A.; Tolstik, T.; Markin, A.; Kozlov, S. Monocyte Activation in Patients with Coronary Heart Disease. Atherosclerosis 2023, 379, S12–S13. [Google Scholar] [CrossRef]
- Lee, J.; Tam, H.; Adler, L.; Ilstad-Minnihan, A.; Macaubas, C.; Mellins, E.D. The MHC Class II Antigen Presentation Pathway in Human Monocytes Differs by Subset and Is Regulated by Cytokines. PLoS ONE 2017, 12, e0183594. [Google Scholar] [CrossRef]
- Westhorpe, C.L.V.; Norman, M.U.; Hall, P.; Snelgrove, S.L.; Finsterbusch, M.; Li, A.; Lo, C.; Tan, Z.H.; Li, S.; Nilsson, S.K.; et al. Effector CD4+ T Cells Recognize Intravascular Antigen Presented by Patrolling Monocytes. Nat. Commun. 2018, 9, 747. [Google Scholar] [CrossRef]
- Jamur, M.C.; Oliver, C. Permeabilization of Cell Membranes. In Immunocytochemical Methods and Protocols; Oliver, C., Jamur, M.C., Eds.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2010; pp. 63–66. ISBN 978-1-59745-324-0. [Google Scholar]
- Joshi, I.; Carney, W.P.; Rock, E.P. Utility of Monocyte HLA-DR and Rationale for Therapeutic GM-CSF in Sepsis Immunoparalysis. Front. Immunol. 2023, 14, 1130214. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahrazoglu, T.; Kluczny, J.I.; Kleimann, P.; Irschfeld, L.-M.; Nienhaus, F.T.; Bönner, F.; Gerdes, N.; Temme, S. Design of a Robust Flow Cytometric Approach for Phenotypical and Functional Analysis of Human Monocyte Subsets in Health and Disease. Biomolecules 2024, 14, 1251. https://doi.org/10.3390/biom14101251
Ahrazoglu T, Kluczny JI, Kleimann P, Irschfeld L-M, Nienhaus FT, Bönner F, Gerdes N, Temme S. Design of a Robust Flow Cytometric Approach for Phenotypical and Functional Analysis of Human Monocyte Subsets in Health and Disease. Biomolecules. 2024; 14(10):1251. https://doi.org/10.3390/biom14101251
Chicago/Turabian StyleAhrazoglu, Talia, Jennifer Isabel Kluczny, Patricia Kleimann, Lisa-Marie Irschfeld, Fabian Theodor Nienhaus, Florian Bönner, Norbert Gerdes, and Sebastian Temme. 2024. "Design of a Robust Flow Cytometric Approach for Phenotypical and Functional Analysis of Human Monocyte Subsets in Health and Disease" Biomolecules 14, no. 10: 1251. https://doi.org/10.3390/biom14101251