Applications of Exosomal miRNAs from Mesenchymal Stem Cells as Skin Boosters

 , , , , and
, , , , and
Abstract
1. Introduction
2. Skin Regeneration and Rejuvenation-Associated miRNAs
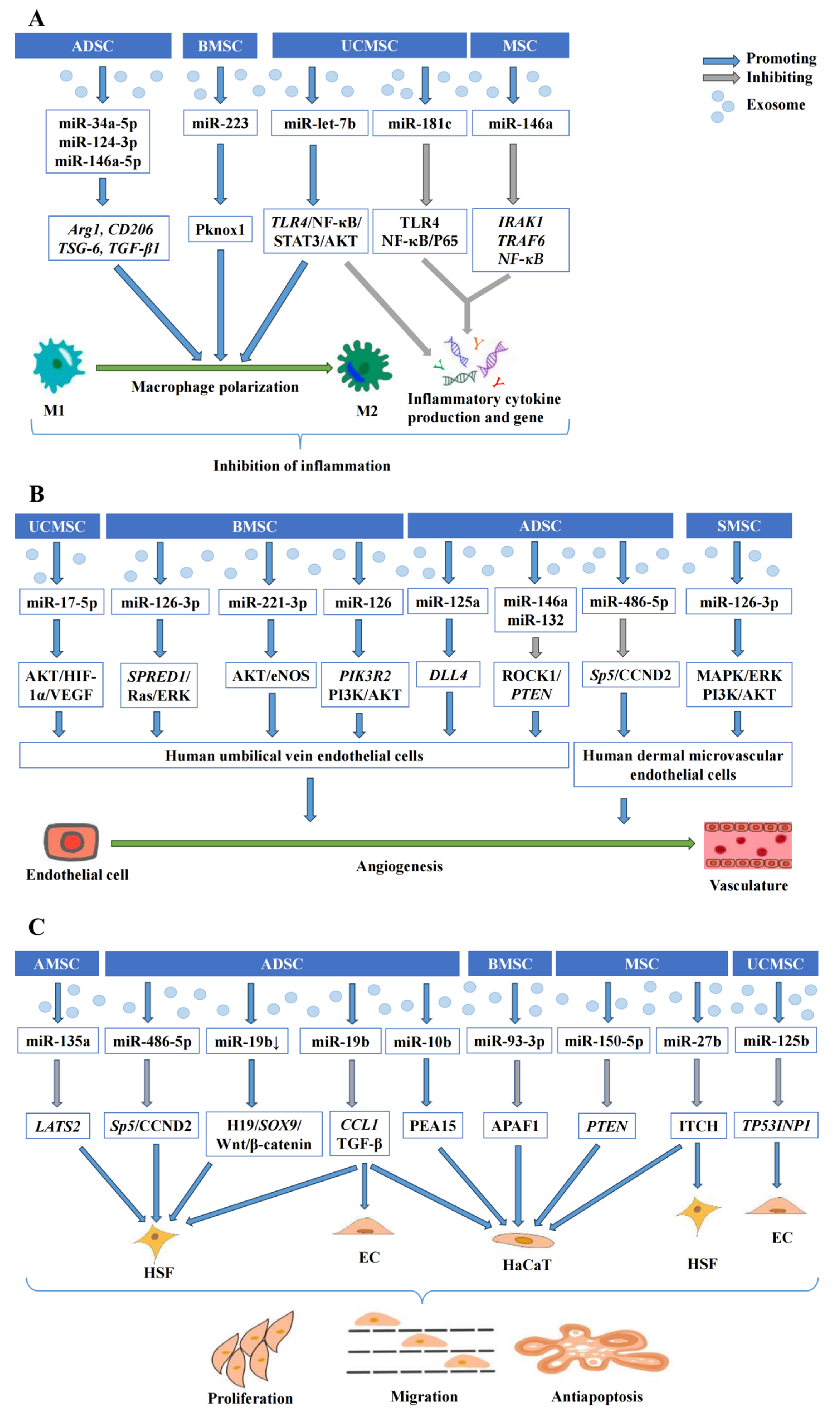
2.1. Wound Healing
2.1.1. Exosomal miRNAs Derived from MSC Modulate Inflammatory Responses
2.1.2. Exosomal miRNAs Derived from MSC Promote Angiogenesis
2.1.3. Exosomal miRNAs Derived from MSC Regulate Cell Proliferation, Migration, and Apoptosis
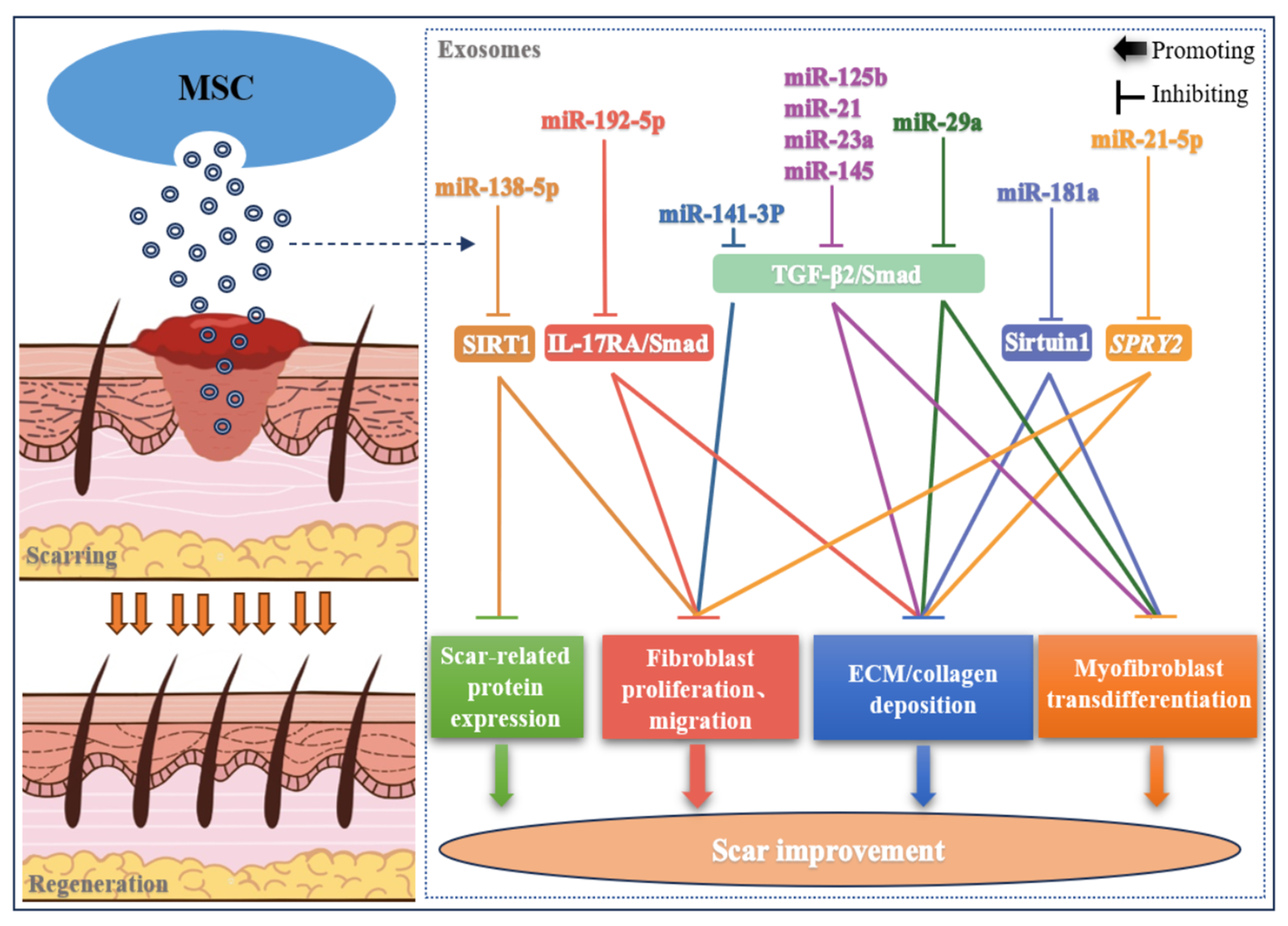
2.2. Skin Scar
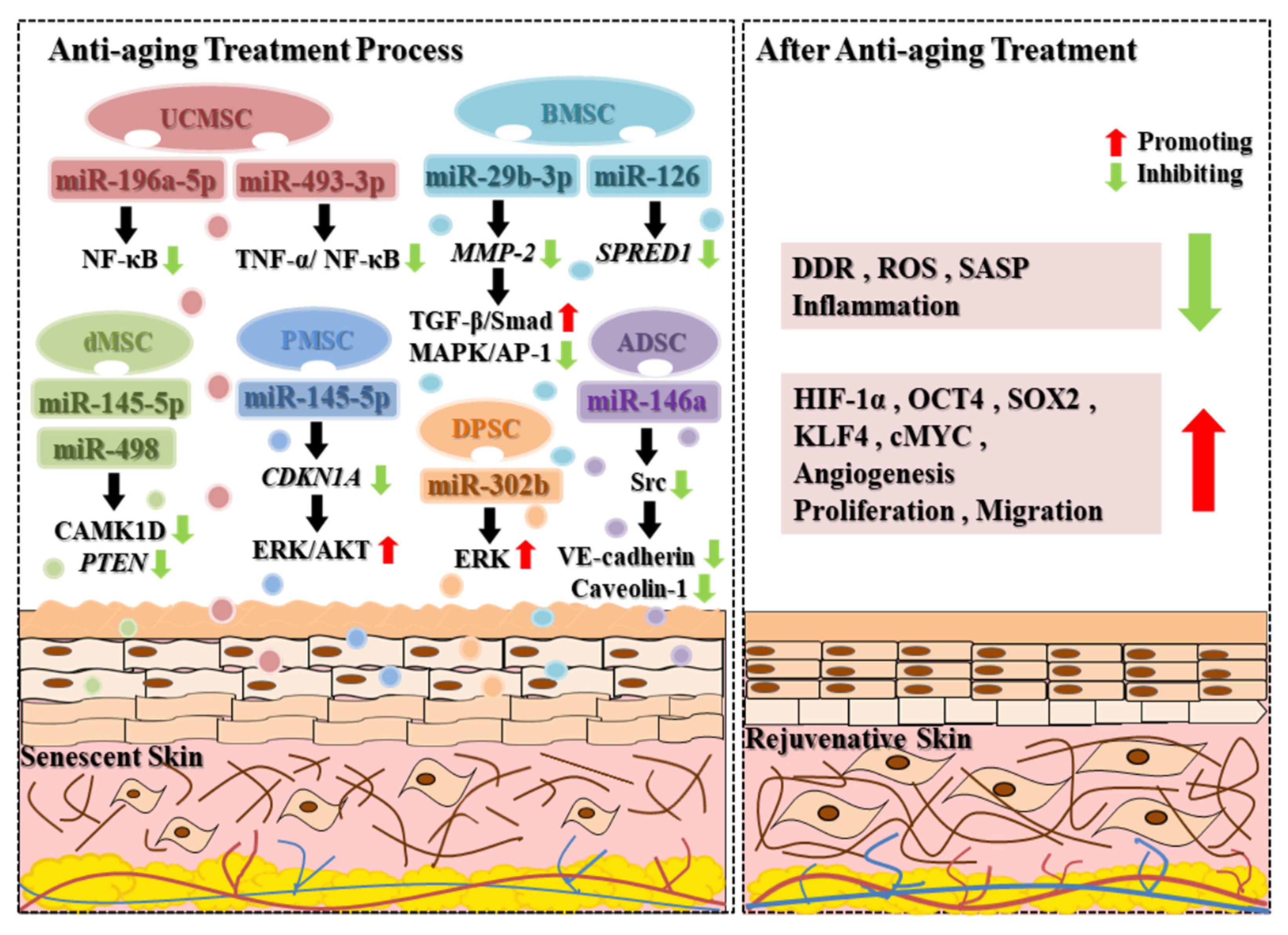
2.3. Anti-Aging
2.4. Hair Loss
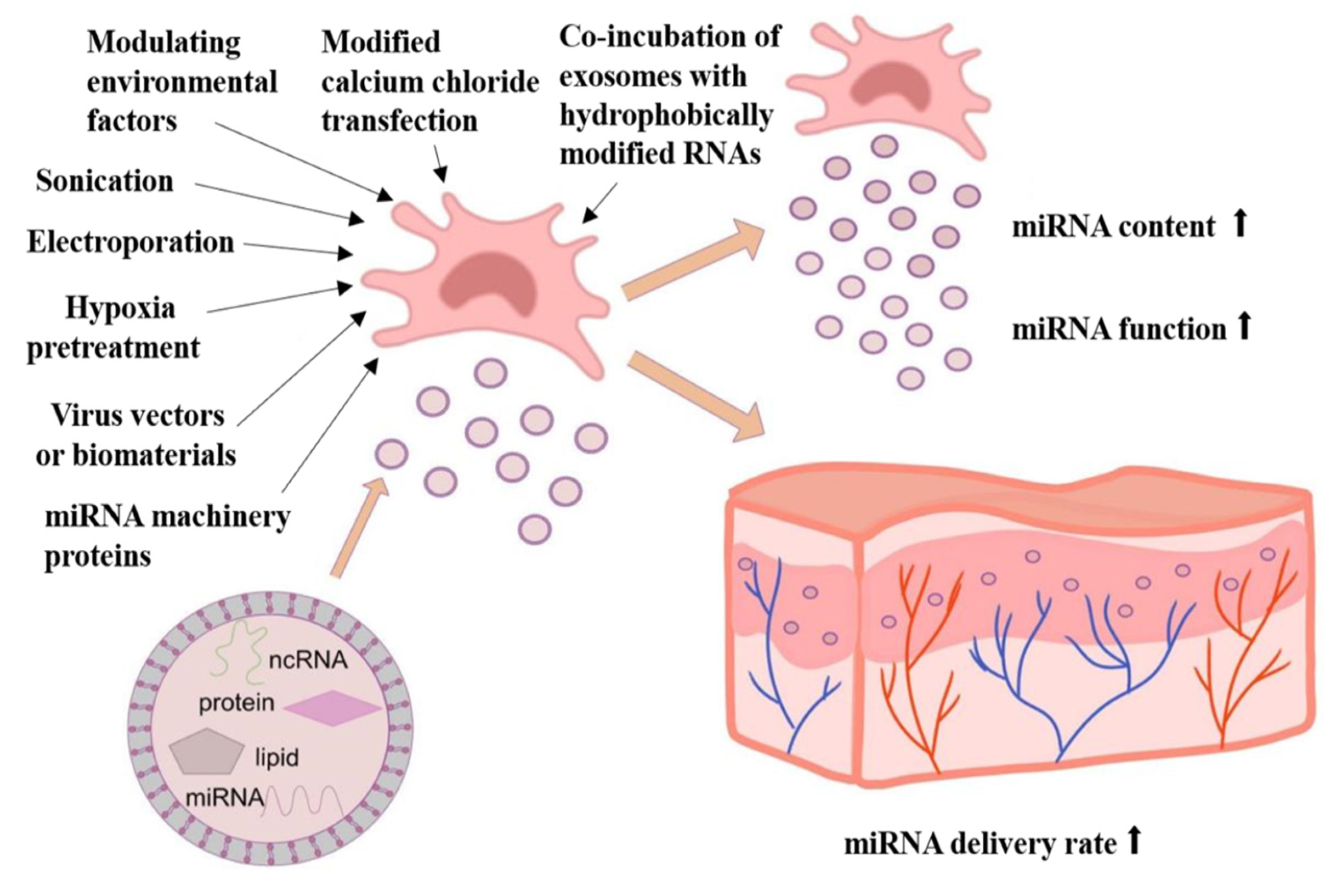
3. Improving Exosomal miRNA Content and Function through Bioengineering Approaches
3.1. Hypoxia-Induced MSCs Produce Wound-Protective Exosome miRNAs
3.2. Improving the miRNA Content and Function by Using Virus Vectors or Biomaterials as Carriers
3.3. Modulative Environmental Factors Regulate Exosomal miRNA Content and Function
3.4. Other Methods to Improve the miRNA Content of Exosomes
4. Conclusions and Prospect
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| MSC | Mesenchymal stem cells |
| EVs | Extracellular vesicles |
| mRNAs | Messenger RNAs |
| miRNAs | MicroRNAs |
| Exos | Exosomes |
| MVs | Microvesicles |
| MSC-Exos | Mesenchymal stem cells-derived exosomes |
| lncRNAs | Long non-coding RNAs |
| UCMSC | Umbilical cord mesenchymal stem cells |
| UCMSC-Exos | Umbilical cord mesenchymal stem cells-derived exosomes |
| NF-κB (NF-κB) | Nuclear transcription factor-kappa B |
| TLR | Toll-like receptor |
| IL-1b | Interleukin-1b |
| TNF-α | Tumor necrosis factor-α |
| IRAK1 | IL-1 receptor-associated kinase 1 |
| TRAF6 | TNF receptor-associated factor 6 |
| BMSC | Bone marrow mesenchymal stem cells |
| ADSC | Adipose mesenchymal stem cells |
| ARG1 | Arginase 1 |
| TSG-6 | Tumor necrosis factor-inducible gene 6 |
| TGF-β1(TGF-β1) | Transforming growth factor beta 1 |
| IFN-γ | Interferon-γ |
| VEGF | Vascular endothelial growth factor |
| MAPK | Mitogen-activated protein kinase |
| M1/2 | Macrophage1/2 |
| LPS | Lipopolysaccharide |
| AKT | Serine/threonine-protein kinase B |
| PI3K | Phosphatidylinositol 3-kinase |
| HIF-1α | Hypoxia inducible factor-1 alpha |
| eNOS | Endothelial nitric oxide synthase |
| PIK3R2 | Phosphoinositol-3 kinase regulatory subunit 2 |
| HUVEC | Human umbilical vein endothelial cells |
| ERK | Extracellular signal-related kinases |
| SPRED1 | Sprouty-related EVH1 domain-containing protein 1 |
| DLL4 | Delta-like 4 |
| ANGPT1 | Angiopoietin1 |
| KDR | Kinase insert domain receptor |
| THBS1 | Thrombospondin-1 |
| VASH1 | Vasohibin-1 |
| CCND2 | Cyclin D2 |
| HMEC | Human dermal microvascular endothelial cells |
| HaCaT | Human keratinocytes |
| APAF1 | Apoptotic protease-activating factor 1 |
| PTEN(PTEN) | Phosphatase and tensin homolog |
| CDK6 | Cyclin dependent kinase 6 |
| HSF | Human skin fibroblasts |
| ECM | Extracellular matrix |
| SIRT1 | Silent information regulator 1 |
| IL-17RA | Interleukin-17 receptor A |
| STAT3 | Signal transducer and activator of transcription 3 |
| PDGF | Platelet-derived growth factor |
| LATS2 | Large tumor suppressor kinase 2 |
| H19 | lncRNA H19 |
| ADSC-Exos | Adipose mesenchymal stem cells-derived exosomes |
| ITCH | Itchy E3 ubiquitin protein ligase |
| SRY | Sex-determining region Y |
| SOX9 | SRY-related high-mobility-group box 9 |
| CCL1 | Chemokine CC motif ligand 1 |
| TP53INP1 | Tumor protein p53 inducible nuclear protein 1 |
| α-SMA | α-smooth muscle actin |
| SASP | Senescence-associated secretory phenotype |
| DDR | DNA damage response |
| MSC-EVs | Mesenchymal stem cells-derived extracellular vesicles |
| PMSC-EVs | Placental mesenchymal stem cells-derived extracellular vesicles |
| ROS | Reactive oxygen species |
| BMSC-EVs | Bone marrow mesenchymal stem cells-derived extracellular vesicles |
| EPC | Endothelial progenitor cells |
| HDF | Human dermal fibroblasts |
| MMP-2 | Matrix metalloproteinase-2 |
| AP-1 | Activator protein-1 |
| DPSC | Dental pulp stem cells |
| UVB | Ultraviolet B |
| OCT4 | Octamer binding transcription factor 4 |
| DPC | Dermal papilla cells |
| HF | Hair follicles |
| DHT | Dihydrotestosterone |
| AGA | Androgen alopecia |
| CNP | Cerium oxide nanoparticles |
| SMSC | Synovial mesenchymal stem cells |
| PMSC | Placenta-derived mesenchymal stem cells |
| NO | Nitric oxide |
| HMGA2 | High mobility group AT-hook 2 |
| Ago2 | Argonaut-2 protein |
| AMSC | Amniotic mesenchymal stem cells |
| dMSC | Decidua mesenchymal stem cells |
| ROCK1 | Rho associated coiled-coil containing protein kinase 1 |
References
- Bonté, F.; Girard, D.; Archambault, J.C.; Desmoulière, A. Skin Changes During Ageing. Biochem. Cell Biol. Ageing Part II Clin. Sci. 2019, 91, 249–280. [Google Scholar] [CrossRef]
- Durai, P.C.; Thappa, D.M.; Kumari, R.; Malathi, M. Aging in elderly: Chronological versus photoaging. Indian J. Dermatol. 2012, 57, 343–352. [Google Scholar] [CrossRef]
- Sen, C.K. Human Wounds and Its Burden: An Updated Compendium of Estimates. Adv. Wound Care 2019, 8, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Olsson, M.; Järbrink, K.; Divakar, U.; Bajpai, R.; Upton, Z.; Schmidtchen, A.; Car, J. The humanistic and economic burden of chronic wounds: A systematic review. Wound Repair Regen. 2018, 27, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Pierpont, Y.N.; Dinh, T.P.; Salas, R.E.; Johnson, E.L.; Wright, T.G.; Robson, M.C.; Payne, W.G. Obesity and Surgical Wound Healing: A Current Review. Int. Sch. Res. Not. 2014, 2014, 638936. [Google Scholar] [CrossRef] [PubMed]
- Prockop, D.J. Marrow stromal cells as stem cells for nonhematopoietic tissues. Science 1997, 276, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Bian, D.; Wu, Y.; Song, G.; Azizi, R.; Zamani, A. The application of mesenchymal stromal cells (MSCs) and their derivative exosome in skin wound healing: A comprehensive review. Stem Cell Res. Ther. 2022, 13, 24. [Google Scholar] [CrossRef] [PubMed]
- Ha, D.H.; Kim, H.K.; Lee, J.; Kwon, H.H.; Park, G.H.; Yang, S.H.; Jung, J.Y.; Choi, H.; Lee, J.H.; Sung, S.; et al. Mesenchymal Stem/Stromal Cell-Derived Exosomes for Immunomodulatory Therapeutics and Skin Regeneration. Cells 2020, 9, 1157. [Google Scholar] [CrossRef]
- Mishra, V.K.; Shih, H.H.; Parveen, F.; Lenzen, D.; Ito, E.; Chan, T.F.; Ke, L.Y. Identifying the Therapeutic Significance of Mesenchymal Stem Cells. Cells 2020, 9, 1145. [Google Scholar] [CrossRef]
- Phinney, D.G.; Di Giuseppe, M.; Njah, J.; Sala, E.; Shiva, S.; St Croix, C.M.; Stolz, D.B.; Watkins, S.C.; Di, Y.P.; Leikauf, G.D.; et al. Mesenchymal stem cells use extracellular vesicles to outsource mitophagy and shuttle microRNAs. Nat. Commun. 2015, 6, 8472. [Google Scholar] [CrossRef]
- Eleuteri, S.; Fierabracci, A. Insights into the Secretome of Mesenchymal Stem Cells and Its Potential Applications. Int. J. Mol. Sci. 2019, 20, 4597. [Google Scholar] [CrossRef] [PubMed]
- Doyle, L.; Wang, M. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [PubMed]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Ludwig, A.-K.; Giebel, B. Exosomes: Small vesicles participating in intercellular communication. Int. J. Biochem. Cell Biol. 2012, 44, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Raik, S.; Kumar, A.; Bhattacharyya, S. Insights into cell-free therapeutic approach: Role of stem cell “soup-ernatant”. Biotechnol. Appl. Biochem. 2017, 65, 104–118. [Google Scholar] [CrossRef] [PubMed]
- Ha, D.H.; Kim, S.D.; Lee, J.; Kwon, H.H.; Park, G.H.; Yang, S.H.; Jung, J.Y.; Lee, J.H.; Park, S.R.; Youn, J.; et al. Toxicological evaluation of exosomes derived from human adipose tissue-derived mesenchymal stem/stromal cells. Regul. Toxicol. Pharmacol. 2020, 115, 104686. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.A.; Hoyt, B.W.; Rowe, C.J.; Dey, D.; Davis, T.A. Alarming Cargo: The Role of Exosomes in Trauma-Induced Inflammation. Biomolecules 2021, 11, 522. [Google Scholar] [CrossRef] [PubMed]
- Najar, M.; Fahmi, H.; Merimi, M. The Medicinal Potential of Mesenchymal Stem/Stromal Cells in Immuno- and Cancer Therapy. Biomolecules 2023, 13, 1171. [Google Scholar] [CrossRef]
- Trzyna, A.; Banaś-Ząbczyk, A. Adipose-Derived Stem Cells Secretome and Its Potential Application in “Stem Cell-Free Therapy”. Biomolecules 2021, 11, 878. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Xu, H.K.; Chen, L.J.; Zhou, S.N.; Li, Y.F.; Xiang, C. Multifunctional role of microRNAs in mesenchymal stem cell-derived exosomes in treatment of diseases. World J. Stem Cells 2020, 12, 1276–1294. [Google Scholar] [CrossRef] [PubMed]
- Ross, K. MiR equal than others: MicroRNA enhancement for cutaneous wound healing. J. Cell. Physiol. 2021, 236, 8050–8059. [Google Scholar] [CrossRef] [PubMed]
- Yan, T.; Huang, L.; Yan, Y.; Zhong, Y.; Xie, H.; Wang, X. Bone marrow mesenchymal stem cell-derived exosome miR-29b-3p alleviates UV irradiation-induced photoaging in skin fibroblast. Photodermatol. Photoimmunol. Photomed. 2023, 39, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Yuan, R.; Dai, X.; Li, Y.; Li, C.; Liu, L. Exosomes from miR-29a-modified adipose-derived mesenchymal stem cells reduce excessive scar formation by inhibiting TGF-β2/Smad3 signaling. Mol. Med. Rep. 2021, 24, 758. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Xu, C.; Zhang, Y.; Xue, C.; Yang, C.; Bi, H.; Qian, X.; Wu, M.; Ji, K.; Zhao, Y.; et al. Umbilical Cord-Derived Mesenchymal Stem Cell-Derived Exosomal MicroRNAs Suppress Myofibroblast Differentiation by Inhibiting the Transforming Growth Factor-β/SMAD2 Pathway During Wound Healing. Stem Cells Transl. Med. 2016, 5, 1425–1439. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Wang, Y.; Ma, K.; Li, Q.; Li, B.; Hu, W.; Fu, X.; Zhang, C. Extracellular Vesicles from Human Umbilical Cord Mesenchymal Stem Cells Facilitate Diabetic Wound Healing Through MiR-17-5p-mediated Enhancement of Angiogenesis. Stem Cell Rev. Rep. 2022, 18, 1025–1040. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Tao, C.; Zhang, A.; Li, X.; Guo, Y.; Wei, H.; Yin, Q.; Li, Q.; Jin, P. Exosomal microRNA-93-3p secreted by bone marrow mesenchymal stem cells downregulates apoptotic peptidase activating factor 1 to promote wound healing. Bioengineered 2022, 13, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Chen, T.; Hao, Y.; Zhang, F.; Tang, X.; Wang, D.; Wei, Z.; Qi, J. Exosomal miR-135a derived from human amnion mesenchymal stem cells promotes cutaneous wound healing in rats and fibroblast migration by directly inhibiting LATS2 expression. Stem Cell Res. Ther. 2020, 11, 56. [Google Scholar] [CrossRef] [PubMed]
- Siu, M.C.; Voisey, J.; Zang, T.; Cuttle, L. MicroRNAs involved in human skin burns, wound healing and scarring. Wound Repair Regen. 2023, 31, 439–453. [Google Scholar] [CrossRef]
- Zhao, W.; Zhang, R.; Zang, C.; Zhang, L.; Zhao, R.; Li, Q.; Yang, Z.; Feng, Z.; Zhang, W.; Cui, R. Exosome Derived from Mesenchymal Stem Cells Alleviates Pathological Scars by Inhibiting the Proliferation, Migration and Protein Expression of Fibroblasts via Delivering miR-138-5p to Target SIRT1. Int. J. Nanomed. 2022, 17, 4023–4038. [Google Scholar] [CrossRef]
- Jiang, Y.; Xu, X.; Xiao, L.; Wang, L.; Qiang, S. The Role of microRNA in the Inflammatory Response of Wound Healing. Front. Immunol. 2022, 13, 852419. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, L.; Yang, J.; Yu, Y.; Chai, J.; Wang, L.; Ma, L.; Yin, H. Exosome Derived From Human Umbilical Cord Mesenchymal Stem Cell Mediates MiR-181c Attenuating Burn-induced Excessive Inflammation. EBioMedicine 2016, 8, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wu, W.; Zhang, L.; Dorset-Martin, W.; Morris, M.W.; Mitchell, M.E.; Liechty, K.W. The role of microRNA-146a in the pathogenesis of the diabetic wound-healing impairment: Correction with mesenchymal stem cell treatment. Diabetes 2012, 61, 2906–2912. [Google Scholar] [CrossRef] [PubMed]
- Bhaumik, D.; Scott, G.K.; Schokrpur, S.; Patil, C.K.; Campisi, J.; Benz, C.C. Expression of microRNA-146 suppresses NF-kappaB activity with reduction of metastatic potential in breast cancer cells. Oncogene 2008, 27, 5643–5647. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Dong, Z.; Cao, Y.; Wang, H.; Liu, S.; Liao, L.; Jin, Y.; Yuan, L.; Li, B. MSC-Derived Exosome Promotes M2 Polarization and Enhances Cutaneous Wound Healing. Stem Cells Int. 2019, 2019, 7132708. [Google Scholar] [CrossRef]
- Ti, D.; Hao, H.; Tong, C.; Liu, J.; Dong, L.; Zheng, J.; Zhao, Y.; Liu, H.; Fu, X.; Han, W. LPS-preconditioned mesenchymal stromal cells modify macrophage polarization for resolution of chronic inflammation via exosome-shuttled let-7b. J. Transl. Med. 2015, 13, 308. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.S.; Kim, S.; Yang, C.E.; Choi, Y.; Song, S.Y.; Kim, H.O. Human Adipose Mesenchymal Stem Cell-Derived Exosomes: A Key Player in Wound Healing. Tissue Eng. Regen. Med. 2021, 18, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.S.; Kim, S. Human adipose mesenchymal stem cells modulate inflammation and angiogenesis through exosomes. Sci. Rep. 2022, 12, 2776. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Liu, W.; Li, J.; Lu, J.; Lu, H.; Jia, W.; Liu, F. Exosomes derived from atorvastatin-pretreated MSC accelerate diabetic wound repair by enhancing angiogenesis via AKT/eNOS pathway. Stem Cell Res. Ther. 2020, 11, 350. [Google Scholar] [CrossRef]
- Zhang, L.; Ouyang, P.; He, G.; Wang, X.; Song, D.; Yang, Y.; He, X. Exosomes from microRNA-126 overexpressing mesenchymal stem cells promote angiogenesis by targeting the PIK3R2-mediated PI3K/Akt signalling pathway. J. Cell. Mol. Med. 2021, 25, 2148–2162. [Google Scholar] [CrossRef]
- Li, M.; Ke, Q.F.; Tao, S.C.; Guo, S.C.; Rui, B.Y.; Guo, Y.P. Fabrication of hydroxyapatite/chitosan composite hydrogels loaded with exosomes derived from miR-126-3p overexpressed synovial mesenchymal stem cells for diabetic chronic wound healing. J. Mater. Chem. B 2016, 4, 6830–6841. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Du, X.; Zou, S.; Fang, Q.; Wu, M.; Li, H.; Shi, B. IFN-γ enhances the therapeutic efficacy of MSCs-derived exosome via miR-126-3p in diabetic wound healing by targeting SPRED1. J. Diabetes 2024, 16, e13465. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhang, L.; Wang, S.; Han, Q.; Zhao, R.C. Exosomes secreted by mesenchymal stem cells promote endothelial cell angiogenesis by transferring miR-125a. J. Cell Sci. 2016, 129, 2182–2189. [Google Scholar] [CrossRef]
- Lu, Y.; Wen, H.; Huang, J.; Liao, P.; Liao, H.; Tu, J.; Zeng, Y. Extracellular vesicle-enclosed miR-486-5p mediates wound healing with adipose-derived stem cells by promoting angiogenesis. J. Cell. Mol. Med. 2020, 24, 9590–9604. [Google Scholar] [CrossRef]
- Qian, L.; Pi, L.; Fang, B.R.; Meng, X.X. Adipose mesenchymal stem cell-derived exosomes accelerate skin wound healing via the lncRNA H19/miR-19b/SOX9 axis. Lab. Investig. 2021, 101, 1254–1266. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Chen, B.; Zhang, X.; Chen, H. Human Adipose-Derived Mesenchymal Stem Cells-Derived Exosomal microRNA-19b Promotes the Healing of Skin Wounds Through Modulation of the CCL1/TGF-β Signaling Axis. Clin. Cosmet. Investig. Dermatol. 2020, 13, 957–971. [Google Scholar] [CrossRef] [PubMed]
- Xiu, C.; Zheng, H.; Jiang, M.; Li, J.; Zhou, Y.; Mu, L.; Liu, W. MSCs-Derived miR-150-5p-Expressing Exosomes Promote Skin Wound Healing by Activating PI3K/AKT Pathway through PTEN. Int. J. Stem Cells 2022, 15, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Xi, Z.; Chen, G.; Liu, K.; Ma, R.; Zhou, C. Extracellular vesicle-carried microRNA-27b derived from mesenchymal stem cells accelerates cutaneous wound healing via E3 ubiquitin ligase ITCH. J. Cell. Mol. Med. 2020, 24, 11254–11271. [Google Scholar] [CrossRef]
- Liao, X.; Yan, F.; Hu, S.; Mu, J.; Li, S.; He, Y.; Tang, M.; Chen, J.; Yu, L.; Sun, J. Adipose mesenchymal stem cell sheets-derived extracellular vesicles-microRNA-10b promote skin wound healing by elevating expression of CDK6. Biomater. Adv. 2022, 136, 212781. [Google Scholar] [CrossRef]
- Zhang, X.F.; Wang, T.; Wang, Z.X.; Huang, K.P.; Zhang, Y.W.; Wang, G.L.; Zhang, H.J.; Chen, Z.H.; Wang, C.Y.; Zhang, J.X.; et al. Hypoxic ucMSC-secreted exosomal miR-125b promotes endothelial cell survival and migration during wound healing by targeting TP53INP1. Mol. Ther.-Nucleic Acids 2021, 26, 347–359. [Google Scholar] [CrossRef]
- Meng, S.; Wei, Q.; Chen, S.; Liu, X.; Cui, S.; Huang, Q.; Chu, Z.; Ma, K.; Zhang, W.; Hu, W.; et al. MiR-141-3p-Functionalized Exosomes Loaded in Dissolvable Microneedle Arrays for Hypertrophic Scar Treatment. Small 2024, 20, e2305374. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yu, W.; Xiao, C.; Su, N.; Han, Y.; Zhai, L.; Hou, C. Exosome from adipose-derived mesenchymal stem cells attenuates scar formation through microRNA-181a/SIRT1 axis. Arch. Biochem. Biophys. 2023, 746, 109733. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, J.; Shi, J.; Liu, K.; Wang, X.; Jia, Y.; He, T.; Shen, K.; Wang, Y.; Liu, J.; et al. Exosomes derived from human adipose mesenchymal stem cells attenuate hypertrophic scar fibrosis by miR-192-5p/IL-17RA/Smad axis. Stem Cell Res. Ther. 2021, 12, 221, Correction to: Stem Cell Res. Ther. 2021, 12, 490. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Kang, L.; Tian, J.; Wu, Y.; Liu, J.; Li, Z.; Wu, X.; Huang, Y.; Gao, B.; Wang, H.; et al. Exosomes Derived from Bone Mesenchymal Stem Cells with the Stimulation of Fe3O4 Nanoparticles and Static Magnetic Field Enhance Wound Healing Through Upregulated miR-21-5p. Int. J. Nanomed. 2020, 15, 7979–7993. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Wei, Q.; Ma, K.; Wang, Y.; Hu, W.; Meng, H.; Li, Q.; Zhang, Y.; Zhang, W.; Li, H.; et al. P-MSC-derived extracellular vesicles facilitate diabetic wound healing via miR-145-5p/CDKN1A-mediated functional improvements of high glucose-induced senescent fibroblasts. Burn. Trauma 2023, 11, tkad010. [Google Scholar] [CrossRef]
- Su, J.L.; Ma, K.; Zhang, C.P.; Fu, X. Effect of human decidua mesenchymal stem cells-derived exosomes on the function of high glucose-induced senescent human dermal fibroblasts and its possible mechanism. Zhonghua Shao Shang Yu Chuang Mian Xiu Fu Za Zhi 2022, 38, 170–183. [Google Scholar] [CrossRef]
- Xiao, X.; Xu, M.; Yu, H.; Wang, L.; Li, X.; Rak, J.; Wang, S.; Zhao, R.C. Mesenchymal stem cell-derived small extracellular vesicles mitigate oxidative stress-induced senescence in endothelial cells via regulation of miR-146a/Src. Signal Transduct. Target. Ther. 2021, 6, 354. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wei, J.; Da Fonseca Ferreira, A.; Wang, H.; Zhang, L.; Zhang, Q.; Bellio, M.A.; Chu, X.M.; Khan, A.; Jayaweera, D.; et al. Rejuvenation of Senescent Endothelial Progenitor Cells by Extracellular Vesicles Derived From Mesenchymal Stromal Cells. JACC Basic Transl. Sci. 2020, 5, 1127–1141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, T.; Tang, Y.; Ren, G.; Zhang, Y.; Ren, X. Concentrated growth factor inhibits UVA-induced photoaging in human dermal fibroblasts via the MAPK/AP-1 pathway. Biosci. Rep. 2020, 40, BSR20193566. [Google Scholar] [CrossRef]
- Gao, W.; Lin, P.; Hwang, E.; Wang, Y.; Yan, Z.; Ngo, H.T.T.; Yi, T.H. Pterocarpus santalinus L. Regulated Ultraviolet B Irradiation-induced Procollagen Reduction and Matrix Metalloproteinases Expression Through Activation of TGF-β/Smad and Inhibition of the MAPK/AP-1 Pathway in Normal Human Dermal Fibroblasts. Photochem. Photobiol. 2018, 94, 139–149. [Google Scholar] [CrossRef]
- Mas-Bargues, C.; Sanz-Ros, J.; Román-Domínguez, A.; Gimeno-Mallench, L.; Inglés, M.; Viña, J.; Borrás, C. Extracellular Vesicles from Healthy Cells Improves Cell Function and Stemness in Premature Senescent Stem Cells by miR-302b and HIF-1α Activation. Biomolecules 2020, 10, 957. [Google Scholar] [CrossRef]
- Chen, L.; Xie, W.; Wu, K.; Meng, Y.; He, Y.; Cai, J.; Jiang, Y.; Zhao, Q.; Yang, Y.; Zhang, M.; et al. Continuous nutrient supply culture strategy controls multivesicular endosomes pathway and anti-photo-aging miRNA cargo loading of extracellular vesicles. J. Tissue Eng. 2023, 14, 20417314231197604. [Google Scholar] [CrossRef]
- Li, Y.; Wang, G.; Wang, Q.; Zhang, Y.; Cui, L.; Huang, X. Exosomes Secreted from Adipose-Derived Stem Cells Are a Potential Treatment Agent for Immune-Mediated Alopecia. J. Immunol. Res. 2022, 2022, 7471246. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Tang, X.; Zhang, X.; Cao, C.; Yu, M.; Wan, M. Adipose Mesenchymal Stromal Cell-Derived Exosomes Carrying MiR-122-5p Antagonize the Inhibitory Effect of Dihydrotestosterone on Hair Follicles by Targeting the TGF-β1/SMAD3 Signaling Pathway. Int. J. Mol. Sci. 2023, 24, 5703. [Google Scholar] [CrossRef]
- Nasser, M.I.; Masood, M.; Adlat, S.; Gang, D.; Zhu, S.; Li, G.; Li, N.; Chen, J.; Zhu, P. Mesenchymal stem cell-derived exosome microRNA as therapy for cardiac ischemic injury. Biomed. Pharmacother. 2021, 143, 112118. [Google Scholar] [CrossRef] [PubMed]
- Gentile, P.; Garcovich, S. Concise Review: Adipose-Derived Stem Cells (ASCs) and Adipocyte-Secreted Exosomal microRNA (A-SE-miR) Modulate Cancer Growth and proMote Wound Repair. J. Clin. Med. 2019, 8, 855. [Google Scholar] [CrossRef]
- Zhao, R.; Liang, H.; Clarke, E.; Jackson, C.; Xue, M. Inflammation in Chronic Wounds. Int. J. Mol. Sci. 2016, 17, 2085. [Google Scholar] [CrossRef] [PubMed]
- Raziyeva, K.; Kim, Y.; Zharkinbekov, Z.; Kassymbek, K.; Jimi, S.; Saparov, A. Immunology of Acute and Chronic Wound Healing. Biomolecules 2021, 11, 700. [Google Scholar] [CrossRef]
- Domenis, R.; Cifù, A.; Quaglia, S.; Pistis, C.; Moretti, M.; Vicario, A.; Parodi, P.C.; Fabris, M.; Niazi, K.R.; Soon-Shiong, P.; et al. Pro inflammatory stimuli enhance the immunosuppressive functions of adipose mesenchymal stem cells-derived exosomes. Sci. Rep. 2018, 8, 13325. [Google Scholar] [CrossRef]
- Wang, S.; Aurora, A.B.; Johnson, B.A.; Qi, X.; McAnally, J.; Hill, J.A.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. The Endothelial-Specific MicroRNA miR-126 Governs Vascular Integrity and Angiogenesis. Dev. Cell 2008, 15, 261–271. [Google Scholar] [CrossRef]
- Guillamat-Prats, R. The Role of MSC in Wound Healing, Scarring and Regeneration. Cells 2021, 10, 1729. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Han, J. Non-coding RNAs in hypertrophic scars and keloids: Current research and clinical relevance: A review. Int. J. Biol. Macromol. 2024, 256 Pt 1, 128334. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Lu, B.; He, J.; Chen, X.; Fu, Q.; Han, H.; Luo, C.; Yin, H.; Qin, Z.; Lyu, D.; et al. Exosomes-loaded thermosensitive hydrogels for corneal epithelium and stroma regeneration. Biomaterials 2022, 280, 121320. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Pan, Y.; Liu, Y.; Li, X.; Tang, L.; Duan, M.; Li, J.; Zhang, G. Exosomes derived from human umbilical cord blood mesenchymal stem cells stimulate regenerative wound healing via transforming growth factor-β receptor inhibition. Stem Cell Res. Ther. 2021, 12, 434. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.Z.; Liu, J.Q.; Yang, L.L.; Fan, L.; He, T.; Su, L.L.; Shi, J.H.; Tang, C.W.; Zheng, Z.; Hu, D.H. Identification of sirtuin 1 as a promising therapeutic target for hypertrophic scars. Br. J. Pharmacol. 2016, 173, 1589–1601. [Google Scholar] [CrossRef] [PubMed]
- Dodig, S.; Čepelak, I.; Pavić, I. Hallmarks of senescence and aging. Biochem. Medica 2019, 29, 030501. [Google Scholar] [CrossRef] [PubMed]
- Malaquin, N.; Martinez, A.; Rodier, F. Keeping the senescence secretome under control: Molecular reins on the senescence-associated secretory phenotype. Exp. Gerontol. 2016, 82, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, F.; Albertini, M.C.; Orciani, M.; Ceka, A.; Cricca, M.; Procopio, A.D.; Bonafè, M. DNA damage response (DDR) and senescence: Shuttled inflamma-miRNAs on the stage of inflamm-aging. Oncotarget 2015, 6, 35509–35521. [Google Scholar] [CrossRef] [PubMed]
- Urbanelli, L.; Buratta, S.; Sagini, K.; Tancini, B.; Emiliani, C. Extracellular Vesicles as New Players in Cellular Senescence. Int. J. Mol. Sci. 2016, 17, 1408. [Google Scholar] [CrossRef]
- Li, X.; Li, C.; Zhang, W.; Wang, Y.; Qian, P.; Huang, H. Inflammation and aging: Signaling pathways and intervention therapies. Signal Transduct. Target. Ther. 2023, 8, 239. [Google Scholar] [CrossRef]
- Casado-Díaz, A.; Quesada-Gómez, J.M.; Dorado, G. Extracellular Vesicles Derived From Mesenchymal Stem Cells (MSC) in Regenerative Medicine: Applications in Skin Wound Healing. Front. Bioeng. Biotechnol. 2020, 8, 146. [Google Scholar] [CrossRef]
- Borghesan, M.; Fafián-Labora, J.; Eleftheriadou, O.; Carpintero-Fernández, P.; Paez-Ribes, M.; Vizcay-Barrena, G.; Swisa, A.; Kolodkin-Gal, D.; Ximénez-Embún, P.; Lowe, R.; et al. Small Extracellular Vesicles Are Key Regulators of Non-cell Autonomous Intercellular Communication in Senescence via the Interferon Protein IFITM3. Cell Rep. 2019, 27, 3956–3971.e6. [Google Scholar] [CrossRef]
- Houri, K.; Mori, T.; Onodera, Y.; Tsujimoto, T.; Takehara, T.; Nakao, S.; Teramura, T.; Fukuda, K. miR-142 induces accumulation of reactive oxygen species (ROS) by inhibiting pexophagy in aged bone marrow mesenchymal stem cells. Sci. Rep. 2020, 10, 3735. [Google Scholar] [CrossRef]
- Vigneron, A.; Vousden, K.H. p53, ROS and senescence in the control of aging. Aging 2010, 2, 471–474. [Google Scholar] [CrossRef]
- Baar, M.P.; Brandt, R.M.C.; Putavet, D.A.; Klein, J.D.D.; Derks, K.W.J.; Bourgeois, B.R.M.; Stryeck, S.; Rijksen, Y.; van Willigenburg, H.; Feijtel, D.A.; et al. Targeted Apoptosis of Senescent Cells Restores Tissue Homeostasis in Response to Chemotoxicity and Aging. Cell 2017, 169, 132–147.e16. [Google Scholar] [CrossRef]
- El-Domyati, M.; Attia, S.; Saleh, F.; Abdel-Wahab, H. Androgenetic alopecia in males: A histopathological and ultrastructural study. J. Cosmet. Dermatol. 2009, 8, 83–91. [Google Scholar] [CrossRef]
- Matsumura, H.; Mohri, Y.; Binh, N.T.; Morinaga, H.; Fukuda, M.; Ito, M.; Kurata, S.; Hoeijmakers, J.; Nishimura, E.K. Hair follicle aging is driven by transepidermal elimination of stem cells via COL17A1 proteolysis. Science 2016, 351, aad4395. [Google Scholar] [CrossRef]
- Sinclair, R.; Chapman, A.; Magee, J. The lack of significant changes in scalp hair follicle density with advancing age. Br. J. Dermatol. 2005, 152, 646–649. [Google Scholar] [CrossRef]
- Alonso, L.; Fuchs, E. The hair cycle. J. Cell Sci. 2006, 119 Pt 3, 391–393. [Google Scholar] [CrossRef]
- Lay, K.; Kume, T.; Fuchs, E. FOXC1 maintains the hair follicle stem cell niche and governs stem cell quiescence to preserve long-term tissue-regenerating potential. Proc. Natl. Acad. Sci. USA 2016, 113, E1506–E1515. [Google Scholar] [CrossRef]
- Van Neste, D.; Tobin, D.J. Hair cycle and hair pigmentation: Dynamic interactions and changes associated with aging. Micron 2004, 35, 193–200. [Google Scholar] [CrossRef]
- Rahmani, W.; Abbasi, S.; Hagner, A.; Raharjo, E.; Kumar, R.; Hotta, A.; Magness, S.; Metzger, D.; Biernaskie, J. Hair follicle dermal stem cells regenerate the dermal sheath, repopulate the dermal papilla, and modulate hair type. Dev. Cell 2014, 31, 543–558. [Google Scholar] [CrossRef]
- Rajendran, R.L.; Gangadaran, P.; Bak, S.S.; Oh, J.M.; Kalimuthu, S.; Lee, H.W.; Baek, S.H.; Zhu, L.; Sung, Y.K.; Jeong, S.Y.; et al. Extracellular vesicles derived from MSCs activates dermal papilla cell in vitro and promotes hair follicle conversion from telogen to anagen in mice. Sci. Rep. 2017, 7, 15560. [Google Scholar] [CrossRef]
- Tang, X.; Cao, C.; Liang, Y.; Han, L.; Tu, B.; Yu, M.; Wan, M. Adipose-Derived Stem Cell Exosomes Antagonize the Inhibitory Effect of Dihydrotestosterone on Hair Follicle Growth by Activating Wnt/β-Catenin Pathway. Stem Cells Int. 2023, 2023, 5548112. [Google Scholar] [CrossRef]
- Hu, S.; Li, Z.; Lutz, H.; Huang, K.; Su, T.; Cores, J.; Dinh, P.C.; Cheng, K. Dermal exosomes containing miR-218-5p promote hair regeneration by regulating β-catenin signaling. Sci. Adv. 2020, 6, eaba1685. [Google Scholar] [CrossRef]
- Nasirishargh, A.; Kumar, P.; Ramasubramanian, L.; Clark, K.; Hao, D.; Lazar, S.V.; Wang, A. Exosomal microRNAs from mesenchymal stem/stromal cells: Biology and applications in neuroprotection. World J. Stem Cells 2021, 13, 776–794. [Google Scholar] [CrossRef]
- Saleh, B.; Dhaliwal, H.K.; Portillo-Lara, R.; Shirzaei Sani, E.; Abdi, R.; Amiji, M.M.; Annabi, N. Local Immunomodulation Using an Adhesive Hydrogel Loaded with miRNA-Laden Nanoparticles Promotes Wound Healing. Small 2019, 15, e1902232. [Google Scholar] [CrossRef]
- Niemiec, S.M.; Louiselle, A.E.; Hilton, S.A.; Dewberry, L.C.; Zhang, L.; Azeltine, M.; Xu, J.; Singh, S.; Sakthivel, T.S.; Seal, S.; et al. Nanosilk Increases the Strength of Diabetic Skin and Delivers CNP-miR146a to Improve Wound Healing. Front. Immunol. 2020, 11, 590285. [Google Scholar] [CrossRef]
- Zgheib, C.; Hilton, S.A.; Dewberry, L.C.; Hodges, M.M.; Ghatak, S.; Xu, J.; Singh, S.; Roy, S.; Sen, C.K.; Seal, S.; et al. Use of Cerium Oxide Nanoparticles Conjugated with MicroRNA-146a to Correct the Diabetic Wound Healing Impairment. J. Am. Coll. Surg. 2019, 228, 107–115. [Google Scholar] [CrossRef]
- Shen, Y.; Xu, G.; Huang, H.; Wang, K.; Wang, H.; Lang, M.; Gao, H.; Zhao, S. Sequential Release of Small Extracellular Vesicles from Bilayered Thiolated Alginate/Polyethylene Glycol Diacrylate Hydrogels for Scarless Wound Healing. ACS Nano 2021, 15, 6352–6368. [Google Scholar] [CrossRef]
- Du, W.; Zhang, K.; Zhang, S.; Wang, R.; Nie, Y.; Tao, H.; Han, Z.; Liang, L.; Wang, D.; Liu, J.; et al. Enhanced proangiogenic potential of mesenchymal stem cell-derived exosomes stimulated by a nitric oxide releasing polymer. Biomaterials 2017, 133, 70–81. [Google Scholar] [CrossRef]
- Yang, K.; Li, D.; Wang, M.; Xu, Z.; Chen, X.; Liu, Q.; Sun, W.; Li, J.; Gong, Y.; Liu, D.; et al. Exposure to blue light stimulates the proangiogenic capability of exosomes derived from human umbilical cord mesenchymal stem cells. Stem Cell Res. Ther. 2019, 10, 358. [Google Scholar] [CrossRef]
- Zhu, Q.; Heon, M.; Zhao, Z.; He, M. Microfluidic engineering of exosomes: Editing cellular messages for precision therapeutics. Lab Chip 2018, 18, 1690–1703. [Google Scholar] [CrossRef]
- Gupta, S.C.; Patchva, S.; Aggarwal, B.B. Therapeutic roles of curcumin: Lessons learned from clinical trials. AAPS J. 2013, 15, 195–218. [Google Scholar] [CrossRef]
- Sun, D.; Zhuang, X.; Xiang, X.; Liu, Y.; Zhang, S.; Liu, C.; Barnes, S.; Grizzle, W.; Miller, D.; Zhang, H.G. A novel nanoparticle drug delivery system: The anti-inflammatory activity of curcumin is enhanced when encapsulated in exosomes. Mol. Ther. 2010, 18, 1606–1614. [Google Scholar] [CrossRef]
- Segal, M.; Biscans, A.; Gilles, M.E.; Anastasiadou, E.; De Luca, R.; Lim, J.; Khvorova, A.; Slack, F.J. Hydrophobically Modified let-7b miRNA Enhances Biodistribution to NSCLC and Downregulates HMGA2 In Vivo. Mol. Ther.-Nucleic Acids 2020, 19, 267–277. [Google Scholar] [CrossRef]
- Didiot, M.C.; Hall, L.M.; Coles, A.H.; Haraszti, R.A.; Godinho, B.M.; Chase, K.; Sapp, E.; Ly, S.; Alterman, J.F.; Hassler, M.R.; et al. Exosome-mediated Delivery of Hydrophobically Modified siRNA for Huntingtin mRNA Silencing. Mol. Ther. 2016, 24, 1836–1847. [Google Scholar] [CrossRef]
- Li, D.; Li, X.I.; Wang, A.; Meisgen, F.; Pivarcsi, A.; Sonkoly, E.; Ståhle, M.; Landén, N.X. MicroRNA-31 Promotes Skin Wound Healing by Enhancing Keratinocyte Proliferation and Migration. J. Investig. Dermatol. 2015, 135, 1676–1685. [Google Scholar] [CrossRef]
- Xiong, Q.H.; Zhao, L.; Wan, G.Q.; Hu, Y.G.; Li, X.L. Engineered BMSCs-Derived Exosomal miR-542-3p Promotes Cutaneous Wound Healing. Endocr. Metab. Immune Disord.-Drug Targets 2023, 23, 336–346. [Google Scholar] [CrossRef]
- Mead, B.; Tomarev, S. Bone Marrow-Derived Mesenchymal Stem Cells-Derived Exosomes Promote Survival of Retinal Ganglion Cells Through miRNA-Dependent Mechanisms. Stem Cells Transl. Med. 2017, 6, 1273–1285. [Google Scholar] [CrossRef]
- Zhang, Y.; Chopp, M.; Liu, X.S.; Katakowski, M.; Wang, X.; Tian, X.; Wu, D.; Zhang, Z.G. Exosomes Derived from Mesenchymal Stromal Cells Promote Axonal Growth of Cortical Neurons. Mol. Neurobiol. 2017, 54, 2659–2673. [Google Scholar] [CrossRef]
- Qing, L.; Chen, H.; Tang, J.; Jia, X. Exosomes and Their MicroRNA Cargo: New Players in Peripheral Nerve Regeneration. Neurorehabilit. Neural Repair 2018, 32, 765–776. [Google Scholar] [CrossRef]
- Nourian Dehkordi, A.; Mirahmadi Babaheydari, F.; Chehelgerdi, M.; Raeisi Dehkordi, S. Skin tissue engineering: Wound healing based on stem-cell-based therapeutic strategies. Stem Cell Res. Ther. 2019, 10, 111. [Google Scholar] [CrossRef]
- Griffiths, T.W.; Watson, R.E.B.; Langton, A.K. Skin ageing and topical rejuvenation strategies. Br. J. Dermatol. 2023, 189 (Suppl. S1), i17–i23. [Google Scholar] [CrossRef]
- Kwon, H.H.; Yang, S.H.; Lee, J.; Park, B.C.; Park, K.Y.; Jung, J.Y.; Bae, Y.; Park, G.H. Combination Treatment with Human Adipose Tissue Stem Cell-derived Exosomes and Fractional CO2 Laser for Acne Scars: A 12-week Prospective, Double-blind, Randomized, Split-face Study. Acta Derm.-Venereol. 2020, 100, adv00310. [Google Scholar] [CrossRef]
- Park, G.H.; Kwon, H.H.; Seok, J.; Yang, S.H.; Lee, J.; Park, B.C.; Shin, E.; Park, K.Y. Efficacy of combined treatment with human adipose tissue stem cell-derived exosome-containing solution and microneedling for facial skin aging: A 12-week prospective, randomized, split-face study. J. Cosmet. Dermatol. 2023, 22, 3418–3426. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Source of MSC | Targets or Pathways | Processes and Effects | References |
|---|---|---|---|---|
| Wound healing | ||||
| miR-181c | UCMSC | TLR4; NF-κB/P65 | lnflammatory cytokine production ↓ | [32] |
| miR-146a | MSC | IRAK1, TRAF6, NF-κB | lnflammatory cytokine production ↓; Inflammatory gene expression ↓ | [33,34] |
| miR-223 | BMSC | Pknox1 | M2-phenotype macrophage polarization ↑ | [35] |
| miR-let-7b | UCMSC | TLR4/NF-κB/ STAT3/AKT | M2-phenotype macrophage polarization ↑; lnflammatory cytokine production ↓ | [36] |
| miR-34a-5p miR-124-3p miR-146a-5p | ADSC | ARG1, CD206, TSG-6, TGF-β1 | M2-phenotype macrophage polarization ↑ | [37] |
| miR-146a miR-132 | ADSC | ROCK1/PTEN | Pro-angiogenic gene expression ↑; Proliferation and tube formation of HUVEC ↑ | [38] |
| miR-17-5p | UCMSC | AKT/HIF-1α/ VEGF | Proliferation, migration, and tube formation of HUVEC ↑ | [26] |
| miR-221-3p | BMSC | AKT/eNOS | Proliferation, migration, and tube formation of HUVEC ↑; VEGF secretion ↑; Granulation tissue formation ↑ | [39] |
| miR-126 | BMSC | PIK3R2; PI3K/AKT | Proliferation, migration, and tube formation of HUVEC ↑ | [40] |
| miR-126-3p | SMSC | MAPK/ERK; PI3K/AKT | Migration of HMEC ↑; Capillary-network formation ↑ | [41] |
| BMSC | SPRED1/Ras/ERK | Proliferation, migration, and tube formation of HUVEC ↑ | [42] | |
| miR-125a | ADSC | DLL4 | Endothelial tip cell formation ↑ | [43] |
| miR-486-5p | ADSC | Sp5/CCND2 | Proliferation and migration of HSF and HMEC ↑; HMEC angiogenesis ↑ | [44] |
| miR-19b↓ | ADSC | H19/SOX9/ Wnt/β-catenin | Proliferation, migration and invasion of HSF ↑; Collagen fibre formation ↑ | [45] |
| miR-19b | ADSC | CCL1/TGF-β | Proliferation and migration of HSF and HaCaT cells ↑; HSF, HaCaT and endothelial cell apoptosis ↓ | [46] |
| miR-135a | AMSC | LATS2 | Proliferation and migration of fibroblasts ↑ | [28] |
| miR-93-3p | BMSC | APAF1 | Proliferation and migration of HaCaT cells ↑; Cellular apoptosis ↓ | [27] |
| miR-150-5p | MSC | PTEN | Proliferation and migration of HaCaT cells ↑ | [47] |
| miR-27b | MSC | ITCH | Proliferation and migration of HSF and HaCaT cells ↑; Collagen fiber proliferation ↑; Epithelialization ↑ | [48] |
| miR-10b | ADSC | PEA15 | Proliferation and migration of HaCaT cells ↑; Cellular apoptosis ↓ | [49] |
| miR-125b | UCMSC | TP53INP1 | Proliferation and migration of endothelial cells ↑; Cellular apoptosis ↓ | [50] |
| Skin scar | ||||
| miR-141-3p | ADSC | TGF-β2/Smad2/3 | Proliferation and migration of hypertrophic scar fibroblasts ↓; Myofibroblast transdifferentiation ↓ | [51] |
| miR-21, miR-23a, miR-125b, miR-145 | UCMSC | TGF-β2/Smad2 | Myofibroblast transdifferentiation ↓; Collagen deposition ↓ | [25] |
| miR-29a | ADSC | TGF-β2/Smad3 | Migrating and proliferating of hypertrophic scar Fibroblasts ↓; Collagen deposition and ECM fibrosis ↓ | [24] |
| miR-181a | ADSC | Sirtuin1 | Myofibroblast transdifferentiation ↓; Collagen deposition ↓ | [52] |
| miR-138-5P | MSC | SIRT1 | Proliferation, migration and protein expression in human skin fibroblasts ↓ | [30] |
| miR-192-5P | ADSC | IL-17RA/Smad | Proliferation and migration of hypertrophic scar fibroblasts ↓; Myofibroblast transdifferentiation ↓; Collagen deposition ↓ | [53] |
| miR-21-5p | BMSC | SPRY2 | Migrating and proliferating of hypertrophic scar fibroblasts ↓; Collagen deposition ↓ | [54] |
| Anti-aging | ||||
| miR-145-5p | PMSC | CDKN1A; ERK/AKT | Proliferation and migration of senescent fibroblasts ↑; Cellular senescence ↓; Cellular apoptosis ↓ | [55] |
| dMSC | CAMK1D; PTEN | Proliferation and migration of senescent dermal fibroblasts↑; Cellular senescence ↓; Cellular apoptosis ↓ | [56] | |
| miR-146a | ADSC | Src kinase; VE-cadherin; Caveolin-1 | Angiogenesis ↑; Cellular senescence ↓; Migration of senescent endothelial cells ↑ | [57] |
| miR-126 | BMSC | Spred-1 | Angiogenesis ↑; Tissue regeneration ↑ | [58] |
| miR-29b-3p | BMSC | MMP-2; TGF-β/Smad; MAPK/AP-1 | Migration of human dermal fibroblasts ↑; Photoaging ↓; Matrix metalloproteinase ↓; Procollagen ↑ | [23,59,60] |
| miR-302b | DPSC | ERK | Cell proliferation ↑; Stemness ↑; Cellular senescence ↓ | [61] |
| miR-498 | dMSC | CAMK1D; PTEN | Proliferation and migration of senescent dermal fibroblasts ↑; Cellular senescence ↓; Cellular apoptosis ↓ | [56] |
| miR-493-3p | UCMSC | TNF-α/NF-κB | Growth and migration of fibroblasts ↑; Procollagen ↑; Oxidative stress levels ↓; Cellular senescence ↓ | [62] |
| miR-196a-5p | UCMSC | NF-κB | ||
| Hair loss | ||||
| miR-22 ↓ | ADSC | Wnt/β-catenin; TNF-α | Hair growth ↑; Hair regeneration ↑; Dermal thickness ↑; Proliferation and migration of DPC ↑; Anti-apoptosis ↑ | [63] |
| miR-122-5p | ADSC | TGF-β1/Smad3 | Hair bulb size ↑; Dermal thickness ↑ | [64] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, J.; Yang, B.; Liu, S.; Xu, Z.; Ding, Z.; Mo, M. Applications of Exosomal miRNAs from Mesenchymal Stem Cells as Skin Boosters. Biomolecules 2024, 14, 459. https://doi.org/10.3390/biom14040459
Zheng J, Yang B, Liu S, Xu Z, Ding Z, Mo M. Applications of Exosomal miRNAs from Mesenchymal Stem Cells as Skin Boosters. Biomolecules. 2024; 14(4):459. https://doi.org/10.3390/biom14040459
Chicago/Turabian StyleZheng, Jinmei, Beibei Yang, Siqi Liu, Zhenfeng Xu, Zhimeng Ding, and Miaohua Mo. 2024. "Applications of Exosomal miRNAs from Mesenchymal Stem Cells as Skin Boosters" Biomolecules 14, no. 4: 459. https://doi.org/10.3390/biom14040459
APA StyleZheng, J., Yang, B., Liu, S., Xu, Z., Ding, Z., & Mo, M. (2024). Applications of Exosomal miRNAs from Mesenchymal Stem Cells as Skin Boosters. Biomolecules, 14(4), 459. https://doi.org/10.3390/biom14040459