
Bactericidal Permeability-Increasing Protein (BPI) Inhibits Mycobacterium tuberculosis Growth
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Cell Culture and Differentiation
2.3. Internalization of BPI
2.3.1. Western Blot
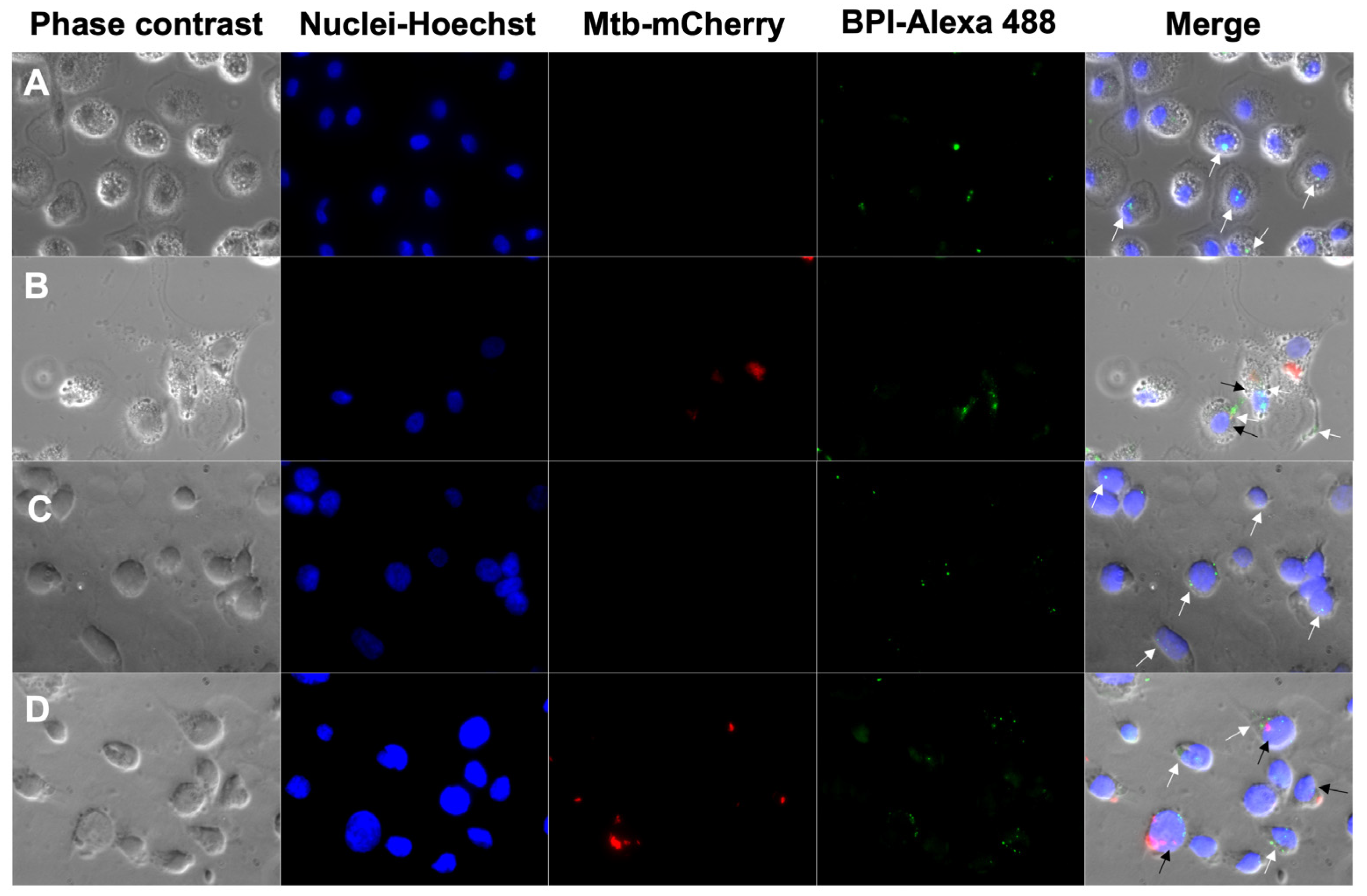
2.3.2. Fluorescence Microscopy
2.4. Intracellular Antimicrobial Activity of BPI
2.5. In Vitro Antimicrobial Activity of BPI
2.6. BPI’s Effect on TNF-α
2.7. Statistical Analyses
3. Results
3.1. Internalization of BPI into Human Macrophages
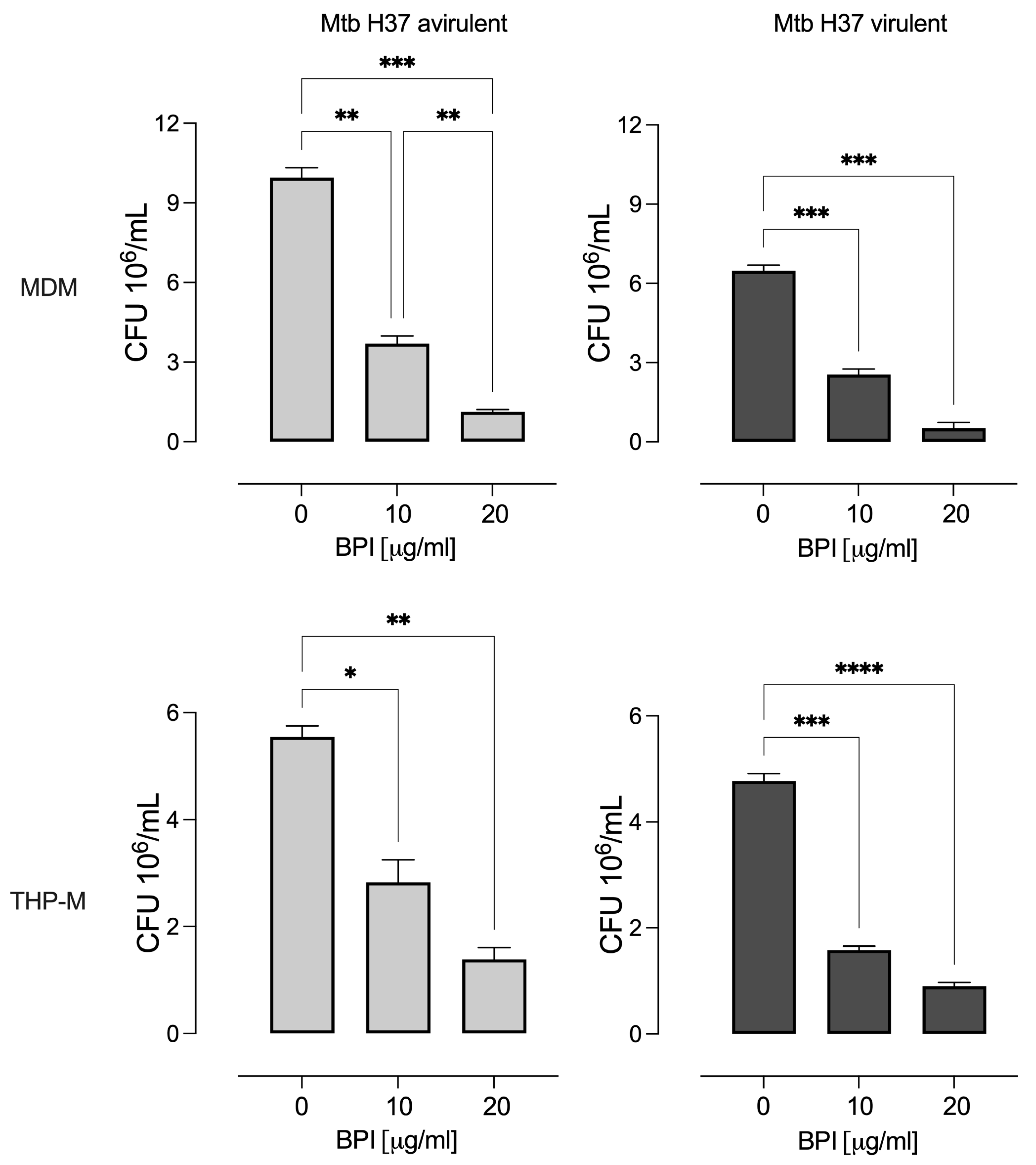
3.2. BPI Inhibits the Intracellular Growth of M. tuberculosis in Macrophages
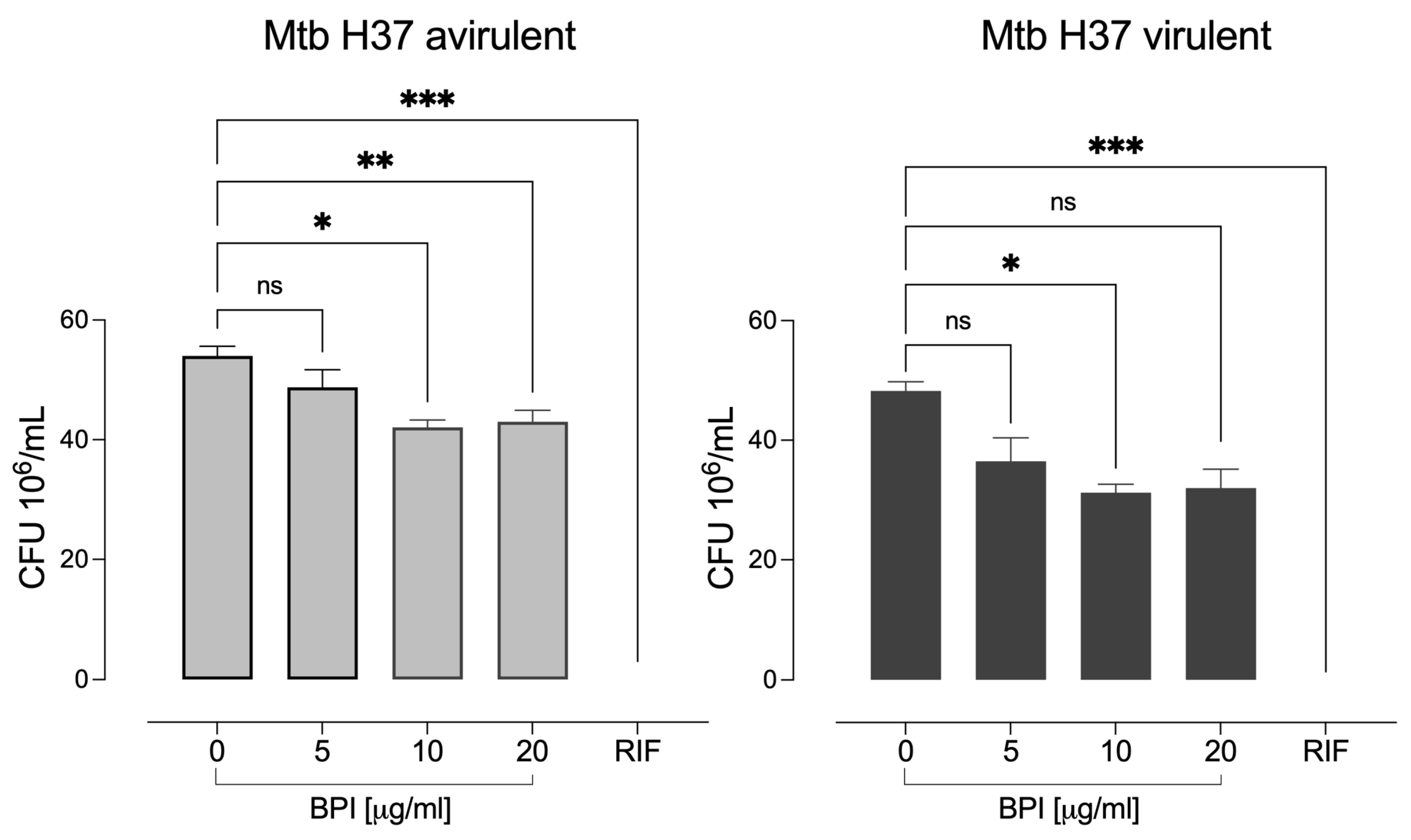
3.3. BPI Shows Inhibitory Activity against M. tuberculosis
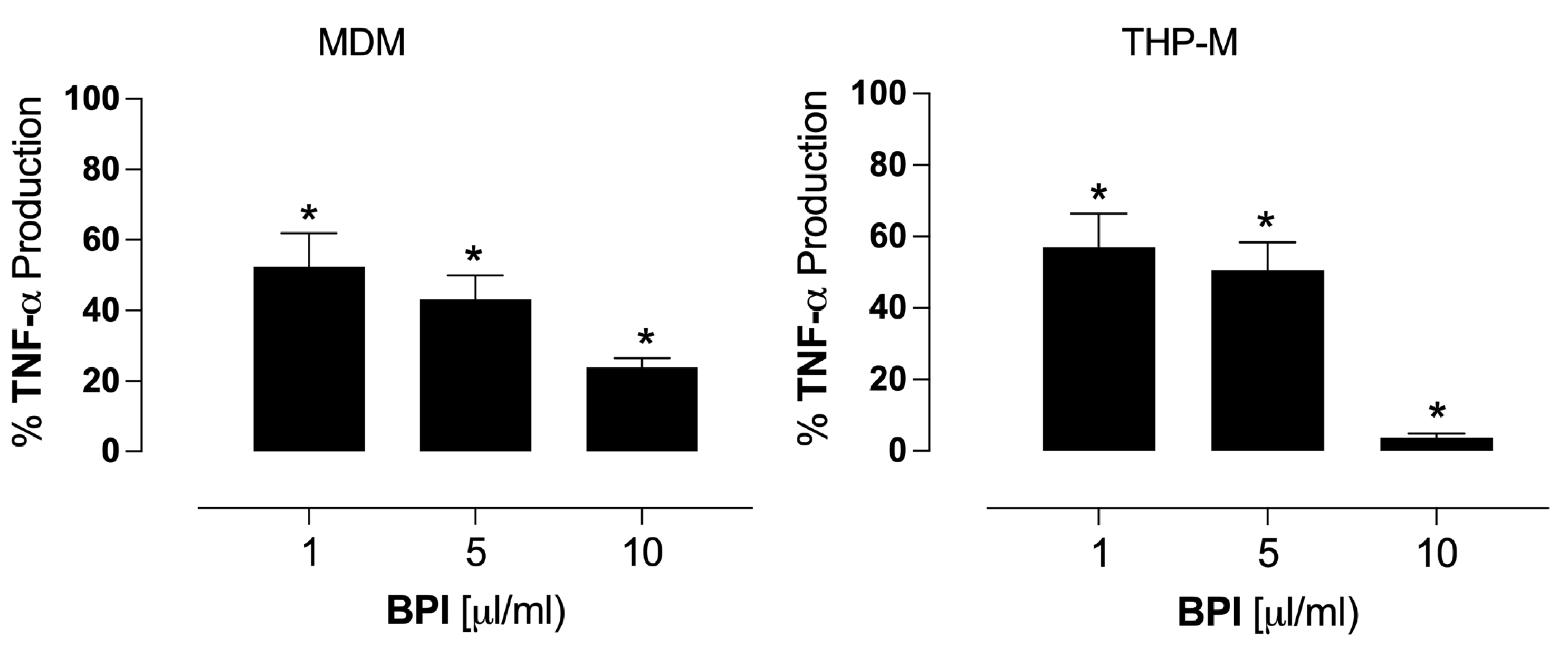
3.4. BPI Inhibits TNF-α Production
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Balakrishnan, A.; Schnare, M.; Chakravortty, D. Of Men Not Mice: Bactericidal/Permeability-Increasing Protein Expressed in Human Macrophages Acts as a Phagocytic Receptor and Modulates Entry and Replication of Gram-Negative Bacteria. Front. Immunol. 2016, 7, 455. [Google Scholar] [CrossRef]
- Weiss, J.; Franson, R.C.; Beckerdite, S.; Schmeidler, K.; Elsbach, P. Partial Characterization and Purification of a Rabbit Granulocyte Factor That Increases Permeability of Escherichia Coli. J. Clin. Investig. 1975, 55, 33–42. [Google Scholar] [CrossRef]
- Iovine, N.M.; Elsbach, P.; Weiss, J. An Opsonic Function of the Neutrophil Bactericidal/Permeability-Increasing Protein Depends on Both Its N- and C-Terminal Domains. Proc. Natl. Acad. Sci. USA 1997, 94, 10973–10978. [Google Scholar] [CrossRef] [PubMed]
- Elsbach, P.; Weiss, J. The Bactericidal/Permeability-Increasing Protein (BPI), a Potent Element in Host-Defense Against Gram-Negative Bacteria and Lipopolysaccharide. Immunobiology 1993, 187, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Theprungsirikul, J.; Skopelja-Gardner, S.; Burns, A.S.; Wierzbicki, R.M.; Rigby, W.F.C. Bactericidal/Permeability-Increasing Protein Preeminently Mediates Clearance of Pseudomonas Aeruginosa In Vivo via CD18-Dependent Phagocytosis. Front. Immunol. 2021, 12, 659523. [Google Scholar] [CrossRef] [PubMed]
- Elsbach, P.; Weiss, J. Role of the Bactericidal/Permeability-Increasing Protein in Host Defence. Curr. Opin. Immunol. 1998, 10, 45–49. [Google Scholar] [CrossRef]
- Balakrishnan, A.; Marathe, S.A.; Joglekar, M.; Chakravortty, D. Bactericidal/Permeability Increasing Protein: A Multifaceted Protein with Functions beyond LPS Neutralization. Innate Immun. 2013, 19, 339–347. [Google Scholar] [CrossRef]
- Beamer, L.J.; Carroll, S.F.; Eisenberg, D. Crystal Structure of Human BPI and Two Bound Phospholipids at 2.4 Angstrom Resolution. Science 1997, 276, 1861–1864. [Google Scholar] [CrossRef]
- Alva, V.; Lupas, A.N. The TULIP Superfamily of Eukaryotic Lipid-Binding Proteins as a Mediator of Lipid Sensing and Transport. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2016, 1861, 913–923. [Google Scholar] [CrossRef]
- Ederer, K.U.; Holzinger, J.M.; Maier, K.T.; Zeller, L.; Werner, M.; Toelge, M.; Gessner, A.; Bülow, S. A Polymorphism of Bactericidal/Permeability-Increasing Protein Affects Its Neutralization Efficiency towards Lipopolysaccharide. Int. J. Mol. Sci. 2022, 23, 1324. [Google Scholar] [CrossRef]
- Ooi, C.E.; Weiss, J.; Doerfler, M.E.; Elsbach, P. Endotoxin-Neutralizing Properties of the 25 KD N-Terminal Fragment and a Newly Isolated 30 KD C-Terminal Fragment of the 55-60 KD Bactericidal/Permeability-Increasing Protein of Human Neutrophils. J. Exp. Med. 1991, 174, 649–655. [Google Scholar] [CrossRef]
- Wang, P.Y.; Kitchens, R.L.; Munford, R.S. Phosphatidylinositides Bind to Plasma Membrane CD14 and Can Prevent Monocyte Activation by Bacterial Lipopolysaccharide. J. Biol. Chem. 1998, 273, 24309–24313. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, H.; Takeuchi, S.; Kubota, K.; Kobayashi, Y.; Kozakai, S.; Ukai, I.; Shichiku, A.; Okubo, M.; Numasaki, M.; Kanemitsu, Y.; et al. Lipopolysaccharide (LPS)-Binding Protein Stimulates CD14-Dependent Toll-like Receptor 4 Internalization and LPS-Induced TBK1-IKKα-IRF3 Axis Activation. J. Biol. Chem. 2018, 293, 10186–10201. [Google Scholar] [CrossRef] [PubMed]
- Gazzano-Santoro, H.; Meszaros, K.; Birr, C.; Carroll, S.F.; Theofan, G.; Horwitz, A.H.; Lim, E.; Aberle, S.; Kasler, H.; Parent, J.B. Competition between RBPI23, a Recombinant Fragment of Bactericidal/Permeability-Increasing Protein, and Lipopolysaccharide (LPS)-Binding Protein for Binding to LPS and Gram-Negative Bacteria. Infect. Immun. 1994, 62, 1185–1191. [Google Scholar] [CrossRef]
- Weiss, J.; Elsbach, P.; Olsson, I.; Odeberg, H. Purification and Characterization of a Potent Bactericidal and Membrane Active Protein from the Granules of Human Polymorphonuclear Leukocytes. J. Biol. Chem. 1978, 253, 2664–2672. [Google Scholar] [CrossRef]
- Holzinger, J.M.; Toelge, M.; Werner, M.; Ederer, K.U.; Siegmund, H.I.; Peterhoff, D.; Blaas, S.H.; Gisch, N.; Brochhausen, C.; Gessner, A.; et al. Scorpionfish BPI Is Highly Active against Multiple Drug-Resistant Pseudomonas Aeruginosa Isolates from People with Cystic Fibrosis. Elife 2023, 12, 86369. [Google Scholar] [CrossRef]
- Chockalingam, A.; McKinney, C.E.; Rinaldi, M.; Zarlenga, D.S.; Bannerman, D.D. A Peptide Derived from Human Bactericidal/Permeability-Increasing Protein (BPI) Exerts Bactericidal Activity against Gram-Negative Bacterial Isolates Obtained from Clinical Cases of Bovine Mastitis. Vet. Microbiol. 2007, 125, 80–90. [Google Scholar] [CrossRef]
- Bülow, S.; Zeller, L.; Werner, M.; Toelge, M.; Holzinger, J.; Entzian, C.; Schubert, T.; Waldow, F.; Gisch, N.; Hammerschmidt, S.; et al. Bactericidal/Permeability-Increasing Protein Is an Enhancer of Bacterial Lipoprotein Recognition. Front. Immunol. 2018, 9, 2768. [Google Scholar] [CrossRef]
- Srivastava, A.; Casey, H.; Johnson, N.; Levy, O.; Malley, R. Recombinant Bactericidal/Permeability-Increasing Protein RBPI21 Protects against Pneumococcal Disease. Infect. Immun. 2007, 75, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Levine, S.M.; Marciniuk, D.D. Global Impact of Respiratory Disease: What Can We Do, Together, to Make a Difference? Chest 2022, 161, 1153–1154. [Google Scholar] [CrossRef]
- Theprungsirikul, J.; Skopelja-Gardner, S.; Rigby, W.F.C. Killing Three Birds with One BPI: Bactericidal, Opsonic, and Anti-Inflammatory Functions. J. Transl. Autoimmun. 2021, 4, 100105. [Google Scholar] [CrossRef] [PubMed]
- Alsayed, S.S.R.; Gunosewoyo, H. Tuberculosis: Pathogenesis, Current Treatment Regimens and New Drug Targets. Int. J. Mol. Sci. 2023, 24, 5202. [Google Scholar] [CrossRef]
- World Health Organization. Available online: https://www.who.int/news-room/fact-sheets/detail/tuberculosis (accessed on 10 January 2024).
- Ruiz, A.; Sarabia, C.; Torres, M.; Juárez, E. Resolvin D1 (RvD1) and Maresin 1 (Mar1) Contribute to Human Macrophage Control of M. Tuberculosis Infection While Resolving Inflammation. Int. Immunopharmacol. 2019, 74, 105694. [Google Scholar] [CrossRef] [PubMed]
- Carroll, P.; Schreuder, L.J.; Muwanguzi-Karugaba, J.; Wiles, S.; Robertson, B.D.; Ripoll, J.; Ward, T.H.; Bancroft, G.J.; Schaible, U.E.; Parish, T. Sensitive Detection of Gene Expression in Mycobacteria under Replicating and Non-Replicating Conditions Using Optimized Far-Red Reporters. PLoS ONE 2010, 5, 9823. [Google Scholar] [CrossRef]
- Guzmán-Beltrán, S.; Rubio-Badillo, M.Á.; Juárez, E.; Hernández-Sánchez, F.; Torres, M. Nordihydroguaiaretic Acid (NDGA) and α-Mangostin Inhibit the Growth of Mycobacterium Tuberculosis by Inducing Autophagy. Int. Immunopharmacol. 2016, 31, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Schön, T.; Werngren, J.; Machado, D.; Borroni, E.; Wijkander, M.; Lina, G.; Mouton, J.; Matuschek, E.; Kahlmeter, G.; Giske, C.; et al. Antimicrobial Susceptibility Testing of Mycobacterium Tuberculosis Complex Isolates—The EUCAST Broth Microdilution Reference Method for MIC Determination. Clin. Microbiol. Infect. 2020, 26, 1488–1492. [Google Scholar] [CrossRef]
- Carranza, C.; Juárez, E.; Torres, M.; Ellner, J.J.; Sada, E.; Schwander, S.K. Mycobacterium Tuberculosis Growth Control by Lung Macrophages and CD8 Cells from Patient Contacts. Am. J. Respir. Crit. Care Med. 2006, 173, 238–245. [Google Scholar] [CrossRef]
- Roca, F.J.; Ramakrishnan, L. TNF Dually Mediates Resistance and Susceptibility to Mycobacteria via Mitochondrial Reactive Oxygen Species. Cell 2013, 153, 521–534. [Google Scholar] [CrossRef]
- Herrera, M.T.; Guzmán-Beltrán, S.; Bobadilla, K.; Santos-Mendoza, T.; Flores-Valdez, M.A.; Gutiérrez-González, L.H.; González, Y. Human Pulmonary Tuberculosis: Understanding the Immune Response in the Bronchoalveolar System. Biomolecules 2022, 12, 1148. [Google Scholar] [CrossRef]
- Burnett, R.J.; Lyden, C.A.; Tindal, C.J.; Cave, C.M.; Marra, M.N.; Solomkin, J.S. Mononuclear Cell Line THP-1 Internalizes Bactericidal/Permeability-Increasing Protein by a Non-Receptor-Mediated Mechanism Consistent with Pinocytosis. Arch. Surg. 1996, 131, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Buccini, D.F.; Cardoso, M.H.; Franco, O.L. Antimicrobial Peptides and Cell-Penetrating Peptides for Treating Intracellular Bacterial Infections. Front. Cell. Infect. Microbiol. 2021, 10, 612931. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.; Olsson, I. Cellular and Subcellular Localization of the Bactericidal/Permeability-Increasing Protein of Neutrophils. Blood 1987, 69, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Skopelja-Gardner, S.; Theprungsirikul, J.; Lewis, K.A.; Hammond, J.H.; Carlson, K.M.; Hazlett, H.F.; Nymon, A.; Nguyen, D.; Berwin, B.L.; Hogan, D.A.; et al. Regulation of Pseudomonas Aeruginosa-Mediated Neutrophil Extracellular Traps. Front. Immunol. 2019, 10, 1670. [Google Scholar] [CrossRef]
- Schultz, H. From Infection to Autoimmunity: A New Model for Induction of ANCA against the Bactericidal/Permeability Increasing Protein (BPI). Autoimmun. Rev. 2007, 6, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Gazzano-Santoro, H.; Parent, J.B.; Grinna, L.; Horwitz, A.; Parsons, T.; Theofan, G.; Elsbach, P.; Weiss, J.; Conlon, P.J. High-Affinity Binding of the Bactericidal/Permeability-Increasing Protein and a Recombinant Amino-Terminal Fragment to the Lipid A Region of Lipopolysaccharide. Infect. Immun. 1992, 60, 4754–4761. [Google Scholar] [CrossRef] [PubMed]
- Wiese, A.; Brandenburg, K.; Carroll, S.F.; Rietschel, E.T.; Seydel, U. Mechanisms of Action of Bactericidal/Permeability-Increasing Protein BPI on Reconstituted Outer Membranes of Gram-Negative Bacteria. Biochemistry 1997, 36, 10311–10319. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, A.H.; Williams, R.E.; Liu, P.S.; Nadell, R. Bactericidal/Permeability-Increasing Protein Inhibits Growth of a Strain of Acholeplasma Laidlawii and L Forms of the Gram-Positive Bacteria Staphylococcus Aureus and Streptococcus Pyogenes. Antimicrob. Agents Chemother. 1999, 43, 2314–2316. [Google Scholar] [CrossRef] [PubMed]
- Bian, Z.C.; Cai, X.H.; Tan, K.A.; Wang, Y.D.; Huang, Z.; Kwan, K.Y.; Xu, P. Identification and Functional Analysis of ToBPI1/LBP and ToBPI2/LBP in Anti-Bacterial Infection of Trachinotus Ovatus. Genes 2023, 14, 826. [Google Scholar] [CrossRef]
- Pinkenburg, O.; Meyer, T.; Bannert, N.; Norley, S.; Bolte, K.; Czudai-Matwich, V.; Herold, S.; Gessner, A.; Schnare, M. The Human Antimicrobial Protein Bactericidal/Permeability-Increasing Protein (BPI) Inhibits the Infectivity of Influenza A Virus. PLoS ONE 2016, 11, 156929. [Google Scholar] [CrossRef]
- Pugin, J.; Heumann, D.; Tomasz, A.; Kravchenko, V.V.; Akamatsu, Y.; Nishijima, M.; Glauser, M.P.; Tobias, P.S.; Ulevitch, R.J. CD14 Is a Pattern Recognition Receptor. Immunity 1994, 1, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Malefane, L.; Maarman, G. Post-Tuberculosis Lung Disease and Inflammatory Role Players: Can We Characterise the Myriad Inflammatory Pathways Involved to Gain a Better Understanding? Chem. Biol. Interact. 2024, 387, 110817. [Google Scholar] [CrossRef] [PubMed]
- Schultz, H.; Weiss, J.P. The Bactericidal/Permeability-Increasing Protein (BPI) in Infection and Inflammatory Disease. Clin. Chim. Acta 2007, 384, 12–23. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guzmán-Beltrán, S.; Juárez, E.; Cruz-Muñoz, B.L.; Páez-Cisneros, C.A.; Sarabia, C.; González, Y. Bactericidal Permeability-Increasing Protein (BPI) Inhibits Mycobacterium tuberculosis Growth. Biomolecules 2024, 14, 475. https://doi.org/10.3390/biom14040475
Guzmán-Beltrán S, Juárez E, Cruz-Muñoz BL, Páez-Cisneros CA, Sarabia C, González Y. Bactericidal Permeability-Increasing Protein (BPI) Inhibits Mycobacterium tuberculosis Growth. Biomolecules. 2024; 14(4):475. https://doi.org/10.3390/biom14040475
Chicago/Turabian StyleGuzmán-Beltrán, Silvia, Esmeralda Juárez, Brenda L. Cruz-Muñoz, Cesar A. Páez-Cisneros, Carmen Sarabia, and Yolanda González. 2024. "Bactericidal Permeability-Increasing Protein (BPI) Inhibits Mycobacterium tuberculosis Growth" Biomolecules 14, no. 4: 475. https://doi.org/10.3390/biom14040475