
Integrated Multi-Omics Analysis of Cerebrospinal Fluid in Postoperative Delirium

 , ,
, ,  , , , ,
, , , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Human Subjects/Study Population
2.2. Delirium Assessment and Matching
2.3. Collection and Processing of Cerebrospinal Fluid
2.4. Proteomics
2.5. Targeted Metabolomics and Untargeted Lipidomics
2.6. Statistical Analysis
2.7. Systems Biology
2.8. Multi-Omics Integration
2.9. Machine Learning
3. Results
3.1. Sample Characteristics
3.2. Molecules Altered in the Delirium Group at PREOP
3.3. Pathway Enrichment Analysis of Lipids and Metabolites
3.4. Signature Prediction of Delirium with Machine Learning
3.5. Multi-Omics Integration
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Inouye, S.K.; Westendorp, R.G.; Saczynski, J.S. Delirium in elderly people. Lancet 2014, 383, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Marcantonio, E.R. Postoperative delirium: A 76-year-old woman with delirium following surgery. JAMA 2012, 308, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Gou, R.Y.; Hshieh, T.T.; Marcantonio, E.R.; Cooper, Z.; Jones, R.N.; Travison, T.G.; Fong, T.G.; Abdeen, A.; Lange, J.; Earp, B.; et al. One-year Medicare costs associated with delirium in older patients undergoing major elective surgery. JAMA Surg. 2021, 156, 430–442. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, J.R. Delirium pathophysiology: An updated hypothesis of the etiology of acute brain failure. Int. J. Geriatr. Psychiatry 2018, 33, 1428–1457. [Google Scholar] [CrossRef]
- Maldonado, J.R. Neuropathogenesis of delirium: Review of current etiologic theories and common pathways. Am. J. Geriatr. Psychiatry 2013, 21, 1190–1222. [Google Scholar] [CrossRef] [PubMed]
- Damkier, H.H.; Brown, P.D.; Praetorius, J. Epithelial pathways in choroid plexus electrolyte transport. Physiology 2010, 25, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Sakka, L.; Coll, G.; Chazal, J. Anatomy and physiology of cerebrospinal fluid. Eur. Ann. Otorhinolaryngol. Head. Neck Dis. 2011, 128, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Lun, M.P.; Monuki, E.S.; Lehtinen, M.K. Development and functions of the choroid plexus-cerebrospinal fluid system. Nat. Rev. Neurosci. 2015, 16, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Spector, R.; Robert Snodgrass, S.; Johanson, C.E. A balanced view of the cerebrospinal fluid composition and functions: Focus on adult humans. Exp. Neurol. 2015, 273, 57–68. [Google Scholar] [CrossRef]
- Aslam, B.; Basit, M.; Nisar, M.A.; Khurshid, M.; Rasool, M.H. Proteomics: Technologies and their applications. J. Chromatogr. Sci. 2017, 55, 182–196. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics by gas chromatography-mass spectrometry: Combined targeted and untargeted profiling. Curr. Protoc. Mol. Biol. 2016, 114, 30.34.31–30.34.32. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Zhang, J.L. Mass-spectrometry-based lipidomics. J. Sep. Sci. 2018, 41, 351–372. [Google Scholar] [CrossRef]
- Tracey, T.J.; Kirk, S.E.; Steyn, F.J.; Ngo, S.T. The role of lipids in the central nervous system and their pathological implications in amyotrophic lateral sclerosis. Semin. Cell Dev. Biol. 2021, 112, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zhang, W.; Liu, J.; Song, Y.; Liu, T.; Li, Z.; Wang, X.; Yang, N.; Li, Y.; Han, D.; et al. Metabolomic and lipidomic profiling of preoperative CSF in elderly hip fracture patients with postoperative delirium. Front. Aging Neurosci. 2020, 12, 570210. [Google Scholar] [CrossRef]
- Tripp, B.A.; Dillon, S.T.; Yuan, M.; Asara, J.M.; Vasunilashorn, S.M.; Fong, T.G.; Metzger, E.D.; Inouye, S.K.; Xie, Z.; Ngo, L.H.; et al. Targeted metabolomics analysis of postoperative delirium. Sci. Rep. 2021, 11, 1521. [Google Scholar] [CrossRef] [PubMed]
- Watne, L.O.; Idland, A.V.; Fekkes, D.; Raeder, J.; Frihagen, F.; Ranhoff, A.H.; Chaudhry, F.A.; Engedal, K.; Wyller, T.B.; Hassel, B. Increased CSF levels of aromatic amino acids in hip fracture patients with delirium suggests higher monoaminergic activity. BMC Geriatr. 2016, 16, 149. [Google Scholar] [CrossRef] [PubMed]
- Poljak, A.; Hill, M.; Hall, R.J.; MacLullich, A.M.; Raftery, M.J.; Tai, J.; Yan, S.; Caplan, G.A. Quantitative proteomics of delirium cerebrospinal fluid. Transl. Psychiatry 2014, 4, e477. [Google Scholar] [CrossRef]
- Vasunilashorn, S.M.; Ngo, L.H.; Chan, N.Y.; Zhou, W.; Dillon, S.T.; Otu, H.H.; Inouye, S.K.; Wyrobnik, I.; Kuchel, G.A.; McElhaney, J.E.; et al. Development of a dynamic multi-protein signature of postoperative delirium. J. Gerontol. A Biol. Sci. Med. Sci. 2019, 74, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Vasunilashorn, S.M.; Dillon, S.T.; Chan, N.Y.; Fong, T.G.; Joseph, M.; Tripp, B.; Xie, Z.; Ngo, L.H.; Lee, C.G.; Elias, J.A.; et al. Proteome-wide analysis using SOMAscan identifies and validates chitinase-3-like protein 1 as a risk and disease marker of delirium among older adults undergoing major elective surgery. J. Gerontol. A Biol. Sci. Med. Sci. 2022, 77, 484–493. [Google Scholar] [CrossRef]
- Xie, Z.; McAuliffe, S.; Swain, C.A.; Ward, S.A.; Crosby, C.A.; Zheng, H.; Sherman, J.; Dong, Y.; Zhang, Y.; Sunder, N.; et al. Cerebrospinal fluid aβ to tau ratio and postoperative cognitive change. Ann. Surg. 2013, 258, 364–369. [Google Scholar] [CrossRef]
- Hirsch, J.; Vacas, S.; Terrando, N.; Yuan, M.; Sands, L.P.; Kramer, J.; Bozic, K.; Maze, M.M.; Leung, J.M. Perioperative cerebrospinal fluid and plasma inflammatory markers after orthopedic surgery. J. Neuroinflammation 2016, 13, 211. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Tang, J.; Liu, C.; Li, X.; Cao, X.; Wang, B.; Dong, R.; Xu, W.; Yu, X.; Wang, M.; et al. Cerebrospinal fluid cholinergic biomarkers are associated with postoperative delirium in elderly patients undergoing Total hip/knee replacement: A prospective cohort study. BMC Anesthesiol. 2020, 20, 246. [Google Scholar] [CrossRef] [PubMed]
- Folstein, M.F.; Folstein, S.E.; McHugh, P.R. “Mini-mental state”. A practical method for grading the cognitive state of patients for the clinician. J. Psychiatr. Res. 1975, 12, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Teng, E.L.; Chui, H.C. The modified mini-mental state (3MS) examination. J. Clin. Psychiatry 1987, 48, 314–318. [Google Scholar] [PubMed]
- Inouye, S.K.; van Dyck, C.H.; Alessi, C.A.; Balkin, S.; Siegal, A.P.; Horwitz, R.I. Clarifying confusion: The confusion assessment method. A new method for detection of delirium. Ann. Intern. Med. 1990, 113, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Albert, M.S.; Levkoff, S.E.; Reilly, C.; Liptzin, B.; Pilgrim, D.; Cleary, P.D.; Evans, D.; Rowe, J.W. The delirium symptom interview: An interview for the detection of delirium symptoms in hospitalized patients. J. Geriatr. Psychiatry Neurol. 1992, 5, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Inouye, S.K.; Marcantonio, E.R.; Kosar, C.M.; Tommet, D.; Schmitt, E.M.; Travison, T.G.; Saczynski, J.S.; Ngo, L.H.; Alsop, D.C.; Jones, R.N. The short-term and long-term relationship between delirium and cognitive trajectory in older surgical patients. Alzheimers Dement. 2016, 12, 766–775. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, P.R. Optimal matching for observational studies. J. Am. Stat. Assoc. 1989, 84, 1024–1032. [Google Scholar] [CrossRef]
- Ngo, L.H.; Inouye, S.K.; Jones, R.N.; Travison, T.G.; Libermann, T.A.; Dillon, S.T.; Kuchel, G.A.; Vasunilashorn, S.M.; Alsop, D.C.; Marcantonio, E.R. Methodologic considerations in the design and analysis of nested case-control studies: Association between cytokines and postoperative delirium. BMC Med. Res. Methodol. 2017, 17, 88. [Google Scholar] [CrossRef]
- Shubin, A.V.; Kollar, B.; Dillon, S.T.; Pomahac, B.; Libermann, T.A.; Riella, L.V. Blood proteome profiling using aptamer-based technology for rejection biomarker discovery in transplantation. Sci. Data 2019, 6, 314. [Google Scholar] [CrossRef]
- Dillon, S.T.; Vasunilashorn, S.M.; Otu, H.H.; Ngo, L.; Fong, T.; Gu, X.; Cavallari, M.; Touroutoglou, A.; Shafi, M.; Inouye, S.K.; et al. Aptamer-based proteomics measuring preoperative cerebrospinal fluid protein alterations associated with postoperative delirium. Biomolecules 2023, 13, 1395. [Google Scholar] [CrossRef] [PubMed]
- Gold, L.; Walker, J.J.; Wilcox, S.K.; Williams, S. Advances in human proteomics at high scale with the SOMAscan proteomics platform. N. Biotechnol. 2012, 29, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Candia, J.; Cheung, F.; Kotliarov, Y.; Fantoni, G.; Sellers, B.; Griesman, T.; Huang, J.; Stuccio, S.; Zingone, A.; Ryan, B.M.; et al. Assessment of variability in the SOMAscan assay. Sci. Rep. 2017, 7, 14248. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Breitkopf, S.B.; Yang, X.; Asara, J.M. A positive/negative ion-switching, targeted mass spectrometry-based metabolomics platform for bodily fluids, cells, and fresh and fixed tissue. Nat. Protoc. 2012, 7, 872–881. [Google Scholar] [CrossRef] [PubMed]
- Luan, H.; Ji, F.; Chen, Y.; Cai, Z. statTarget: A streamlined tool for signal drift correction and interpretations of quantitative mass spectrometry-based omics data. Anal. Chim. Acta 2018, 1036, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.; Hart, P. Nearest neighbor pattern classification. IEEE Trans. Inf. Theory 1967, 13, 21–27. [Google Scholar] [CrossRef]
- De Livera, A.M.; Olshansky, G.; Simpson, J.A.; Creek, D.J. NormalizeMets: Assessing, selecting and implementing statistical methods for normalizing metabolomics data. Metabolomics 2018, 14, 54. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Society. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Hoaglin, D. Understanding Robust and Exploratory Data Analysis; Taylor & Francis: Boca Raton, FL, USA, 2001. [Google Scholar]
- Xia, J.; Wishart, D.S. Using MetaboAnalyst 3.0 for comprehensive metabolomics data analysis. Curr. Protoc. Bioinform. 2016, 55, 14.10.11–14.10.91. [Google Scholar] [CrossRef]
- Lipid Pathway Enrichment Analysis (LIPEA). Available online: https://hyperlipea.org (accessed on 18 October 2020).
- Tripp, B.; Otu, H.H. Integration of multi-omics data using probabilistic graph models and external knowledge. Curr. Bioinform. 2021, 16, 1–11. [Google Scholar] [CrossRef]
- Chanumolu, S.K.; Albahrani, M.; Can, H.; Otu, H.H. KEGG2Net: Deducing gene interaction networks and acyclic graphs from KEGG pathways. EMBnetjournal 2021, 26, e949. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Brunk, E.; Sahoo, S.; Zielinski, D.C.; Altunkaya, A.; Drager, A.; Mih, N.; Gatto, F.; Nilsson, A.; Preciat Gonzalez, G.A.; Aurich, M.K.; et al. Recon3D enables a three-dimensional view of gene variation in human metabolism. Nat. Biotechnol. 2018, 36, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Aimo, L.; Liechti, R.; Hyka-Nouspikel, N.; Niknejad, A.; Gleizes, A.; Götz, L.; Kuznetsov, D.; David, F.P.; van der Goot, F.G.; Riezman, H.; et al. The SwissLipids knowledgebase for lipid biology. Bioinformatics 2015, 31, 2860–2866. [Google Scholar] [CrossRef] [PubMed]
- Friedman, N.; Goldszmidt, M.; Wyner, A.J. Data Analysis with Bayesian Networks: A Bootstrap Approach. arXiv 1999, arXiv:1301.6695v1. [Google Scholar]
- Scutari, M.; Nagarajan, R. Identifying significant edges in graphical models of molecular networks. Artif. Intell. Med. 2013, 57, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Sneath, P.H.A.; Sokal, R.R. Numerical Taxonomy: The Principles and Practice of Numerical Classification; W. H. Freeman: San Francisco, CA, USA, 1973. [Google Scholar]
- Pearson, K. On lines and planes of closest fit to systems of points in space. Lond. Edinb. Dublin Philos. Mag. J. Sci. 1901, 2, 559–572. [Google Scholar] [CrossRef]
- Hsu, C.-W.; Lin, C.-J. A comparison of methods for multiclass support vector machines. IEEE Trans. Neural Netw. 2002, 13, 415–425. [Google Scholar] [CrossRef]
- Farrell, A.; Wang, G.; Rush, S.A.; Martin, J.A.; Belant, J.L.; Butler, A.B.; Godwin, D. Machine learning of large-scale spatial distributions of wild turkeys with high-dimensional environmental data. Ecol. Evol. 2019, 9, 5938–5949. [Google Scholar] [CrossRef]
- Rau, A.; Manansala, R.; Flister, M.J.; Rui, H.; Jaffrézic, F.; Laloë, D.; Auer, P.L. Individualized multi-omic pathway deviation scores using multiple factor analysis. Biostatistics 2022, 23, 362–379. [Google Scholar] [CrossRef]
- Zou, H.; Hastie, T. Regularization and variable selection via the Elastic Net. J. R. Stat. Soc. Ser. B Stat. Methodol. 2005, 67, 301–320. [Google Scholar] [CrossRef]
- Sammut, C.; Webb, G.I. Leave-one-out cross-validation. In Encyclopedia of Machine Learning; Sammut, C., Webb, G.I., Eds.; Springer US: Boston, MA, USA, 2010; pp. 600–601. [Google Scholar]
- Bradley, A.P. The use of the area under the ROC curve in the evaluation of machine learning algorithms. Pattern Recognit. 1997, 30, 1145–1159. [Google Scholar] [CrossRef]
- Gao, J.; Xu, K.; Liu, H.; Liu, G.; Bai, M.; Peng, C.; Li, T.; Yin, Y. Impact of the gut microbiota on intestinal immunity mediated by tryptophan metabolism. Front. Cell Infect. Microbiol. 2018, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Palfrey, H.A.; Pathak, R.; Kadowitz, P.J.; Gettys, T.W.; Murthy, S.N. The metabolism and significance of homocysteine in nutrition and health. Nutr. Metab. 2017, 14, 78. [Google Scholar] [CrossRef] [PubMed]
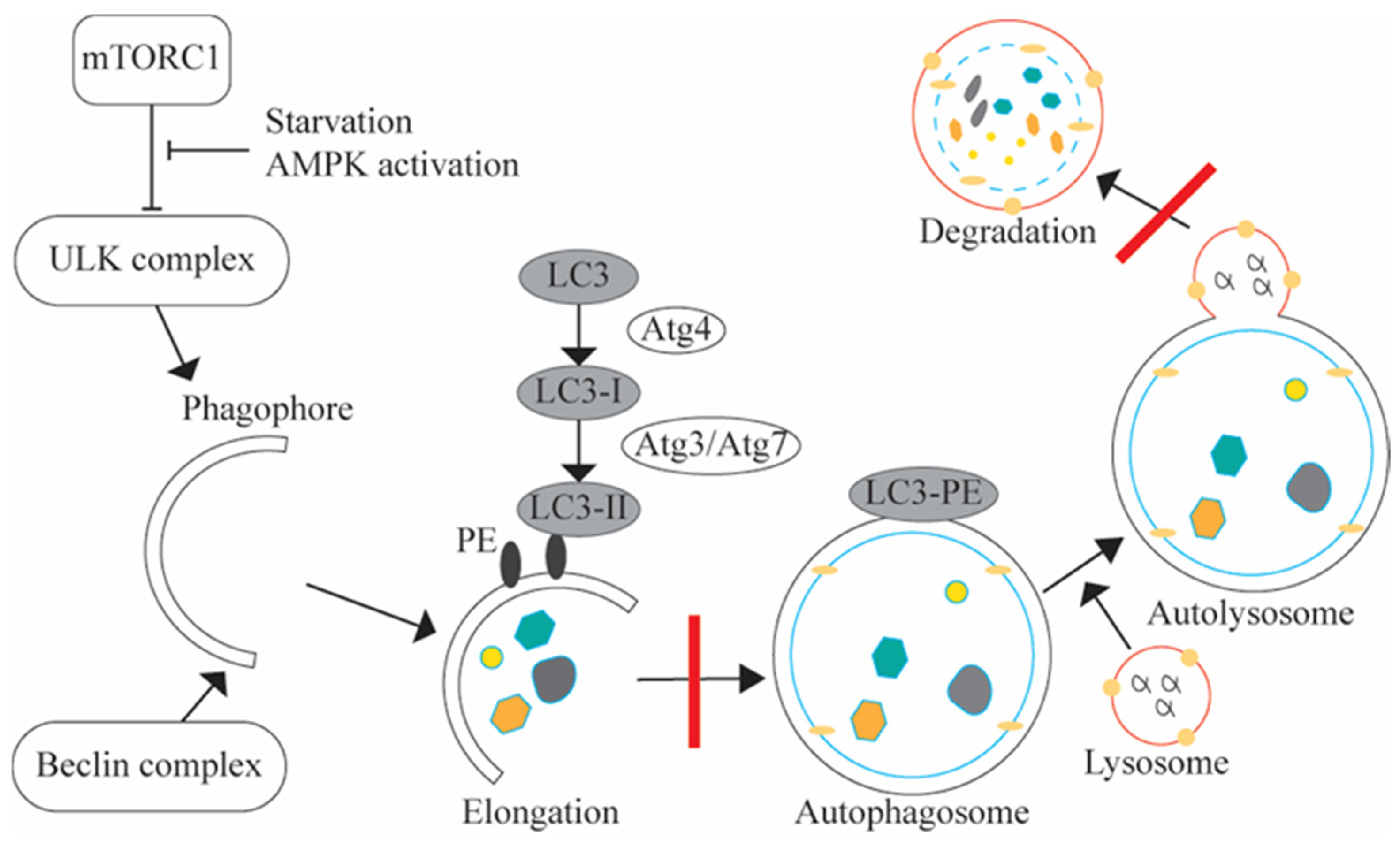
- Hernandez-Diaz, S.; Soukup, S.F. The role of lipids in autophagy and its implication in neurodegeneration. Cell Stress 2020, 4, 167–186. [Google Scholar] [CrossRef] [PubMed]
- Graham, S.F.; Chevallier, O.P.; Elliott, C.T.; Hölscher, C.; Johnston, J.; McGuinness, B.; Kehoe, P.G.; Passmore, A.P.; Green, B.D. Untargeted metabolomic analysis of human plasma indicates differentially affected polyamine and L-arginine metabolism in mild cognitive impairment subjects converting to Alzheimer’s disease. PLoS ONE 2015, 10, e0119452. [Google Scholar] [CrossRef] [PubMed]
- Ravaglia, G.; Forti, P.; Maioli, F.; Bianchi, G.; Martelli, M.; Talerico, T.; Servadei, L.; Zoli, M.; Mariani, E. Plasma amino acid concentrations in patients with amnestic mild cognitive impairment or Alzheimer disease. Am. J. Clin. Nutr. 2004, 80, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Amand, J.; Fehlmann, T.; Backes, C.; Keller, A. DynaVenn: Web-based computation of the most significant overlap between ordered sets. BMC Bioinform. 2019, 20, 743. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.; Charych, E.; Lee, A.; Möller, T. Kynurenines in CNS disease: Regulation by inflammatory cytokines. Front. Neurosci. 2014, 8, 12. [Google Scholar] [CrossRef]
- Loretz, N.; Becker, C.; Hochstrasser, S.; Metzger, K.; Beck, K.; Mueller, J.; Gross, S.; Vincent, A.; Amacher, S.A.; Sutter, R.; et al. Activation of the kynurenine pathway predicts mortality and neurological outcome in cardiac arrest patients: A validation study. J. Crit. Care 2022, 67, 57–65. [Google Scholar] [CrossRef]
- Lim, C.K.; Brew, B.J.; Sundaram, G.; Guillemin, G.J. Understanding the roles of the kynurenine pathway in multiple sclerosis progression. Int. J. Tryptophan Res. 2010, 3, 157–167. [Google Scholar] [CrossRef]
- Gulaj, E.; Pawlak, K.; Bien, B.; Pawlak, D. Kynurenine and its metabolites in Alzheimer’s disease patients. Adv. Med. Sci. 2010, 55, 204–211. [Google Scholar] [CrossRef]
- Teruya, T.; Chen, Y.J.; Kondoh, H.; Fukuji, Y.; Yanagida, M. Whole-blood metabolomics of dementia patients reveal classes of disease-linked metabolites. Proc. Natl. Acad. Sci. USA 2021, 118, e2022857118. [Google Scholar] [CrossRef] [PubMed]
- Min, S.W.; Chen, X.; Tracy, T.E.; Li, Y.; Zhou, Y.; Wang, C.; Shirakawa, K.; Minami, S.S.; Defensor, E.; Mok, S.A.; et al. Critical role of acetylation in tau-mediated neurodegeneration and cognitive deficits. Nat. Med. 2015, 21, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Tracy, T.E.; Gan, L. Acetylated tau in Alzheimer’s disease: An instigator of synaptic dysfunction underlying memory loss: Increased levels of acetylated tau blocks the postsynaptic signaling required for plasticity and promotes memory deficits associated with tauopathy. Bioessays 2017, 39, 1600224. [Google Scholar] [CrossRef]
- Connell, E.; Le Gall, G.; Pontifex, M.G.; Sami, S.; Cryan, J.F.; Clarke, G.; Müller, M.; Vauzour, D. Microbial-derived metabolites as a risk factor of age-related cognitive decline and dementia. Mol. Neurodegener. 2022, 17, 43. [Google Scholar] [CrossRef]
- Herrmann, K.M.; Weaver, L.M. The shikimate pathway. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 473–503. [Google Scholar] [CrossRef]
- Jenkins, T.A.; Nguyen, J.C.D.; Polglaze, K.E.; Bertrand, P.P. Influence of tryptophan and serotonin on mood and cognition with a possible role of the gut-brain axis. Nutrients 2016, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Garcez, F.B.; Garcia de Alencar, J.C.; Fernandez, S.S.M.; Avelino-Silva, V.I.; Sabino, E.C.; Martins, R.C.R.; Franco, L.A.M.; Lima Ribeiro, S.M.; Possolo de Souza, H.; Avelino-Silva, T.J. Association between gut microbiota and delirium in acutely ill older adults. J. Gerontol. Ser. A 2023, 78, 1320–1327. [Google Scholar] [CrossRef]
- Zhang, Y.; Baldyga, K.; Dong, Y.; Song, W.; Villanueva, M.; Deng, H.; Mueller, A.; Houle, T.T.; Marcantonio, E.R.; Xie, Z. The association between gut microbiota and postoperative delirium in patients. Transl. Psychiatry 2023, 13, 156. [Google Scholar] [CrossRef]
- Ticinesi, A.; Parise, A.; Nouvenne, A.; Cerundolo, N.; Prati, B.; Meschi, T. The possible role of gut microbiota dysbiosis in the pathophysiology of delirium in older persons. Microbiome Res. Rep. 2023, 2, 19. [Google Scholar] [CrossRef] [PubMed]
- Tynkkynen, J.; Chouraki, V.; van der Lee, S.J.; Hernesniemi, J.; Yang, Q.; Li, S.; Beiser, A.; Larson, M.G.; Sääksjärvi, K.; Shipley, M.J.; et al. Association of branched-chain amino acids and other circulating metabolites with risk of incident dementia and Alzheimer’s disease: A prospective study in eight cohorts. Alzheimers Dement. 2018, 14, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Dayon, L.; Guiraud, S.P.; Corthésy, J.; Da Silva, L.; Migliavacca, E.; Tautvydaitė, D.; Oikonomidi, A.; Moullet, B.; Henry, H.; Métairon, S.; et al. One-carbon metabolism, cognitive impairment and CSF measures of Alzheimer pathology: Homocysteine and beyond. Alzheimers Res. Ther. 2017, 9, 43. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.D.; Refsum, H.; Bottiglieri, T.; Fenech, M.; Hooshmand, B.; McCaddon, A.; Miller, J.W.; Rosenberg, I.H.; Obeid, R. Homocysteine and dementia: An international consensus statement. J. Alzheimers Dis. 2018, 62, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Mei, X.; Tang, T.; Wang, M.; Wei, X.; Zheng, H.; Cao, J.; Zheng, H.; Cody, K.; Xiong, L.; et al. Preoperative homocysteine modifies the association between postoperative C-reactive protein and postoperative delirium. Front. Aging Neurosci. 2022, 14, 963421. [Google Scholar] [CrossRef] [PubMed]
- Codogno, P.; Mehrpour, M.; Proikas-Cezanne, T. Canonical and non-canonical autophagy: Variations on a common theme of self-eating? Nat. Rev. Mol. Cell Biol. 2012, 13, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Dang, X.; Huan, X.; Du, X.; Chen, X.; Bi, M.; Yan, C.; Jiao, Q.; Jiang, H. Correlation of ferroptosis and other types of cell death in neurodegenerative diseases. Neurosci. Bull. 2022, 38, 938–952. [Google Scholar] [CrossRef] [PubMed]
- Castillo, P.E.; Younts, T.J.; Chávez, A.E.; Hashimotodani, Y. Endocannabinoid signaling and synaptic function. Neuron 2012, 76, 70–81. [Google Scholar] [CrossRef]
- Wallace, M.; Morris, C.; O’Grada, C.M.; Ryan, M.; Dillon, E.T.; Coleman, E.; Gibney, E.R.; Gibney, M.J.; Roche, H.M.; Brennan, L. Relationship between the lipidome, inflammatory markers and insulin resistance. Mol. BioSystems 2014, 10, 1586–1595. [Google Scholar] [CrossRef]
- Vasunilashorn, S.M.; Dillon, S.T.; Inouye, S.K.; Ngo, L.H.; Fong, T.G.; Jones, R.N.; Travison, T.G.; Schmitt, E.M.; Alsop, D.C.; Freedman, S.D.; et al. High c-reactive protein predicts delirium incidence, duration, and feature severity after major non-cardiac surgery. J. Am. Geriatr. Soc. 2017, 65, e109–e116. [Google Scholar] [CrossRef]
- Dillon, S.T.; Vasunilashorn, S.M.; Ngo, L.; Otu, H.H.; Inouye, S.K.; Jones, R.N.; Alsop, D.C.; Kuchel, G.A.; Metzger, E.D.; Arnold, S.E.; et al. Higher c-reactive protein levels predict postoperative delirium in older patients undergoing major elective surgery: A longitudinal nested case-control study. Biol. Psychiatry 2017, 81, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Vasunilashorn, S.M.; Ngo, L.; Inouye, S.K.; Libermann, T.A.; Jones, R.N.; Alsop, D.C.; Guess, J.; Jastrzebski, S.; McElhaney, J.E.; Kuchel, G.A.; et al. Cytokines and postoperative delirium in older patients undergoing major elective surgery. J. Gerontol. A Biol. Sci. Med. Sci. 2015, 70, 1289–1295. [Google Scholar] [CrossRef] [PubMed]
- Kar, A.; Mehrotra, S.; Chatterjee, S. CD38: T cell immuno-metabolic modulator. Cells 2020, 9, 1716. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Huang, Y.; Xing, S.; Chen, C.; Shen, D.; Chen, J. Aβ promotes CD38 expression in senescent microglia in Alzheimer’s disease. Biol. Res. 2022, 55, 10. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, S.; Privat, A.L.; Bressac, L.; Toulorge, D. CD38 in neurodegeneration and neuroinflammation. Cells 2020, 9, 471. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.L. Neuroprotection by glucose-6-phosphate dehydrogenase and the pentose phosphate pathway. J. Cell Biochem. 2019, 120, 14285–14295. [Google Scholar] [CrossRef]
- Tu, D.; Gao, Y.; Yang, R.; Guan, T.; Hong, J.S.; Gao, H.M. The pentose phosphate pathway regulates chronic neuroinflammation and dopaminergic neurodegeneration. J. Neuroinflammation 2019, 16, 255. [Google Scholar] [CrossRef]
- Stocki, P.; Chapman, D.C.; Beach, L.A.; Williams, D.B. Depletion of cyclophilins B and C leads to dysregulation of endoplasmic reticulum redox homeostasis. J. Biol. Chem. 2014, 289, 23086–23096. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, M.; Zhang, C.W.; Singh, B.K.; Sinha, R.A.; Moe, K.T.; DeSilva, D.A.; Yen, P.M. Hyperhomocysteinemia causes ER stress and impaired autophagy that is reversed by Vitamin B supplementation. Cell Death Dis. 2016, 7, e2513. [Google Scholar] [CrossRef]
- Fassbender, K.; Mielke, O.; Bertsch, T.; Nafe, B.; Fröschen, S.; Hennerici, M. Homocysteine in cerebral macroangiography and microangiopathy. Lancet 1999, 353, 1586–1587. [Google Scholar] [CrossRef]
- Diaz-Arrastia, R. Homocysteine and neurologic disease. Arch. Neurol. 2000, 57, 1422–1427. [Google Scholar] [CrossRef] [PubMed]
- Price, B.R.; Wilcock, D.M.; Weekman, E.M. Hyperhomocysteinemia as a risk factor for vascular contributions to cognitive impairment and dementia. Front. Aging Neurosci. 2018, 10, 350. [Google Scholar] [CrossRef] [PubMed]
- Evers, S.; Koch, H.G.; Grotemeyer, K.H.; Lange, B.; Deufel, T.; Ringelstein, E.B. Features, symptoms, and neurophysiological findings in stroke associated with hyperhomocysteinemia. Arch. Neurol. 1997, 54, 1276–1282. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.E.; Mart, M.F.; Cunningham, C.; Shehabi, Y.; Girard, T.D.; MacLullich, A.M.J.; Slooter, A.J.C.; Ely, E.W. Delirium. Nat. Rev. Dis. Primers 2020, 6, 90. [Google Scholar] [CrossRef]
- Fong, T.G.; Inouye, S.K. The inter-relationship between delirium and dementia: The importance of delirium prevention. Nat. Rev. Neurol. 2022, 18, 579–596. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipids | Metabolites |
|---|---|
| LPE (17:1) | L-Tryptophan_D3_pos |
| PE (15:0) (18:1) | DL-Valine_D8_pos |
| PG (15:0) (18:1) | DL-Alanine_D3_pos |
| PI (15:0) (18:1) | DL-Alanine_D3_pos |
| (a) | |||||||
| Metabolite | p-Value | BH p-Value | tFC | Metabolite | p-Value | BH p-Value | tFC |
| deoxyadenosine | 0.0008 | 0.0557 | 2.46 | ornithine | 0.0481 | 0.2451 | 1.27 |
| shikimate 1 | 0.0154 | 0.2114 | 2.25 | D-gluconate | 0.0481 | 0.2451 | 1.26 |
| indoleacrylic acid 1 | 0.0302 | 0.2451 | 2.02 | kynurenine | 0.0334 | 0.2451 | 1.25 |
| S-adenosyl-L-homocysteine | 0.0005 | 0.0540 | 1.97 | deoxyinosine | 0.0481 | 0.2451 | 1.25 |
| methylnicotinamide | 0.0481 | 0.2451 | 1.84 | acetyllysine | 0.0348 | 0.2451 | 1.25 |
| anthranilate 1 | 0.0210 | 0.2451 | 1.81 | N-acetyl-glutamate | 0.0007 | 0.0632 | 1.23 |
| 1,3-diphopshateglycerate | 0.0002 | 0.0418 | 1.80 | D-sedoheptulose-1-7-phosphate 1 | 0.0160 | 0.2180 | 1.25 |
| N-acetyl spermidine | 0.0077 | 0.1290 | 1.70 | aspartate | 0.0481 | 0.2451 | 1.17 |
| glucose-6-phosphate | 0.0050 | 0.1175 | 1.66 | 2-hydroxygluterate | 0.0067 | 0.1175 | 1.16 |
| hydroxyphenylacetic acid 1 | 0.0481 | 0.2451 | 1.64 | Succinate 1 | 0.0481 | 0.2451 | 1.14 |
| 7-methylguanosine | 0.0035 | 0.1175 | 1.62 | phenylpropiolic acid | 0.0023 | 0.1023 | 1.13 |
| S-ribosyl-L-homocysteine 1 | 0.0059 | 0.1175 | 1.58 | 2-oxo-4-methylthiobutanoate | 0.0038 | 0.1376 | 1.13 |
| fructose-6-phosphate | 0.0052 | 0.1175 | 1.56 | deoxyribose-phosphate | 0.0342 | 0.3151 | 1.13 |
| SBP | 0.0481 | 0.2451 | 1.49 | arginine | 0.0481 | 0.2451 | 1.11 |
| D-glucono-delta-lactone-6-phosphate | 0.0481 | 0.2451 | 1.48 | methylmalonic acid | 0.0033 | 0.1175 | 1.10 |
| 3-methylphenylacetic acid 1 | 0.0077 | 0.1462 | 1.45 | asparagine | 0.0154 | 0.2114 | 1.09 |
| nicotinamide riboside | 0.0431 | 0.1462 | 1.45 | dGMP | 0.0290 | 0.2857 | 1.01 |
| uracil | 0.0070 | 0.1175 | 1.41 | trehalose-sucrose | 0.0389 | 0.2451 | 0.90 |
| 1-Methyladenosine 1 | 0.0135 | 0.1847 | 1.45 | 3-hydroxybuterate | 0.0485 | 0.3365 | −0.81 |
| guanine | 0.0481 | 0.2451 | 1.39 | dTTP | 0.0432 | 0.3269 | −1.13 |
| uridine | 0.0054 | 0.1175 | 1.37 | alanine | 0.0481 | 0.2451 | −1.15 |
| carbamoyl phosphate | 0.0240 | 0.2749 | 1.33 | dihydroxy-acetone-phosphate | 0.0481 | 0.2451 | −1.43 |
| homoserine | 0.0481 | 0.2451 | 1.32 | 2-deoxyglucose-6-phosphate | 0.0481 | 0.2451 | −1.46 |
| ribose-phosphate | 0.0328 | 0.2451 | 1.31 | N-Acetylputrescine | 0.0429 | 0.3151 | −1.54 |
| acadesine | 0.0481 | 0.2451 | 1.30 | 2,3-Diphosphoglyceric acid | 0.0154 | 0.2114 | −1.67 |
| N6-Acetyl-L-lysine | 0.0226 | 0.2451 | 1.29 | ||||
| (b) | |||||||
| Lipid | p-Value | BH p-Value | tFC | Lipid | p-value | BH p-value | tFC |
| PC (36:7e) | 0.038 | 0.269 | 1.52 | DG (18:3e) | 0.038 | 0.269 | −1.33 |
| PE (16:1e) (22:6) | 0.038 | 0.269 | 1.49 | PI (18:0) (20:4) | 0.038 | 0.269 | −1.38 |
| BiotinylPE (32:5) | 0.004 | 0.269 | 1.46 | TG (16:0) (16:0) (17:0) | 0.038 | 0.269 | −1.39 |
| PIP2 (31:6e) | 0.002 | 0.269 | 1.32 | TG (6:0) (6:0) (14:1) | 0.038 | 0.269 | −1.44 |
| DG (8:0) (12:2) | 0.050 | 0.377 | 1.27 | TG (12:1e) (6:0) (6:0) | 0.002 | 0.269 | −1.46 |
| DG (4:0) (10:3) | 0.011 | 0.269 | 1.20 | PE (18:0) (22:6) | 0.011 | 0.269 | −1.47 |
| DG (4:0) (11:3) | 0.038 | 0.269 | 1.12 | CerP (d36:3+O) | 0.016 | 0.338 | −1.53 |
| PC (17:0) (14:1) | 0.038 | 0.269 | 1.07 | PE (16:1e) (20:4) | 0.004 | 0.269 | −1.57 |
| PC (16:1e) (18:1) | 0.030 | 0.377 | −0.91 | PC (32:0e) | 0.038 | 0.269 | −1.62 |
| CL (15:0) (18:1) (15:0) (18:1) | 0.030 | 0.269 | −1.16 | PC (32:1e) | 0.038 | 0.269 | −1.64 |
| PE (40:5e) | 0.038 | 0.269 | −1.19 | PE (18:0) (20:3) | 0.011 | 0.269 | −1.74 |
| DG (6:0) (11:3) | 0.038 | 0.269 | −1.23 | SM (d38:2) | 0.038 | 0.269 | −1.80 |
| ChE (20:4) | 0.038 | 0.269 | −1.31 | PC (35:1) | 0.038 | 0.269 | −2.09 |
| (c) | |||||||
| Protein | p-Value | BH p-Value | tFC | Protein | p-Value | BH p-Value | tFC |
| ACAN | 0.0126 | 0.5077 | 1.79 | MICA | 0.0113 | 0.5077 | 0.90 |
| CFL1 | 0.0407 | 0.5077 | 1.62 | HAPLN1 | 0.0424 | 0.5077 | 0.86 |
| CXCL11 | 0.0465 | 0.5077 | 1.46 | PRKCA | 0.0351 | 0.5077 | −1.02 |
| H2AFZ | 0.0191 | 0.5077 | 1.44 | PTPN6 | 0.0440 | 0.5077 | −1.04 |
| MUC1 | 0.0370 | 0.5077 | 1.39 | IGFBP2 | 0.0285 | 0.5077 | −1.07 |
| NAMPT | 0.0287 | 0.5077 | 1.36 | FSTL1 | 0.0370 | 0.5077 | −1.12 |
| INS | 0.0185 | 0.5077 | 1.35 | CTSD | 0.0208 | 0.5077 | −1.15 |
| CD97 | 0.0058 | 0.5077 | 1.32 | PROC | 0.0148 | 0.5077 | −1.21 |
| ICOS | 0.0275 | 0.5077 | 1.31 | CCL28 | 0.0086 | 0.5077 | −1.21 |
| PARK7 | 0.0278 | 0.5077 | 1.31 | CHRDL1 | 0.0258 | 0.5077 | −1.22 |
| FAM107B | 0.0279 | 0.5077 | 1.29 | MSN | 0.0142 | 0.5077 | −1.24 |
| CD38 | 0.0478 | 0.5077 | 1.29 | MMP14 | 0.0278 | 0.5077 | −1.29 |
| NGF | 0.0079 | 0.5077 | 1.20 | CCL2 | 0.0300 | 0.5077 | −1.34 |
| PPIF | 0.0152 | 0.5077 | 1.17 | CXCL6 | 0.0167 | 0.5077 | −1.36 |
| THPO | 0.0100 | 0.5077 | 1.12 | GNLY | 0.0453 | 0.5077 | −1.43 |
| DCN | 0.0127 | 0.5077 | 1.09 | CTSV | 0.0370 | 0.5077 | −1.47 |
| Characteristics | Proteomics | Metabolomics | ||
|---|---|---|---|---|
| D (n = 24) | C (n = 24) | D (n = 18) | C (n = 18) | |
| Age, M (SD) (Range) | 73.0 (4.9) (65–81) | 72.6 (5.5) (64–83) | 72.9 (5.1) (65–81) | 73.0 (5.7) (65–83) |
| Female, n (%) | 11 (46) | 11 (46) | 10 (56) | 10 (56) |
| MMSE, M (SD) | 27.2 (2.0) | 27.5 (1.7) | 27.3 (2.1) | 27.7 (1.4) |
| Absolute difference in year of surgery between D and C, mean (SD) | 0.8 (0.8) | 1.2 (1.2) | ||
| Characteristics | Lipidomics | Multi-omics | ||
| D (n= 16) | C (n = 16) | D (n = 15) | C (n = 15) | |
| Age, M (SD) (Range) | 72.9 (4.6) (65–81) | 72.6 (5.3) (64–82) | 73.0 (4.8) (65–81) | 72.3 (5.4) (64–82) |
| Female, n (%) | 9 (56) | 9 (56) | 8 (53) | 8 (53) |
| MMSE, M (SD) | 27.2 (2.2) | 27.8 (1.5) | 27.4 (2.1) | 27.9 (1.3) |
| Absolute difference in year of surgery between D and C, mean (SD) | 1.0 (0.8) | 0.9 (0.8) | ||
| Molecular Type | All 1 | Used for Analysis | QC Approach | <10% CV 2 n (%) |
|---|---|---|---|---|
| Lipids | 161 | 26 | Pooled QC samples [15,34] | 159 (98.8) |
| Metabolites | 219 | 51 | Pooled QC samples [15,34] | 219 (100) |
| Proteins | 1305 | 32 | Multi-step QC process established by manufacturer [33] | N/A |
| Lipid Class | Total (n = 26) | Down (n = 18) | Up (n = 8) |
|---|---|---|---|
| Phosphatidylcholine | 6 | 4 | 2 |
| Diacylglycerols | 5 | 2 | 3 |
| Phosphatidylethanolamines | 5 | 4 | 1 |
| Triacylglycerol | 3 | 3 | 0 |
| Biotintl-PE | 1 | 0 | 1 |
| Ceramide-1-phosphates | 1 | 1 | 0 |
| Cholesteryl esters | 1 | 1 | 0 |
| Cardiolipins | 1 | 1 | 0 |
| Phosphatidylinositol | 1 | 1 | 0 |
| Phosphatidylinositol bisphosphate | 1 | 0 | 1 |
| Sphingomyelin | 1 | 1 | 0 |
| Pathway | Total | Exp | Hits | Raw p-Value | FDR |
|---|---|---|---|---|---|
| Pentose phosphate pathway | 22 | 0.64 | 7 | 1.28 × 10−6 | 0.00011 |
| Arginine biosynthesis | 14 | 0.41 | 5 | 2.65 × 10−5 | 0.00111 |
| Glycolysis/Gluconeogenesis | 26 | 0.75 | 5 | 0.00067 | 0.0188 |
| Alanine, aspartate, and glutamate metabolism | 28 | 0.81 | 5 | 0.00096 | 0.0202 |
| Butanoate metabolism | 15 | 0.43 | 3 | 0.00805 | 0.113 |
| Nicotinate and nicotinamide metabolism | 15 | 0.43 | 3 | 0.00805 | 0.113 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tripp, B.A.; Dillon, S.T.; Yuan, M.; Asara, J.M.; Vasunilashorn, S.M.; Fong, T.G.; Inouye, S.K.; Ngo, L.H.; Marcantonio, E.R.; Xie, Z.; et al. Integrated Multi-Omics Analysis of Cerebrospinal Fluid in Postoperative Delirium. Biomolecules 2024, 14, 924. https://doi.org/10.3390/biom14080924
Tripp BA, Dillon ST, Yuan M, Asara JM, Vasunilashorn SM, Fong TG, Inouye SK, Ngo LH, Marcantonio ER, Xie Z, et al. Integrated Multi-Omics Analysis of Cerebrospinal Fluid in Postoperative Delirium. Biomolecules. 2024; 14(8):924. https://doi.org/10.3390/biom14080924
Chicago/Turabian StyleTripp, Bridget A., Simon T. Dillon, Min Yuan, John M. Asara, Sarinnapha M. Vasunilashorn, Tamara G. Fong, Sharon K. Inouye, Long H. Ngo, Edward R. Marcantonio, Zhongcong Xie, and et al. 2024. "Integrated Multi-Omics Analysis of Cerebrospinal Fluid in Postoperative Delirium" Biomolecules 14, no. 8: 924. https://doi.org/10.3390/biom14080924
APA StyleTripp, B. A., Dillon, S. T., Yuan, M., Asara, J. M., Vasunilashorn, S. M., Fong, T. G., Inouye, S. K., Ngo, L. H., Marcantonio, E. R., Xie, Z., Libermann, T. A., & Otu, H. H. (2024). Integrated Multi-Omics Analysis of Cerebrospinal Fluid in Postoperative Delirium. Biomolecules, 14(8), 924. https://doi.org/10.3390/biom14080924