Abstract
Skin aging is influenced by intrinsic and extrinsic factors that progressively impair skin functionality over time. Investigating the skin aging process requires thorough research using innovative technologies. This review explores the use of in vitro human 3D culture models, serving as valuable alternatives to animal ones, in skin aging research. The aim is to highlight the benefits and necessity of improving the methodology in analyzing the molecular mechanisms underlying human skin aging. Traditional 2D models, including monolayers of keratinocytes, fibroblasts, or melanocytes, even if providing cost-effective and straightforward methods to study critical processes such as extracellular matrix degradation, pigmentation, and the effects of secretome on skin cells, fail to replicate the complex tissue architecture with its intricated interactions. Advanced 3D models (organoid cultures, “skin-on-chip” technologies, reconstructed human skin, and 3D bioprinting) considerably enhance the physiological relevance, enabling a more accurate representation of skin aging and its peculiar features. By reporting the advantages and limitations of 3D models, this review highlights the importance of using advanced in vitro systems to develop practical anti-aging preventive and reparative approaches and improve human translational research in this field. Further exploration of these technologies will provide new opportunities for previously unexplored knowledge on skin aging.
1. Introduction
To effectively study skin aging, it is essential to have suitable models. Over the years, in vivo, in vitro, and in silico approaches have been developed for pharmaceutical and cosmetic testing. Due to growing ethical and economic concerns, alternative in vitro models have been designed to replace or reduce animal experiments guided by the ethics of the ”3 R” principles: replacement, reduction, and refinement [1]. These “principles” were formulated to assist researchers in discovering and using the latest available techniques and to encourage the development of new tools and methods, defining the fundamental objectives of an innovative science [2]. Animal models are subject to ethical restrictions and show noticeable genetic differences from humans. These issues have further increased the need for alternative in vitro systems mimicking natural skin’s structural and functional hallmarks. Therefore, the development of alternative models has gained a high priority following the European ban on animal testing for cosmetic products (2009/1223/EU), the REACH guideline for chemicals (2006/1907/EC), and the recommendation to follow the 3 R principle for research (2010/63/EU) [3].
Overall, the technical advances of 3D models may aid in uncovering the underlying causes of skin aging, which may further fuel the discovery of suitable treatments. The aim of this review is to describe the 3D experimental models currently proposed for investigating the molecular mechanisms underlying human skin aging [4] (i.e., upregulation of stress regulatory proteins, reactive oxygen species (ROS) accumulation, defects in integrin expression, increased collagen breakdown, telomere shortening, secretory-associated senescence phenotype (SASP)) and exploring preventive and reparative approaches.
2. From Basic 2D to Complex 3D Models
Two-dimensional in vitro models are a cornerstone in biomedical research, since they allow recreating a simple and controllable environment to study cellular behavior, drug responses, and disease molecular mechanisms. They also show the important advantages of being cost-effective and easy to maintain. By providing an accessible and economical platform and representing a quick and repeatable model to evaluate skin responses, 2D models facilitate experimental replication and high-throughput screening, making them indispensable in the initial research and drug development phases [5,6,7].
However, the currently used in vitro 2D skin aging models have no negligible limitations. Firstly, they do not fully replicate the complex structure and functionality of human skin because they lack its three-dimensional structure, which includes distinct layers such as the epidermis, dermis, and subcutaneous tissue. Particularly, the aged skin undergoes various structural and biochemical alterations that 2D models cannot absolutely replicate. These features, such as the spatial 3D organization, can affect the accuracy of experimental outcomes. Moreover, 2D models fail to mimic the dynamic interactions between skin cells, immune cells, and the extracellular matrix [8].
One of the crucial functions of the skin is to act as a barrier against environmental factors, another item improperly replicated by 2D models, limiting studies on permeability and drug delivery. The 2D cells interact only with nearby cells within the horizontal plane and are uniformly and directly exposed to stimuli or drugs without the action of other cellular types, limiting their ability to reproduce chemical and biological factors, i.e., the pH and oxygen gradients of the in vivo microenvironment, and cellular interactions [9]. The limitations in mimicking the true barrier function and tissue environment of human skin can result in unreliable data regarding drug absorption, efficacy, and toxicity when using 2D models. Also, mechanical properties of the skin, such as firmness and elasticity, are not replicated in 2D models. Aging skin physiologically changes in these properties, and the inability to mimic them accurately in 2D models can affect the relevance of experimental outcomes. In the dermis, human skin aging is characterized by reduced collagen synthesis, decreased mitochondrial function, lower ATP levels, and increased nuclear factor-kappa B (NF-κB) signaling, which is associated with inflammation and damaged skin barrier function [10]. The absence of immune cells and blood vessels in 2D models strongly limits the study of inflammatory and vascular responses in the skin aging process [11].
Moreover, due to their simplified structure, 2D skin models may not be suitable for long-term studies, as they often fail to maintain cellular viability and function over extended periods, which is essential for chronic toxicity and prolonged treatment studies. Additionally, cells in 2D cultures can often exhibit different morphology, proliferation rates, and differentiation patterns compared to those in vivo. Hence, although the 2D skin models are generally less expensive and easier to be established in cell biology research and easy to be analyzed by several molecular, biochemical, and image-based assays than 3D models, they may not always provide the necessary accuracy for studies of translational relevance. This can lead to higher costs and efforts in later stages of research and development. All these limitations highlight the need for more advanced and realistic skin models, such as 3D organotypic cultures and “Skin-on-Chip” technologies [12,13], that could better replicate human aged skin for research purposes to transfer the results to the clinic. The timeline in Figure 1 depicts the most significant advances in developing skin 3D models.

Figure 1.
Timeline of the different advances in the development of skin 3D models. Pseudo-3D models: Bell et al., 1979 [14], 1981 [15,16]; Li et al., 2002 [17]. Organoids: Rheinwald et al., 1975 [18]; Itoh et al., 2011 [19], 2013 [20], Guenou et al., 2009 [21], Takahashi et al., 2006 [22]; Lee et al., 2020 [23]. Reconstructed human skin (RHS): Fentem and Botham 2002 [24]; Liu et al., 2007 [25]; Diekmann et al., 2016 [26]. Skin-on-chip: Lim et al., 2018 [27]; Jeong et al., 2021 [28]; Pauty et al., 2021 [29]. Three-dimensional bioprinting: Lee et al., 2009 [30]; Min et al., 2018 [31]. Skin microbiota model: Kim et al., 2021 [32]; Rikken et al., 2023 [33].
Figure 1.
Timeline of the different advances in the development of skin 3D models. Pseudo-3D models: Bell et al., 1979 [14], 1981 [15,16]; Li et al., 2002 [17]. Organoids: Rheinwald et al., 1975 [18]; Itoh et al., 2011 [19], 2013 [20], Guenou et al., 2009 [21], Takahashi et al., 2006 [22]; Lee et al., 2020 [23]. Reconstructed human skin (RHS): Fentem and Botham 2002 [24]; Liu et al., 2007 [25]; Diekmann et al., 2016 [26]. Skin-on-chip: Lim et al., 2018 [27]; Jeong et al., 2021 [28]; Pauty et al., 2021 [29]. Three-dimensional bioprinting: Lee et al., 2009 [30]; Min et al., 2018 [31]. Skin microbiota model: Kim et al., 2021 [32]; Rikken et al., 2023 [33].

3. 3D Models as Innovative Tool for Studying Skin Aging
Generally, 3D cell culture models represent a significant innovation with respect to traditional 2D models and serve as a suitable alternative to animal models in industrial applications and experimental research, providing a more accurate representation of the in vivo environment. Three-dimensional skin aged models can simulate the multi-layered structure of human skin and allow the physiological cell–cell and cell–matrix interaction in three dimensions, mimicking the natural architecture and organization of tissues. For the first time, in the middle of the 20th century, Medawar defined the method of growing adult mammalian skin epithelial cells from skin explants. He achieved this by incubating the skin epithelium in a specific medium supplemented with serum, maintained in gaseous equilibrium with air or an air-oxygen mixture, and the cells that formed it were able to proliferate and migrate [34].
Another important factor is that 3D skin models can have the simultaneous presence of both aged cells and non-aged cells as well as immune cells, vascular components, and, not least, appendages (e.g., sweat glands, sebaceous glands, hair follicles, and vessel-like channels)—all reviewed in Mori et al. [35]. These features, increasing the model complexity, enhance the soundness of experimental results and understanding of how cell communication may be involved in the aging process of tissues [36].
Developing 3D in vitro models for skin aging, recreating an aged phenotype, poses a significant issue since aging is a multifaceted process that involves numerous intrinsic and extrinsic factors and complex biological and molecular changes that are difficult to replicate in standard, shorter-term in vitro models. Overall, 3D skin aged models represent a significant advancement in studying skin aging, offering a more accurate and comprehensive representation of these complex processes [37]. Also, innovative skin aging systems provide information on approaches that could slow down the aging process by extending the healthy life span.
Since, as said before, aging is an irreversible biological process that takes decades in an organism, a crucial factor to consider for generating an aged skin model is the culture time of the cells involved. In this regard, Dos Santos et al. studied the chronological aging of the epidermis, developing an in vitro 3D model of skin aging showing the age-related phenotype (decrease in the epidermis thickness, a reduced expression of hyaluronan and its surface receptor CD44, etc.) [38]. Although this model is very similar to in vivo aged skin, the long culture time to establish it is a disadvantage for the high-throughput generation of skin equivalents [37]. To overcome this hurdle, bioengineered skin models have been created with the direct incorporation of senescent cells that stop dividing but remain metabolically active, added with other normal cells, providing a more accurate representation of the skin microenvironment and the complex interactions between all cells. It is known that senescent cells strongly influence the surrounding tissue by secreting pro-inflammatory factors, chemokines, cytokines, and proteases, known to induce the SASP [39]. The aged in vitro model (cultured skin equivalents) was generated through the co-culture of keratinocytes and senescent fibroblasts on a scaffold made of collagen, glycosaminoglycan, and chitosan. To simulate the senescence phenotype, fibroblasts were previously exposed to mitomycin-C that induced premature senescence by triggering DNA damage and producing reactive oxygen species (ROS). The mitomycin-C-treated fibroblast showed an aged-related phenotype characterized by a reduced proliferative ability, decreased expression of filaggrin, an essential protein for the correct formation and function of the skin protective barrier in the epidermis and lowered elastin and collagen in dermal tissue, and increased secretion of pro-inflammatory cytokines [26].
Moreover, to increase the efficiency of 3D aged skin models, more recent work has been carried out incorporating increasing ratios of hydrogen peroxide (H2O2)-induced premature senescent fibroblasts together with normal fibroblasts into the collagen matrix. This skin equivalent, named “senoskin”, recapitulated several key hallmarks of skin aging, such as reduced collagen production, increased levels of pro-inflammatory cytokines, and an altered extracellular matrix (ECM) composition [40].
Victorelli et al. suggested further improvement of 3D skin models since they incorporated ultraviolet (UV)-induced senescent melanocytes in skin equivalents. This model even more mimicked the structure and function of native human skin and allowed a better understanding of how these cells affect the microenvironment in a setting that closely resembles in vivo conditions. Senescent melanocytes expressed various inflammatory and SASP markers, which are known to induce skin aging by influencing nearby non-senescent cells and the ECM. This process induced telomere dysfunction and decreased proliferation of surrounding cells (keratinocytes and fibroblasts) through CXC chemokine ligand 3 (CXCL3)-dependent mitochondrial ROS, thereby contributing to skin aging [41].
An alternative way to develop 3D aged skin equivalent is by the incorporation of advanced glycation end-products (AGEs) into dermal–epidermal compartments. The study of Markiewicz et al. utilized a commercially available full-thickness skin equivalent (a dermal–epidermal skin model with a fully developed basement membrane (EpiDerm-FT™)) treated with methylglyoxal (MGO), a highly reactive dicarbonyl compound and a key intermediate in AGE formation, in culture media. The MGO-induced AGEs in the 3D skin model resulted in significant changes, including a rapid aged phenotype induction characterized by the increase in AGE formation, which interfered with the skin’s structural integrity, decreased collagen expression, and reduced stratification of the epidermis. Of note, in this 3D skin model, external treatment with derivatives of resveratrol reduced the AGEs’ generation, decreased the expression of aging and oxidative stress markers, and restored collagen levels and epidermal integrity [42]. Lee and colleagues established an in vitro 3D skin model from EFT-400 full-thickness reconstituted skin tissue (EpiDermFT) in which human keratinocytes and fibroblasts were sequentially cultured on a matrix used to support cell growth in three dimensions, allowing them to create the epidermal and the dermal layer, respectively. To induce the aging process through glycation, the new 3D skin was exposed to glyoxal (GO). In order to assess the degree of glycation within the skin equivalent, the levels of Nε-(carboxymethyl)lysine, a well-known marker of glycation, were evaluated. This 3D skin model, imitating the effects of glycation on skin aging, could be a good tool to screen the antiglycation activity of topical formulations, counteracting the effects of glycation and slowing the aging process [43].
Exogenous stressors, such as UV radiation, are significant environmental factors contributing to skin aging, a process known as photoaging. Only the ultraviolet type A (UVA) (320–400 nm) and ultraviolet type B (UVB) (280–320 nm) can pass through the atmosphere, reaching the skin. Direct exposure of skin to sunlight or UV radiation led to harmful effects, including sunburn, inflammation, photo-immunosuppression, and even skin cancers [44]. Additionally, UV radiation is a primary cause of premature skin aging since the UVA exposure accumulates ROS that activate the NF-κB and mitogen-activated protein kinase (MAPK) signaling pathways [45] and increase the downstream molecule tumor necrosis factor alpha (TNF-α) and matrix metalloproteinases (MMPs) expression, leading to ECM degradation (alteration in collagen and elastic fibers in the dermis). All these events can be measured in deep wrinkle formation, skin laxity, and hyperpigmentation, all features of photoaged skin [46]. In view of this, Casale et al. established a human full-thickness skin model (named Endo-HSE) exposed to UVA radiation as an advanced in vitro model to examine the downstream effects of UVA damage and test the efficacy of photoprotectants, such as sunscreens, antioxidants, and retinoic acid. In detail, fibroblasts were obtained from human tissue biopsies, cultured on gelatin microcarriers, then transferred to a maturation chamber for six weeks to form disk-shaped tissue constructs. Subsequently, to create epithelium, keratinocytes and melanocytes were plated onto a newly formed dermal layer for one week in submerged conditions and then lifted to the air–liquid interface for up to 14 days. The new 3D skin model was treated with UVA radiation by using a UVA lamp in order to simulate the solar exposure and has measured significant oxidative stress in both the dermis and epidermis and increased senescence markers expression (such as p63, Ki67, and activated caspase 3) in the epidermal layer, a reduced self-renewal capacity of skin stem cells were measured, and also the modified remodeling of collagen architecture in the dermis was notably observed, all changes associated with aging [47].
4. Overview of 3D Aged Skin Models
The 3D innovative skin models, described in the following paragraphs highlighting their peculiar features, are listed in Figure 2a. Figure 2b summarizes the microstructure of obtained skin by applying these models.
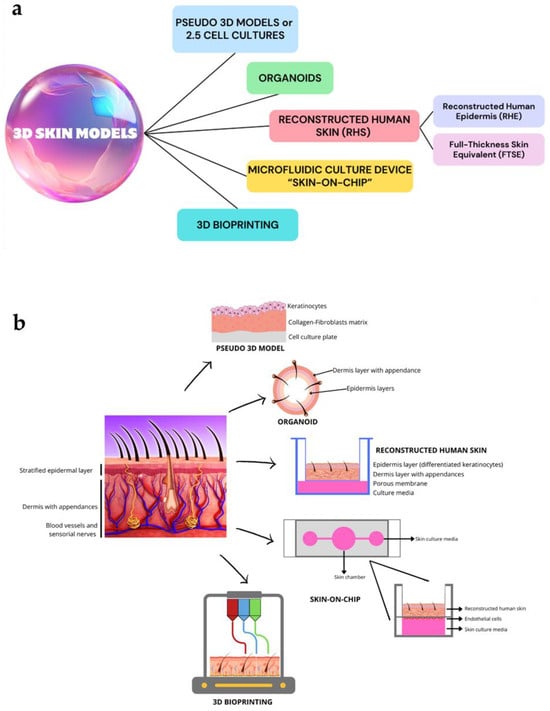
Figure 2.
A schematic overview of the main 3D skin models (a) and different microstructure of skin obtained by each modeling technology (b). These images were created with www.canva.com, accessed on 12 August 2024.
4.1. Pseudo-3D Systems
It is known that skin aging leads to decreased elasticity, loss of dermal matrix integrity and cell contraction, and an impaired migration ability. A valid cellular model of aged skin should also consider these features, which are significantly compromised in senescent cells compared to younger ones. In this scenario, it is necessary to specify the presence of models named “pseudo-3D systems” or alternatively called “2.5D cell cultures”. These are hybrid models borderline between 2D and 3D in which cells are grown in a bidimensional condition and are plated on top of a thick layer on ECM-coated culture and, therefore, do not fully reflect the complexity of 3D systems, but at the same time, try to recreate multiple physiological conditions. However, these cultures do not entirely replicate an in vivo environment, as the cells are still embedded into the culture medium and lack direct and complete contact with the ECM on their surface. Cell migration in the 2D space is primarily driven by adhesion and traction forces. However, in a three-dimensional microenvironment, additional factors such as biochemical, steric, and mechanical influences play a significant role in in vivo stiffness and then in cell migration [48]. The reduced functional ability in terms of the cell migration of senescent fibroblasts has been recently verified using a 2.5D model. Human dermal fibroblasts (FF-95) derived from foreskin tissue were induced to express an aged phenotype following replicative senescence, and then they were mixed with a solution of type I bovine collagen to evaluate their migration. Senescent fibroblasts, when placed in 2.5D culture, exhibited a significant reduction in contractility within collagen gels compared to younger cells, highlighting their weakened mechanical properties. The contractility reduction is due to alterations in cytoskeletal organization, which are crucial for maintaining structure and function in young skin [49]. To sum up, although pseudo-3D models offer valuable insights into the behavior of aged cells, they have the critical limitation of not perfectly replicating the complexity of in vivo environments. These models have underscored the impaired mechanical properties and migration abilities of senescent fibroblasts, emphasizing cytoskeletal changes as a crucial factor in aging.
4.2. Organoid Cultures
Three-dimensional structures that self-organize due to the natural tendency of adherent cells to aggregate themselves, known as organoids, appear to be a significant breakthrough for scientific research. Sato et al., in 2009, created for the first time organoids from intestinal stem cells [50]. Subsequently, they have been developed from various tissues, including the lungs, brain, stomach, liver, pancreas, prostate, mammary gland, fallopian tubes, taste buds, lungs, salivary glands, esophagus, epididymis, tongue, lacrimal gland, and thyroid (all reviewed in Suarez-Martinez et al. [9]).
Organoids derive from stem cells such as induced pluripotent stem cells (iPSCs), adult stem cells (ASCs), or embryonic stem cells (ESCs), and they develop into organotypic structures when cultured in a 3D matrix with specific growth factors. This process involves stepwise differentiation protocols that mimic gastrulation and organogenesis signals, and by using various growth factors or inhibitor molecules, the formation of specific germ layers, such as endoderm, mesoderm, and ectoderm, could be induced [51]. iPSCs used to create organoids are generated by reprogramming adult cells to a pluripotent state, allowing them to differentiate into any cell type [52]. Regarding ASCs, the cells are isolated directly from skin tissue and expanded in culture and can be induced to form skin organoids by providing an appropriate 3D scaffold and a cocktail of growth factors promoting skin differentiation [53].
Notably, organoids can form lumens, gland-like structures, and other organ-specific features [23]. Hence, organoids are miniaturized organs with a 3D structure and multiple cell layers composed of tissue-specific cell types and stem cells, exhibiting a unique intrinsic organization [54]. They can be expanded over long periods, genetically modified, and cryopreserved while maintaining their phenotypic characteristics [55,56].
Organoids substantially differ from spheroids, another three-dimensional cell culture with a spherical morphology, generally composed of a single somatic cell type (including cancer cells, primary cells, or cell lines). When the attachment to a substrate is precluded, the cells are able to self-aggregate and generate the spheroid that better mimics cell–cell and cell–matrix interactions than 2D cultures, although they lack the capacity to replicate the more complex structure or function of an organ [57]. Spheroids are commonly used in cancer research to assess tumor growth, invasion, and drug resistance or are used in drug screening and in studies of cell–cell and cell–matrix interactions [58]. Of note, organoids are used in developmental biology to study organogenesis and tissue differentiation, in disease modeling to understand the mechanisms involved in different diseases, such as genetic disorders, infectious diseases, and cancer within a specific organ context, and are employed in regenerative and personalized medicine to develop and test new therapies. Finally, organoids serve as platforms for studying complex organ functions and interactions with their microenvironment [59]. Organoids derived from patient samples, known as patient-derived organoids (PDOs), have shown promise in cancer research for personalized medicine, as they can more accurately replicate the genetic landscape and drug response of the original tumors compared to traditional 2D cultures [60]. Furthermore, PDOs represent a potential preclinical model tool to identify novel biomarkers of aging and design personalized approaches for the prevention and treatment of age-related diseases [61].
Regarding the skin models, the concept of utilizing a more complex in vitro skin culture system was first introduced in 1975 by Rheinwald et al., who tested a self-organizing approach to produce stratified squamous epithelium through the serial co-cultivation of primary human keratinocytes and irradiated mouse fibroblasts. This innovative discovery paved the way for the in vitro culture of self-organizing skin tissue [18]. In the early 2000s, 3D self-organized layered epidermal equivalents obtained from ESCs and iPSCs were developed, representing a significant breakthrough in the field of skin organoids [19,20,21,22]. Now, 3D skin equivalents can faithfully replicate the right structure and composition of native skin, including the epidermis, dermis, and also skin appendages [62,63]. In 2011, Itoh et al. produced 3D skin equivalents by seeding keratinocytes obtained by fibroblast-derived iPSC and stimulated with a specific growth factor onto a support matrix containing human feeder fibroblasts [19]. The epithelial differentiation of iPSCs into keratinocytes was achieved using retinoic acid and bone morphogenetic protein 4 (BMP4) to prevent neural differentiation [64]. The organoids obtained with this experimental protocol closely resembled the complex layering of native epidermis due to the presence of stratification of iPSC-derived keratinocytes [65]. In 2020, Lee et al. generated an almost complete self-organized skin system in vitro differentiated from iPSCs, forming a skin organoid that recapitulated many appendage structures, including hair follicles [23]. Interesting studies provided valuable results into the development and functionality of skin-related organoids, specifically focusing on mature skin systems of appendages (sebaceous and sweat glands, respectively) [66,67]. The first study highlights that the expression of B lymphocyte-induced nuclear maturation protein 1 (Blimp1) on hair follicle stem cells and cells that reside in the sebaceous gland base is responsible for generating sebaceous gland organoids. These organoids exhibit key characteristics of sebaceous glands, such as lipid secretion, making them a helpful tool for researching gland biology and skin conditions like acne [66]. The second one developed functional sweat gland organoids, which are crucial for thermoregulation and skin hydration. They generated organoids from mouse sweat gland-derived stem cells cultured in a 3D matrix and specific growth factors [67].
To obtain organoids for aging research, Pitrez et al. recently described how reprogramming somatic cells into iPSCs then differentiated in specific cell types was efficient. Moreover, an alternative strategy to investigate aging mechanisms is the transdifferentiation from one somatic cell type to another obtained from aged or age-related disorder-affected donors. However, the challenges in accurately mimicking the aging process and translating findings to in vivo are still ongoing [36]. iPSC-derived skin organoids initially resemble early fetal skin structures and then develop more mature features after extended culture. However, they are not effective in simulating the complex and dynamic changes that occur during aging in vitro [68]. Therefore, based on the previous work [38], Sun et al. hypothesized that prolonging the time of the organoid culture in vitro as much as possible or obtaining skin organoids generated from fibroblasts exposed to mitomycin C could represent two more feasible methods to induce aging [69]. More recently, Kim and colleagues described in detail the procedure for obtaining skin organoids. They developed a peculiar model of skin organoids with hair follicles to evaluate the skin’s response to environmental factors like solar ultraviolet exposure [70]. Skin organoids are generated from human iPSCs, which can differentiate into various skin cell types. The process begins with the induction of iPSCs to form embryoid bodies, which are then induced to differentiate into skin cells, including keratinocytes, melanocytes, and dermal fibroblasts, using specific supplements added in the culture medium. After 85 days, the newly formed skin organoids were divided into eight equal parts, which were placed on polymerized collagen I-coated transwell culture inserts in a humidified incubator for 3–4 weeks. To promote further epidermal maturation, the skin organoids were cultured in an air–liquid interface culture condition at 37 °C for six days, replacing the medium daily. To induce photodamages and other signs of aging, skin organoids were irradiated for 20 min, twice, with a 2 h interval between exposures with a combination of UV-A and B, that closely mimic natural solar radiation. This procedure was repeated every two days, resulting in three UV exposures. The findings indicated skin barrier disruption, a notable increase in epidermal thickness and of the number of sunburn cells, identified by fragmentation or condensation of cell nuclei. Moreover, an increased skin pigmentation due to the higher number of melanocytes compared to control skin organoids, downregulation of genes related to skin barrier function (filaggrin and loricrin), and extracellular matrix degradation (significant decrease in type I collagen expression and an increase in MMP-1 expression) were observed. Of interest, a detrimental effect of UV radiation on hair follicles was described [70].
An important discovery highlights the potential of skin organoids in therapeutic applications, in particular for skin regeneration and wound healing, which are closely related to skin aging and are affected by the changes that occur in the skin over time [71]. Kim et al. generated skin organoids using iPSCs obtained from HLA-homozygous cord blood mononuclear cells and peripheral blood mononuclear cells with the aim of overcoming immune rejection, a significant hurdle in clinical applications of stem cell therapies. To test the functional abilities of the skin organoids, they were transplanted into immunodeficient mice with skin lesions. Following a good integration with the host tissue, they promoted healing, restoring normal skin functions [72].
Although organoids are defined as useful 3D skin models in vitro, there remain many issues to overcome. The main limitations include the absence of physiological communication between tissues, the incomplete development of vascularity, and impaired formation of complex neural networks and groups of immune cells that are often not fully integrated [73,74]. In addition, two specific limitations for organoids of aged skin are related to the low reprogramming efficiency in iPSCs of cells obtained from aged donors as compared to those of young subjects, and to the potential loss of some epigenetic markers of aging during the reprogramming process [75,76]. Furthermore, it is important to highlight that there is not currently a universally shared and standardized protocol for skin organoid generation, since organoids often exhibit high heterogeneity [77]. The possibility of having biobanks of organoids related to various pathologies could be a way to overcome this obstacle. However, the application of organoids in aging research holds great promise, enhancing our understanding of aging, identifying potential molecular markers for clinical evaluation of aging interventions, and suggesting strategies to combat aging and age-related diseases. Continued advancements in organoid technology and methods will provide a valuable tool for modeling the changes that occur with advancing age in human skin and for studying the development of age-associated skin conditions.
4.3. Reconstructed Human Skin (RHS)
The term “Reconstructed Human Skin” (RHS) refers to laboratory-grown skin models (dermal equivalent) mimicking the structure and function of human skin. These 3D models may consist of either just a skin layer or multi-layers (epidermis and dermis), depending on the cell type used and the proposed application. The RHS model is widely used in experimental research to study basic skin biology and disease mechanisms and to test pharmacological and cosmetic approaches. In light of this, we can identify the reconstructed human epidermis (RHE) model as an advanced in vitro 3D system that tries to replicate the fully differentiated epidermis without a dermal compartment. This model is composed by culturing only keratinocytes (from primary cultures or cell lines) on a supportive and inert matrix (i.e., collagen solution), allowing them to differentiate and create a multi-layered structure that is very similar to natural epidermis. Keratinocytes are plated into transwell chambers and shortly cultivated under submerged conditions; subsequently, the cultures are exposed to the air–liquid interface, which induces keratinocyte differentiation [78,79]. After two weeks of culture, a structure like the epidermis is characterized by classical cellular stratification (epithelium with basal, spiny, granular, and corneal layers), and the physiological properties of the human epidermis, that resembles a physiological epidermis. The RHE models are extensively used for dermatological research, including studies on barrier function, irritation, and drug permeability, and are used to test anti-aging treatments and products. They provide a more ethical and cost-effective alternative to animal testing and are valuable for understanding skin physiology and pathology. Several RHE are commercially available and supplied by various companies (listed in the review of Hofmann et al. [80]). Moreover, “pigmented RHE models”, which also incorporate melanocytes of different phototypes in the basal layer of the epidermal construct, can be either produced or purchased [25]. These systems had been utilized to study in a highly controlled environment the phototoxicity, pigmentation disorders, and test the effectiveness of commercially available cosmetic products such as sunscreens [81]. Hall and colleagues recently reported that pigmented RHE models can achieve realistic epidermal pigmentation through the process of melanocore transfer between melanocytes and keratinocytes in the epidermis [82]. The RHE model was also employed to investigate how the aging process can influence the amount, function, and microRNA (miRNA) content of the extracellular vesicles (EVs) released from the keratinocytes of young or aged skin samples. The results demonstrated that aged keratinocytes release more EVs compared to non-senescent keratinocytes; EVs from aged cells showed a specific microRNA profile characterized by an elevated level of miR-30a, central in the regulation of human epidermal barrier functions, and finally, inhibited the proliferation of young keratinocytes, and impaired the initial stages of wound healing in mice [83].
The RHS model has been improved by creating the full-thickness skin equivalent (FTSE). FTSE refers specifically to a type of RHS that replicates both skin layers (epidermal and dermal) simultaneously, thus providing a more complex model of human skin. FTSE is therefore formed by keratinocytes, fibroblasts, and extracellular components of the matrix, and it is for this reason that the FTSE model is used in wound healing studies, skin regeneration, and advanced dermatological research. Specifically, a keratinocytes’ layer is seeded on top of a fibroblast-loaded matrix and initially cultured under submerged conditions. After a short period, the culture is exposed to air, like the RHE protocol. Interestingly, skin aging can be induced in FTSE models by treatment with the cytostatic drug mitomycin C. Diekmann et al. developed and characterized a 3D skin model that accurately reflected the structural and functional aspects of aged skin. They created reconstructed skin co-culturing fibroblasts and keratinocytes on a collagen–glycosaminoglycan–chitosan scaffold mimicking the ECM, and aging was induced by the exposition of fibroblasts to mitomycin-C. This model exhibited key markers of aged skin, including increased expression of AGEs, decreased collagen I, and an increase in MMP-1 activity [26].
In summary, the literature documents the significant advancements and applications of RHS models in experimental research. The development of these sophisticated models allows for studying skin aging processes, thus providing a precious tool for understanding skin physiology and pathology and testing pharmacological and cosmetic treatments.
4.4. The Microfluidic Culture Device Called “Skin-on-Chip”
Nowadays, “Skin-on-Chip” (SoC) technology represents a significant advancement in the 3D skin models’ field. It overcomes the limitations of animal models by offering reproducible and reliable platforms for ex vivo modeling, ensuring the physiological relevance of in vivo skin [84]. In more detail, SoC is an advanced microfluidic culture device that integrates living cells into a micro-engineered environment to accurately mimic the dynamic environment, simulating the movement of fluids present in the human skin. Zhang et al. described this model as follows “Skin-on-Chip is to culture skin tissues within a microfluidic system, which can control many physical and biochemical parameters such as medium flow, mechanical force and gradients of biochemicals, mimicking the 3D microenvironments of the natural human skin” [85]. Key features of SoC devices are a microfluidic system that allows the continuous perfusion of fluids and a dynamic environment that mimics various physiological conditions such as mechanical stress, nutrient gradients, and fluid flow. Having the ability to adjust the physical parameters to control the cellular microenvironment, SoC technology has promoted the transition from static to dynamic 3D models that more accurately recapitulate human physiology, including the integration of vasculature and the immune system [84]. Constant and in-depth experimental research has enabled the creation of perfusable vascular channels in 3D skin-equivalent models (composed of normal human dermal fibroblasts and normal human epidermal keratinocytes). These models are developed by seeding endothelial cells into the dermal layer to induce vascularization, forming capillary networks [35].
Actually, few articles on the use of 3D aged skin models placed in microfluidic systems are available. Lim et al. induced aging in fibroblast/keratinocyte skin equivalents constructed on a microfluidic cell culture device using magnetic stretching with the aim of inducing aging in the skin equivalent and forming “Wrinkled Skin on the Chip” (WSOC) [27]. Briefly, a co-culture of human fibroblasts and keratinocytes was generated: a collagen solution containing fibroblast cells was placed in the SoC chamber in a CO2 incubator for 1 h to form a gel. Subsequently, to develop the stratum corneum of the epidermis, human keratinocytes were sprinkled onto the collagen layer and then incubated for 1 h for cell adhesion. Finally, the 3D model was subjected to uniaxial stretching using attractive and repulsive forces between an electromagnet and a permanent magnet. The mechanical stimulus reduced the proliferation of fibroblasts and keratinocytes and decreased the production of collagen, fibronectin, and keratin. Due to the lower production of these proteins, the skin equivalents could not maintain the stratum corneum or withstand the tensile stress of magnetic stretching, resulting in wrinkle formation [27]. WSOC can be effectively used to test the effect of anti-wrinkle formulations and cosmetics before or after the in vivo approaches.
In the work of Jeong et al., for the first time, an in vitro aged FTSE has been developed into an advanced SoC technology that used mechanical stimulus reflective of the circadian rhythm. Primary human fibroblasts were suspended for five days in collagen I solution to create the dermal layer; next, primary keratinocyte cultures were left to attach to the newly formed dermal layer before the air exposure phase, in which cells are induced to differentiate. To simulate the aging process, a periodic mechanical stimulation that mimicked the circadian rhythm for over 28 days was applied. This methodology induced aging in in vitro skin models, as observed by a reduced contraction, decreased epidermal layer thickness, and increased β-galactosidase expression. By combining an FTSE with dynamic mechanical stimuli reflecting the circadian rhythm, this study provided a more accurate and physiologically relevant platform for investigating skin aging [28]. Another study on a 3D model revealed the morphological changes in blood vessels induced by aging fibroblasts. The 3D model integrated human blood vessels, formed within the collagen gel matrix by human umbilical vein endothelial cells (HUVECs), into a pre-formed channel combined with young or senescent fibroblasts. Senescent fibroblasts exerted excessive traction stress on the surrounding ECM, leading to significant mechanical rearrangement of ECM fibers and promoting angiogenesis through the SASP phenotype. To confirm this result, rapamycin, an SASP inhibitor, counteracted the secretory phenotype of senescent fibroblasts, modulating their impact on blood vessels [29].
With promising applications in simulating the skin aging process and evaluating anti-aging treatments, these cutting-edge models offer new opportunities for more relevant studies of skin physiology and its pathological changes, owing to their ability to dynamically simulate human skin’s physiological and mechanical conditions. The main advantages and disadvantages/limitations of the SoC systems are listed in Table 1.

Table 1.
Advantages and disadvantages and/or limitations of Skin-on-Chip technology.
4.5. 3D Bioprinting Models for Skin Aging as a Viable Alternative to Traditional Animal Testing
A bioprinted model refers to a 3D structure created using bioprinting, an advanced technology that can form biological structures with a complex architecture closely resembling a physiological microenvironment. Bioinks, which are mandatory to build 3D models, are composed of living cells (such as fibroblasts, keratinocytes, adipocytes, endothelial cells, and melanocytes [90]), preferably derived from primary cultures and autologous stem cells [91] and growth factors [92] combined with biomaterials (aqueous formulations as a hydrogel). These biomaterials form a solid post-printing structure that mimics the extracellular matrix (ECM) and supports the cells within, ensuring their proliferation and differentiation. In recent years, significant progress has been made in developing and applying bioprinting technology and using organoids as a component of bioink. They recreate miniaturized tumor clusters with hierarchically organized cells, thus allowing a better simulation of the intrinsic characteristics of the tumor microenvironment [93]. Polymers used in bioinks can be natural or synthetic (such as collagen, dextran, fibrin, agarose–chitosan, gelatin, hyaluronic acid (HA), hydroxyapatite, alginate, methylcellulose, polylactic acid (PLA), polyglycolide (PGA), poly(lactic-co-glycolic acid) (PLGA), and polyethylene glycol (PEG) [94]). All these biological components are homogenously deposited on a substrate following the natural layer-by-layer distribution by different approaches, such as inkjet, extrusion, and laser-assisted technology [95].
Bioprinting technology has made significant strides in recent years, demonstrating the ability to create tissue constructs that closely replicate a range of tissues and organs, such as liver, kidney, bone, heart, skin, neurons, and vascular systems [95]. This advanced technology is also currently applied in cancer research [96]. For example, bioink loaded with lymphoid cells isolated from patients with chronic lymphocytic leukaemia has been recently established [97]. Moreover, in a very recent work, a new 3D bioprinting technique has been developed to recreate the natural structure of human corneal stroma using adipose tissue stem cells [98].
The 3D bioprinted skin, a human cell-based full-thickness skin model, has recently emerged as an advanced technology in tissue engineering, regenerative medicine, and the study of skin aging since it gives the potential to recreate complex tissue structures that closely resemble physiological skin with all dermal layers, blood vessels, nerves, muscles, and skin appendages (sebaceous, sweat, and mammary glands, hair follicles, nerves, and fingernails) [90,99,100,101]. It enables the precise placement of different cell types and biomaterials in a spatially organized manner, closely replicating the architecture of natural tissues. This innovative approach allows the creation of skin equivalents that always more accurately mimic the structural and functional characteristics of natural skin and that can be used as clinical-grade surgical grafts, for the development of topical formulations, dermal toxicology research, and for testing anti-aging therapies [102]. Lee et al. were the first to create a 3D collagen hydrogel structure that mimics stratified skin by bioprinting two distinct layers of cells: inner fibroblasts and outer keratinocytes [30].
Ansaf and colleagues have reported on the technical procedure to obtain a 3D bioprinting model [102]. A notable advantage of this approach was that the layer-by-layer deposition ensures obtaining a printed skin closely like the physiological organization of human skin. In fact, the first layer deposited is the epidermal one, composed of keratinocytes, and the dermal layer containing fibroblasts is the deepest layer. After printing, the newly formed skin construct is left to grow in a cellular-appropriate environment so as to promote cell–cell interaction to form functional tissue.
A very recent work generated an advanced immunocompetent 3D bioprinted human skin model to assess skin sensitization. The authors incorporated macrophages into photopolymerizable bioink created by blending silk fibroin methacrylate, gelatin methacrylate, and photoactivated human platelet releasate. The bioink was deposited within a transwell system to create the 3D skin construct, where an artificial basement membrane supported both an epidermal layer and a vascularized dermal layer. Cultured in an air–liquid interface, the printed construct showed a differentiated keratinocyte layer and dermal ECM remodelling by fibroblasts and endothelial cells, with the incorporation of macrophages further enhancing the model’s physiological relevance [103].
Even if 3D bioprinted skin is a very recent technology, the scientific literature reports on this approach employed for skin aging studies are growing [92,101,104,105]. Further studies are needed to develop 3D printed models specifically for studying skin aging. However, a recent review explored the role of 3D bioprinting to understand the mechanism of therapy-induced senescence (TIS) (or tumor dormancy) in cancer [106]. TIS is a particular condition in which both tumor and normal cells, in response to radiation and chemotherapy, become senescent. The accumulation of senescent cells in tumors can, paradoxically, promote tumor relapse, metastasis, and resistance to therapy due to the expression of the SASP phenotype [107]. The 3D bioprinting recreating a 3D structure that mimics the in vivo tumor microenvironment allows studying more deeply the cellular interactions between different cell types, including senescent and non-senescent tumor cells, within a realistic ECM.
A printed skin model made by a bioink, composed of keratinocytes, fibroblasts, and melanocytes, suspended in a biocompatible hydrogel matrix has been generated to obtain a 3D skin model showing a visible pigmentation. Melanocytes were sequentially printed onto the dermal layer along with keratinocytes to induce skin pigmentation and then the new skin equivalent was exposed to the air–liquid interface [31].
In summary, the advancements in 3D bioprinted skin models represent the most significant leap forward in tissue engineering and regenerative medicine, offering unprecedented capabilities to replicate natural skin’s complex structures and functions. Although the scientific literature is still emerging, these models show great promise in accurately mimicking physiological skin and studying its aging processes. The use of various bioinks, together with innovative techniques like integrating macrophages and assessing stimulus-induced senescence, highlights the potential of these models for both clinical applications and in-depth research. The continued development and refinement of these technologies will be crucial to deepen our understanding of skin aging and assessing therapeutic interventions.
A summary of the highlights of 3D cell culture models used in human skin aging research is represented in Figure 3.
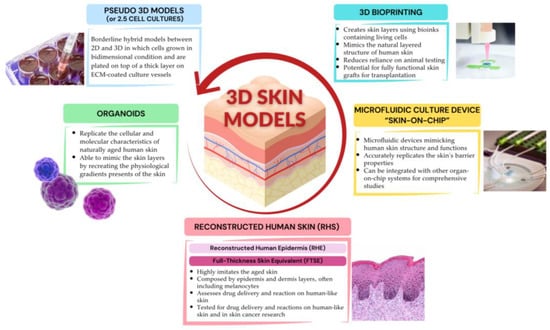
Figure 3.
Highlights of 3D cellular models used in human skin aging research. Created with www.canva.com, accessed on 24 July 2024.
4.6. Future Directions for 3D Aged Skin Model Research: The Crucial Role of Microbiota in Enhancing the Realism and Functionality of 3D Skin Models
A very interesting advantage that should not be underestimated in the use of 3D skin models is the inclusion of the skin microbiota. The skin is colonized by several bacteria that can change significantly throughout life, forming a complex ecosystem with skin cells and closely interacting with the host’s immune system to regulate immune responses [108,109]. It is well known that the microbiota significantly impacts the biology of in vivo skin, contributing to the maintenance of the outermost layer of the epidermis, the stratum corneum, which results from specialized keratinocyte differentiation [110] and cornification processes. In this layer, the complex microbial ecosystem can promote keratinocyte differentiation and preserve the integrity of the healthy barrier [111]. The skin microbiota interacts with keratinocytes through different pattern-recognition receptor-dependent mechanisms (toll-like receptors (TLRs)), inducing the secretion of cytokines and other immune mediators [112]. Since the crucial role of the microbiota in the skin is absent in 3D models, it does not fully mimic skin physiology and skin–microbiota interactions. Establishing a realistic microbiota allows the understanding of the dynamic interactions between skin cells and microbes, even in long-term studies. Excluding the 2D in vitro models, which are inadequate for developing systems that can accurately replicate the relationship between all skin cells and microbiota since they lack stratum corneum and excluding the animal models characterized by their own microbiota not quite similar to those of human skin [113,114], recent advancements aim to establish and maintain microbiota onto more complex human skin models. Rikken and colleagues recently established a cost-effective methodology that is easily usable with 3D skin models with different cell types and bacteria, which is an alternative to experimental animal models in preclinical research [33]. Landemaine et al. treated a 3D model with Staphylococcus epidermidis as a single strain and with skin microbiota from a healthy individual and observed that colonization with a complete microbiota is less invasive, better tolerated, and promotes keratinocyte proliferation and cohesion more effectively than single strains, highlighting the importance of integrating the microbiota into 3D skin models to increase the soundness of the results [115]. Similar conclusions were later presented by Loomis et al., who used a commercial 3D skin tissue equivalent (EpiDerm) co-cultured at the air–tissue interface with bacteria isolated from swabs of healthy human skin. They found that the presence of a microbial community had a more significant effect (changes in epidermal thickness, epidermal cell proliferation, and filaggrin production) on human skin (epidermal layer) than individual taxa [116]. This result underlines the importance of a complete microbiota in studies involving the use of 3D models.
The benefits of in vitro 3D models with integrated microbiota could be significant in the field of dermatology, basic biology, and are valuable for testing the effects of drugs and cosmetic products. Moreover, 3D skin models using aged patient-derived cells and microbiota could represent a cutting-edge technique for developing and studying targeted, personalized interventions.
To the best of our knowledge, there is only one work investigating the influence of the microbiota in a 3D model of aged skin. Briefly, a commercially available human 3D skin full-thickness model composed of keratinocytes and fibroblasts was exposed to Polyinosinic-polycytidylic acid (Poly(I:C)), an agonist of toll-like receptor 3 (TLR3) and retinoic acid-inducible gene I (RIG-I)-like receptors), to induce aging of the skin model. Then, the aged 3D skin model was treated with supernatant derived from the Streptococcus cultures (Streptococcus pneumoniae, Streptococcus infantis, and Streptococcus thermophilus) collected from young women’s faces by sterilized skin tape. In the aged 3D model, the exposure to Streptococcus supernatants restored the skin barrier in terms of elasticity increase, hydration, a decrease in desquamation, collagen upregulation, and improved lipid synthesis. All these features could be attributed to spermidine, a polyamine secreted by Streptococcus cultures [32].
The skin microbial composition predicts chronological age in adults, as it is significant in aged subjects. A very recent article investigated the complex relationship between skin microbiota and the signs of skin aging. Using a comprehensive multi-study analysis, the authors identified specific microbial characteristics that correlate with various markers of aging on the skin [117]. However, they do not use 3D models. Future research incorporating patient-derived aged fibroblasts, immune cells, and host microbiota could create more sophisticated 3D skin models that reflect the individual characteristics of older patients, mimicking the physiological interactions between the epidermis, dermis, and immune cells.
In Table 2, the experimental works using different types of 3D skin models exclusively for the study of aging research are listed.
Table 2.
Summary of experimental studies using 3D skin models in aging research.
5. Conclusions
Three-dimensional in vitro skin models represent a significant advancement in the study of skin aging, providing a more accurate and comprehensive understanding of this complex process. Traditional 2D models, still widely used today, fail to replicate human skin’s intricate architecture and cell–cell interactions. Of note, 3D models, such as organoids, skin-on-chip, reconstructed human skin, and 3D bioprinting, offer experimental conditions closer to in vivo environments (Figure 2). Specifically, 3D models allow for more in-depth studies of physiological changes in the structure and function of the skin over time, including collagen degradation, reduced cellular vitality, and altered immune responses. Additionally, 3D models derived from aged patient cells could help develop new personalized anti-aging treatments. In addition, integrating skin microbiota into these models further enhances their realism and clinical translatability.
Despite promising advancements in this field, challenges remain, such as standardizing protocols, incorporating all relevant cell types, and recreating the exact physiological conditions of aged skin. Research and development are essential to overcome these obstacles and fully realize the potential of 3D skin models for studying aged skin. In summary, using advanced 3D skin models holds great promise for improving our understanding of skin aging and developing effective treatments, advancing scientific knowledge and clinical applications in the field.
Author Contributions
Conceptualization, F.L. and P.P.; methodology, F.L. and P.P.; writing—original draft preparation, F.L., F.R.A., A.C., V.C., S.A., B.C. and P.P.; writing—review and editing, B.C. and P.P.; supervision P.P.; funding acquisition, B.C. and P.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research and the APC were funded by the Department of Life, Health, and Environmental Sciences, University of L’Aquila, grants “FFO MeSVA 2022” and “FFO MeSVA 2023”.
Acknowledgments
The authors sincerely thank Maria Grazia Cifone for her valuable advice and for reviewing the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Russell, W.M. The development of the three Rs concept. Altern. Lab. Anim. 1995, 23, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Hubrecht, R.C.; Carter, E. The 3Rs and Humane Experimental Technique: Implementing Change. Animals 2019, 9, 754. [Google Scholar] [CrossRef] [PubMed]
- Henkler, F.; Tralau, T.; Tentschert, J.; Kneuer, C.; Haase, A.; Platzek, T.; Luch, A.; Gotz, M.E. Risk assessment of nanomaterials in cosmetics: A European union perspective. Arch. Toxicol. 2012, 86, 1641–1646. [Google Scholar] [CrossRef]
- Sanchez, M.M.; Bagdasarian, I.A.; Darch, W.; Morgan, J.T. Organotypic cultures as aging associated disease models. Aging-Us 2022, 14, 9338–9383. [Google Scholar] [CrossRef]
- Augello, F.R.; Lombardi, F.; Artone, S.; Ciafarone, A.; Altamura, S.; Di Marzio, L.; Cifone, M.G.; Palumbo, P.; Giuliani, M.; Cinque, B. Evaluation of the Effectiveness of an Innovative Polycomponent Formulation on Adult and Aged Human Dermal Fibroblasts. Biomedicines 2023, 11, 2410. [Google Scholar] [CrossRef]
- Quiles, J.; Cabrera, M.; Jones, J.; Tsapekos, M.; Caturla, N. In Vitro Determination of the Skin Anti-Aging Potential of Four-Component Plant-Based Ingredient. Molecules 2022, 27, 8101. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, I.C.; Meng, S.; Xu, J. LincRNA-EPS Promotes Proliferation of Aged Dermal Fibroblast by Inducing CCND1. Int. J. Mol. Sci. 2024, 25, 7677. [Google Scholar] [CrossRef] [PubMed]
- Maione, A.G.; Brudno, Y.; Stojadinovic, O.; Park, L.K.; Smith, A.; Tellechea, A.; Leal, E.C.; Kearney, C.J.; Veves, A.; Tomic-Canic, M.; et al. Three-dimensional human tissue models that incorporate diabetic foot ulcer-derived fibroblasts mimic in vivo features of chronic wounds. Tissue Eng. Part C Methods 2015, 21, 499–508. [Google Scholar] [CrossRef]
- Suarez-Martinez, E.; Suazo-Sanchez, I.; Celis-Romero, M.; Carnero, A. 3D and organoid culture in research: Physiology, hereditary genetic diseases and cancer. Cell Biosci. 2022, 12, 39. [Google Scholar] [CrossRef]
- Baker, P.; Huang, C.; Radi, R.; Moll, S.B.; Jules, E.; Arbiser, J.L. Skin Barrier Function: The Interplay of Physical, Chemical, and Immunologic Properties. Cells 2023, 12, 2745. [Google Scholar] [CrossRef]
- Ueck, C.; Volksdorf, T.; Houdek, P.; Vidal, Y.S.S.; Sehner, S.; Ellinger, B.; Lobmann, R.; Larena-Avellaneda, A.; Reinshagen, K.; Ridderbusch, I.; et al. Comparison of In-Vitro and Ex-Vivo Wound Healing Assays for the Investigation of Diabetic Wound Healing and Demonstration of a Beneficial Effect of a Triterpene Extract. PLoS ONE 2017, 12, e0169028. [Google Scholar] [CrossRef] [PubMed]
- Zoio, P.; Oliva, A. Skin-on-a-Chip Technology: Microengineering Physiologically Relevant In Vitro Skin Models. Pharmaceutics 2022, 14, 682. [Google Scholar] [CrossRef] [PubMed]
- Hayden, P.J.; Harbell, J.W. Special review series on 3D organotypic culture models: Introduction and historical perspective. In Vitro Cell. Dev. Biol. Anim. 2021, 57, 95–103. [Google Scholar] [CrossRef]
- Bell, E.; Ivarsson, B.; Merrill, C. Production of a tissue-like structure by contraction of collagen lattices by human fibroblasts of different proliferative potential in vitro. Proc. Natl. Acad. Sci. USA 1979, 76, 1274–1278. [Google Scholar] [CrossRef]
- Bell, E.; Ehrlich, H.P.; Buttle, D.J.; Nakatsuji, T. Living tissue formed in vitro and accepted as skin-equivalent tissue of full thickness. Science 1981, 211, 1052–1054. [Google Scholar] [CrossRef] [PubMed]
- Bell, E.; Ehrlich, H.P.; Sher, S.; Merrill, C.; Sarber, R.; Hull, B.; Nakatsuji, T.; Church, D.; Buttle, D.J. Development and use of a living skin equivalent. Plast. Reconstr. Surg. 1981, 67, 386–392. [Google Scholar] [CrossRef]
- Li, W.J.; Laurencin, C.T.; Caterson, E.J.; Tuan, R.S.; Ko, F.K. Electrospun nanofibrous structure: A novel scaffold for tissue engineering. J. Biomed. Mater. Res. 2002, 60, 613–621. [Google Scholar] [CrossRef]
- Rheinwald, J.G.; Green, H. Serial cultivation of strains of human epidermal keratinocytes: The formation of keratinizing colonies from single cells. Cell 1975, 6, 331–343. [Google Scholar] [CrossRef]
- Itoh, M.; Kiuru, M.; Cairo, M.S.; Christiano, A.M. Generation of keratinocytes from normal and recessive dystrophic epidermolysis bullosa-induced pluripotent stem cells. Proc. Natl. Acad. Sci. USA 2011, 108, 8797–8802. [Google Scholar] [CrossRef]
- Itoh, M.; Umegaki-Arao, N.; Guo, Z.; Liu, L.; Higgins, C.A.; Christiano, A.M. Generation of 3D skin equivalents fully reconstituted from human induced pluripotent stem cells (iPSCs). PLoS ONE 2013, 8, e77673. [Google Scholar] [CrossRef]
- Guenou, H.; Nissan, X.; Larcher, F.; Feteira, J.; Lemaitre, G.; Saidani, M.; Del Rio, M.; Barrault, C.C.; Bernard, F.X.; Peschanski, M.; et al. Human embryonic stem-cell derivatives for full reconstruction of the pluristratified epidermis: A preclinical study. Lancet 2009, 374, 1745–1753. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Rabbani, C.C.; Gao, H.; Steinhart, M.R.; Woodruff, B.M.; Pflum, Z.E.; Kim, A.; Heller, S.; Liu, Y.; Shipchandler, T.Z.; et al. Hair-bearing human skin generated entirely from pluripotent stem cells. Nature 2020, 582, 399–404. [Google Scholar] [CrossRef]
- Fentem, J.H.; Botham, P.A. ECVAM’s activities in validating alternative tests for skin corrosion and irritation. Altern. Lab. Anim. 2002, 30 (Suppl. 2), 61–67. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Suwa, F.; Wang, X.; Takemura, A.; Fang, Y.R.; Li, Y.; Zhao, Y.; Jin, Y. Reconstruction of a tissue-engineered skin containing melanocytes. Cell Biol. Int. 2007, 31, 985–990. [Google Scholar] [CrossRef]
- Diekmann, J.; Alili, L.; Scholz, O.; Giesen, M.; Holtkotter, O.; Brenneisen, P. A three-dimensional skin equivalent reflecting some aspects of in vivo aged skin. Exp. Dermatol. 2016, 25, 56–61. [Google Scholar] [CrossRef]
- Lim, H.Y.; Kim, J.; Song, H.J.; Kim, K.; Choi, K.C.; Park, S.; Sung, G.Y. Development of wrinkled skin-on-a-chip (WSOC) by cyclic uniaxial stretching. J. Ind. Eng. Chem. 2018, 68, 238–245. [Google Scholar] [CrossRef]
- Jeong, S.; Kim, J.; Jeon, H.M.; Kim, K.; Sung, G.Y. Development of an Aged Full-Thickness Skin Model Using Flexible Skin-on-a-Chip Subjected to Mechanical Stimulus Reflecting the Circadian Rhythm. Int. J. Mol. Sci. 2021, 22, 2788. [Google Scholar] [CrossRef] [PubMed]
- Pauty, J.; Nakano, S.; Usuba, R.; Nakajima, T.; Johmura, Y.; Omori, S.; Sakamoto, N.; Kikuchi, A.; Nakanishi, M.; Matsunaga, Y.T. A 3D tissue model-on-a-chip for studying the effects of human senescent fibroblasts on blood vessels. Biomater. Sci. 2021, 9, 199–211. [Google Scholar] [CrossRef]
- Lee, W.; Debasitis, J.C.; Lee, V.K.; Lee, J.H.; Fischer, K.; Edminster, K.; Park, J.K.; Yoo, S.S. Multi-layered culture of human skin fibroblasts and keratinocytes through three-dimensional freeform fabrication. Biomaterials 2009, 30, 1587–1595. [Google Scholar] [CrossRef]
- Min, D.; Lee, W.; Bae, I.H.; Lee, T.R.; Croce, P.; Yoo, S.S. Bioprinting of biomimetic skin containing melanocytes. Exp. Dermatol. 2018, 27, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Kim, M.; Kim, M.; Park, C.; Yoon, Y.; Lim, D.H.; Yeo, H.; Kang, S.; Lee, Y.G.; Beak, N.I.; et al. Spermidine-induced recovery of human dermal structure and barrier function by skin microbiome. Commun. Biol. 2021, 4, 231. [Google Scholar] [CrossRef] [PubMed]
- Rikken, G.; Meesters, L.D.; Jansen, P.A.M.; Rodijk-Olthuis, D.; van Vlijmen-Willems, I.; Niehues, H.; Smits, J.P.H.; Olah, P.; Homey, B.; Schalkwijk, J.; et al. Novel methodologies for host-microbe interactions and microbiome-targeted therapeutics in 3D organotypic skin models. Microbiome 2023, 11, 227. [Google Scholar] [CrossRef]
- Medawar, P.B. The cultivation of adult mammalian skin epithelium in vitro. Q. J. Microsc. Sci. 1948, 89, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Mori, N.; Morimoto, Y.; Takeuchi, S. Skin integrated with perfusable vascular channels on a chip. Biomaterials 2017, 116, 48–56. [Google Scholar] [CrossRef]
- Pitrez, P.R.; Monteiro, L.M.; Borgogno, O.; Nissan, X.; Mertens, J.; Ferreira, L. Cellular reprogramming as a tool to model human aging in a dish. Nat. Commun. 2024, 15, 1816. [Google Scholar] [CrossRef]
- Costello, L.; Dicolandrea, T.; Tasseff, R.; Isfort, R.; Bascom, C.; von Zglinicki, T.; Przyborski, S. Tissue engineering strategies to bioengineer the ageing skin phenotype in vitro. Aging Cell 2022, 21, e13550. [Google Scholar] [CrossRef]
- Dos Santos, M.; Metral, E.; Boher, A.; Rousselle, P.; Thepot, A.; Damour, O. In vitro 3-D model based on extending time of culture for studying chronological epidermis aging. Matrix Biol. 2015, 47, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Lammermann, I.; Terlecki-Zaniewicz, L.; Weinmullner, R.; Schosserer, M.; Dellago, H.; de Matos Branco, A.D.; Autheried, D.; Sevcnikar, B.; Kleissl, L.; Berlin, I.; et al. Blocking negative effects of senescence in human skin fibroblasts with a plant extract. NPJ Aging Mech. Dis. 2018, 4, 4. [Google Scholar] [CrossRef]
- Weinmullner, R.; Zbiral, B.; Becirovic, A.; Stelzer, E.M.; Nagelreiter, F.; Schosserer, M.; Lammermann, I.; Liendl, L.; Lang, M.; Terlecki-Zaniewicz, L.; et al. Organotypic human skin culture models constructed with senescent fibroblasts show hallmarks of skin aging. NPJ Aging Mech. Dis. 2020, 6, 4. [Google Scholar] [CrossRef]
- Victorelli, S.; Lagnado, A.; Halim, J.; Moore, W.; Talbot, D.; Barrett, K.; Chapman, J.; Birch, J.; Ogrodnik, M.; Meves, A.; et al. Senescent human melanocytes drive skin ageing via paracrine telomere dysfunction. EMBO J. 2019, 38, e101982. [Google Scholar] [CrossRef]
- Markiewicz, E.; Jerome, J.; Mammone, T.; Idowu, O.C. Anti-Glycation and Anti-Aging Properties of Resveratrol Derivatives in the in-vitro 3D Models of Human Skin. Clin. Cosmet. Investig. Dermatol. 2022, 15, 911–927. [Google Scholar] [CrossRef]
- Lee, K.H.; Ng, Y.P.; Cheah, P.S.; Lim, C.K.; Toh, M.S. Molecular characterization of glycation-associated skin ageing: An alternative skin model to study in vitro antiglycation activity of topical cosmeceutical and pharmaceutical formulations. Br. J. Dermatol. 2017, 176, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Yang, T.; Yu, D.; Xiong, H.; Zhang, S. Current insights and future perspectives of ultraviolet radiation (UV) exposure: Friends and foes to the skin and beyond the skin. Environ. Int. 2024, 185, 108535. [Google Scholar] [CrossRef]
- Chen, T.; Hou, H.; Fan, Y.; Wang, S.; Chen, Q.; Si, L.; Li, B. Protective effect of gelatin peptides from pacific cod skin against photoaging by inhibiting the expression of MMPs via MAPK signaling pathway. J. Photochem. Photobiol. B. 2016, 165, 34–41. [Google Scholar] [CrossRef]
- Gu, Y.; Han, J.; Jiang, C.; Zhang, Y. Biomarkers, oxidative stress and autophagy in skin aging. Ageing Res. Rev. 2020, 59, 101036. [Google Scholar] [CrossRef] [PubMed]
- Casale, C.; Imparato, G.; Urciuolo, F.; Rescigno, F.; Scamardella, S.; Escolino, M.; Netti, P.A. Engineering a human skin equivalent to study dermis remodelling and epidermis senescence in vitro after UVA exposure. J. Tissue Eng. Regen. Med. 2018, 12, 1658–1669. [Google Scholar] [CrossRef]
- Bouzos, E.; Asuri, P. Sandwich Culture Platforms to Investigate the Roles of Stiffness Gradients and Cell-Matrix Adhesions in Cancer Cell Migration. Cancers 2023, 15, 1729. [Google Scholar] [CrossRef] [PubMed]
- Rebehn, L.; Khalaji, S.; KleinJan, F.; Kleemann, A.; Port, F.; Paul, P.; Huster, C.; Nolte, U.; Singh, K.; Kwapich, L.; et al. The weakness of senescent dermal fibroblasts. Proc. Natl. Acad. Sci. USA 2023, 120, e2301880120. [Google Scholar] [CrossRef]
- Sato, T.; Vries, R.G.; Snippert, H.J.; van de Wetering, M.; Barker, N.; Stange, D.E.; van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J.; et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef]
- Hu, J.L.; Todhunter, M.E.; LaBarge, M.A.; Gartner, Z.J. Opportunities for organoids as new models of aging. J. Cell Biol. 2018, 217, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Koehler, K.R. Skin organoids: A new human model for developmental and translational research. Exp. Dermatol. 2021, 30, 613–620. [Google Scholar] [CrossRef]
- Zhao, Z.; Chen, X.; Dowbaj, A.M.; Sljukic, A.; Bratlie, K.; Lin, L.; Fong, E.L.S.; Balachander, G.M.; Chen, Z.; Soragni, A.; et al. Organoids. Nat. Rev. Methods Primers 2022, 2, 94. [Google Scholar] [CrossRef]
- Hong, Y.J.; Do, J.T. Neural Lineage Differentiation From Pluripotent Stem Cells to Mimic Human Brain Tissues. Front. Bioeng. Biotechnol. 2019, 7, 400. [Google Scholar] [CrossRef]
- Drost, J.; Clevers, H. Organoids in cancer research. Nat. Rev. Cancer 2018, 18, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. Modeling Development and Disease with Organoids. Cell 2016, 165, 1586–1597. [Google Scholar] [CrossRef] [PubMed]
- Ryu, N.E.; Lee, S.H.; Park, H. Spheroid Culture System Methods and Applications for Mesenchymal Stem Cells. Cells 2019, 8, 1620. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, F.; Augello, F.R.; Artone, S.; Ciafarone, A.; Topi, S.; Cifone, M.G.; Cinque, B.; Palumbo, P. Involvement of Cyclooxygenase-2 in Establishing an Immunosuppressive Microenvironment in Tumorspheres Derived from TMZ-Resistant Glioblastoma Cell Lines and Primary Cultures. Cells 2024, 13, 258. [Google Scholar] [CrossRef]
- Silva-Pedrosa, R.; Salgado, A.J.; Ferreira, P.E. Revolutionizing Disease Modeling: The Emergence of Organoids in Cellular Systems. Cells 2023, 12, 930. [Google Scholar] [CrossRef]
- Qu, S.; Xu, R.; Yi, G.; Li, Z.; Zhang, H.; Qi, S.; Huang, G. Patient-derived organoids in human cancer: A platform for fundamental research and precision medicine. Mol. Biomed. 2024, 5, 6. [Google Scholar] [CrossRef]
- Torrens-Mas, M.; Perello-Reus, C.; Navas-Enamorado, C.; Ibarguen-Gonzalez, L.; Sanchez-Polo, A.; Segura-Sampedro, J.J.; Masmiquel, L.; Barcelo, C.; Gonzalez-Freire, M. Organoids: An Emerging Tool to Study Aging Signature across Human Tissues. Modeling Aging with Patient-Derived Organoids. Int. J. Mol. Sci. 2021, 22, 547. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, M.; Koehler, K.R.; Shafiee, A. Biofabrication of Human Skin with Its Appendages. Adv. Healthc. Mater. 2022, 11, e2201626. [Google Scholar] [CrossRef] [PubMed]
- Cavallero, S.; Neves Granito, R.; Stockholm, D.; Azzolin, P.; Martin, M.T.; Fortunel, N.O. Exposure of Human Skin Organoids to Low Genotoxic Stress Can Promote Epithelial-to-Mesenchymal Transition in Regenerating Keratinocyte Precursor Cells. Cells 2020, 9, 1912. [Google Scholar] [CrossRef]
- Metallo, C.M.; Ji, L.; de Pablo, J.J.; Palecek, S.P. Retinoic acid and bone morphogenetic protein signaling synergize to efficiently direct epithelial differentiation of human embryonic stem cells. Stem Cells 2008, 26, 372–380. [Google Scholar] [CrossRef]
- Miyake, T.; Shimada, M.; Matsumoto, Y.; Okino, A. DNA Damage Response After Ionizing Radiation Exposure in Skin Keratinocytes Derived from Human-Induced Pluripotent Stem Cells. Int. J. Radiat. Oncol. Biol. Phys. 2019, 105, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Feldman, A.; Mukha, D.; Maor, I.I.; Sedov, E.; Koren, E.; Yosefzon, Y.; Shlomi, T.; Fuchs, Y. Blimp1(+) cells generate functional mouse sebaceous gland organoids in vitro. Nat. Commun. 2019, 10, 2348. [Google Scholar] [CrossRef] [PubMed]
- Diao, J.; Liu, J.; Wang, S.; Chang, M.; Wang, X.; Guo, B.; Yu, Q.; Yan, F.; Su, Y.; Wang, Y. Sweat gland organoids contribute to cutaneous wound healing and sweat gland regeneration. Cell Death Dis. 2019, 10, 238. [Google Scholar] [CrossRef]
- Rossi, G.; Manfrin, A.; Lutolf, M.P. Progress and potential in organoid research. Nat. Rev. Genet. 2018, 19, 671–687. [Google Scholar] [CrossRef]
- Sun, X.; Sun, F.; Zhang, Y.; Qu, J.; Zhang, W.; Liu, G.-H. A narrative review of organoids for investigating organ aging: Opportunities and challenges. J. Bio-XResearch 2023, 6, 3–14. [Google Scholar] [CrossRef]
- Kim, M.J.; Ahn, H.J.; Kong, D.; Lee, S.; Kim, D.H.; Kang, K.S. Modeling of solar UV-induced photodamage on the hair follicles in human skin organoids. J. Tissue Eng. 2024, 15, 20417314241248753. [Google Scholar] [CrossRef]
- Ding, X.; Kakanj, P.; Leptin, M.; Eming, S.A. Regulation of the Wound Healing Response during Aging. J. Investig. Dermatol. 2021, 141, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Ju, J.H. Generation of 3D Skin Organoid from Cord Blood-derived Induced Pluripotent Stem Cells. J. Vis. Exp. 2019, 146, e59297. [Google Scholar] [CrossRef]
- Oceguera-Yanez, F.; Avila-Robinson, A.; Woltjen, K. Differentiation of pluripotent stem cells for modeling human skin development and potential applications. Front. Cell Dev. Biol. 2022, 10, 1030339. [Google Scholar] [CrossRef] [PubMed]
- Dash, B.C.; Korutla, L.; Vallabhajosyula, P.; Hsia, H.C. Unlocking the Potential of Induced Pluripotent Stem Cells for Wound Healing: The Next Frontier of Regenerative Medicine. Adv. Wound Care 2022, 11, 622–638. [Google Scholar] [CrossRef]
- Lo Sardo, V.; Ferguson, W.; Erikson, G.A.; Topol, E.J.; Baldwin, K.K.; Torkamani, A. Influence of donor age on induced pluripotent stem cells. Nat. Biotechnol. 2017, 35, 69–74. [Google Scholar] [CrossRef]
- Mahmoudi, S.; Brunet, A. Aging and reprogramming: A two-way street. Curr. Opin. Cell Biol. 2012, 24, 744–756. [Google Scholar] [CrossRef]
- Kim, J.; Koo, B.K.; Knoblich, J.A. Human organoids: Model systems for human biology and medicine. Nat. Rev. Mol. Cell Biol. 2020, 21, 571–584. [Google Scholar] [CrossRef]
- Asselineau, D.; Bernard, B.A.; Bailly, C.; Darmon, M.; Prunieras, M. Human epidermis reconstructed by culture: Is it “normal”? J. Investig. Dermatol. 1986, 86, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Prunieras, M.; Regnier, M.; Woodley, D. Methods for cultivation of keratinocytes with an air-liquid interface. J. Investig. Dermatol. 1983, 81, 28s–33s. [Google Scholar] [CrossRef]
- Hofmann, E.; Schwarz, A.; Fink, J.; Kamolz, L.P.; Kotzbeck, P. Modelling the Complexity of Human Skin In Vitro. Biomedicines 2023, 11, 794. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, J.E.; Kim, B.J.; Cho, K.H. In vitro phototoxicity test using artificial skin with melanocytes. Photodermatol. Photoimmunol. Photomed. 2007, 23, 73–80. [Google Scholar] [CrossRef]
- Hall, M.J.; Lopes-Ventura, S.; Neto, M.V.; Charneca, J.; Zoio, P.; Seabra, M.C.; Oliva, A.; Barral, D.C. Reconstructed human pigmented skin/epidermis models achieve epidermal pigmentation through melanocore transfer. Pigment. Cell Melanoma Res. 2022, 35, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Nedachi, T.; Bonod, C.; Rorteau, J.; Chinoune, W.; Ishiuchi, Y.; Hughes, S.; Gillet, B.; Bechetoille, N.; Sigaudo-Roussel, D.; Lamartine, J. Chronological aging impacts abundance, function and microRNA content of extracellular vesicles produced by human epidermal keratinocytes. Aging 2023, 15, 12702–12722. [Google Scholar] [CrossRef]
- Cho, S.W.; Malick, H.; Kim, S.J.; Grattoni, A. Advances in Skin-on-a-Chip Technologies for Dermatological Disease Modeling. J. Investig. Dermatol. 2024, 144, 1707–1715. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Sito, L.; Mao, M.; He, J.; Zhang, Y.S.; Zhao, X. Current advances in skin-on-a-chip models for drug testing. Microphysiol. Syst. 2018, 2, 4. [Google Scholar] [CrossRef]
- Abaci, H.E.; Gledhill, K.; Guo, Z.Y.; Christiano, A.M.; Shuler, M.L. Pumpless microfluidic platform for drug testing on human skin equivalents. Lab Chip 2015, 15, 882–888. [Google Scholar] [CrossRef]
- Sutterby, E.; Thurgood, P.; Baratchi, S.; Khoshmanesh, K.; Pirogova, E. Microfluidic Skin-on-a-Chip Models: Toward Biomimetic Artificial Skin. Small 2020, 16, e2002515. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Getschman, A.E.; Hwang, S.; Volkman, B.F.; Klonisch, T.; Levin, D.; Zhao, M.; Santos, S.; Liu, S.; Cheng, J.; et al. Investigations on T cell transmigration in a human skin-on-chip (SoC) model. Lab Chip 2021, 21, 1527–1539. [Google Scholar] [CrossRef]
- Risueno, I.; Valencia, L.; Jorcano, J.L.; Velasco, D. Skin-on-a-chip models: General overview and future perspectives. APL Bioeng. 2021, 5, 030901. [Google Scholar] [CrossRef]
- Liu, H.; Xing, F.; Yu, P.; Zhe, M.; Duan, X.; Liu, M.; Xiang, Z.; Ritz, U. A review of biomacromolecule-based 3D bioprinting strategies for structure-function integrated repair of skin tissues. Int. J. Biol. Macromol. 2024, 268, 131623. [Google Scholar] [CrossRef]
- Ong, C.S.; Yesantharao, P.; Huang, C.Y.; Mattson, G.; Boktor, J.; Fukunishi, T.; Zhang, H.; Hibino, N. 3D bioprinting using stem cells. Pediatr. Res. 2018, 83, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.L.; Qi, J.T.Z.; Yeong, W.Y.; Naing, M.W. Proof-of-concept: 3D bioprinting of pigmented human skin constructs. Biofabrication 2018, 10, 025005. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Luo, Y.; Ma, Y.; Wang, P.; Yao, R. Converging bioprinting and organoids to better recapitulate the tumor microenvironment. Trends Biotechnol. 2024, 42, 648–663. [Google Scholar] [CrossRef] [PubMed]
- Arslan-Yildiz, A.; El Assal, R.; Chen, P.; Guven, S.; Inci, F.; Demirci, U. Towards artificial tissue models: Past, present, and future of 3D bioprinting. Biofabrication 2016, 8, 014103. [Google Scholar] [CrossRef]
- Matai, I.; Kaur, G.; Seyedsalehi, A.; McClinton, A.; Laurencin, C.T. Progress in 3D bioprinting technology for tissue/organ regenerative engineering. Biomaterials 2020, 226, 119536. [Google Scholar] [CrossRef]
- Li, Y.; Liu, J.; Xu, S.; Wang, J. 3D Bioprinting: An Important Tool for Tumor Microenvironment Research. Int. J. Nanomedicine 2023, 18, 8039–8057. [Google Scholar] [CrossRef]
- Ribezzi, D.; Barbaglio, F.; Pinos, R.; Bonetti, L.; Farè, S.; Scielzo, C. Design of a Novel Bioink Suitable for the 3d Printing of Lymphoid Cells. Front. Biomater. Sci. 2022, 28, S545. [Google Scholar] [CrossRef]
- Puistola, P.; Miettinen, S.; Skottman, H.; Moro, A. Novel strategy for multi-material 3D bioprinting of human stem cell based corneal stroma with heterogenous design. Mater. Today Bio 2024, 24, 100924. [Google Scholar] [CrossRef]
- Kang, M.S.; Jang, J.; Jo, H.J.; Kim, W.H.; Kim, B.; Chun, H.J.; Lim, D.; Han, D.W. Advances and Innovations of 3D Bioprinting Skin. Biomolecules 2022, 13, 55. [Google Scholar] [CrossRef]
- Zhang, Y.; Enhejirigala; Yao, B.; Li, Z.; Song, W.; Li, J.; Zhu, D.; Wang, Y.; Duan, X.; Yuan, X.; et al. Using bioprinting and spheroid culture to create a skin model with sweat glands and hair follicles. Burns Trauma. 2021, 9, tkab013. [Google Scholar] [CrossRef]
- Weng, T.T.; Zhang, W.; Xia, Y.L.; Wu, P.; Yang, M.; Jin, R.H.; Xia, S.Z.; Wang, J.L.; You, C.G.; Han, C.M.; et al. 3D bioprinting for skin tissue engineering: Current status and perspectives. J. Tissue Eng. 2021, 12, 20417314211028574. [Google Scholar] [CrossRef]
- Ansaf, R.B.; Ziebart, R.; Gudapati, H.; Simoes Torigoe, R.M.; Victorelli, S.; Passos, J.; Wyles, S.P. 3D bioprinting-a model for skin aging. Regen. Biomater. 2023, 10, rbad060. [Google Scholar] [CrossRef] [PubMed]
- Bhar, B.; Das, E.; Manikumar, K.; Mandal, B.B. 3D Bioprinted Human Skin Model Recapitulating Native-Like Tissue Maturation and Immunocompetence as an Advanced Platform for Skin Sensitization Assessment. Adv. Healthc. Mater. 2024, 13, e2303312. [Google Scholar] [CrossRef] [PubMed]
- Olejnik, A.; Semba, J.A.; Kulpa, A.; Danczak-Pazdrowska, A.; Rybka, J.D.; Gornowicz-Porowska, J. 3D Bioprinting in Skin Related Research: Recent Achievements and Application Perspectives. ACS Synth. Biol. 2022, 11, 26–38. [Google Scholar] [CrossRef]
- Sun, W.; Wang, B.; Yang, T.; Yin, R.; Wang, F.; Zhang, H.; Zhang, W. Three-Dimensional Bioprinted Skin Microrelief and Its Role in Skin Aging. Biomimetics 2024, 9, 366. [Google Scholar] [CrossRef] [PubMed]
- Shboul, S.A.; DeLuca, V.J.; Dweiri, Y.A.; Saleh, T. Can 3D bioprinting solve the mystery of senescence in cancer therapy? Ageing Res. Rev. 2022, 81, 101732. [Google Scholar] [CrossRef]
- Prasanna, P.G.; Citrin, D.E.; Hildesheim, J.; Ahmed, M.M.; Venkatachalam, S.; Riscuta, G.; Xi, D.; Zheng, G.; Deursen, J.V.; Goronzy, J.; et al. Therapy-Induced Senescence: Opportunities to Improve Anticancer Therapy. J. Natl. Cancer Inst. 2021, 113, 1285–1298. [Google Scholar] [CrossRef]
- Szabo, K.; Bolla, B.S.; Erdei, L.; Balogh, F.; Kemeny, L. Are the Cutaneous Microbiota a Guardian of the Skin’s Physical Barrier? The Intricate Relationship between Skin Microbes and Barrier Integrity. Int. J. Mol. Sci. 2023, 24, 5962. [Google Scholar] [CrossRef]
- Boxberger, M.; Cenizo, V.; Cassir, N.; La Scola, B. Challenges in exploring and manipulating the human skin microbiome. Microbiome 2021, 9, 125. [Google Scholar] [CrossRef]
- Meisel, J.S.; Sfyroera, G.; Bartow-McKenney, C.; Gimblet, C.; Bugayev, J.; Horwinski, J.; Kim, B.; Brestoff, J.R.; Tyldsley, A.S.; Zheng, Q.; et al. Commensal microbiota modulate gene expression in the skin. Microbiome 2018, 6, 20. [Google Scholar] [CrossRef]
- Bolla, B.S.; Erdei, L.; Urban, E.; Burian, K.; Kemeny, L.; Szabo, K. Cutibacterium acnes regulates the epidermal barrier properties of HPV-KER human immortalized keratinocyte cultures. Sci. Rep. 2020, 10, 12815. [Google Scholar] [CrossRef] [PubMed]
- Pivarcsi, A.; Bodai, L.; Rethi, B.; Kenderessy-Szabo, A.; Koreck, A.; Szell, M.; Beer, Z.; Bata-Csorgoo, Z.; Magocsi, M.; Rajnavolgyi, E.; et al. Expression and function of Toll-like receptors 2 and 4 in human keratinocytes. Int. Immunol. 2003, 15, 721–730. [Google Scholar] [CrossRef]
- Larson, P.J.; Chong, D.; Fleming, E.; Oh, J. Challenges in Developing a Human Model System for Skin Microbiome Research. J. Investig. Dermatol. 2021, 141, 228–231.e4. [Google Scholar] [CrossRef]
- Emmert, H.; Rademacher, F.; Glaser, R.; Harder, J. Skin microbiota analysis in human 3D skin models-”Free your mice”. Exp. Dermatol. 2020, 29, 1133–1139. [Google Scholar] [CrossRef] [PubMed]
- Landemaine, L.; Cenizo, V.; Lemaire, G.J.; Portes, P. 961 Colonization of a 3D skin model with a complete microbiota is more beneficial to the skin barrier than with Staphylococcus epidermidis alone. J. Investig. Dermatol. 2018, 138, S163. [Google Scholar] [CrossRef]
- Loomis, K.H.; Wu, S.K.; Ernlund, A.; Zudock, K.; Reno, A.; Blount, K.; Karig, D.K. A mixed community of skin microbiome representatives influences cutaneous processes more than individual members. Microbiome 2021, 9, 22. [Google Scholar] [CrossRef]
- Myers, T.; Bouslimani, A.; Huang, S.; Hansen, S.T.; Clavaud, C.; Azouaoui, A.; Ott, A.; Gueniche, A.; Bouez, C.; Zheng, Q.; et al. A multi-study analysis enables identification of potential microbial features associated with skin aging signs. Front. Aging 2023, 4, 1304705. [Google Scholar] [CrossRef]
- Janson, D.; Rietveld, M.; Willemze, R.; El Ghalbzouri, A. Effects of serially passaged fibroblasts on dermal and epidermal morphogenesis in human skin equivalents. Biogerontology 2013, 14, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Adamus, J.; Aho, S.; Meldrum, H.; Bosko, C.; Lee, J.M. p16INK4A influences the aging phenotype in the living skin equivalent. J. Investig. Dermatol. 2014, 134, 1131–1133. [Google Scholar] [CrossRef]
- Wedel, S.; Martic, I.; Hrapovic, N.; Fabre, S.; Madreiter-Sokolowski, C.T.; Haller, T.; Pierer, G.; Ploner, C.; Jansen-Dürr, P.; Cavinato, M. tBHP treatment as a model for cellular senescence and pollution-induced skin aging. Mech. Ageing Dev. 2020, 190, 111318. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, S.; Luo, D.; Chen, D.; Zhou, H.; Zhang, S.; Chen, X.; Lu, W.; Liu, W. Systematic Study of Resveratrol Nanoliposomes Transdermal Delivery System for Enhancing Anti-Aging and Skin-Brightening Efficacy. Molecules 2023, 28, 2738. [Google Scholar] [CrossRef] [PubMed]
- Wedel, S.; Martic, I.; Navarro, L.G.; Ploner, C.; Pierer, G.; Jansen-Dürr, P.; Cavinato, M. Depletion of growth differentiation factor 15 (GDF15) leads to mitochondrial dysfunction and premature senescence in human dermal fibroblasts. Aging Cell 2023, 22, e13752. [Google Scholar] [CrossRef]
- Klinngam, W.; Rungkamoltip, P.; Wongwanakul, R.; Joothamongkhon, J.; Du, A.M.S.; Khongkow, M.; Asawapirom, U.; Iempridee, T.; Ruktanonchai, U. Skin Rejuvenation Efficacy and Safety Evaluation of Kaempferia parviflora Standardized Extract (BG100) in Human 3D Skin Models and Clinical Trial. Biomolecules 2024, 14, 776. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Jung, Y.S.; Cho, Y.R.; Seo, J.W.; Lim, W.C.; Nam, T.G.; Lim, T.G.; Byun, S. Oral Administration of Prevents UVB-Induced Skin Aging through Targeting the c-Raf Signaling Axis. Antioxidants 2021, 10, 1663. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.O.; Smith, A.; Liu, Y.; Du, P.; Blumberg, J.B.; Garlick, J. Photoprotection by pistachio bioactives in a 3-dimensional human skin equivalent tissue model. Int. J. Food Sci. Nutr. 2017, 68, 712–718. [Google Scholar] [CrossRef]
- Yonezawa, T.; Momota, R.; Iwano, H.; Zhao, S.; Hakozaki, T.; Soh, C.; Sawaki, S.; Toyama, K.; Oohashi, T. Unripe peach (Prunus persica) extract ameliorates damage from UV irradiation and improved collagen XVIII expression in 3D skin model. J. Cosmet. Dermatol. 2019, 18, 1507–1515. [Google Scholar] [CrossRef]
- Fernandez-Carro, E.; Remacha, A.R.; Orera, I.; Lattanzio, G.; Garcia-Barrios, A.; del Barrio, J.; Alcaine, C.; Ciriza, J. Human Dermal Decellularized ECM Hydrogels as Scaffolds for 3D In Vitro Skin Aging Models. Int. J. Mol. Sci. 2024, 25, 4020. [Google Scholar] [CrossRef]
- Min, D.; Ahn, Y.; Lee, H.K.; Jung, W.; Kim, H.J. A novel optical coherence tomography-based in vitro method of anti-aging skin analysis using 3D skin wrinkle mimics. Skin Res. Technol. 2023, 29, e13354. [Google Scholar] [CrossRef]
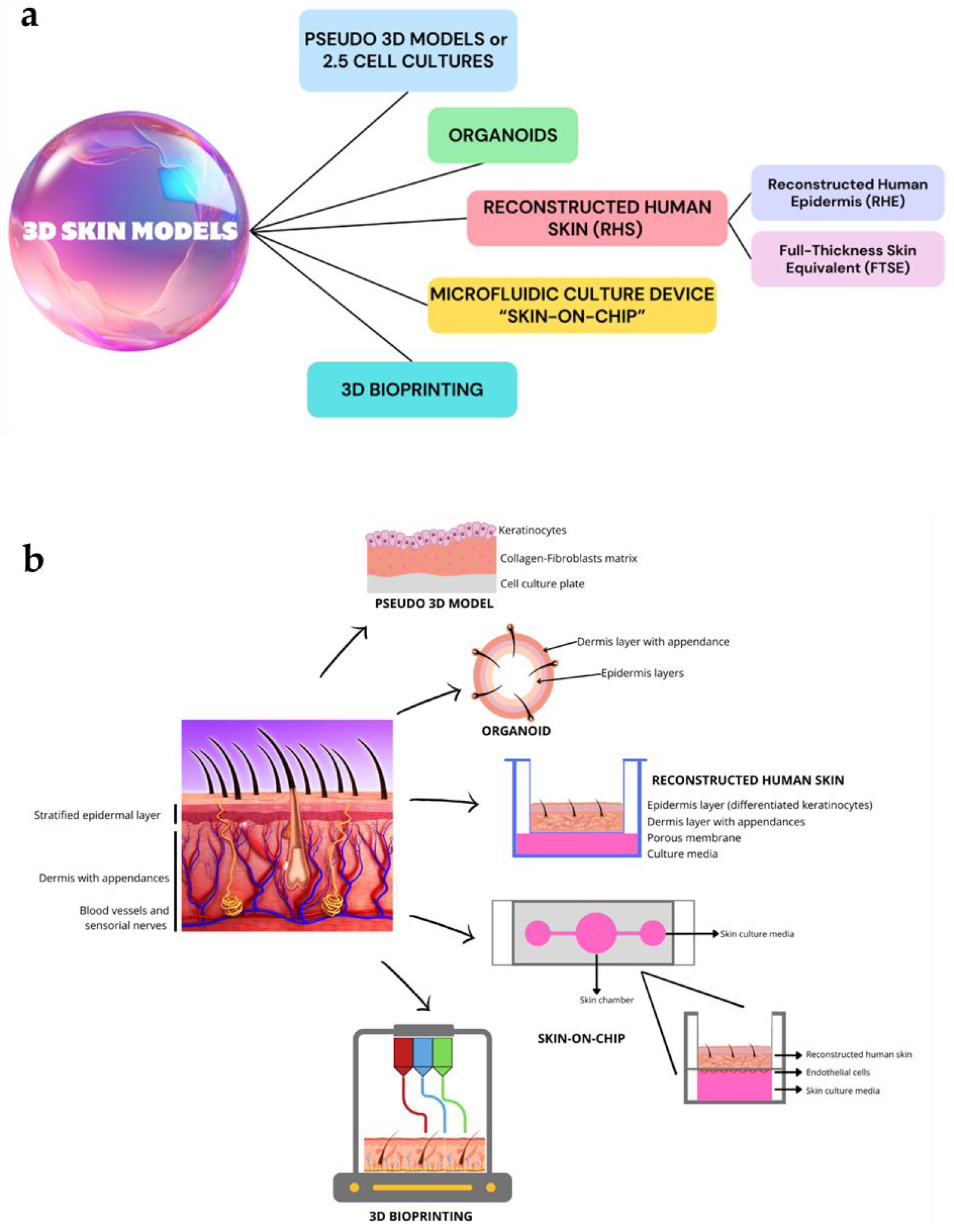
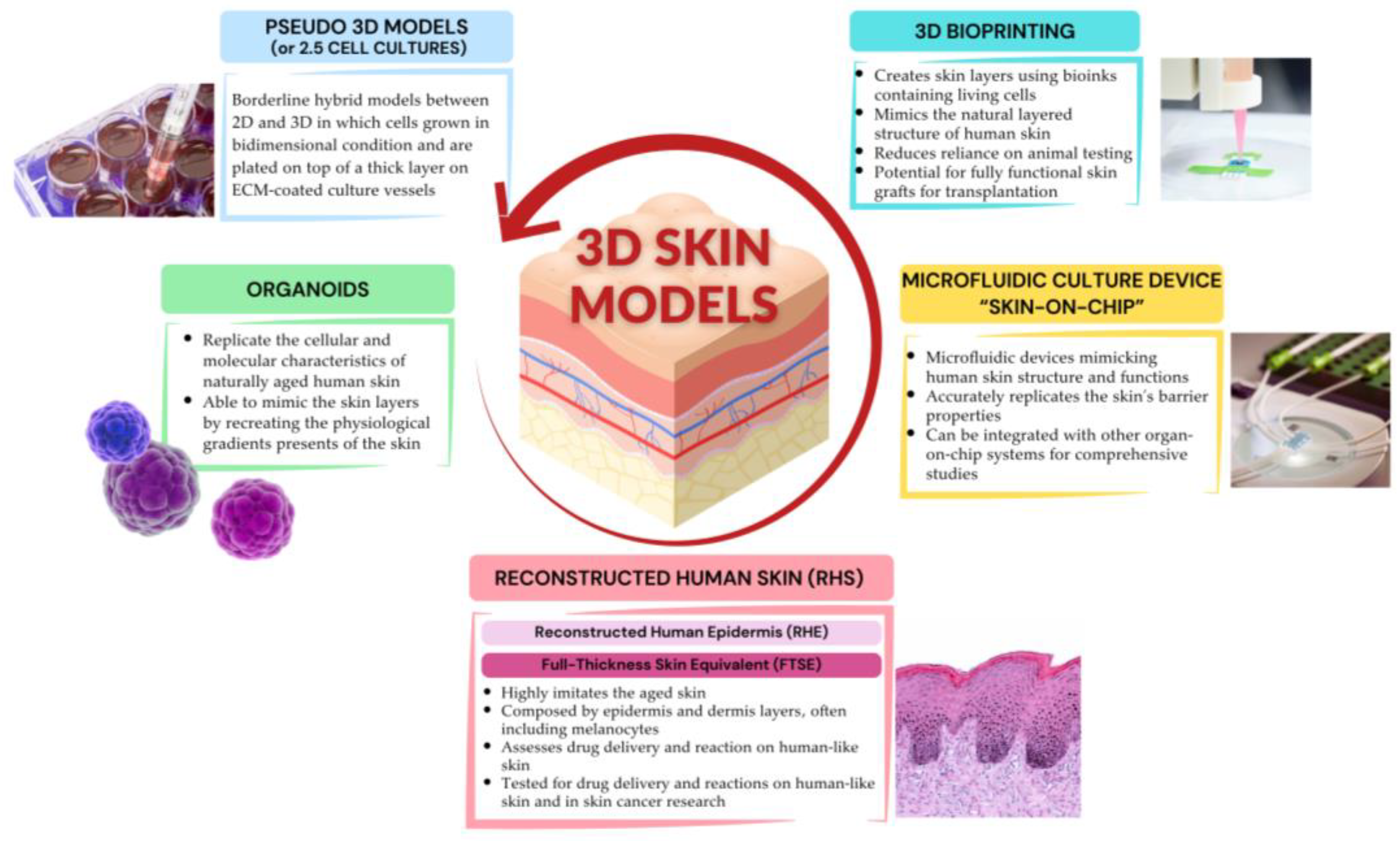
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).