

Planarian Mucus: A Novel Source of Pleiotropic Cytotoxic and Cytostatic Agents against Cancer Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Planarian Rearing
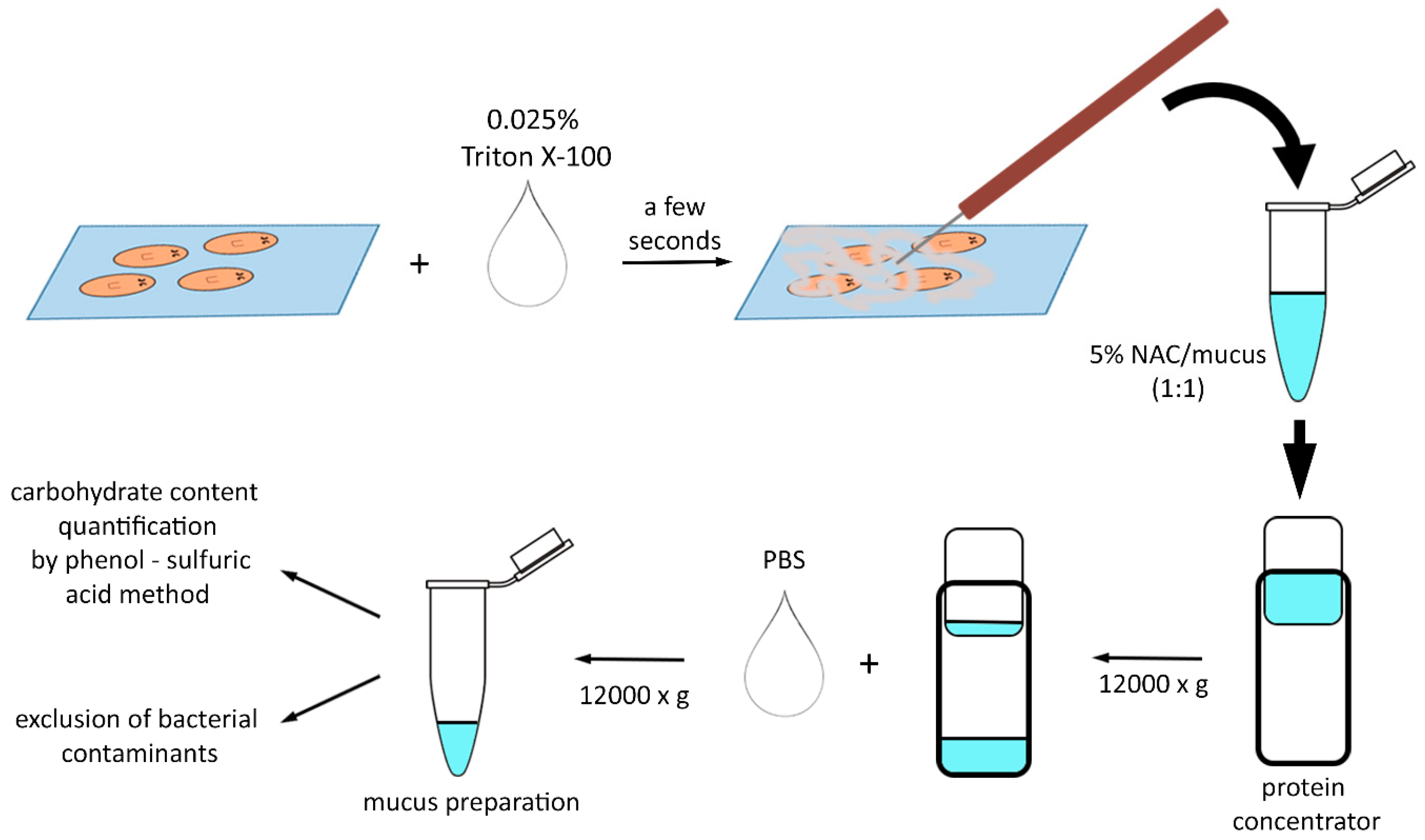
2.2. Planarian Mucus Preparation, Quantification, Storage and Treatment
2.3. Cell Cultures
2.4. Crystal Violet Assay
2.5. Propidium Iodide Viability Assay
2.6. Cell Cycle Analysis
2.7. JC1 Assay
2.8. Annexin V/PI Assay
2.9. Analysis of Lipid Content
2.10. Reactive Oxygen Species (ROS) Detection
2.11. Phospho-Histone 3 Immunostaining
2.12. Transmission Electron Microscopy
2.13. Statistical Analysis
3. Results
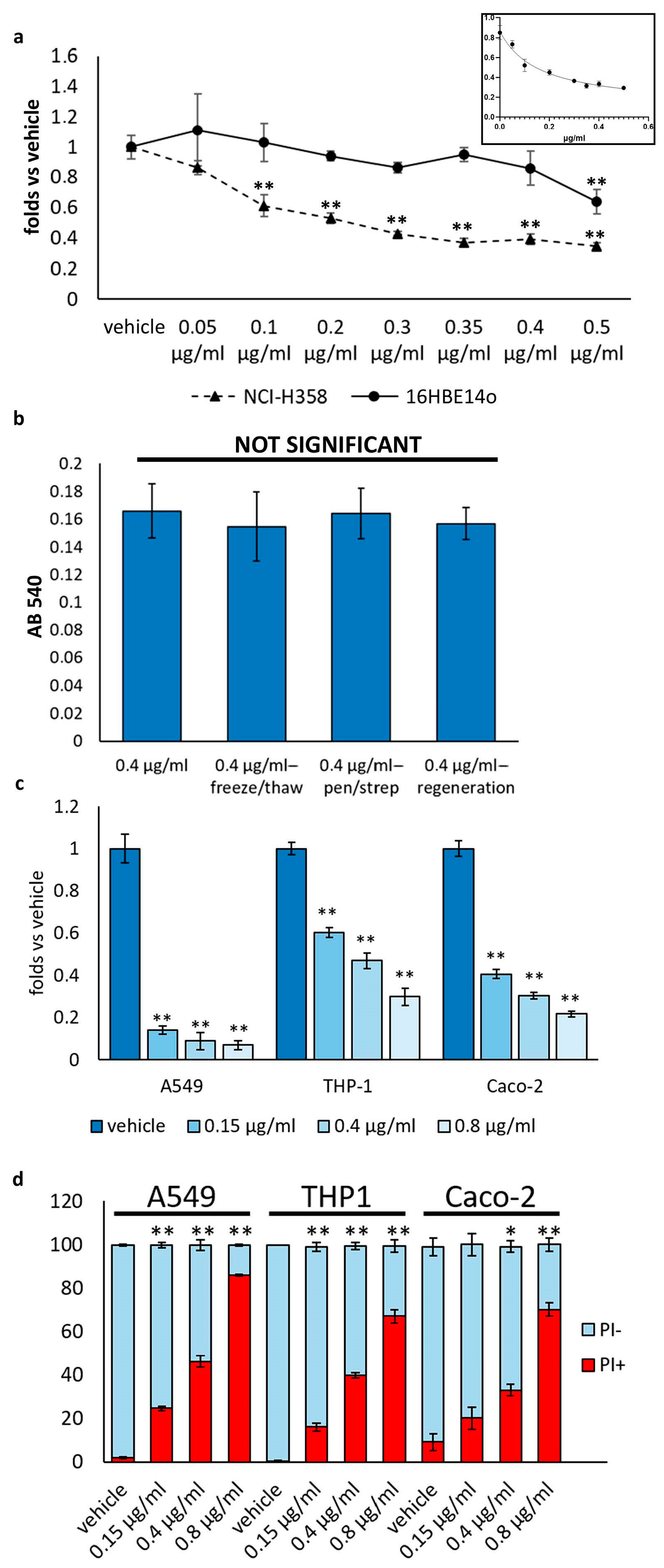
3.1. D. japonica Mucus Exhibited Inhibitory Effect on the Growth of Human Carcinoma Cells
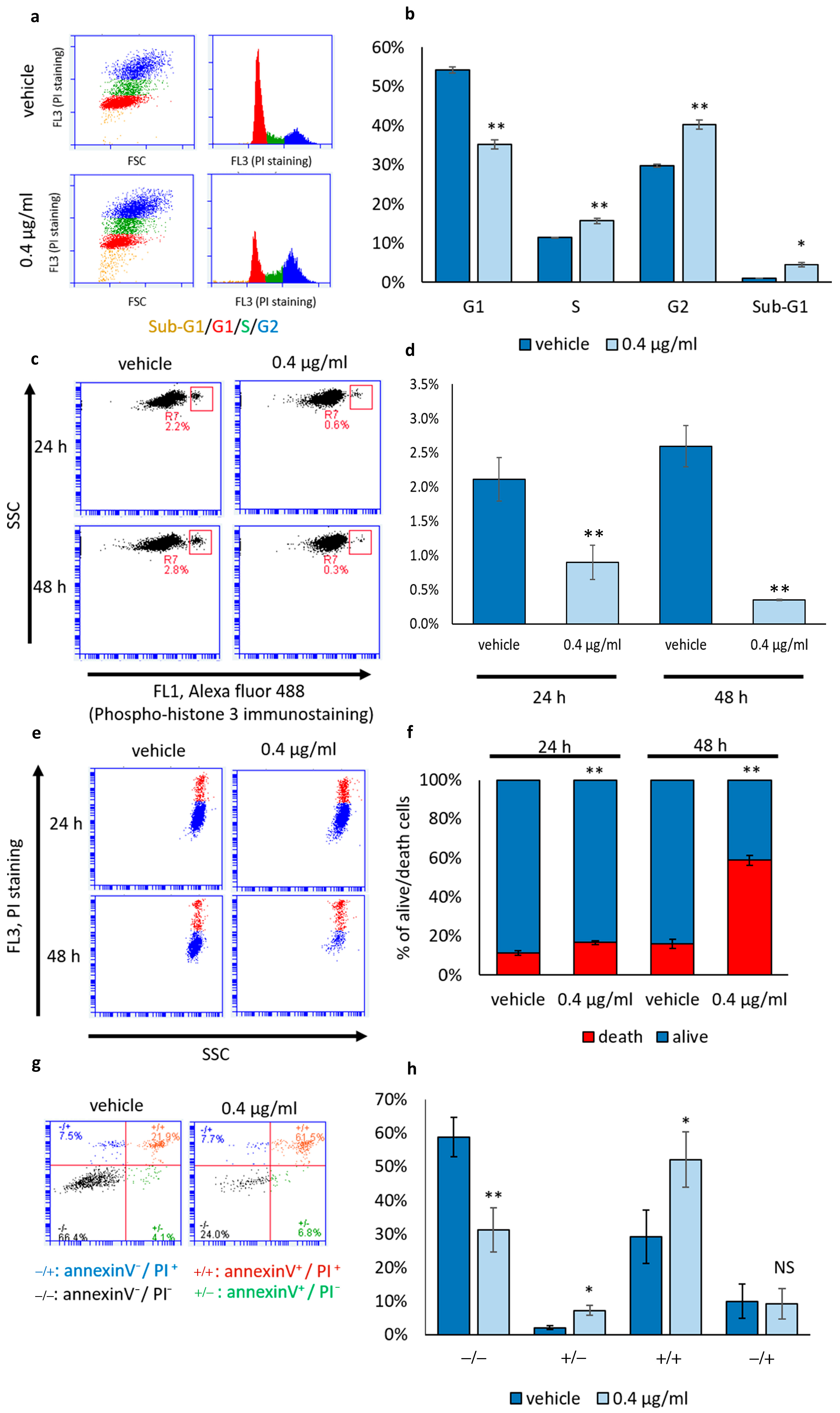
3.2. Mucus Treatment Induces Both Cytostatic and Cytotoxic Effects on NCI-H358 Cells
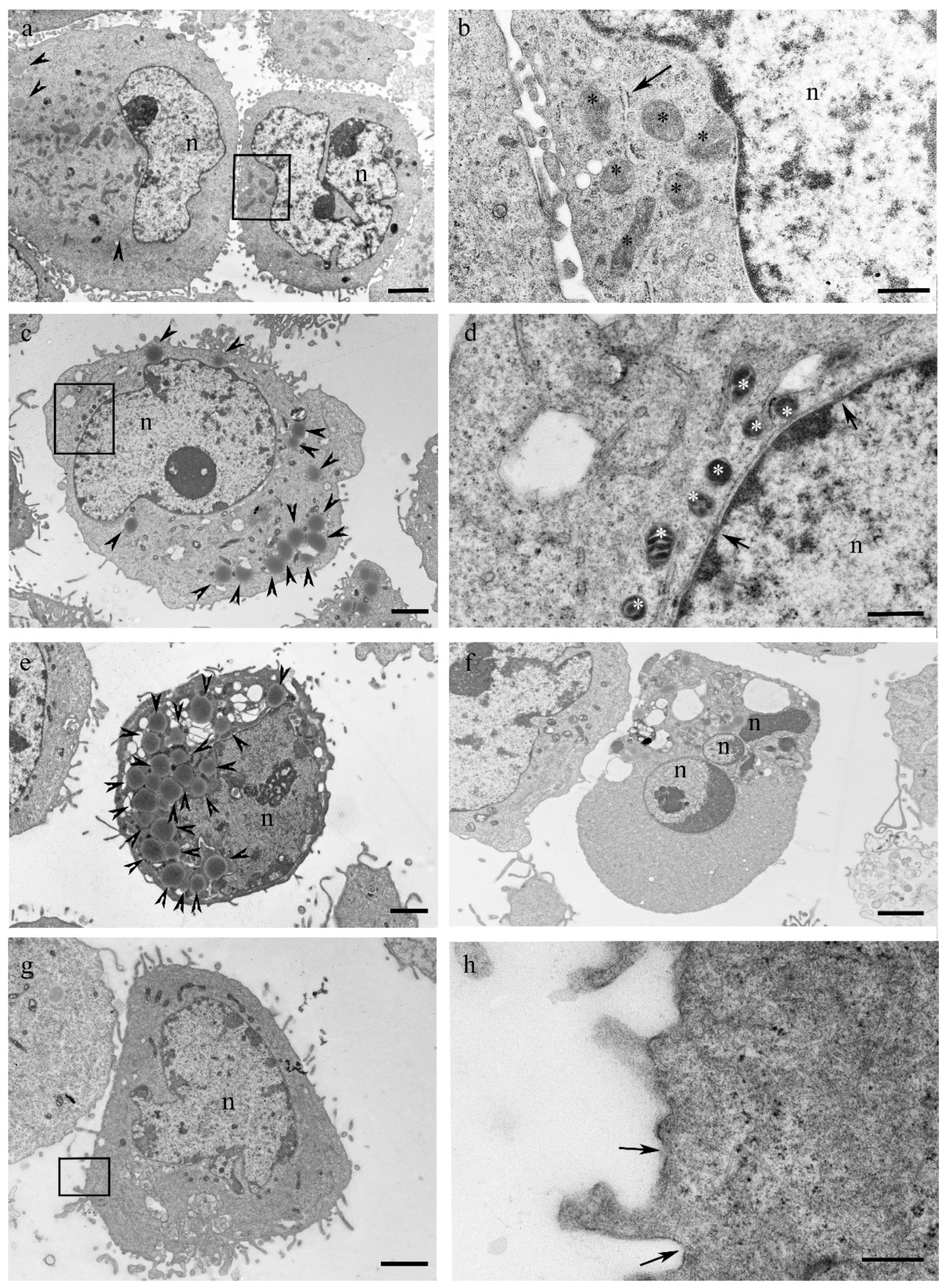
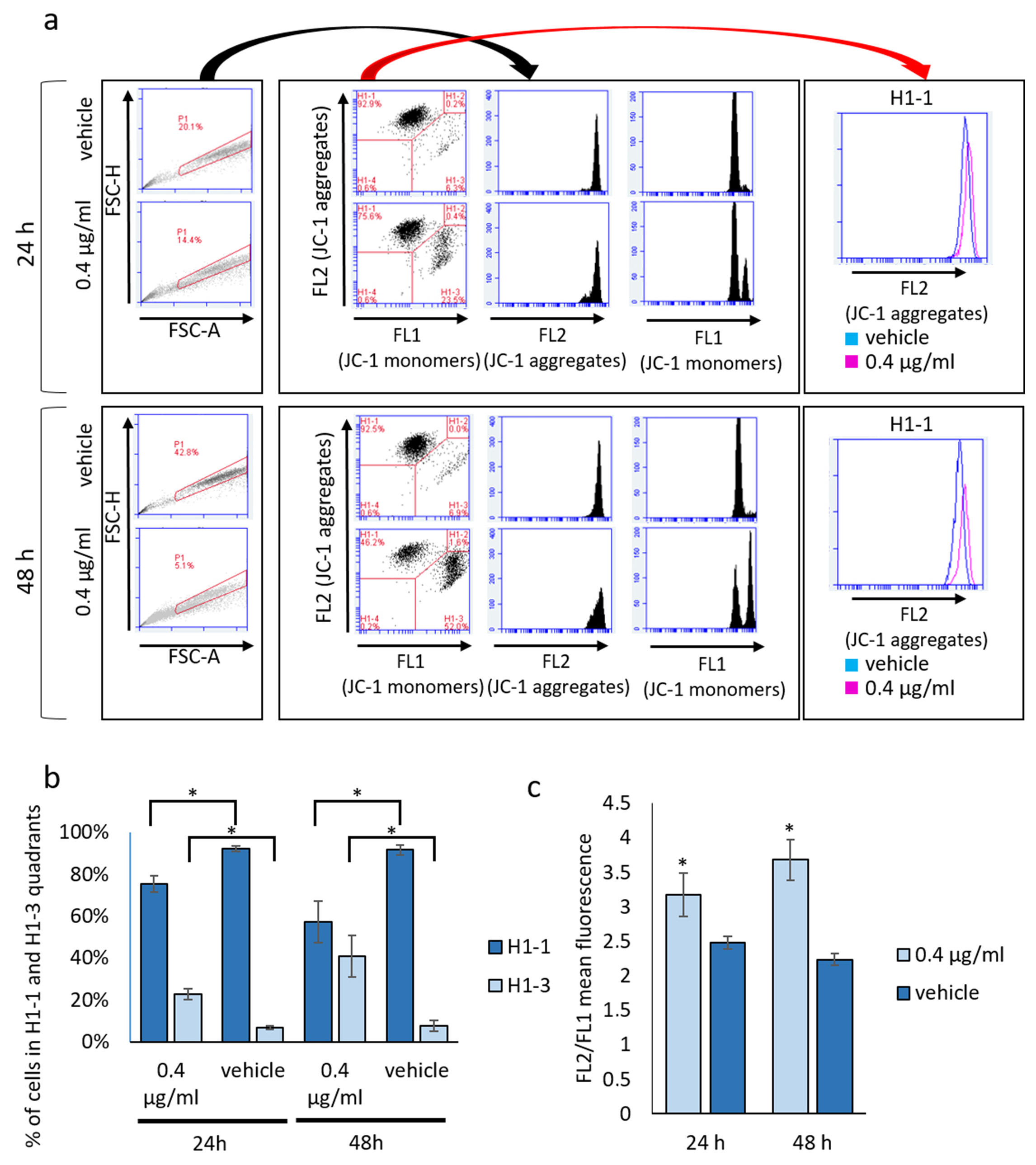
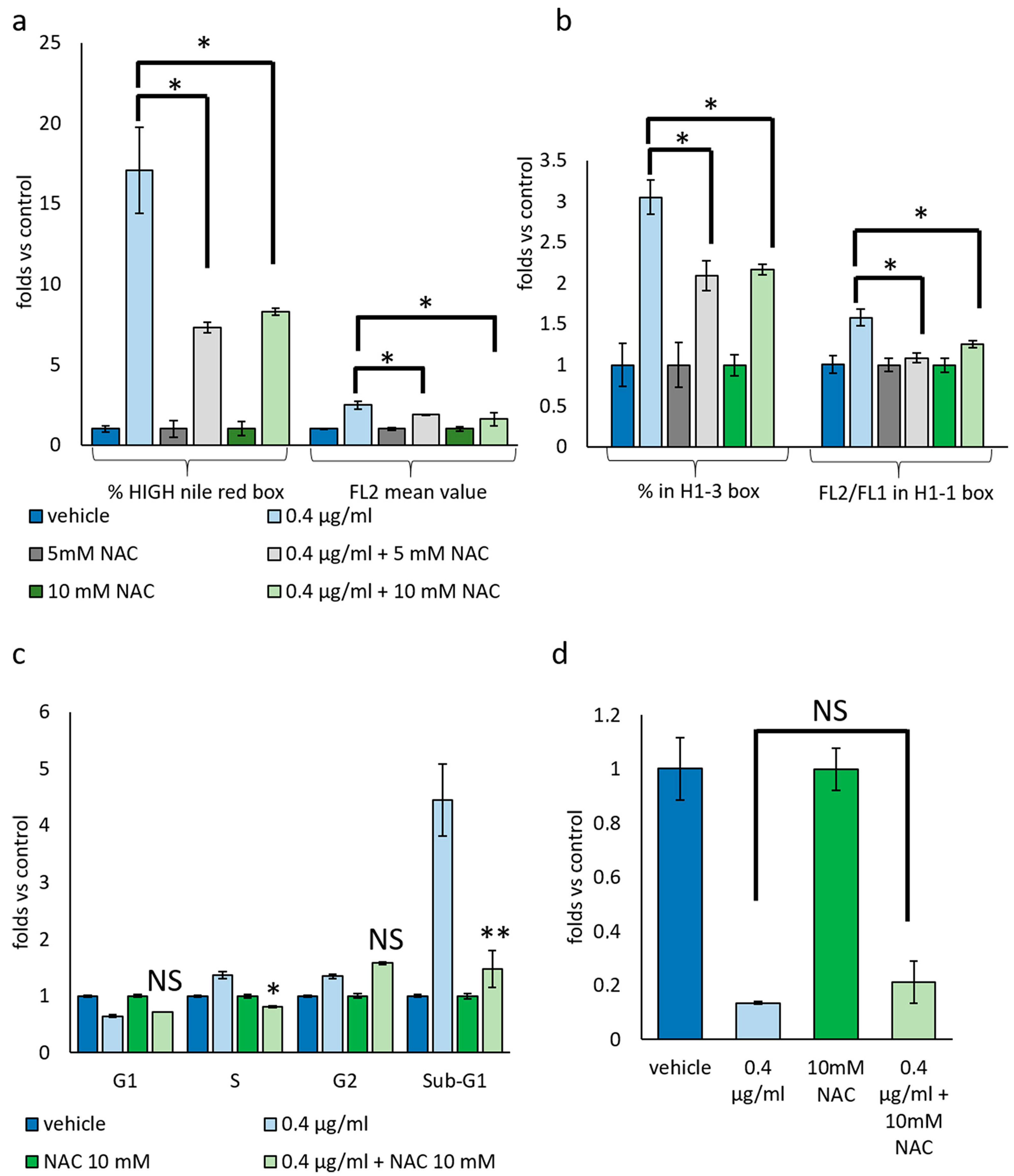
3.3. Mucus Treatment Induces Mixed Cell Death Modalities, Accumulation of Lipid Droplets and Mitochondrial Membrane Reorganization
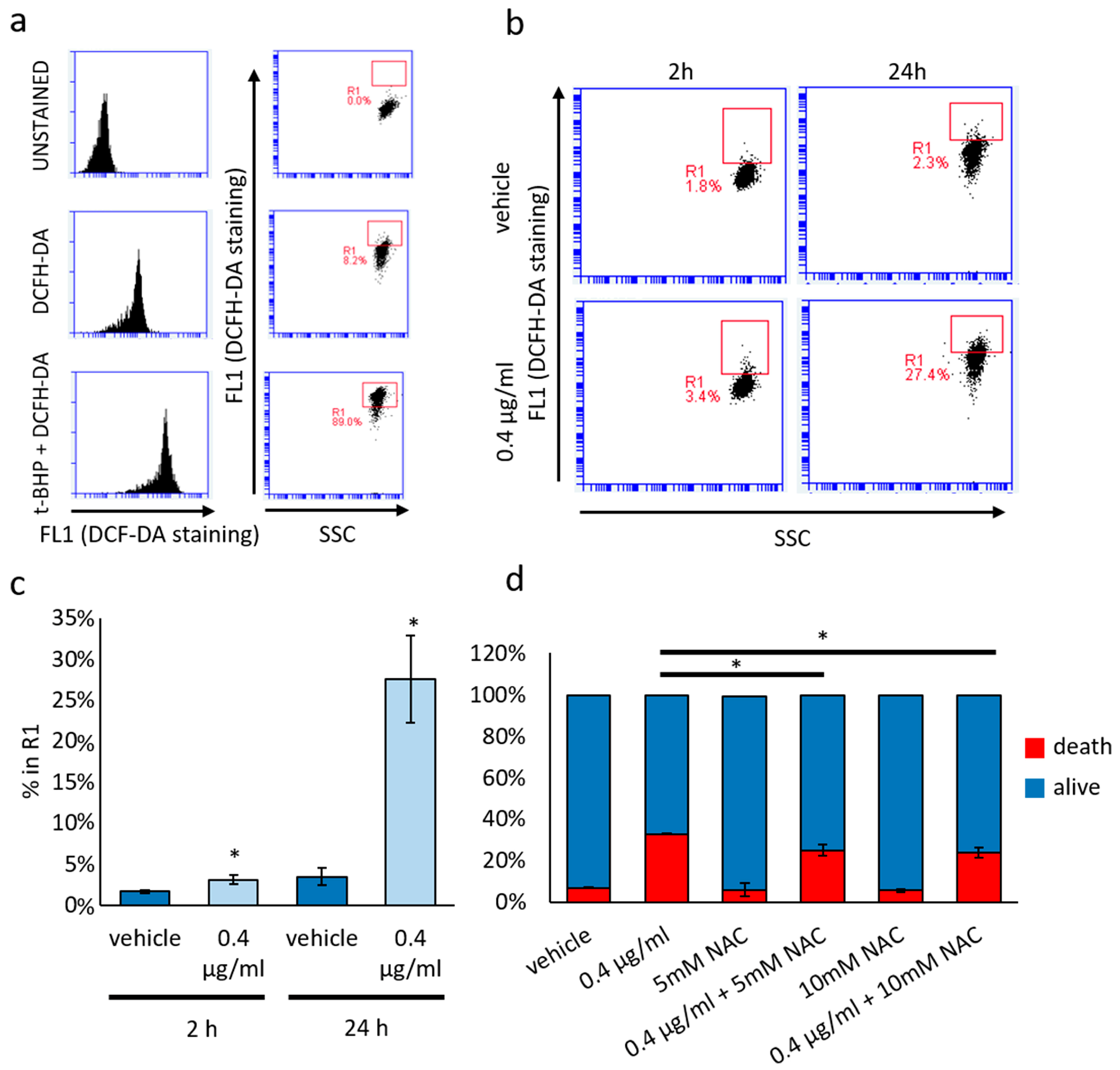
3.4. Cytotoxic and Not Cytostatic Effect Produced by Mucus Is Mediated by Reactive Oxygen Species
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Corey, E.J. Editorial review for comprehensive natural products chemistry. In Comprehensive Natural Products Chemistry; Barton, D.H.R., Nakanishi, K., Meth-Cohn, O., Eds.; Elsevier: Amsterdam, NY, USA, 1999. [Google Scholar]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; International Natural Product Sciences Taskforce; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Feher, M.; Schmidt, J.M. Property distributions: Differences between drugs, natural products, and molecules from combinatorial chemistry. J. Chem. Inf. Comput. Sci. 2003, 43, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Ma, Q.; Ye, L.; Piao, G. The Traditional Medicine and Modern Medicine from Natural Products. Molecules 2016, 29, 559. [Google Scholar] [CrossRef]
- Cechinel, F.V. Natural Products as Source of Molecules with Therapeutic Potential: Research & Development, Challenges and Perspectives; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Cragg, G.M.; Pezzuto, J.M. Natural Products as a Vital Source for the Discovery of Cancer Chemotherapeutic and Chemopreventive Agents. Med. Princ. Pract. 2016, 25, 41–59. [Google Scholar] [CrossRef]
- Chaachouay, N.; Zidane, L. Plant-Derived Natural Products: A Source for Drug Discovery and Development. Drugs Drug Candidates 2024, 3, 184–207. [Google Scholar] [CrossRef]
- Nasim, N.; Sandeep, I.S.; Mohanty, S. Plant-derived natural products for drug discovery: Current approaches and prospects. Nucleus 2022, 65, 399–411. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2023, 22, 275–325. [Google Scholar] [CrossRef]
- Rossi, L.; Salvetti, A. Planarian stem cell niche, the challenge for understanding tissue regeneration. Semin. Cell Dev. Biol. 2019, 87, 30–36. [Google Scholar] [CrossRef]
- Alessandra, S.; Rossi, L. Planarian Stem Cell Heterogeneity. Adv. Exp. Med. Biol. 2019, 1123, 39–54. [Google Scholar] [CrossRef]
- Ireland, D.; Bochenek, V.; Chaiken, D.; Rabeler, C.; Onoe, S.; Soni, A.; Collins, E.S. Dugesia japonica is the best suited of three planarian species for high-throughput toxicology screening. Chemosphere 2020, 253, 126718. [Google Scholar] [CrossRef]
- Hagstrom, D.; Cochet-Escartin, O.; Zhang, S.; Khuu, C.; Collins, E.M. Freshwater Planarians as an Alternative Animal Model for Neurotoxicology. Toxicol. Sci. 2015, 147, 270–285. [Google Scholar] [CrossRef] [PubMed]
- Hagstrom, D.; Cochet-Escartin, O.; Collins, E.M. Planarian brain regeneration as a model system for developmental neurotoxicology. Regeneration 2016, 3, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Suleiman, S.; Di Fiore, R.; Cassar, A.; Formosa, M.M.; Calleja-Agius, J.; Schembri-Wismayer, P. Anticancer effects of an extract from a local planarian species on human acute myeloid leukemia HL-60 cells in vitro. Biomed. Pharmacother. 2020, 130, 110549. [Google Scholar] [CrossRef]
- Pedersen, K.J. Slime-secreting cells of planarians. Ann. N. Y. Acad. Sci. 1963, 106, 424–443. [Google Scholar] [CrossRef]
- Hayes, M.J. Sulphated glycosaminoglycans support an assortment of planarian rhabdite structures. Biol. Open 2017, 6, 571–581. [Google Scholar] [CrossRef]
- Gao, L.; Li, A.; Li, N.; Liu, X.; Deng, H.; Zhao, B.; Pang, Q. Innate and intrinsic immunity in planarians. Invertebr. Surviv. J. 2017, 4, 443–452. [Google Scholar]
- Martin, G.G. A New Function of Rhabdites: Mucus Production for Ciliary Gliding. Zoomorphologie 1978, 91, 235–248. [Google Scholar] [CrossRef]
- Wilden, B.; Majdi, N.; Kuhlicke, U.; Neu, T.R.; Traunspurger, W. Flatworm mucus as the base of a food web. BMC Ecol. 2019, 19, 15. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, K.J. Some features of the fine structure and histochemistry of planarian subepidermal gland cells. Z. Für Zellforsch. Und Mikrosk. Anat. 1953, 50, 121–142. [Google Scholar] [CrossRef]
- Bowen, I.D.; Ryder, T.A.; Winters, C. The distribution of oxidizable mucosubstances and polysaccharides in the planarian Polycelis tenuis Iijima. Cell Tissue Res. 1975, 161, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Bocchinfuso, D.G.; Taylor, P.; Ross, E.; Ignatchenko, A.; Ignatchenko, V.; Kislinger, T.; Pearson, B.J.; Moran, M.F. Proteomic profiling of the planarian Schmidtea mediterranea and its mucous reveals similarities with human secretions and those predicted for parasitic flatworms. Mol. Cell. Proteom. 2012, 11, 681–691. [Google Scholar] [CrossRef] [PubMed]
- E-Kobon, T.; Thongararm, P.; Roytrakul, S.; Meesuk, L.; Chumnanpuen, P. Prediction of anticancer peptides against MCF-7 breast cancer cells from the peptidomes of Achatina fulica mucus fractions. Comput. Struct. Biotechnol. J. 2015, 14, 49–57. [Google Scholar] [CrossRef]
- Stabili, L.; Schirosi, R.; Parisi, M.G.; Piraino, S.; Cammarata, M. The Mucus of Actinia equina (Anthozoa, Cnidaria): An Unexplored Resource for Potential Applicative Purposes. Mar. Drugs 2015, 13, 5276–5296. [Google Scholar] [CrossRef]
- Balestrini, L.; Di Donfrancesco, A.; Rossi, L.; Marracci, S.; Isolani, M.E.; Bianucci, A.M.; Batistoni, R. The natural compound sanguinarine perturbs the regenerative capabilities of planarians. Int. J. Dev. Biol. 2017, 61, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Masuko, T.; Minami, A.; Iwasaki, N.; Majima, T.; Nishimura, S.; Lee, Y.C. Carbohydrate analysis by a phenol-sulfuric acid method in microplate format. Anal. Biochem. 2005, 339, 69–72. [Google Scholar] [CrossRef]
- Smiley, S.T.; Reers, M.; Mottola-Hartshorn, C.; Lin, M.; Chen, A.; Smith, T.W.; Steele, G.D., Jr.; Chen, L.B. Intracellular heterogeneity in mitochondrial membrane potentials revealed by a J-aggregate-forming lipophilic cation JC-1. Proc. Natl. Acad. Sci. USA 1991, 88, 3671–3675. [Google Scholar] [CrossRef]
- Pietkiewicz, S.; Schmidt, J.H.; Lavrik, I.N. Quantification of apoptosis and necroptosis at the single cell level by a combination of Imaging Flow Cytometry with classical Annexin V/propidium iodide staining. J. Immunol. Methods 2015, 423, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Miao, E.A.; Rajan, J.V.; Aderem, A. Caspase-1-induced pyroptotic cell death. Immunol. Rev. 2011, 243, 206–214. [Google Scholar] [CrossRef]
- Demuynck, R.; Efimova, I.; Naessens, F.; Krysko, D.V. Immunogenic ferroptosis and where to find it? J. Immunother. Cancer 2021, 9, e003430. [Google Scholar] [CrossRef]
- Miyake, S.; Murai, S.; Kakuta, S.; Uchiyama, Y.; Nakano, H. Identification of the hallmarks of necroptosis and ferroptosis by transmission electron microscopy. Biochem. Biophys. Res. Commun. 2020, 527, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Beatty, A.; Singh, T.; Tyurina, Y.Y.; Tyurin, V.A.; Samovich, S.; Nicolas, E.; Maslar, K.; Zhou, Y.; Cai, K.Q.; Tan, Y.; et al. Ferroptotic cell death triggered by conjugated linolenic acids is mediated by ACSL1. Nat. Commun. 2021, 12, 2244. [Google Scholar] [CrossRef]
- Zou, Y.; Palte, M.J.; Deik, A.A.; Li, H.; Eaton, J.K.; Wang, W.; Tseng, Y.Y.; Deasy, R.; Kost-Alimova, M.; Dančík, V.; et al. A GPX4-dependent cancer cell state underlies the clear-cell morphology and confers sensitivity to ferroptosis. Nat. Commun. 2019, 10, 1617. [Google Scholar] [CrossRef] [PubMed]
- Danielli, M.; Perne, L.; Jarc Jovičić, E.; Petan, T. Lipid droplets and polyunsaturated fatty acid trafficking: Balancing life and death. Front. Cell Dev. Biol. 2023, 11, 1104725. [Google Scholar] [CrossRef]
- Oropesa, M.; de la Mata, M.; Maraver, J.G.; Cordero, M.D.; Cotán, D.; Rodríguez-Hernández, A.; Domínguez-Moñino, I.; de Miguel, M.; Navas, P.; Sánchez-Alcázar, J.A. Apoptotic microtubule network organization and maintenance depend on high cellular ATP levels and energized mitochondria. Apoptosis 2011, 16, 404–424. [Google Scholar] [CrossRef]
- Kim, J.M.; Bae, H.R.; Park, B.S.; Lee, J.M.; Ahn, H.B.; Rho, J.H.; Yoo, K.W.; Park, W.C.; Rho, S.H.; Yoon, H.S.; et al. Early mitochondrial hyperpolarization and intracellular alkalinization in lactacystin-induced apoptosis of retinal pigment epithelial cells. J. Pharmacol. Exp. Ther. 2003, 305, 474–481. [Google Scholar] [CrossRef]
- Li, P.F.; Dietz, R.; von Harsdorf, R. p53 regulates mitochondrial membrane potential through reactive oxygen species and induces cytochrome c-independent apoptosis blocked by Bcl-2. EMBO J. 1999, 18, 6027–6036. [Google Scholar] [CrossRef]
- Marzo, I.; Brenner, C.; Zamzami, N.; Jürgensmeier, J.M.; Susin, S.A.; Vieira, H.L.; Prévost, M.C.; Xie, Z.; Matsuyama, S.; Reed, J.C.; et al. Bax and adenine nucleotide translocator cooperate in the mitochondrial control of apoptosis. Science 1998, 281, 2027–2031. [Google Scholar] [CrossRef]
- Rowe, L.A.; Degtyareva, N.; Doetsch, P.W. DNA damage-induced reactive oxygen species (ROS) stress response in Saccharomyces cerevisiae. Free Radic. Biol. Med. 2008, 45, 1167–1177. [Google Scholar] [CrossRef]
- Lagunin, A.; Filimonov, D.; Poroikov, V. Multi-targeted natural products evaluation based on biological activity prediction with PASS. Curr. Pharm. Des. 2010, 16, 1703–1717. [Google Scholar] [CrossRef]
- Muhammad, N.; Usmani, D.; Tarique, M.; Naz, H.; Ashraf, M.; Raliya, R.; Tabrez, S.; Zughaibi, T.A.; Alsaieedi, A.; Hakeem, I.J.; et al. The Role of Natural Products and Their Multitargeted Approach to Treat Solid Cancer. Cells 2022, 11, 2209. [Google Scholar] [CrossRef] [PubMed]
- Chinnadurai, R.K.; Khan, N.; Meghwanshi, G.K.; Ponne, S.; Althobiti, M.; Kumar, R. Current research status of anti-cancer peptides: Mechanism of action, production, and clinical applications. Biomed. Pharmacother. 2023, 164, 114996. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, D.; Veiga, A.S.; Castanho, M.A. From antimicrobial to anticancer peptides. A review. Front. Microbiol. 2013, 4, 294. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.V.; Yedery, R.D.; Aranha, C. Antimicrobial peptides: Premises and promises. Int. J. Antimicrob. Agents 2004, 24, 536–547. [Google Scholar] [CrossRef]
- Matusiewicz, M.; Kosieradzka, I.; Niemiec, T.; Grodzik, M.; Antushevich, H.; Strojny, B.; Gołębiewska, M. In Vitro Influence of Extracts from Snail Helix aspersa Müller on the Colon Cancer Cell Line Caco-2. Int. J. Mol. Sci. 2018, 19, 1064. [Google Scholar] [CrossRef]
- Harris, F.; Dennison, S.R.; Singh, J.; Phoenix, D.A. On the selectivity and efficacy of defense peptides with respect to cancer cells. Med. Res. Rev. 2013, 33, 190–234. [Google Scholar] [CrossRef]
- Shyu, P., Jr.; Wong, X.F.A.; Crasta, K.; Thibault, G. Dropping in on lipid droplets: Insights into cellular stress and cancer. Biosci. Rep. 2018, 38, BSR20180764. [Google Scholar] [CrossRef]
- Lee, H.; Horbath, A.; Kondiparthi, L.; Meena, J.K.; Lei, G.; Dasgupta, S.; Liu, X.; Zhuang, L.; Koppula, P.; Li, M.; et al. Cell cycle arrest induces lipid droplet formation and confers ferroptosis resistance. Nat. Commun. 2024, 15, 79. [Google Scholar] [CrossRef]
- Piel, J. Metabolites from symbiotic bacteria. Nat. Prod. Rep. 2009, 26, 338–362. [Google Scholar] [CrossRef]
- König, G.M.; Kehraus, S.; Seibert, S.F.; Abdel-Lateff, A.; Müller, D. Natural products from marine organisms and their associated microbes. ChemBioChem 2006, 7, 229–238. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gambino, G.; Da Pozzo, E.; Salvetti, A.; Rossi, L. Planarian Mucus: A Novel Source of Pleiotropic Cytotoxic and Cytostatic Agents against Cancer Cells. Biomolecules 2024, 14, 1075. https://doi.org/10.3390/biom14091075
Gambino G, Da Pozzo E, Salvetti A, Rossi L. Planarian Mucus: A Novel Source of Pleiotropic Cytotoxic and Cytostatic Agents against Cancer Cells. Biomolecules. 2024; 14(9):1075. https://doi.org/10.3390/biom14091075
Chicago/Turabian StyleGambino, Gaetana, Eleonora Da Pozzo, Alessandra Salvetti, and Leonardo Rossi. 2024. "Planarian Mucus: A Novel Source of Pleiotropic Cytotoxic and Cytostatic Agents against Cancer Cells" Biomolecules 14, no. 9: 1075. https://doi.org/10.3390/biom14091075