
Genome Mining for Diazo-Synthesis-Related Genes in Streptomyces sp. CS057 Unveiled the Cryptic Biosynthetic Gene Cluster crx for the Novel 3,4-AHBA-Derived Compound Crexazone 2
 ,
,  ,
,  , , , ,
, , , ,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions, Plasmids, and DNA Manipulations
2.2. Generation of Mutants
2.2.1. Generation of ΔAHBA Mutant
2.2.2. Generation of Mutants by Gene Replacement Using Gibson Assembly Plasmids
- pHZ1358ΔNRPS was used to delete orf4, generating the ΔNRPS mutant. It was constructed by Gibson assembly between pHZ1358 digested with NheI and XbaI; fragment A (2095 bp), containing the 5′-end of orf4, orf5, and orf6 and amplified with oligonucleotides AdNRPS FW and RV; fragment B (2554 bp), containing the 3′-end of orf4, and orf1 to orf3, amplified with oligonucleotides BdNRPS FW and RV; and the apramycin cassette, amplified from pUO9090 with oligonucleotides ApraGibsFW and RV.
- pHZ1358Δ11 was used to delete crxA1, generating the ΔcrxA1 mutant. Fragment A (2337 bp), containing orf10 and the 5′-end of crxA1, was amplified with oligonucleotides d11AFW and RV; fragment B (2627 bp), containing the 3′-end of crxA1, crxA2, crxB, and the 3′-end of crxC, was amplified with oligonucleotides d11BFW and RV.
- pHZ1358Δ14 was used to delete crxC, generating the ΔcrxC mutant. Fragment A (2181 bp), containing the 5′-end of crxC and crxA3, was amplified with oligonucleotides d14AFW and RV; fragment B (3662 bp), containing the 3′-end of crxC and crxA1 to crxB, was amplified with oligonucleotides d14BFW and RV.
- pHZ1358Δ16 was used to delete crxD, generating the ΔcrxD mutant. Fragment A (2512 bp), containing crxE, was amplified with oligonucleotides d16AFW and RV; fragment B (2655 bp), containing the 3′-end of crxD, crxA3, and the 5′-end of crxC, was amplified with oligonucleotides d16BFW and RV.
- pHZ1358Δ17 was used to delete crxE, generating the ΔcrxE mutant. Fragment A (1993 bp), containing the 5′-end of crxE, crxK, and crxI, was amplified with oligonucleotides d17AFW and RV; fragment B (2343 bp), containing the 5′-end of crxA3, crxD, and the 3′-end of crxE, was amplified with oligonucleotides d17BFW and RV.
- pHZ1358Δ22 was used to delete crxM, generating the ΔcrxM mutant. Fragment A (3035 bp), containing crxR1, crxF, and the 5′-end of crxM, was amplified with oligonucleotides d22AFW and RV; fragment B (2907 bp), conatining the 3′-end of crxM, crxN, and the 3′-end of crxH, was amplified with oligonucleotides d22BFW and RV.
- pHZ1358ΔTetR was used to delete crxR2, generating the ΔTetR mutant. Fragment A (2600 bp), containing the 5′-end of crxR1, crxT, crxP, and the 5′-end of crxR2, was amplified with oligonucleotides dTetRAFW and RV; fragment B (2700 bp), containing the 3′-end of crxR2, orf28 and the 3′-end of orf29, was amplified with oligonucleotides dTetRBFW and RV.
2.3. Generation of Recombinant Strains
- pSETEcHLuxR24 was constructed to overexpress crxR1, encoding a LuxR regulator under the control of the ermE*p promoter. CrxR1 (2539 bp) was amplified with the oligonucleotides eLuxR24XbaIFW and eLuxR24EcoRVRV and cloned into the XbaI and EcoRV sites of pSETEcH.
- pSETEcHLuxRA was constructed to overexpress the LuxR regulator encoded by orfA under the control of the ermE*p promoter. orfA (1049 bp), amplified with the oligonucleotides eLuxRAEcoRVFW and eLuxRAEcoRVRV, and cloned into the EcoRV site of pSETEcH.
- pSETEcHLuxR24A was constructed to overexpress both LuxR regulators encoded by orfA and crxR1 under the control of the ermE*p promoter orfA (1049 bp), amplified with the oligonucleotides orfApSETHGibsFWXbaI and orfApSETHGibsRVcorf24 and crxR1 (2539 bp), and amplified with oligonucleotides orf24FW and orf24gibsRVEcoRV. Both fragments were then cloned by Gibson assembly into pSETEcH digested with XbaI and EcoRV.
- pSETxkLuxR24 was constructed to overexpress crxR1 under the control of the kasOp* promoter. CrxR1 (2612 bp) was amplified with oligonucleotides FW24c3pSETxkBamHI and RV24c5pSETxkEcoRV and cloned by Gibson assembly into pSETxk digested with BamHI and EcoRV.
- pOJ260pLuxR24 was constructed to insert the ermE*p promoter upstream crxR1. CrxR1 (2539 bp) was amplified with oligonucleotides eLuxR24XbaIFW and eLuxR24EcoRVRV and cloned into the XbaI and EcoRV sites of pOJ260p.
- pSETxKTetR was constructed to overexpress the TetR regulator encoded by crxR2 under the control of the kasOp* promoter. CrxR2 (770 bp), amplified with the oligonucleotides kTetRcpSETxkBamHIFW and kTetRcpSETxkEcoRVRV, and cloned by Gibson assembly into pSETxk, digested with BamHI and EcoRV.
- pOJ260pNRPS was constructed to insert the ermE*p promoter upstream of the NRPS-coding gene orf4. This plasmid was generated by cloning a 5′-end fragment of orf4 amplified by PCR with the oligonucleotides pNRPSXbaIFW and pNRPSEcoRIRV into the XbaI and EcoRI sites of pOJ260p.
- pSETEcHc11LuxR24 was constructed to complement the ΔcrxA1 mutant. CrxA1 (1200 bp) was amplified with the oligonucleotides cORF11Lux24c5Lux24EcoRVFW and cORF11Lux24c5pSETHEcoRVRV and cloned by Gibson assembly into pSETEcHLuxR24 digested with EcoRV downstream of crxR1.
2.4. Chromatografic Analysis, Isolation and Structural Determination of Crexazones
2.4.1. Ultraperformance Liquid Chromatography (UPLC)
2.4.2. Isolation of Crexazones and Compound Dereplication Based on LC-DAD-HRMS Analyses
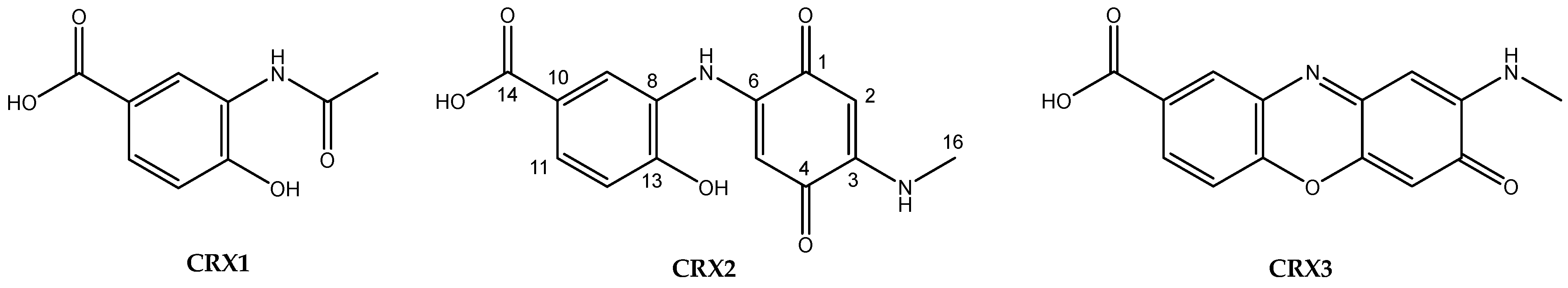
2.4.3. Structural Elucidation of Crexazone 2 (CRX2)
2.5. In Vitro Bioactivity Tests
3. Results
3.1. Genome Mining of Streptomyces Strains for N-N Bond Formation Coding Genes
3.2. Analysis of the NRPS Cluster in Region 28 from Streptomyces CS057
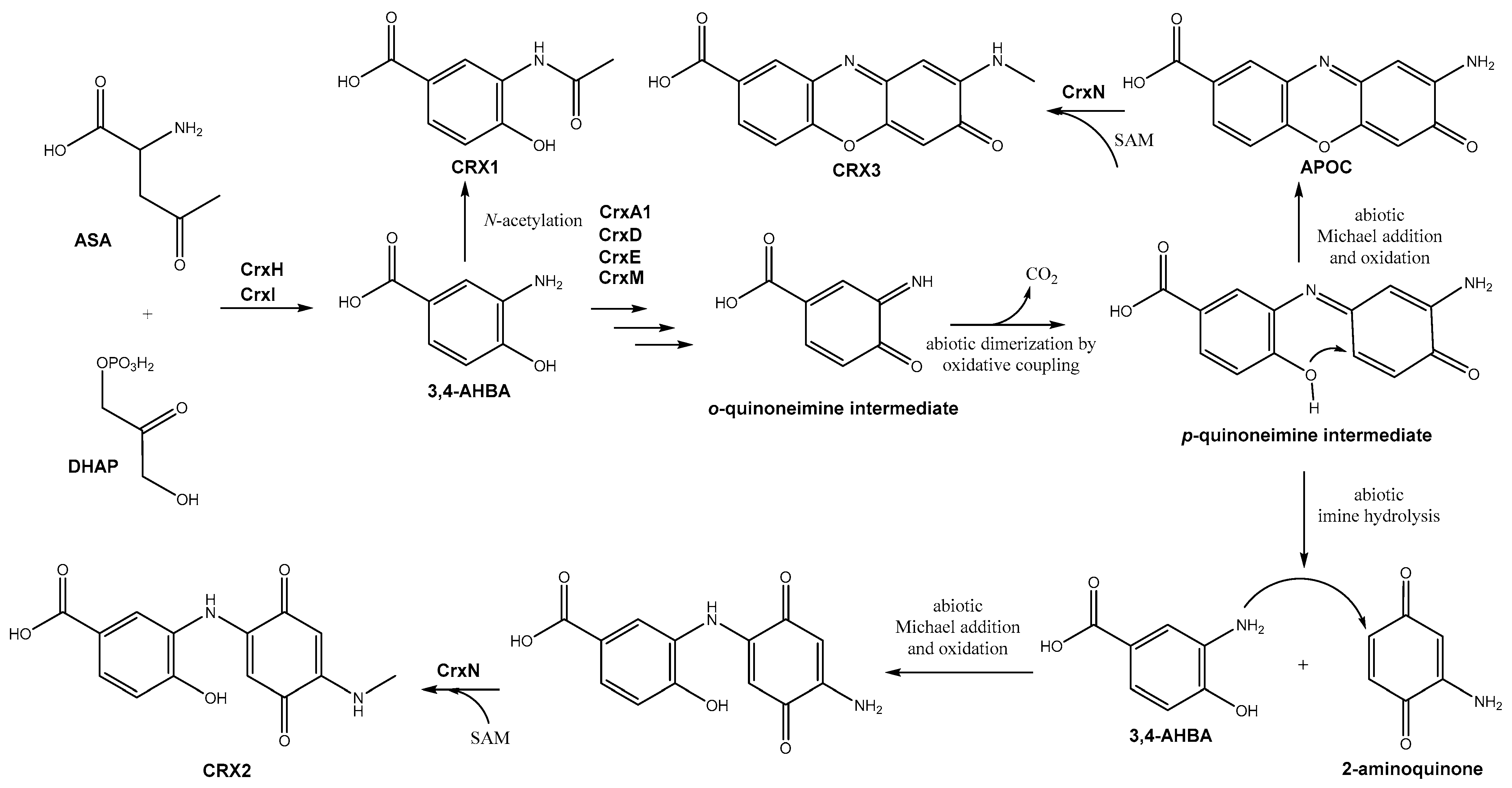
3.3. The crx Biosynthetic Gene Cluster and Its Encoded Compounds
- (i)
- Inactivation/overexpression of crxR2. Region 28 contains the TetR-like regulatory gene crxR2 (orf27) (Figure 1). TetR regulators usually act as repressors, although some can act as activators [53,54]. To evaluate its putative role in region 28, crxR2 was deleted and replaced by the apramycin resistance gene using plasmid pHZ1358ΔTetR (Table 1). The resultant mutant ΔTetR was genetically confirmed by PCR: using oligonucleotides dTetRIFW/ApraI and dTetRDRV/ApraD (Table S1), a 2600 bp fragment A and a 2700 bp fragment B were amplified from the mutant and not from the WT strain (Figure S5). Additionally, crxR2 was also overexpressed in the WT strain using pSETxkTetR (Table 1). The metabolite profiles of the resultant recombinant strain WT-kTetR and of the ΔTetR mutant were compared to those of the WT (Figure S6). No differences were observed, indicating that the TetR regulator CrxR2 is not involved in the regulation of region 28 under the tested conditions.
- (ii)
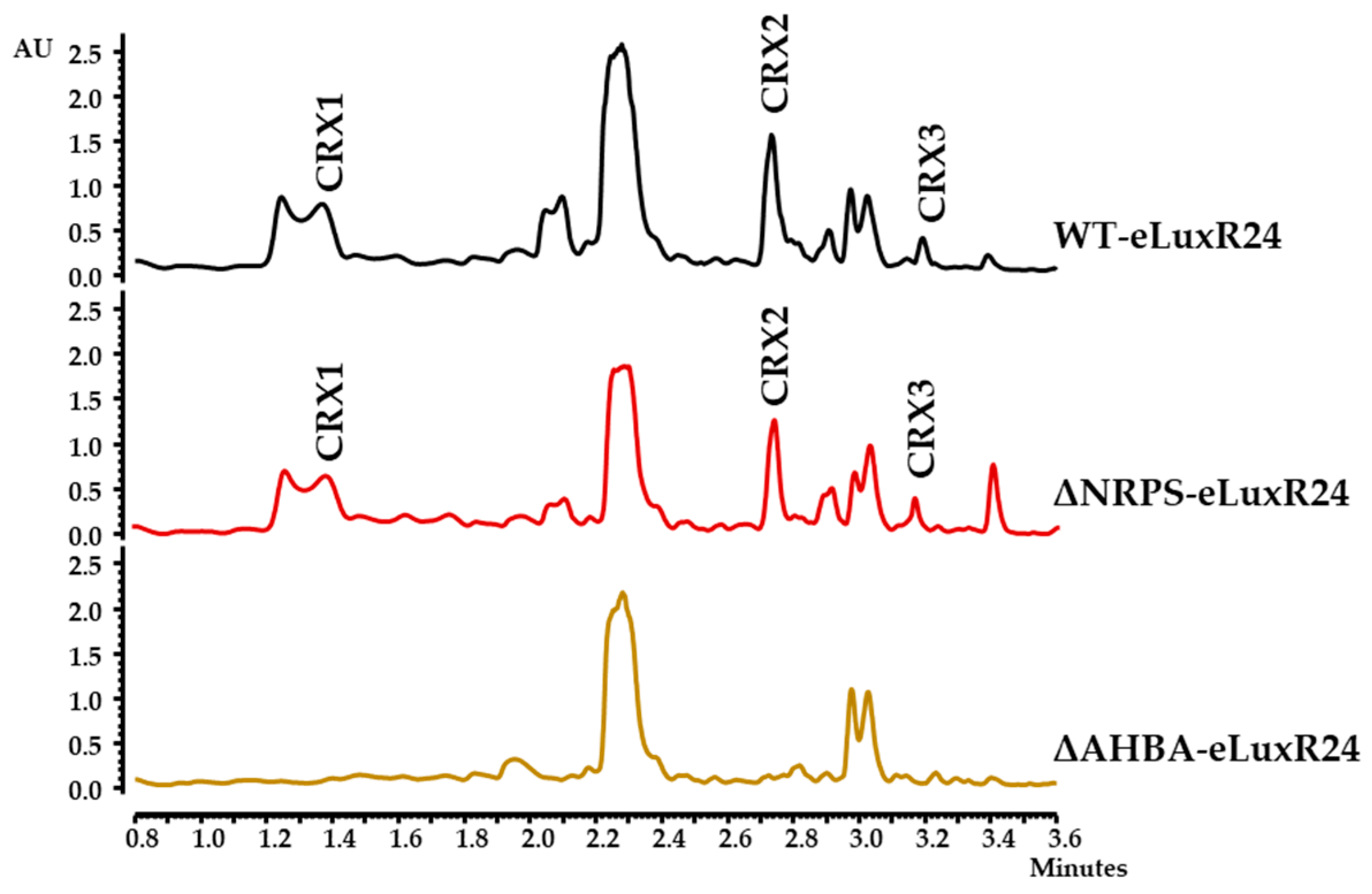
- Overexpression of orfA and/or crxR1. Region 28 contains two orfs encoding LuxR-type activators, orfA located at the “NRPS region” and crxR1 (orf24) at the “AHBA region” (Figure 1a). These two genes were individually overexpressed in the WT strain. The resultant recombinant strains WT-eLuxRA and WT-eLuxR24 (Table 1) were cultivated in R5A medium for 6 days, and their metabolite profiles were compared with those of the WT strain. Expression of orfA did not lead to the production of additional compounds (Figure 2). However, upon expressing crxR1, a series of differential peaks were detected (Figure 2). Compounds in these peaks were named crexazones (CRX) (see below). Co-expression of orfA and crxR1 (strain WT-eLuxR24A; Table 1) did not result in the production of additional peaks compared to those produced by the WT-eLuxR24 strain (Figure 2). These results indicated that the production of those differential peaks only requires the expression of crxR1. To evaluate a potential improvement in the production of those compounds, the ermE*p promoter was inserted upstream of crxR1 in the chromosome of WT using plasmid pOJ260pLuxR24 (Table 1). The right insertion of this promoter in the resultant recombinant strain WT-pLuxR24 was confirmed by PCR using oligonucleotides M13FW and eLuxR24DRV (Table S1), which amplified a 3400 bp DNA fragment from the recombinant strain and not from the WT strain (Figure S7). Moreover, crxR1 was overexpressed in trans under the control of the kasOp* promoter, using plasmid pSETxkLuxR24 (Table 1), generating the WT-kLuxR24 strain. Culture extracts from both strains did not show any additional differential peaks. However, some of the already detected compounds were produced in higher amounts: compounds CRX1 and CRX2 in WT-kLuxR24 and CRX1 in WT-pLuxR24 (Figure 2).
3.4. Determination of the Chemical Structure of Crexazones
3.5. Generation of Mutants in Genes Related to N-N Bond Formation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Klenk, H.-P.; Clément, C.; Ouhdouch, Y.; Van Wezel, G.P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Wink, J.; Mohammadipanah, F.; Hamedi, J. (Eds.) Biology and Biotechnology of Actinobacteria; Springer International Publishing: Cham, Switzerland, 2017; ISBN 978-3-319-60338-4. [Google Scholar]
- Donald, L.; Pipite, A.; Subramani, R.; Owen, J.; Keyzers, R.A.; Taufa, T. Streptomyces: Still the biggest producer of new natural secondary metabolites, a current perspective. Microbiol. Res. 2022, 13, 418–465. [Google Scholar] [CrossRef]
- Quinn, G.A.; Banat, A.M.; Abdelhameed, A.M.; Banat, I.M. Streptomyces from traditional medicine: Sources of new innovations in antibiotic discovery. J. Med. Microbiol. 2020, 69, 1040–1048. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug. Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Olano, C.; Méndez, C.; Salas, J.A. Molecular insights on the biosynthesis of antitumour compounds by Actinomycetes. Microb. Biotechnol. 2011, 4, 144–164. [Google Scholar] [CrossRef] [PubMed]
- Jagannathan, S.V.; Manemann, E.M.; Rowe, S.E.; Callender, M.C.; Soto, W. Marine Actinomycetes, new sources of biotechnological products. Mar. Drugs 2021, 19, 365. [Google Scholar] [CrossRef]
- Sivalingam, P.; Hong, K.; Pote, J.; Prabakar, K. Extreme environment Streptomyces: Potential sources for new antibacterial and anticancer drug leads? Int. J. Microbiol. 2019, 2019, 5283948. [Google Scholar] [CrossRef]
- Seipke, R.F.; Kaltenpoth, M.; Hutchings, M.I. Streptomyces as symbionts: An emerging and widespread theme? FEMS Microbiol. Rev. 2012, 36, 862–876. [Google Scholar] [CrossRef]
- Katsuyama, Y.; Matsuda, K. Recent advance in the biosynthesis of nitrogen–nitrogen bond–containing natural products. Curr. Opin. Chem. Biol. 2020, 59, 62–68. [Google Scholar] [CrossRef]
- Chen, L.; Deng, Z.; Zhao, C. Nitrogen–nitrogen bond formation reactions involved in Natural Product biosynthesis. ACS Chem. Biol. 2021, 16, 559–570. [Google Scholar] [CrossRef]
- García-Gutiérrez, C.; Pérez-Victoria, I.; Montero, I.; Fernández-De La Hoz, J.; Malmierca, M.G.; Martín, J.; Salas, J.A.; Olano, C.; Reyes, F.; Méndez, C. Unearthing a cryptic biosynthetic gene cluster for the piperazic acid-bearing depsipeptide Diperamycin in the ant-dweller Streptomyces sp. CS113. Int. J. Mol. Sci. 2024, 25, 2347. [Google Scholar] [CrossRef] [PubMed]
- He, H.-Y.; Niikura, H.; Du, Y.-L.; Ryan, K.S. Synthetic and biosynthetic routes to nitrogen–nitrogen bonds. Chem. Soc. Rev. 2022, 51, 2991–3046. [Google Scholar] [CrossRef] [PubMed]
- Sugai, Y.; Katsuyama, Y.; Ohnishi, Y. A Nitrous acid biosynthetic pathway for diazo group formation in Bacteria. Nat. Chem. Biol. 2016, 12, 73–75. [Google Scholar] [CrossRef]
- Waldman, A.J.; Balskus, E.P. Discovery of a diazo-forming enzyme in Cremeomycin biosynthesis. J. Org. Chem. 2018, 83, 7539–7546. [Google Scholar] [CrossRef]
- Kawai, S.; Yamada, A.; Du, D.; Sugai, Y.; Katsuyama, Y.; Ohnishi, Y. Identification and analysis of the biosynthetic gene cluster for the hydrazide-containing aryl polyene Spinamycin. ACS Chem. Biol. 2023, 18, 1821–1828. [Google Scholar] [CrossRef]
- Kawai, S.; Sugaya, Y.; Hagihara, R.; Tomita, H.; Katsuyama, Y.; Ohnishi, Y. Complete biosynthetic pathway of Alazopeptin, a tripeptide consisting of two molecules of 6-diazo-5-oxo-L-norleucine and one molecule of alanine. Angew. Chem. Int. Ed. 2021, 60, 10319–10325. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.-L.; Candra, H.; Pang, L.M.; Xiong, J.; Ding, Y.; Tran, H.T.; Low, Z.J.; Ye, H.; Liu, M.; Zheng, J.; et al. Biosynthesis of Tasikamides via pathway coupling and diazonium-mediated hydrazone formation. J. Am. Chem. Soc. 2022, 144, 1622–1633. [Google Scholar] [CrossRef]
- Kawai, S.; Hagihara, R.; Shin-ya, K.; Katsuyama, Y.; Ohnishi, Y. Bacterial Avenalumic acid biosynthesis includes substitution of an aromatic amino group for hydride by nitrous acid dependent diazotization. Angew. Chem. Int. Ed. 2022, 61, e202211728. [Google Scholar] [CrossRef]
- Del Rio Flores, A.; Twigg, F.F.; Du, Y.; Cai, W.; Aguirre, D.Q.; Sato, M.; Dror, M.J.; Narayanamoorthy, M.; Geng, J.; Zill, N.A.; et al. Biosynthesis of Triacsin featuring an N-hydroxytriazene pharmacophore. Nat. Chem. Biol. 2021, 17, 1305–1313. [Google Scholar] [CrossRef]
- Blodgett, J.A.V.; Oh, D.-C.; Cao, S.; Currie, C.R.; Kolter, R.; Clardy, J. Common biosynthetic origins for polycyclic tetramate macrolactams from phylogenetically diverse bacteria. Proc. Natl. Acad. Sci. USA 2010, 107, 11692–11697. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.J.; Oh, D.-C.; Yuceer, M.C.; Klepzig, K.D.; Clardy, J.; Currie, C.R. Bacterial protection of beetle-fungus mutualism. Science 2008, 322, 63. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.; Munnoch, J.T.; Devine, R.; Holmes, N.A.; Seipke, R.F.; Wilkinson, K.A.; Wilkinson, B.; Hutchings, M.I. Formicamycins, antibacterial polyketides produced by Streptomyces formicae isolated from african Tetraponera plant-ants. Chem. Sci. 2017, 8, 3218–3227. [Google Scholar] [CrossRef] [PubMed]
- Malmierca, M.G.; González-Montes, L.; Pérez-Victoria, I.; Sialer, C.; Braña, A.F.; García Salcedo, R.; Martín, J.; Reyes, F.; Méndez, C.; Olano, C.; et al. Searching for glycosylated natural products in Actinomycetes and identification of novel macrolactams and angucyclines. Front. Microbiol. 2018, 9, 39. [Google Scholar] [CrossRef]
- Malmierca, M.G.; Pérez-Victoria, I.; Martín, J.; Reyes, F.; Méndez, C.; Salas, J.A.; Olano, C. New Sipanmycin analogues generated by combinatorial biosynthesis and mutasynthesis approaches relying on the substrate flexibility of key enzymes in the biosynthetic pathway. Appl. Environ. Microbiol. 2020, 86, e02453-19. [Google Scholar] [CrossRef]
- Ceniceros, A.; Cuervo, L.; Méndez, C.; Salas, J.A.; Olano, C.; Malmierca, M.G. A multidisciplinary approach to Unraveling the natural product biosynthetic potential of a Streptomyces strain collection isolated from leaf-cutting ants. Microorganisms 2021, 9, 2225. [Google Scholar] [CrossRef]
- Cuervo, L.; Álvarez-García, S.; Salas, J.A.; Méndez, C.; Olano, C.; Malmierca, M.G. The volatile organic compounds of Streptomyces spp.: An in-depth analysis of their antifungal properties. Microorganisms 2023, 11, 1820. [Google Scholar] [CrossRef]
- Prado-Alonso, L.; Pérez-Victoria, I.; Malmierca, M.G.; Montero, I.; Rioja-Blanco, E.; Martín, J.; Reyes, F.; Méndez, C.; Salas, J.A.; Olano, C. Colibrimycins, novel halogenated hybrid polyketide synthase-nonribosomal peptide synthetase (PKS-NRPS) compounds produced by Streptomyces Sp. strain CS147. Appl. Environ. Microbiol. 2022, 88, e0183921. [Google Scholar] [CrossRef]
- Kieser, T.; Bibb, M.J.; Buttner, M.J.; Chater, K.F.; Hopwood, D.A. Practical Streptomyces Genetics; Kieser, T., Ed.; Innes: Norwich, UK, 2000; ISBN 978-0-7084-0623-6. [Google Scholar]
- Sánchez, L.; Braña, A.F. Cell density influences antibiotic biosynthesis in Streptomyces clavuligerus. Microbiology 1996, 142, 1209–1220. [Google Scholar] [CrossRef]
- Hobbs, G.; Frazer, C.M.; Gardner, D.C.J.; Cullum, J.A.; Oliver, S.G. Dispersed growth of Streptomyces in liquid culture. Appl. Microbiol. Biotechnol. 1989, 31, 272–277. [Google Scholar] [CrossRef]
- Fernández, E.; Weißbach, U.; Reillo, C.S.; Braña, A.F.; Méndez, C.; Rohr, J.; Salas, J.A. Identification of two genes from Streptomyces argillaceus encoding glycosyltransferases involved in transfer of a disaccharide during biosynthesis of the antitumor drug Mithramycin. J. Bacteriol. 1998, 180, 4929–4937. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D.W. (Eds.) Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Peláez, A.I.; Ribas-Aparicio, R.M.; Gómez, A.; Rodicio, M.R. Structural and functional characterization of the recR gene of Streptomyces. Mol. Gen. Genom. 2001, 265, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; He, X.; Liang, J.; Zhou, X.; Deng, Z. Analysis of Functions in Plasmid pHZ1358 Influencing its genetic and structural stability in Streptomyces lividans 1326. Appl. Microbiol. Biotechnol. 2009, 82, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Olano, C.; Wilkinson, B.; Sanchez, C.; Moss, S.; Sheridan, R.; Math, V.; Weston, A.; Brana, A.; Martin, C.; Oliynyk, M. Biosynthesis of the angiogenesis inhibitor Borrelidin by Streptomyces parvulus Tü4055. Cluster analysis and assignment of functions. Chem. Biol. 2004, 11, 87–97. [Google Scholar]
- Cuervo, L.; Malmierca, M.G.; García-Salcedo, R.; Méndez, C.; Salas, J.A.; Olano, C.; Ceniceros, A. Co-expression of transcriptional regulators and housekeeping genes in Streptomyces spp.: A strategy to optimize metabolite production. Microorganisms 2023, 11, 1585. [Google Scholar] [CrossRef]
- Altschul, S. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Augustijn, H.E.; Reitz, Z.L.; Biermann, F.; Alanjary, M.; Fetter, A.; Terlouw, B.R.; Metcalf, W.W.; Helfrich, E.J.N.; et al. antiSMASH 7.0: New and improved predictions for detection, regulation, chemical structures and visualisation. Nucleic. Acids. Res. 2023, 51, W46–W50. [Google Scholar] [CrossRef]
- Pérez-Victoria, I.; Martín, J.; Reyes, F. Combined LC/UV/MS and NMR Strategies for the Dereplication of Marine Natural Products. Planta Med. 2016, 82, 857–871. [Google Scholar] [CrossRef]
- Martín, J.; Crespo, G.; González-Menéndez, V.; Pérez-Moreno, G.; Sánchez-Carrasco, P.; Pérez-Victoria, I.; Ruiz-Pérez, L.M.; González-Pacanowska, D.; Vicente, F.; Genilloud, O.; et al. MDN-0104, an antiplasmodial betaine lipid from Heterospora chenopodii. J. Nat. Prod. 2014, 77, 2118–2123. [Google Scholar] [CrossRef]
- Dictionary of Natural Products on USB v. 32:2; CRC Press: Boca Raton, FL, USA, 2023.
- Waldman, A.J.; Pechersky, Y.; Wang, P.; Wang, J.X.; Balskus, E.P. The Cremeomycin biosynthetic gene cluster encodes a pathway for diazo Formation. ChemBioChem 2015, 16, 2172–2175. [Google Scholar] [CrossRef]
- Guo, Y.Y.; Li, Z.H.; Xia, T.Y.; Du, Y.L.; Mao, X.M.; Li, Y.-Q. Molecular mechanism of azoxy bond formation for Azoxymycins biosynthesis. Nat. Commun. 2019, 10, 4420. [Google Scholar] [CrossRef]
- Zheng, Z.; Xiong, J.; Bu, J.; Ren, D.; Lee, Y.; Yeh, Y.; Lin, C.; Parry, R.; Guo, Y.; Liu, H. Reconstitution of the final steps in the biosynthesis of Valanimycin reveals the origin of its characteristic azoxy moiety. Angew. Chem. Int. Ed. 2024, 63, e202315844. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.L.; Rohac, R.; Mitchell, A.J.; Boal, A.K.; Balskus, E.P. An N-nitrosating metalloenzyme constructs the pharmacophore of Streptozotocin. Nature 2019, 566, 94–99. [Google Scholar] [CrossRef]
- Matsuda, K.; Tomita, T.; Shin-ya, K.; Wakimoto, T.; Kuzuyama, T.; Nishiyama, M. Discovery of unprecedented hydrazine-forming machinery in bacteria. J. Am. Chem. Soc. 2018, 140, 9083–9086. [Google Scholar] [CrossRef] [PubMed]
- Aparicio, J.F.; Molnár, I.; Schwecke, T.; König, A.; Haydock, S.F.; Ee Khaw, L.; Staunton, J.; Leadlay, P.F. Organization of the biosynthetic gene cluster for Rapamycin in Streptomyces hygroscopicus: Analysis of the enzymatic domains in the modular polyketide synthase. Gene 1996, 169, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Stachelhaus, T.; Mootz, H.D.; Marahiel, M.A. The specificity-conferring code of adenylation domains in nonribosomal peptide synthetases. Chem. Biol. 1999, 6, 493–505. [Google Scholar] [CrossRef]
- Khayatt, B.I.; Overmars, L.; Siezen, R.J.; Francke, C. Classification of the adenylation and acyl-transferase activity of NRPS and PKS systems using ensembles of substrate specific hidden markov models. PLoS ONE 2013, 8, e62136. [Google Scholar] [CrossRef]
- Bloudoff, K.; Schmeing, T.M. Structural and functional aspects of the nonribosomal peptide synthetase condensation domain superfamily: Discovery, dissection and diversity. Biochim. Et Biophys. Acta (BBA)-Proteins Proteom. 2017, 1865, 1587–1604. [Google Scholar] [CrossRef]
- Uguru, G.C.; Stephens, K.E.; Stead, J.A.; Towle, J.E.; Baumberg, S.; McDowall, K.J. Transcriptional Activation of the pathway-specific regulator of the Actinorhodin biosynthetic genes in Streptomyces coelicolor. Mol. Microbiol. 2005, 58, 131–150. [Google Scholar] [CrossRef]
- Li, X.; Yu, T.; He, Q.; McDowall, K.J.; Jiang, B.; Jiang, Z.; Wu, L.; Li, G.; Li, Q.; Wang, S.; et al. Binding of a biosynthetic intermediate to AtrA modulates the production of Lindamycin by Streptomyces globisporus. Mol. Microbiol. 2015, 96, 1257–1271. [Google Scholar] [CrossRef]
- Suzuki, H.; Ohnishi, Y.; Horinouchi, S. GriC and GriD constitute a carboxylic acid reductase involved in Grixazone biosynthesis in Streptomyces griseus. J. Antibiot. 2007, 60, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.; Molloy, B.; Pérez-Victoria, I.; Montero, I.; Braña, A.F.; Olano, C.; Arca, S.; Martín, J.; Reyes, F.; Salas, J.A.; et al. Uncovering the cryptic gene cluster ahb for 3-amino-4-hydroxybenzoate derived ahbamycins, by searching SARP regulator encoding genes in the Streptomyces argillaceus genome. Int. J. Mol. Sci. 2023, 24, 8197. [Google Scholar] [CrossRef] [PubMed]
- Abdelfattah, M.S. A new bioactive aminophenoxazinone alkaloid from a marine-derived Actinomycete. Nat. Prod. Res. 2013, 27, 2126–2131. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Ohnishi, Y.; Furusho, Y.; Sakuda, S.; Horinouchi, S. Novel benzene ring biosynthesis from C(3) and C(4) primary metabolites by two enzymes. J. Biol. Chem. 2006, 281, 36944–36951. [Google Scholar] [CrossRef] [PubMed]
- Taber, W.A.; Nancy, N.G.; Harry, L.Y.J. Structure and synthesis of texazone, 2-(N-methylamino)-3H-phenoxazin-3-one-8-carboxylic acid, an Actinomycete metabolite. J. Antibiot. 1983, 36, 688–694. [Google Scholar]
- Katsuyama, Y.; Sato, Y.; Sugai, Y.; Higashiyama, Y.; Senda, M.; Senda, T.; Ohnishi, Y. Crystal structure of the nitrosuccinate lyase CreD in complex with fumarate provides insights into the catalytic mechanism for nitrous acid elimination. FEBS J. 2018, 285, 1540–1555. [Google Scholar] [CrossRef]
- Becerril, A.; Pérez-Victoria, I.; Martín, J.M.; Reyes, F.; Salas, J.A.; Méndez, C. Biosynthesis of Largimycins in Streptomyces argillaceus Involves Transient β-Alkylation and Cryptic Halogenation Steps Unprecedented in the Leinamycin Family. ACS Chem. Biol. 2022, 17, 2320–2331. [Google Scholar] [CrossRef]
- Adak, S.; Moore, B.S. Cryptic halogenation reactions in natural product biosynthesis. Nat. Prod. Rep. 2021, 38, 1760–1774. [Google Scholar] [CrossRef]
- Hagihara, R.; Katsuyama, Y.; Sugai, Y.; Onaka, H.; Ohnishi, Y. Novel desferrioxamine derivatives synthesized using the secondary metabolism-specific nitrous acid biosynthetic pathway in Streptomyces davawensis. J. Antibiot. 2018, 71, 911–919. [Google Scholar] [CrossRef]
- Menéndez, N.; Nur-e-Alam, M.; Fischer, C.; Braña, A.F.; Salas, J.A.; Rohr, J.; Méndez, C. Deoxysugar transfer during chromomycin A3 biosynthesis in Streptomyces griseus subsp. griseus: New derivatives with antitumor activity. Appl. Environ. Microbiol. 2006, 72, 167–177. [Google Scholar] [CrossRef]
- Suzuki, H.; Furusho, Y.; Higashi, T.; Ohnishi, Y.; Horinouchi, S. A Novel O-Aminophenol Oxidase Responsible for Formation of the Phenoxazinone Chromophore of Grixazone. J. Biol. Chem. 2006, 281, 824–833.66. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.; Mathur, P. p-Quinoneimine as an intermediate in the oxidative coupling of 2-amino-5-methylphenol to 4a,7-dimethyldihydro-2-aminophenoxazinone catalyzed by a monomeric copper(II) complex. Catal. Commun. 2014, 55, 1–5. [Google Scholar] [CrossRef]
- Corbett, J.F. Benzoquinone imines. Part II. Hydrolysis of p-benzoquinone monoimine and p-benzoquinone di-imine. J. Chem. Soc. B 1969, 213–216. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mutant Strain | Plasmid | Deleted Genes |
| ΔAHBA | pHZ1358ΔAHBA | orf14-23 |
| ΔNRPS | pHZ1358ΔNRPS | orf4 |
| ΔcrxA1 | pHZ1358Δ11 | crxA1 |
| ΔcrxC | pHZ1358Δ14 | crxC |
| ΔcrxD | pHZ1358Δ16 | crxD |
| ΔcrxE | pHZ1358Δ17 | crxE |
| ΔcrxM | pHZ1358Δ22 | crxM |
| ΔTetR | pHZ1358ΔTetR | crxR2 |
| Recombinant Strain | Plasmid | Expressed Genes |
| WT-eLuxR24 | pSETEcHLuxR24 | crxR1 |
| WT-eLuxRA | pSETEcHLuxRA | orfA |
| Wt-eLuxR24A | pSETEcHLuxR24A | orfA, crxR1 |
| WT-kLuxR24 | pSETxkLuxR24 | crxR1 |
| WT-pLuxR24 | pOJ260pLuxR24 | crxR1 |
| WT-kTetR | pSETxkTetR | crxR2 |
| WT-pNRPS | pOJ260pNRPS | orf4 |
| WT-pNRPS-eLuxR24 | pOJ260pNRPS | orf4 |
| pSETEcHLuxR24 | crxR1 | |
| ΔAHBA-eLuxR24 | pSETEcHLuxR24 | crxR1 |
| ΔNRPS-eLuxR24 | pSETEcHLuxR24 | crxR1 |
| ΔcrxA1-eLuxR24 | pSETEcHLuxR24 | crxR1 |
| ΔcrxC-eLuxR24 | pSETEcHLuxR24 | crxR1 |
| ΔcrxD-eLuxR24 | pSETEcHLuxR24 | crxR1 |
| ΔcrxE-eLuxR24 | pSETEcHLuxR24 | crxR1 |
| ΔcrxM-eLuxR24 | pSETEcHLuxR24 | crxR1 |
| ΔcrxA1c | pSETEcHc11LuxR24 | crxR1, crxA1 |
| Position | δC | δH (mult, J in Hz) |
|---|---|---|
| 1 | 177.1, C | |
| 2 | 92.2, CH | 5.31, s |
| 3 | 151.5, C | |
| 4 | 179.4, C | |
| 5 | 95.5, CH | 5.50, s |
| 6 | 147.1, C | |
| 7 | 8.83, s | |
| 8 | 125.0, C | |
| 9 | 124.0, CH | 7.78, d (1.9) |
| 10 | 121.8, C | |
| 11 | 128.3, CH | 7.69, dd (8.4, 1.9) |
| 12 | 115.7, CH | 7.03, d (8.4) |
| 13 | 154.3, C | |
| 14 | 166.7, C | |
| 15 | 7.95, q (5.3) | |
| 16 | 29.0, CH3 | 2.78, d (5.3) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prado-Alonso, L.; Ye, S.; Pérez-Victoria, I.; Montero, I.; Riesco, P.; Ortiz-López, F.J.; Martín, J.; Olano, C.; Reyes, F.; Méndez, C. Genome Mining for Diazo-Synthesis-Related Genes in Streptomyces sp. CS057 Unveiled the Cryptic Biosynthetic Gene Cluster crx for the Novel 3,4-AHBA-Derived Compound Crexazone 2. Biomolecules 2024, 14, 1084. https://doi.org/10.3390/biom14091084
Prado-Alonso L, Ye S, Pérez-Victoria I, Montero I, Riesco P, Ortiz-López FJ, Martín J, Olano C, Reyes F, Méndez C. Genome Mining for Diazo-Synthesis-Related Genes in Streptomyces sp. CS057 Unveiled the Cryptic Biosynthetic Gene Cluster crx for the Novel 3,4-AHBA-Derived Compound Crexazone 2. Biomolecules. 2024; 14(9):1084. https://doi.org/10.3390/biom14091084
Chicago/Turabian StylePrado-Alonso, Laura, Suhui Ye, Ignacio Pérez-Victoria, Ignacio Montero, Pedro Riesco, Francisco Javier Ortiz-López, Jesús Martín, Carlos Olano, Fernando Reyes, and Carmen Méndez. 2024. "Genome Mining for Diazo-Synthesis-Related Genes in Streptomyces sp. CS057 Unveiled the Cryptic Biosynthetic Gene Cluster crx for the Novel 3,4-AHBA-Derived Compound Crexazone 2" Biomolecules 14, no. 9: 1084. https://doi.org/10.3390/biom14091084