
A Perspective on the Enhancer Dependent Bacterial RNA Polymerase

{kind=link}
{kind=link}
Abstract
:1. Introduction
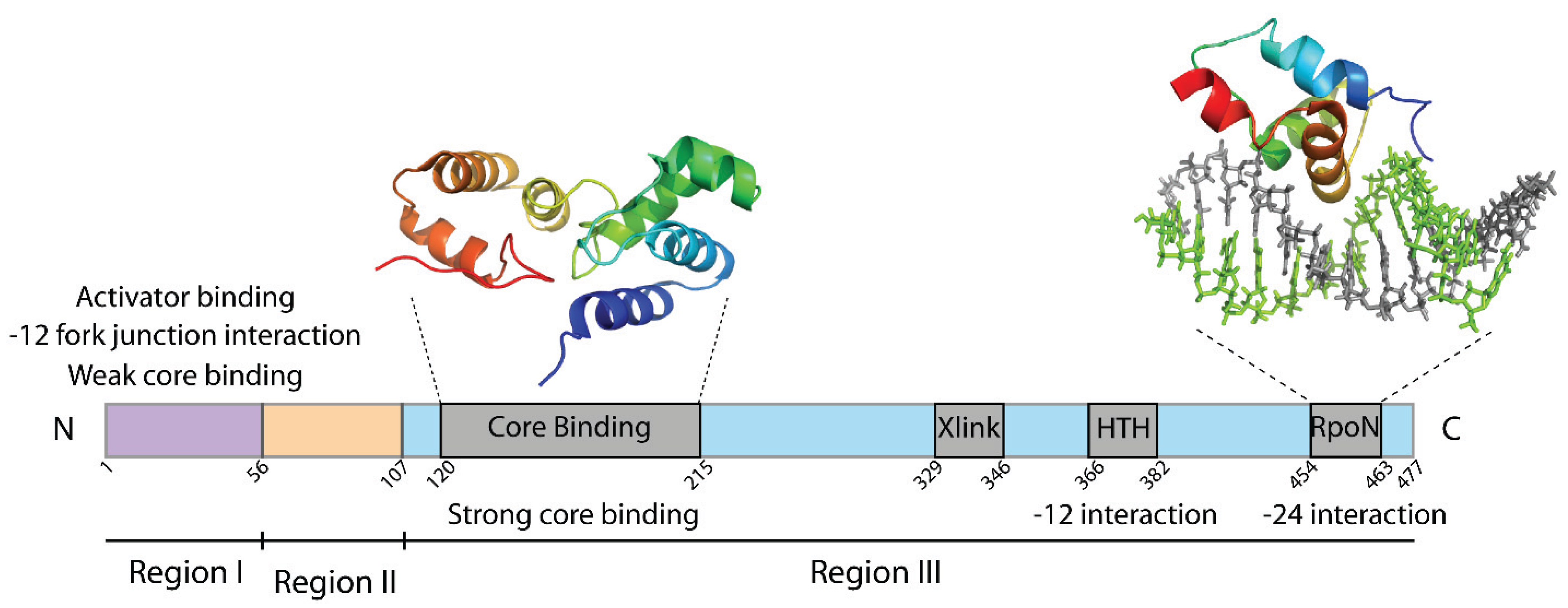
2. The Sigma54 Factor

3. Promoter Recognition
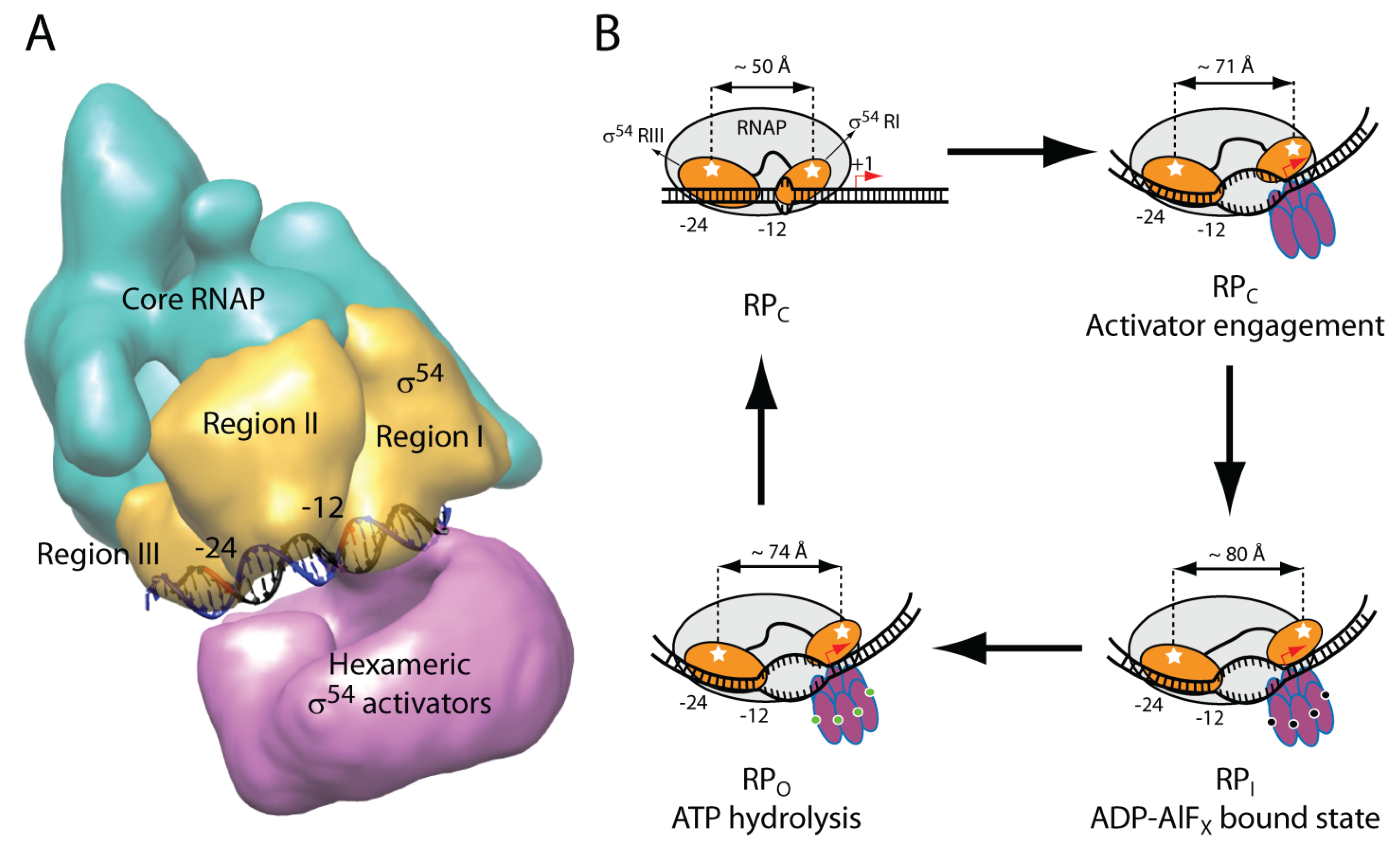
4. Activator Remodel of the Sigma54 Holoenzyme

5. Signaling for Sigma54 Dependent Transcription
6. Conclusions and a Final Perspective
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Merrick, M.J. In a class of its own—The RNA polymerase sigma factor sigma 54 (sigma N). Mol. Microbiol. 1993, 10, 903–909. [Google Scholar] [CrossRef] [PubMed]
- Bush, M.; Dixon, R. The role of bacterial enhancer binding proteins as specialized activators of sigma54-dependent transcription. Microbiol. Mol. Biol. Rev. 2012, 76, 497–529. [Google Scholar] [CrossRef] [PubMed]
- Joly, N.; Zhang, N.; Buck, M.; Zhang, X. Coupling AAA protein function to regulated gene expression. Biochim. Biophys. Acta 2012, 1823, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Francke, C.; Groot Kormelink, T.; Hagemeijer, Y.; Overmars, L.; Sluijter, V.; Moezelaar, R.; Siezen, R.J. Comparative analyses imply that the enigmatic sigma factor 54 is a central controller of the bacterial exterior. BMC Genomics 2011. [Google Scholar] [CrossRef]
- Dixon, R.; Kahn, D. Genetic regulation of biological nitrogen fixation. Nat. Rev. Microbiol. 2004, 2, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.A.; Grimm, D.; Henion, A.K.; Elias, A.F.; Stewart, P.E.; Rosa, P.A.; Gherardini, F.C. Borrelia burgdorferi sigma54 is required for mammalian infection and vector transmission but not for tick colonization. Proc. Natl. Acad. Sci. USA 2005, 102, 5162–5167. [Google Scholar] [CrossRef] [PubMed]
- Fouts, D.E.; Abramovitch, R.B.; Alfano, J.R.; Baldo, A.M.; Buell, C.R.; Cartinhour, S.; Chatterjee, A.K.; D’Ascenzo, M.; Gwinn, M.L.; Lazarowitz, S.G.; et al. Genomewide identification of Pseudomonas syringae pv. Tomato DC3000 promoters controlled by the hrpL alternative sigma factor. Proc. Natl. Acad. Sci. USA 2002, 99, 2275–2280. [Google Scholar] [CrossRef]
- Studholme, D.J.; Buck, M. The biology of enhancer-dependent transcriptional regulation in bacteria: Insights from genome sequences. FEMS Microbiol. Lett. 2000, 186, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, A.J.; Millikan, D.S.; Campbell, J.M.; Visick, K.L. Vibrio fischeri sigma54 controls motility, biofilm formation, luminescence, and colonization. Appl. Environ. Microbiol. 2004, 70, 2520–2524. [Google Scholar] [CrossRef] [PubMed]
- Kustu, S.; Santero, E.; Keener, J.; Popham, D.; Weiss, D. Expression of sigma 54 (NtrA)-dependent genes is probably united by a common mechanism. Microbiol. Rev. 1989, 53, 367–376. [Google Scholar] [PubMed]
- Popham, D.L.; Szeto, D.; Keener, J.; Kustu, S. Function of a bacterial activator protein that binds to transcriptional enhancers. Science 1989, 243, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Cannon, W.; Chaney, M.; Buck, M. Characterisation of holoenzyme lacking sigman regions I and II. Nucleic Acids Res. 1999, 27, 2478–2486. [Google Scholar] [CrossRef] [PubMed]
- Syed, A.; Gralla, J.D. Isolation and properties of enhancer-bypass mutants of sigma 54. Mol. Microbiol. 1997, 23, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Doucleff, M.; Malak, L.T.; Pelton, J.G.; Wemmer, D.E. The C-terminal RpoN domain of sigma54 forms an unpredicted helix-turn-helix motif similar to domains of sigma70. J. Biol. Chem. 2005, 280, 41530–41536. [Google Scholar] [CrossRef] [PubMed]
- Doucleff, M.; Pelton, J.G.; Lee, P.S.; Nixon, B.T.; Wemmer, D.E. Structural basis of DNA recognition by the alternative sigma-factor, sigma54. J. Mol. Biol. 2007, 369, 1070–1078. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Gralla, J.D. Roles for the C-terminal region of sigma 54 in transcriptional silencing and DNA binding. J. Biol. Chem. 2001, 276, 8979–8986. [Google Scholar] [CrossRef] [PubMed]
- Friedman, L.J.; Mumm, J.P.; Gelles, J. RNA polymerase approaches its promoter without long-range sliding along DNA. Proc. Natl. Acad. Sci. USA 2013, 110, 9740–9745. [Google Scholar] [CrossRef] [PubMed]
- Friedman, L.J.; Gelles, J. Mechanism of transcription initiation at an activator-dependent promoter defined by single-molecule observation. Cell 2012, 148, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Tintut, Y.; Wang, J.T.; Gralla, J.D. Abortive cycling and the release of polymerase for elongation at the sigma 54-dependent glnAP2 promoter. J. Biol. Chem. 1995, 270, 24392–24398. [Google Scholar] [CrossRef] [PubMed]
- Burrows, P.C.; Severinov, K.; Ishihama, A.; Buck, M.; Wigneshweraraj, S.R. Mapping sigma 54-RNA polymerase interactions at the −24 consensus promoter element. J. Biol. Chem. 2003, 278, 29728–29743. [Google Scholar] [CrossRef] [PubMed]
- Bose, D.; Pape, T.; Burrows, P.C.; Rappas, M.; Wigneshweraraj, S.R.; Buck, M.; Zhang, X. Organization of an activator-bound RNA polymerase holoenzyme. Mol. Cell 2008, 32, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Joly, N.; Buck, M. A common feature from different subunits of a homomeric AAA+ protein contacts three spatially distinct transcription elements. Nucleic Acids Res. 2012, 40, 9139–9152. [Google Scholar] [CrossRef] [PubMed]
- Sysoeva, T.A.; Chowdhury, S.; Guo, L.; Nixon, B.T. Nucleotide-induced asymmetry within ATPase activator ring drives sigma54-RNAP interaction and ATP hydrolysis. Genes Dev. 2013, 27, 2500–2511. [Google Scholar] [PubMed]
- Sharma, A.; Leach, R.N.; Gell, C.; Zhang, N.; Burrows, P.C.; Shepherd, D.A.; Wigneshweraraj, S.; Smith, D.A.; Zhang, X.; Buck, M.; et al. Domain movements of the enhancer-dependent sigma factor drive DNA delivery into the RNA polymerase active site: Insights from single molecule studies. Nucleic Acids Res. 2014, 42, 5177–5190. [Google Scholar] [CrossRef]
- Zhang, N.; Gordiyenko, Y.; Joly, N.; Lawton, E.; Robinson, C.V.; Buck, M. Subunit dynamics and nucleotide-dependent asymmetry of an AAA+ transcription complex. J. Mol. Biol. 2014, 426, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Burrows, P.C.; Joly, N.; Buck, M. A prehydrolysis state of an AAA+ ATPase supports transcription activation of an enhancer-dependent RNA polymerase. Proc. Natl. Acad. Sci. USA 2010, 107, 9376–9381. [Google Scholar] [CrossRef] [PubMed]
- Lawton, E.; Jovanovic, M.; Joly, N.; Waite, C.; Zhang, N.; Wang, B.; Burrows, P.; Buck, M. Determination of the self-association residues within a homomeric and a heteromeric AAA+ enhancer binding protein. J. Mol. Biol. 2014, 426, 1692–1710. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.R.; Barton, G.; Pan, Z.; Buck, M.; Wigneshweraraj, S. Nitrogen stress response and stringent response are coupled in Escherichia coli. Nat. Commun. 2014. [Google Scholar] [CrossRef]
- Zhao, K.; Liu, M.; Burgess, R.R. Promoter and regulon analysis of nitrogen assimilation factor, sigma54, reveal alternative strategy for E. coli MG1655 flagellar biosynthesis. Nucleic Acids Res. 2010, 38, 1273–1283. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, N.; Buck, M. A Perspective on the Enhancer Dependent Bacterial RNA Polymerase. Biomolecules 2015, 5, 1012-1019. https://doi.org/10.3390/biom5021012
Zhang N, Buck M. A Perspective on the Enhancer Dependent Bacterial RNA Polymerase. Biomolecules. 2015; 5(2):1012-1019. https://doi.org/10.3390/biom5021012
Chicago/Turabian StyleZhang, Nan, and Martin Buck. 2015. "A Perspective on the Enhancer Dependent Bacterial RNA Polymerase" Biomolecules 5, no. 2: 1012-1019. https://doi.org/10.3390/biom5021012
APA StyleZhang, N., & Buck, M. (2015). A Perspective on the Enhancer Dependent Bacterial RNA Polymerase. Biomolecules, 5(2), 1012-1019. https://doi.org/10.3390/biom5021012