
Innovative Biochemometric Approach to the Metabolite and Biological Profiling of the Balkan Thistle (Cirsium appendiculatum Griseb.), Asteraceae


 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
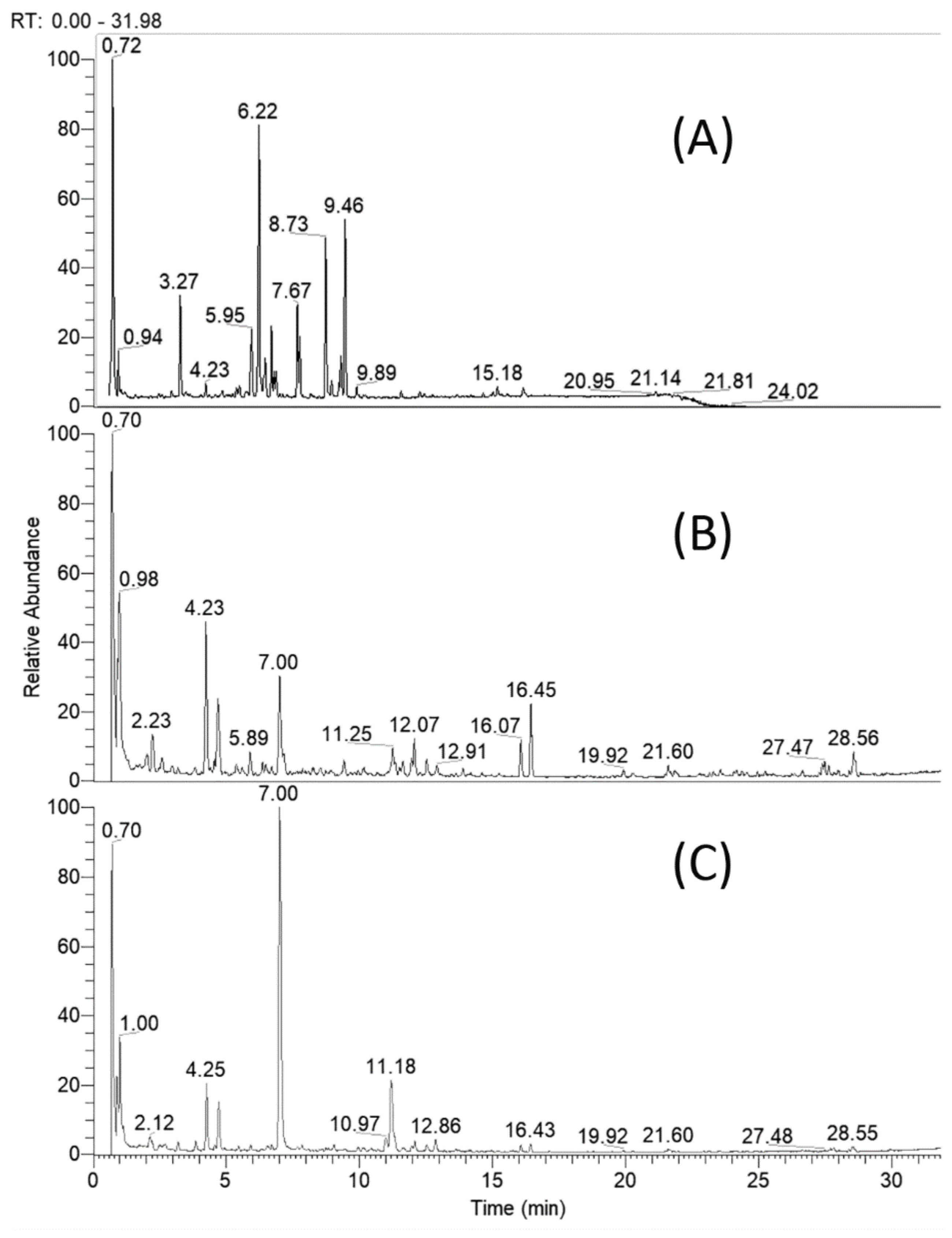
2.1. UHPLC–HRMS Profiling of Specialized Natural Products in Cirsium appendiculatum Extracts
2.1.1. Carboxylic (Including Hydroxybenzoic, Hydroxycinnamic and Acylquinic) Acids and Their Glycosides
2.1.2. Flavonoids, Flavones and Flavonols
2.1.3. Methoxylated Flavonoids
2.1.4. Free Fatty Acids
2.2. Total Content of Phenolics and Flavonoids
2.3. Antioxidant Properties
2.4. Enzyme Inhibitory Effects
2.5. Supervised Multivariate Analysis
2.6. Relationship between Chemical Compounds and Biological Activities
3. Materials and Methods
3.1. Plant Material
3.2. Sample Extraction
3.3. UHPLC–HRMS
3.4. Total Phenolics and Flavonoid Contents Determination
3.5. Biological Activities Determination
3.6. Data Processing
3.6.1. Univariate Analysis
3.6.2. Supervised Multivariate Analysis
3.6.3. Pearson Correlation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wolfender, J.-L.; Litaudon, M.; Touboul, D.; Queiroz, E.F. Innovative omics-based approaches for prioritisation and targeted isolation of natural products–new strategies for drug discovery. Nat. Prod. Rep. 2019, 36, 855–868. [Google Scholar] [CrossRef] [Green Version]
- Gevrenova, R.; Zheleva-Dimitrova, D.; Balabanova, V.; Voynikov, Y.; Sinan, K.I.; Mahomoodally, M.F.; Zengin, G. Integrated phytochemistry, bio-functional potential and multivariate analysis of Tanacetum macrophyllum (Waldst. & Kit.) Sch. Bip. and Telekia speciosa (Schreb.) Baumg.(Asteraceae). Ind. Crop. Prod. 2020, 155, 112817. [Google Scholar]
- Kadereit, J.W.; Jeffrey, C. Flowering Plants, Eudicots: Asterales. The Families and Genera of Vascular Plants, Volume VIII; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Yildiz, B.; Arabaci, T.; Dirmenci, T.; Köstekci, S. A taxonomic revision of the genus Cirsium Mill. sect. Cirsium (Asteraceae: Cardueae) in Turkey. Turk. J. Bot. 2016, 40, 514–530. [Google Scholar] [CrossRef]
- Kim, M.S.; Nam, M.; Hwang, G.S. Metabolic alterations in two Cirsium species identified at distinct phenological stages using UPLC-QTOF/MS. Phytochem. Anal. 2018, 29, 77–86. [Google Scholar] [CrossRef]
- Lee, S.; Lee, D.-H.; Kim, J.-C.; Um, B.H.; Sung, S.H.; Jeong, L.S.; Kim, Y.K.; Kim, S.-N. Pectolinarigenin, an aglycone of pectolinarin, has more potent inhibitory activities on melanogenesis than pectolinarin. Biochem. Biophys. Res. Commun. 2017, 493, 765–772. [Google Scholar] [CrossRef]
- Abbet, C.; Slacanin, I.; Corradi, E.; De Mieri, M.; Hamburger, M.; Potterat, O. Comprehensive analysis of Cirsium spinosissimum Scop., a wild alpine food plant. Food Chem. 2014, 160, 165–170. [Google Scholar] [CrossRef]
- Jordon-Thaden, I.E.; Louda, S.M. Chemistry of Cirsium and Carduus: A role in ecological risk assessment for biological control of weeds? Biochem. Syst. Ecol. 2003, 31, 1353–1396. [Google Scholar] [CrossRef] [Green Version]
- Nazaruk, J.; Chłędzik, S.; Strawa, J.; Bazydło, K.; Wajs-Bonikowska, A. Chemical Composition and Antioxidant Activity of Cirsium vulgare Inflorescences. Nat. Prod. Commun. 2017, 12, 1934578X1701200414. [Google Scholar] [CrossRef] [Green Version]
- Do, J.-C.; Jung, K.-Y.; Son, K.-H. Isolation of pectolinarin from the aerial parts of Cirsium nipponicum. Korean J. Pharmacogn. 1994, 25, 73–75. [Google Scholar]
- Dutta, C.; Ray, L.P.; Roy, D. Taraxasterol and its derivatives from Cirsium arvense. Phytochemistry 1972, 11, 2267–2269. [Google Scholar] [CrossRef]
- N. Stoyanov Stefanov, B.K. Flora Republicae Bulgaricae; Nauka i Izkustvo (Science and Art State Publishing House): Sofia, Bulgaria, 1967; Volume 2, pp. 1123–1128. [Google Scholar]
- Wolfender, J.L.; Queiroz, E.F.; Allard, P.M. Massive metabolite profiling of natural extracts for a rational prioritization of bioactive natural products: A paradigm shift in pharmacognosy. Food Front. 2020, 1, 105–106. [Google Scholar] [CrossRef] [Green Version]
- Termentzi, A.; Zervou, M.; Kokkalou, E. Isolation and structure elucidation of novel phenolic constituents from Sorbus domestica fruits. Food Chem. 2009, 116, 371–381. [Google Scholar] [CrossRef]
- Chahdoura, H.; João, C.; Barros, L.; Santos-Buelga, C.; Ferreira, I.C.; Achour, L. Phytochemical characterization and antioxidant activity of Opuntia microdasys (Lehm.) Pfeiff flowers in different stages of maturity. J. Funct. Foods 2014, 9, 27–37. [Google Scholar] [CrossRef]
- Ak, G.; Gevrenova, R.; Sinan, K.I.; Zengin, G.; Zheleva, D.; Mahomoodally, M.F.; Senkardes, I.; Brunetti, L.; Leone, S.; Di Simone, S.C. Tanacetum vulgare L.(Tansy) as an effective bioresource with promising pharmacological effects from natural arsenal. Food Chem. Toxicol. 2021, 153, 112268. [Google Scholar] [CrossRef]
- Gai, X.-H. Research progress on chemical constituents of Coptidis Rhizoma and its pharmacological activities. Chin. Tradit. Herb. Drugs 2018, 4919–4927. [Google Scholar]
- Münzenberger, B.; Heilemann, J.; Strack, D.; Kottke, I.; Oberwinkler, F. Phenolics of mycorrhizas and non-mycorrhizal roots of Norway spruce. Planta 1990, 182, 142–148. [Google Scholar] [CrossRef]
- Lee, S.R.; Clardy, J.; Senger, D.R.; Cao, S.; Kim, K.H. Iridoid and phenylethanoid glycosides from the aerial part of Barleria lupulina. Rev. Bras. Farmacogn. 2016, 26, 281–284. [Google Scholar] [CrossRef]
- Puppala, M.; Ponder, J.; Suryanarayana, P.; Reddy, G.B.; Petrash, J.M.; LaBarbera, D.V. The isolation and characterization of β-glucogallin as a novel aldose reductase inhibitor from Emblica officinalis. PLoS ONE 2012, 7, e31399. [Google Scholar] [CrossRef] [Green Version]
- Clifford, M.N.; Wu, W.; Kirkpatrick, J.; Kuhnert, N. Profiling the chlorogenic acids and other caffeic acid derivatives of herbal Chrysanthemum by LC− MS n. J. Agric. Food Chem. 2007, 55, 929–936. [Google Scholar] [CrossRef]
- Jaiswal, R.; Kiprotich, J.; Kuhnert, N. Determination of the hydroxycinnamate profile of 12 members of the Asteraceae family. Phytochemistry 2011, 72, 781–790. [Google Scholar] [CrossRef]
- Justesen, U. Collision-induced fragmentation of deprotonated methoxylated flavonoids, obtained by electrospray ionization mass spectrometry. J. Mass Spectrom. 2001, 36, 169–178. [Google Scholar] [CrossRef]
- Ren, D.; Ran, L.; Yang, C.; Xu, M.; Yi, L. Integrated strategy for identifying minor components in complex samples combining mass defect, diagnostic ions and neutral loss information based on ultra-performance liquid chromatography-high resolution mass spectrometry platform: Folium Artemisiae Argyi as a case study. J. Chromatogr. 2018, 1550, 35–44. [Google Scholar]
- Zhang, X.; Liang, C.; Li, C.; Bu, M.; Bu, L.; Xiao, Y.; Sun, H.; Zhang, L. Simultaneous qualitative and quantitative study of main compounds in Commelina communis linn. by UHPLC–Q-TOF-MS-MS and HPLC–ESI-MS-MS. J. Chromatogr. Sci. 2018, 56, 582–594. [Google Scholar] [CrossRef]
- De Souza, M.B., Jr.; Pinto, J.C.; Lima, E.L. Control of a chaotic polymerization reactor: A neural network based model predictive approach. Polym. Eng. Sci. 1996, 36, 448–457. [Google Scholar] [CrossRef]
- Rashmi, H.B.; Negi, P.S. Phenolic acids from vegetables: A review on processing stability and health benefits. Food Res. Int. 2020, 136, 109298. [Google Scholar] [CrossRef]
- Nazaruk, J. Antioxidant activity and total phenolic content in Cirsium five species from north–east region of Poland. Fitoterapia 2008, 79, 194–196. [Google Scholar] [CrossRef]
- Llorent-Martínez, E.J.; Zengin, G.; Sinan, K.I.; Polat, R.; Canlı, D.; Picot-Allain, M.C.N.; Mahomoodally, M.F. Impact of different extraction solvents and techniques on the biological activities of Cirsium yildizianum (Asteraceae: Cynareae). Ind. Crop. Prod. 2020, 144, 112033. [Google Scholar] [CrossRef]
- Sánchez-Rangel, J.C.; Benavides, J.; Heredia, J.B.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. The Folin–Ciocalteu assay revisited: Improvement of its specificity for total phenolic content determination. Anal. Methods 2013, 5, 5990–5999. [Google Scholar] [CrossRef]
- Wu, L.; Wu, W.; Cai, Y.; Li, C.; Wang, L. HPLC fingerprinting-based multivariate analysis of phenolic compounds in mango leaves varieties: Correlation to their antioxidant activity and in silico α-glucoidase inhibitory ability. J. Pharm. Biomed. Anal. 2020, 191, 113616. [Google Scholar] [CrossRef]
- Ramsay, R.R.; Tipton, K.F. Assessment of enzyme inhibition: A review with examples from the development of monoamine oxidase and cholinesterase inhibitory drugs. Molecules 2017, 22, 1192. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.; Chang, S.K. Digestive enzyme inhibition activity of the phenolic substances in selected fruits, vegetables and tea as compared to black legumes. J. Funct. Foods 2017, 38, 644–655. [Google Scholar] [CrossRef]
- Papoutsis, K.; Zhang, J.; Bowyer, M.C.; Brunton, N.; Gibney, E.R.; Lyng, J. Fruit, vegetables, and mushrooms for the preparation of extracts with α-amylase and α-glucosidase inhibition properties: A review. Food Chem. 2020, 338, 128119. [Google Scholar]
- Yener, I.; Kocakaya, S.O.; Ertas, A.; Erhan, B.; Kaplaner, E.; Oral, E.V.; Yilmaz-Ozden, T.; Yilmaz, M.A.; Ozturk, M.; Kolak, U. Selective in vitro and in silico enzymes inhibitory activities of phenolic acids and flavonoids of food plants: Relations with oxidative stress. Food Chem. 2020, 327, 127045. [Google Scholar] [CrossRef]
- Liao, Z.; Chen, X.; Wu, M. Antidiabetic effect of flavones from Cirsium japonicum DC in diabetic rats. Arch. Pharmacal Res. 2010, 33, 353–362. [Google Scholar] [CrossRef]
- Zengin, G.; Ceylan, R.; Sinan, K.I.; Ak, G.; Uysal, S.; Mahomoodally, M.F.; Lobine, D.; Aktumsek, A.; Cziáky, Z.; Jeko, J. Network analysis, chemical characterization, antioxidant and enzyme inhibitory effects of foxglove (Digitalis cariensis Boiss. ex Jaub. & Spach): A novel raw material for pharmaceutical applications. J. Pharm. Biomed. Anal. 2020, 191, 113614. [Google Scholar]
- Trabelsi, N.; Falleh, H.; Jallali, I.; Daly, A.B.; Hajlaoui, H.; Smaoui, A.; Abdelly, C.; Ksouri, R. Variation of phenolic composition and biological activities in Limoniastrum monopetalum L. organs. Acta Physiol. Plant. 2012, 34, 87–96. [Google Scholar] [CrossRef]
- Fernandez, C.; Monnier, Y.; Ormeño, E.; Baldy, V.; Greff, S.; Pasqualini, V.; Mévy, J.-P.; Bousquet-Mélou, A. Variations in allelochemical composition of leachates of different organs and maturity stages of Pinus halepensis. J. Chem. Ecol. 2009, 35, 970–979. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.A. Rosacea, reactive oxygen species, and azelaic acid. J. Clin. Aesthetic Dermatol. 2009, 2, 26. [Google Scholar]
- Zheleva-Dimitrova, D.; Gevrenova, R.; Zaharieva, M.M.; Najdenski, H.; Ruseva, S.; Lozanov, V.; Balabanova, V.; Yagi, S.; Momekov, G.; Mitev, V. HPLC-UV and LC–MS Analyses of Acylquinic Acids in Geigeria alata (DC) Oliv. & Hiern. and their Contribution to Antioxidant and Antimicrobial Capacity. Phytochem. Anal. 2017, 28, 176–184. [Google Scholar]
- Majewska, M.; Skrzycki, M.; Podsiad, M.; Czeczot, H. Evaluation of antioxidant potential of flavonoids: An in vitro study. Acta Pol. Pharm. 2011, 68, 611–615. [Google Scholar]
- Petruk, G.; Di Lorenzo, F.; Imbimbo, P.; Silipo, A.; Bonina, A.; Rizza, L.; Piccoli, R.; Monti, D.M.; Lanzetta, R. Protective effect of Opuntia ficus-indica L. cladodes against UVA-induced oxidative stress in normal human keratinocytes. Bioorg. Med. Chem. Lett. 2017, 27, 5485–5489. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.-S.; Han, S.-M.; Kim, S.-Y.; Bae, Y.-S.; Kang, I.-J. Isolation of acetylcholinesterase inhibitors from the flowers of Chrysanthemum indicum Linne. Food Sci. Biotechnol. 2007, 16, 265–269. [Google Scholar]
- Yu, Q.; Fan, L.; Duan, Z. Five individual polyphenols as tyrosinase inhibitors: Inhibitory activity, synergistic effect, action mechanism, and molecular docking. Food Chem. 2019, 297, 124910. [Google Scholar] [CrossRef]
- Lim, H.; Son, K.H.; Chang, H.W.; Bae, K.; Kang, S.S.; Kim, H.P. Anti-inflammatory activity of pectolinarigenin and pectolinarin isolated from Cirsium chanroenicum. Biol. Pharm. Bull. 2008, 31, 2063–2067. [Google Scholar] [CrossRef] [Green Version]
- Schallreuter, K.; Wood, J. A possible mechanism of action for azelaic acid in the human epidermis. Arch. Dermatol. Res. 1990, 282, 168–171. [Google Scholar] [CrossRef]
- Zengin, G.; Aktumsek, A. Investigation of antioxidant potentials of solvent extracts from different anatomical parts of Asphodeline anatolica E. Tuzlaci: An endemic plant to Turkey. Afr. J. Tradit. Complementary Altern. Med. 2014, 11, 481–488. [Google Scholar] [CrossRef] [Green Version]
- Grochowski, D.M.; Uysal, S.; Aktumsek, A.; Granica, S.; Zengin, G.; Ceylan, R.; Locatelli, M.; Tomczyk, M. In vitro enzyme inhibitory properties, antioxidant activities, and phytochemical profile of Potentilla thuringiaca. Phytochem. Lett. 2017, 20, 365–372. [Google Scholar] [CrossRef]
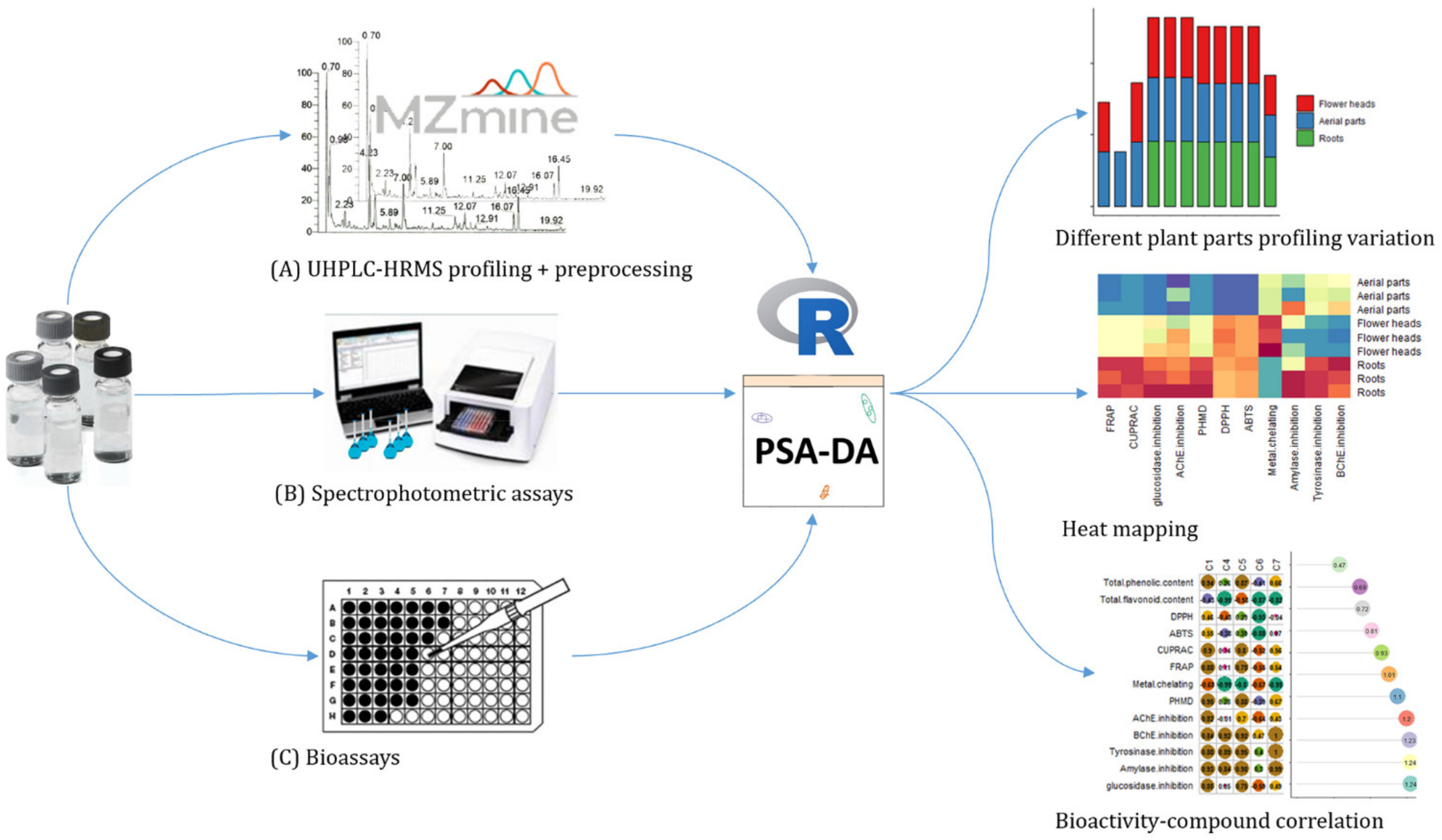




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| № | Identified/Tentatively Annotated Compound | Molecular Formula | Exact Mass [M-H]− | tR (Min) | Δ ppm | Distribution | Level of Identification (CAWG) |
|---|---|---|---|---|---|---|---|
| Carboxylic (including hydroxybenzoic and hydroxycinnamic) acids | |||||||
| 1. | protocatechuic acid a | C7H6O4 | 153.0179 | 2.15 | −7.986 | 1, 2 | 1 |
| 2. | dihydroxybenzoic acid | C7H6O4 | 153.0181 | 3.44 | −8.182 | 2, 3 | 2 |
| 3. | gentisic acid a | C7H6O4 | 153.0179 | 3.86 | −9.685 | 2 | 1 |
| 4. | vanillic acid a | C8H8O4 | 167.0338 | 4.77 | −6.837 | 1, 2, 3 | 1 |
| 5. | caffeic acid a | C9H8O4 | 179.0340 | 3.51 | −5.317 | 1, 2, 3 | 1 |
| 6. | quinic acid | C7H12O6 | 191.0551 | 3.18 | −5.032 | 1, 2, 3 | 2 |
| 7. | eucomic acid | C11H12O6 | 239.0557 | 3.38 | −0.717 | 1, 2, 3 | 2 |
| 8. | caffeoyl-syringic acid | C18H16O8 | 359.0985 | 2.32 | 0.390 | 1, 2, 3 | 4 |
| Hydroxybenzoic and hydroxycinnamc acids glycosides | |||||||
| 9. | 4-hydroxyphenylacetic acid O-β-D-glucoside | C14H18O8 | 313.0933 | 2.18 | 1.467 | 2 | 2 |
| 10. | vanillic acid O-deoxyhexoside | C14H18O8 | 313.0934 | 3.25 | 1.467 | 2, 3 | 2 |
| 11. | gentisic acid O-hexoside | C14H20O8 | 315.1087 | 1.92 | 0.601 | 1, 2, 3 | 2 |
| 12. | p-hydroxybenzoic acid O-hexoside | C14H20O8 | 315.1086 | 2.10 | 0.029 | 1, 2, 3 | 2 |
| 13. | vanillic acid O-hexoside | C14H18O9 | 329.0885 | 1.71 | 2.020 | 1, 2, 3 | 2 |
| 14. | leonuriside A | C14H20O9 | 331.1037 | 1.44 | 0.739 | 1, 2, 3 | 2 |
| 15. | gallic acid O-hexoside | C13H16O10 | 331.0676 | 1.58 | 1.601 | 2 | 2 |
| Acylquinic acids | |||||||
| 16. | 1-p-coumaroylquinic acid | C16H18O8 | 337.0932 | 4.61 | 1.007 | 1, 2 | 2 |
| 17. | 3-p-coumaroylquinic acid | C16H18O8 | 337.0935 | 3.01 | 1.748 | 2 | 2 |
| 18. | 1-caffeoylquinic acid | C16H18O9 | 353.0880 | 2.27 | 0.410 | 1, 2 | 2 |
| 19. | neochlorogenic (3-caffeoylquinic) acid | C16H18O9 | 353.0878 | 3.21 | −0.015 | 1, 2, 3 | 1 |
| 20. | chlorogenic (5-caffeoylquinic) acid a | C16H18O9 | 353.0874 | 3.94 | −1.233 | 1, 2, 3 | 1 |
| 21. | 4-caffeoylquinic acid | C16H18O9 | 353.0879 | 6.27 | 0.155 | 1, 2, 3 | 2 |
| 22. | 3,4-dicaffeoylquinic acid a | C25H24O12 | 515.1199 | 5.73 | 0.836 | 1, 2, 3 | 1 |
| 23. | 1,5-dicaffeoylquinic acid a | C25H24O12 | 515.1191 | 5.91 | −0.697 | 1, 2, 3 | 1 |
| 24. | 3,5-dicaffeoylquinic acid | C25H24O12 | 515.1199 | 6.08 | 0.720 | 1, 2, 3 | 1 |
| 25. | 4,5-dicaffeoylquinic acid | C25H24O12 | 515.1191 | 6.25 | −0.697 | 1, 2, 3 | 1 |
| 26. | 1,3,5-tricaffeoylquinic acid | C34H30O15 | 677.1512 | 5.15 | - | 1, 2, 3 | 1 |
| Flavonoids | |||||||
| 27. | apigenin a | C15H9O5 | 269.0459 | 8.58 | 1.313 | 1, 3 | 1 |
| 28. | genkwanin a | C16H12O5 | 283.0608 | 11.41 | −1.543 | 2 | 1 |
| 29. | acacetin | C16H12O5 | 283.0615 | 11.40 | 1.142 | 1, 3 | 2 |
| 30. | luteolin a | C15H10O6 | 285.0404 | 7.55 | −0.075 | 1, 3 | 1 |
| 31. | hispidulin (scutellarein-6-methyl ether) a | C16H12O6 | 299.0561 | 8. 81 | −0.172 | 1, 2, 3 | 1 |
| 32. | diosmetin | C16H12O6 | 299.0560 | 9.28 | −0.272 | 1 | 1 |
| 33. | quercetin a | C15H9O6 | 301.0354 | 7.61 | 1.11 | 1 | 1 |
| 34. | pectolinarigenin | C17H14O6 | 313.0722 | 12.26 | 1.305 | 1, 2, 3 | 2 |
| 35. | nepetin (6-methoxyluteolin) | C16H11O7 | 315.0514 | 8.09 | 1.251 | 1, 3 | 2 |
| 36. | cirsiliol | C17H14O7 | 329.0669 | 8.87 | 0.772 | 1 | 2 |
| 37. | apigenin 7-O-glucoside a | C21H20O10 | 431.0988 | 6.06 | 0.835 | 1 | 1 |
| 38. | kaempferol 3-O-deoxyhexoside | C21H20O10 | 431.0983 | 6.60 | −0.232 | 1, 2 | 2 |
| 39. | apigenin O-hexuronide | C21H18O11 | 445.0770 | 6.45 | −0.347 | 1, 2, 3 | 2 |
| 40. | kaempferol 3-O-glucoside a | C21H20O11 | 447.0935 | 5.63 | 0.571 | 1, 2 | 1 |
| 41. | luteolin 7-O-glucoside a | C21H19O11 | 447.0934 | 6.04 | 0.281 | 1, 2, 3 | 1 |
| 42. | luteolin 7-O-hexuronide | C21H18O12 | 461.0734 | 5.37 | 1.911 | 1, 3 | 2 |
| 43. | diosmetin 7-O-hexoside | C22H22O11 | 461.1092 | 6.30 | 0.684 | 1, 2, 3 | 2 |
| 44. | hispidulin 7-O-hexoside | C22H22O11 | 461.1093 | 6.67 | 0.966 | 1, 2, 3 | 2 |
| 45. | hispidulin-O-hexuronide | C22H20O12 | 475.0882 | 6.33 | 0.002 | 1, 3 | 2 |
| 46. | pectolinarigenin-O-hexoside | C23H24O11 | 475.1247 | 8.11 | 0.159 | 1 | 2 |
| 47. | nepetin-O-hexoside | C22H21O12 | 477.1040 | 5.65 | 0.316 | 1, 3 | 2 |
| 48. | nepetin-O-hexuronide | C22H20O13 | 491.0835 | 6.32 | 0.725 | 1 | 2 |
| 49. | acaciin (acacetin 7-O-rutinoside) a | C28H32O14 | 591.1730 | 7.59 | 3.622 | 1, 2, 3 | 1 |
| 50. | kaempferol 3-O-rutinoside a | C27H30O15 | 593.1532 | 5.40 | 3.383 | 1, 3 | 1 |
| 51. | hispidulin 7-O-rutinoside | C28H32O15 | 607.1675 | 6.34 | 1.049 | 1, 2, 3 | 2 |
| 52. | pectolinarin (pectolinarigenin 7-O- rutinoside) a | C29H34O15 | 621.1824 | 7.67 | −0.199 | 1, 2, 3 | 1 |
| Free fatty acids | |||||||
| 53. | nonanedioic acid (azelaic acid) | C9H16O4 | 187.0967 | 6.32 | −4.502 | 1, 2, 3 | 2 |
| 54. | 3-hydroxysuberic acid | C8H14O5 | 189.0758 | 4.64 | −5.483 | 1, 2, 3 | 2 |
| 55. | 3-hydroxyazelaic acid | C9H16O5 | 203.0918 | 6.25 | −3.677 | 1, 2, 3 | 2 |
| 56. | 2-dodecenoic acid | C12H20O4 | 227.1287 | 9.46 | −0.715 | 2, 3 | 2 |
| 57. | 9,13-dyhidroxyoctadeca-9,11,13-trienoic acid | C18H30O4 | 309.2074 | 12.76 | 0.768 | 2, 3 | 2 |
| 58. | 11,12-dyhidroxyoctadeca-9,13,15-trienoic acid | C18H30O4 | 309.2075 | 12.91 | −0.332 | 2 | 2 |
| 59. | 9,10-dyhidroxyoctadeca-12,14,16-trienoic acid | C18H30O4 | 309.2074 | 10.81 | 1.835 | 2 | 2 |
| 60. | 9,13-dyhidroxyoctadeca-11,13-dienoic acid | C18H32O4 | 311.2231 | 13.67 | 0.859 | 1, 2, 3 | 2 |
| 61. | 9,10-dyhidroxyoctadeca-9-enoic acid | C18H34O4 | 313.2388 | 13.79 | 0.885 | 3 | 2 |
| Parts | Total Phenolic Content (mgGAE/g) | Total Flavonoid Content (mgRE/g) | DPPH• (mgTE/g) | ABTS•+ (mgTE/g) | CUPRAC (mgTE/g) | FRAP (mg TE/g) | PHMD (mmolTE/g) | Metal Chelating (mgEDTAE/g) |
|---|---|---|---|---|---|---|---|---|
| Flower heads | 71.75 ± 1.47 b | 46.59 ± 0.89 a | 101.79 ± 0.15 a | 224.57 ± 0.57 a | 356.97 ± 11.52 b | 169.60 ± 0.84 b | 1.71 ± 0.07 b | 32.53 ± 3.51 a |
| Aerial parts | 26.02 ± 1.49 c | 2.64 ± 0.08 c | 70.25 ± 1.91 c | 124.16 ± 4.73 b | 103.77 ± 5.89 c | 69.98 ± 2.01 c | 0.74 ± 0.01 c | 9.42 ± 0.54 b |
| Roots | 143.62 ± 2.99 a | 3.99 ± 0.06 b | 97.95 ± 0.60 b | 224.59 ± 0.33 a | 618.36 ± 5.17 a | 269.89 ± 8.50 a | 3.36 ± 0.15 a | na |
| Parts | AChE Inhibition (mgGALAE/g) | BChE Inhibition (mgGALAE/g) | Tyrosinase (mgKAE/g) | Amylase (mmolACAE/g) | Glucosidase (mmolACAE/g) |
|---|---|---|---|---|---|
| Flower heads | 4.40 ± 0.40 a | 1.54 ± 0.07 c | 97.78 ± 0.76 c | 0.60 ± 0.01 a | 0.41 ± 0.10 b |
| Aerial parts | 3.52 ± 0.31 b | 2.67 ± 0.34 b | 110.61 ± 0.79 b | 0.61 ± 0.06 a | na |
| Roots | 4.93 ± 0.25 a | 3.80 ± 0.26 a | 127.99 ± 0.68 a | 0.62 ± 0.04 a | 0.72 ± 0.07 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheleva-Dimitrova, D.; Zengin, G.; Ak, G.; Sinan, K.I.; Mahomoodally, M.F.; Gevrenova, R.; Balabanova, V.; Stefanova, A.; Nedialkov, P.; Voynikov, Y. Innovative Biochemometric Approach to the Metabolite and Biological Profiling of the Balkan Thistle (Cirsium appendiculatum Griseb.), Asteraceae. Plants 2021, 10, 2046. https://doi.org/10.3390/plants10102046
Zheleva-Dimitrova D, Zengin G, Ak G, Sinan KI, Mahomoodally MF, Gevrenova R, Balabanova V, Stefanova A, Nedialkov P, Voynikov Y. Innovative Biochemometric Approach to the Metabolite and Biological Profiling of the Balkan Thistle (Cirsium appendiculatum Griseb.), Asteraceae. Plants. 2021; 10(10):2046. https://doi.org/10.3390/plants10102046
Chicago/Turabian StyleZheleva-Dimitrova, Dimitrina, Gokhan Zengin, Gunes Ak, Kouadio Ibrahime Sinan, Mohamad Fawzi Mahomoodally, Reneta Gevrenova, Vessela Balabanova, Alexandra Stefanova, Paraskev Nedialkov, and Yulian Voynikov. 2021. "Innovative Biochemometric Approach to the Metabolite and Biological Profiling of the Balkan Thistle (Cirsium appendiculatum Griseb.), Asteraceae" Plants 10, no. 10: 2046. https://doi.org/10.3390/plants10102046
APA StyleZheleva-Dimitrova, D., Zengin, G., Ak, G., Sinan, K. I., Mahomoodally, M. F., Gevrenova, R., Balabanova, V., Stefanova, A., Nedialkov, P., & Voynikov, Y. (2021). Innovative Biochemometric Approach to the Metabolite and Biological Profiling of the Balkan Thistle (Cirsium appendiculatum Griseb.), Asteraceae. Plants, 10(10), 2046. https://doi.org/10.3390/plants10102046