
Anti-Inflammatory Effects of Huberia peruviana Cogn. Methanol Extract by Inhibiting Src Activity in the NF-κB Pathway

 , and
, and 
Abstract
:1. Introduction
2. Results
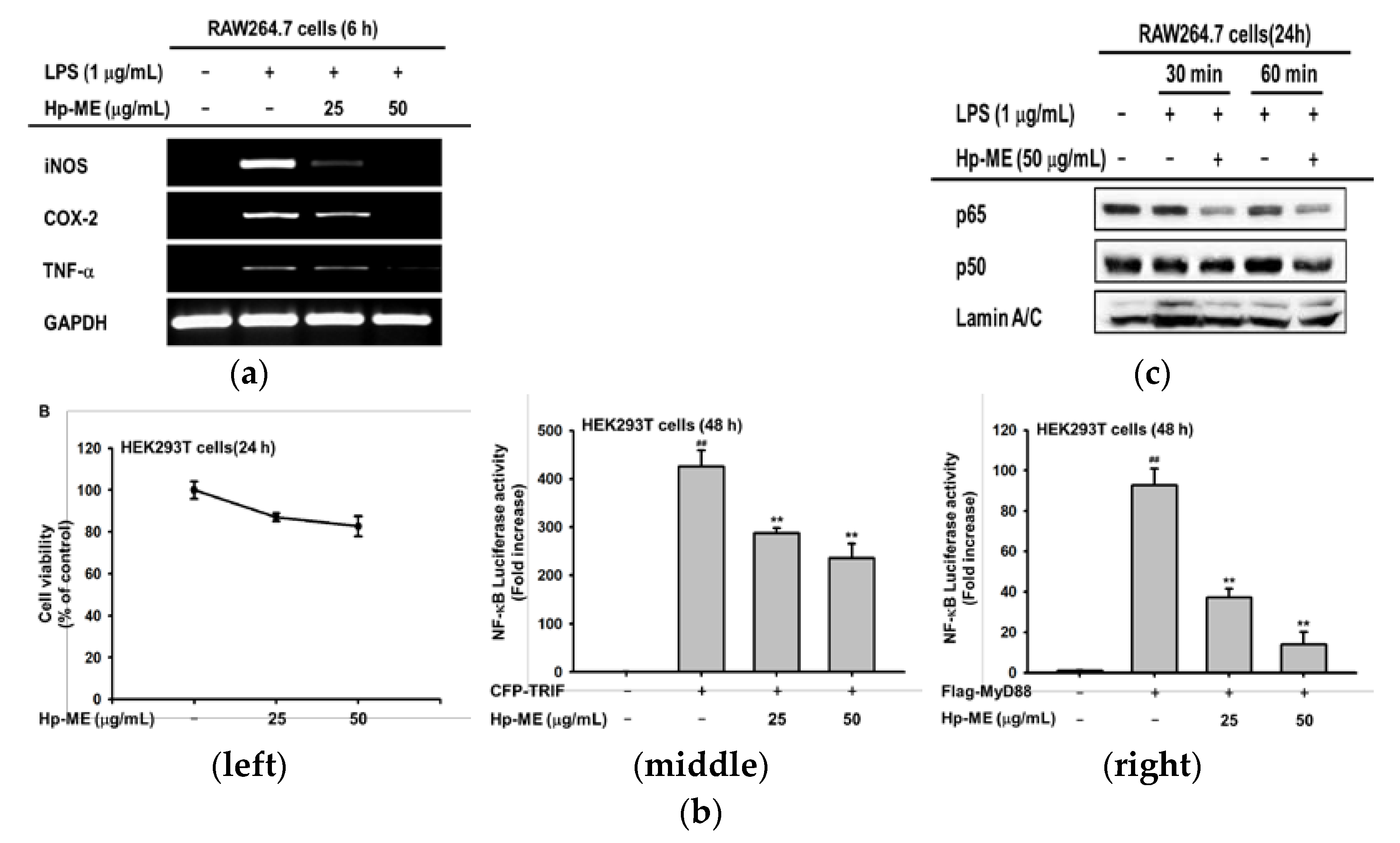
2.1. Hp-ME Reduces NO Production in Inflammatory Responses Mediated by TLR
2.2. Hp-ME Has an Anti-Inflammatory Effect at the mRNA Expression and Transcriptional Level
2.3. Hp-ME Disrupts Phosphorylation of Src Kinase in the Intracellular NF-κB Signaling Pathway
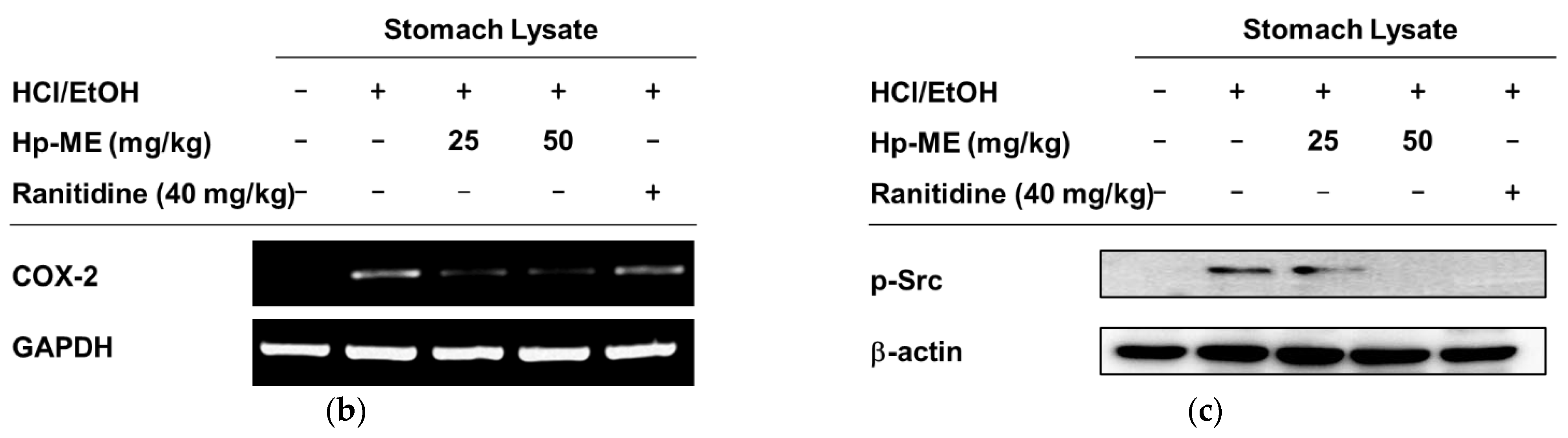
2.4. Ulcerative Lesions Were Alleviated by Hp-ME in an HCl/EtOH-Induced Gastritis Mouse Model
2.5. Lung Injury Was Attenuated by Hp-ME in an LPS-Induced ALI Mouse Model
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Plant Extraction
4.3. Cell Culture
4.4. Nitric Oxide (NO) Production Assay
4.5. Liquid Chromatography–Mass Spectrometry (LC-MS)
4.6. Cell Viability Assay
4.7. Analysis of mRNA Expression Level Using Semiquantitative and Quantitative Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.8. Luciferase Reporter Gene Activity Assay
4.9. Preparation of Total Cell and Nuclear Lysates
4.10. Western Blotting Analysis
4.11. In Vivo HCl/EtOH-Induced Acute Gastritis Mouse Model
4.12. In Vivo LPS-Induced ALI Model
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schett, G.; Neurath, M.F. Resolution of chronic inflammatory disease: Universal and tissue-specific concepts. Nat. Commun. 2018, 9, 3261. [Google Scholar] [CrossRef]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef]
- Marques, R.E.; Marques, P.E.; Guabiraba, R.; Teixeira, M.M. Exploring the Homeostatic and Sensory Roles of the Immune System. Front. Immunol. 2016, 7, 125. [Google Scholar] [CrossRef] [Green Version]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacomini, E.; Iona, E.; Ferroni, L.; Miettinen, M.; Fattorini, L.; Orefici, G.; Julkunen, I.; Coccia, E.M. Infection of human macrophages and dendritic cells with Mycobacterium tuberculosis induces a differential cytokine gene expression that modulates T cell response. J. Immunol. 2001, 166, 7033–7041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escamilla-Tilch, M.; Filio-Rodriguez, G.; Garcia-Rocha, R.; Mancilla-Herrera, I.; Mitchison, N.A.; Ruiz-Pacheco, J.A.; Sanchez-Garcia, F.J.; Sandoval-Borrego, D.; Vazquez-Sanchez, E.A. The interplay between pathogen-associated and danger-associated molecular patterns: An inflammatory code in cancer? Immunol. Cell Biol. 2013, 91, 601–610. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansson Kvarnhammar, A.; Tengroth, L.; Adner, M.; Cardell, L.O. Innate immune receptors in human airway smooth muscle cells: Activation by TLR1/2, TLR3, TLR4, TLR7 and NOD1 agonists. PLoS ONE 2013, 8, e68701. [Google Scholar] [CrossRef] [Green Version]
- Brikos, C.; O’Neill, L.A. Signalling of toll-like receptors. Handb. Exp. Pharmacol. 2008, 183, 21–50. [Google Scholar] [CrossRef]
- Song, C.; Hong, Y.H.; Park, J.G.; Kim, H.G.; Jeong, D.; Oh, J.; Sung, G.H.; Hossain, M.A.; Taamalli, A.; Kim, J.H.; et al. Suppression of Src and Syk in the NF-kappaB signaling pathway by Olea europaea methanol extract is leading to its anti-inflammatory effects. J. Ethnopharmacol. 2019, 235, 38–46. [Google Scholar] [CrossRef]
- Limtrakul, P.; Yodkeeree, S.; Pitchakarn, P.; Punfa, W. Suppression of Inflammatory Responses by Black Rice Extract in RAW 264.7 Macrophage Cells via Downregulation of NF-kB and AP-1 Signaling Pathways. Asian Pac. J. Cancer Prev. 2015, 16, 4277–4283. [Google Scholar] [CrossRef] [Green Version]
- Krishnamoorthy, S.; Honn, K.V. Inflammation and disease progression. Cancer Metastasis Rev. 2006, 25, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Baumgratz, J.F.A.J.N. Three new species of Huberia (Melastomataceae) from Peru. Novon 1999, 9, 139–146. [Google Scholar] [CrossRef]
- Al-Harbi, N.O.; Imam, F.; Al-Harbi, M.M.; Ansari, M.A.; Zoheir, K.M.; Korashy, H.M.; Sayed-Ahmed, M.M.; Attia, S.M.; Shabanah, O.A.; Ahmad, S.F. Dexamethasone Attenuates LPS-induced Acute Lung Injury through Inhibition of NF-kappaB, COX-2, and Pro-inflammatory Mediators. Immunol. Investig. 2016, 45, 349–369. [Google Scholar] [CrossRef]
- Matute-Bello, G.; Downey, G.; Moore, B.B.; Groshong, S.D.; Matthay, M.A.; Slutsky, A.S.; Kuebler, W.M. Acute Lung Injury in Animals Study, G. An official American Thoracic Society workshop report: Features and measurements of experimental acute lung injury in animals. Am. J. Respir. Cell Mol. Biol. 2011, 44, 725–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardinal-Fernandez, P.; Lorente, J.A.; Ballen-Barragan, A.; Matute-Bello, G. Acute Respiratory Distress Syndrome and Diffuse Alveolar Damage. New Insights on a Complex Relationship. Ann. Am. Thorac. Soc. 2017, 14, 844–850. [Google Scholar] [CrossRef]
- Duan, L.; Rao, X.; Sigdel, K.R. Regulation of Inflammation in Autoimmune Disease. J. Immunol. Res. 2019, 2019, 7403796. [Google Scholar] [CrossRef] [Green Version]
- Hanada, T.; Yoshimura, A. Regulation of cytokine signaling and inflammation. Cytokine Growth Factor Rev. 2002, 13, 413–421. [Google Scholar] [CrossRef]
- Nathan, C. Points of control in inflammation. Nature 2002, 420, 846–852. [Google Scholar] [CrossRef]
- Ward, P.A.; Lentsch, A.B. The acute inflammatory response and its regulation. Arch. Surg. 1999, 134, 666–669. [Google Scholar] [CrossRef] [Green Version]
- Korhonen, R.; Lahti, A.; Kankaanranta, H.; Moilanen, E. Nitric oxide production and signaling in inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 471–479. [Google Scholar] [CrossRef]
- Jeong, D.; Yi, Y.S.; Sung, G.H.; Yang, W.S.; Park, J.G.; Yoon, K.; Yoon, D.H.; Song, C.; Lee, Y.; Rhee, M.H.; et al. Anti-inflammatory activities and mechanisms of Artemisia asiatica ethanol extract. J. Ethnopharmacol. 2014, 152, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Park, J.G.; Sung, G.H.; Yang, S.; Yang, W.S.; Kim, E.; Kim, J.H.; Ha, V.T.; Kim, H.G.; Yi, Y.S.; et al. Kaempferol, a dietary flavonoid, ameliorates acute inflammatory and nociceptive symptoms in gastritis, pancreatitis, and abdominal pain. Mol. Nutr. Food Res. 2015, 59, 1400–1405. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Gang, D.; Yu, X.; Hu, Y.; Yue, Y.; Cheng, W.; Pan, X.; Zhang, P. Genistein: The potential for efficacy in rheumatoid arthritis. Clin. Rheumatol. 2013, 32, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Moscatelli, V.; Hnatyszyn, O.; Acevedo, C.; Megias, J.; Alcaraz, M.J.; Ferraro, G. Flavonoids from Artemisia copa with anti-inflammatory activity. Planta Med. 2006, 72, 72–74. [Google Scholar] [CrossRef] [Green Version]
- Aktan, F. iNOS-mediated nitric oxide production and its regulation. Life Sci. 2004, 75, 639–653. [Google Scholar] [CrossRef]
- Morita, I. Distinct functions of COX-1 and COX-2. Prostaglandins Other Lipid Mediat. 2002, 68–69, 165–175. [Google Scholar] [CrossRef]
- Schottelius, A.J.; Baldwin, A.S., Jr. A role for transcription factor NF-kappa B in intestinal inflammation. Int. J. Colorectal Dis. 1999, 14, 18–28. [Google Scholar] [CrossRef]
- Roskoski, R., Jr. Src protein-tyrosine kinase structure and regulation. Biochem. Biophys. Res. Commun. 2004, 324, 1155–1164. [Google Scholar] [CrossRef]
- Dudonne, S.; Vitrac, X.; Coutiere, P.; Woillez, M.; Merillon, J.M. Comparative study of antioxidant properties and total phenolic content of 30 plant extracts of industrial interest using DPPH, ABTS, FRAP, SOD, and ORAC assays. J. Agric. Food Chem. 2009, 57, 1768–1774. [Google Scholar] [CrossRef]
- Lorz, L.R.; Kim, D.; Kim, M.Y.; Cho, J.Y. Panax ginseng-derived fraction BIOGF1K reduces atopic dermatitis responses via suppression of mitogen-activated protein kinase signaling pathway. J. Ginseng Res. 2020, 44, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Choi, E.; Hong, Y.H.; Kim, H.; Jang, Y.J.; Lee, J.S.; Choung, E.S.; Woo, B.Y.; Hong, Y.D.; Lee, S.; et al. Syk/NF-kappaB-targeted anti-inflammatory activity of Melicope accedens (Blume) T.G. Hartley methanol extract. J. Ethnopharmacol. 2021, 271, 113887. [Google Scholar] [CrossRef]
- Rahmawati, L.; Aziz, N.; Oh, J.; Hong, Y.H.; Woo, B.Y.; Hong, Y.D.; Manilack, P.; Souladeth, P.; Jung, J.H.; Lee, W.S.; et al. Cissus subtetragona Planch. Ameliorates Inflammatory Responses in LPS-induced Macrophages, HCl/EtOH-induced Gastritis, and LPS-induced Lung Injury via Attenuation of Src and TAK1. Molecules 2021, 26, 6073. [Google Scholar] [CrossRef]
- Lee, J.O.; Kim, J.H.; Kim, S.; Kim, M.Y.; Hong, Y.H.; Kim, H.G.; Cho, J.Y. Gastroprotective effects of the nonsaponin fraction of Korean Red Ginseng through cyclooxygenase-1 upregulation. J. Ginseng Res. 2020, 44, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.; Kim, E.; Kim, J.H.; Yoon, K.; Kim, S.; Lee, J.; Cho, J.Y. AKT1-targeted proapoptotic activity of compound K in human breast cancer cells. J. Ginseng Res. 2019, 43, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; Kim, J.H.; Jeong, D.; Hong, Y.H.; Park, S.H.; Yang, Y.; Jang, Y.J.; Kim, J.H.; Cho, J.Y. 3-Deazaadenosine, an S-adenosylhomocysteine hydrolase inhibitor, attenuates lipopolysaccharide-induced inflammatory responses via inhibition of AP-1 and NF-kappaB signaling. Biochem. Pharmacol. 2020, 182, 114264. [Google Scholar] [CrossRef]
- Kim, H.G.; Choi, S.; Lee, J.; Hong, Y.H.; Jeong, D.; Yoon, K.; Yoon, D.H.; Sung, G.H.; Lee, S.; Hong, S.; et al. Src Is a Prime Target Inhibited by Celtis choseniana Methanol Extract in Its Anti-Inflammatory Action. Evid. Based Complement. Altern. Med. 2018, 2018, 3909038. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Score per Field | ||
|---|---|---|---|
| 0 | 1 | 2 | |
| A. Neutrophils in the alveolar space | None | 1–5 | >5 |
| B. Neutrophils in the interstitial space | None | 1–5 | >5 |
| C. Hyaline membranes | None | 3 | >3 |
| D. Alveolar septal thickening | <2X | 2X–4X | >4X |
| Gene | Direction | Sequences (5′ → 3′) |
|---|---|---|
| iNOS | Forward Reverse | TGCCAGGGTCACAACTTTACA ACCCCAAGCAAGACTTGGAC |
| COX-2 | Forward Reverse | TGAGTACCGCAACGCTTCT TGGGAGGCACTTGCATTGAT |
| TNF-α | Forward Reverse | TTGACCTCAGCGCTGAGTTG CCTGTAGCCCACGTCGTAGC |
| GAPDH | Forward Reverse | GAAGGTCGGTGTGAACGGAT AGTGATGGCATGGACTGTGG |
| Gene | Direction | Sequences (5′ → 3′) |
|---|---|---|
| TNF-α | Forward Reverse | TGCCTATGTCTCAGCCTCTT GAGGCCATTTGGGAACTTCT |
| GAPDH | Forward Reverse | TGTGAACGGATTTGGCCGTA ACTGTGCCGTTGAATTGCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.A.; Lee, C.Y.; Mitra, A.; Kim, H.; Woo, B.Y.; Hong, Y.D.; Noh, J.K.; Yi, D.-K.; Kim, H.G.; Cho, J.Y. Anti-Inflammatory Effects of Huberia peruviana Cogn. Methanol Extract by Inhibiting Src Activity in the NF-κB Pathway. Plants 2021, 10, 2335. https://doi.org/10.3390/plants10112335
Kim SA, Lee CY, Mitra A, Kim H, Woo BY, Hong YD, Noh JK, Yi D-K, Kim HG, Cho JY. Anti-Inflammatory Effects of Huberia peruviana Cogn. Methanol Extract by Inhibiting Src Activity in the NF-κB Pathway. Plants. 2021; 10(11):2335. https://doi.org/10.3390/plants10112335
Chicago/Turabian StyleKim, Seung A, Chae Young Lee, Ankita Mitra, Haeyeop Kim, Byoung Young Woo, Yong Deog Hong, Jin Kyoung Noh, Dong-Keun Yi, Han Gyung Kim, and Jae Youl Cho. 2021. "Anti-Inflammatory Effects of Huberia peruviana Cogn. Methanol Extract by Inhibiting Src Activity in the NF-κB Pathway" Plants 10, no. 11: 2335. https://doi.org/10.3390/plants10112335
APA StyleKim, S. A., Lee, C. Y., Mitra, A., Kim, H., Woo, B. Y., Hong, Y. D., Noh, J. K., Yi, D.-K., Kim, H. G., & Cho, J. Y. (2021). Anti-Inflammatory Effects of Huberia peruviana Cogn. Methanol Extract by Inhibiting Src Activity in the NF-κB Pathway. Plants, 10(11), 2335. https://doi.org/10.3390/plants10112335