
The Combination of Untargeted Metabolomics and Machine Learning Predicts the Biosynthesis of Phenolic Compounds in Bryophyllum Medicinal Plants (Genus Kalanchoe)

 , ,
, , 
 ,
,  and
and
Abstract
:1. Introduction
2. Results
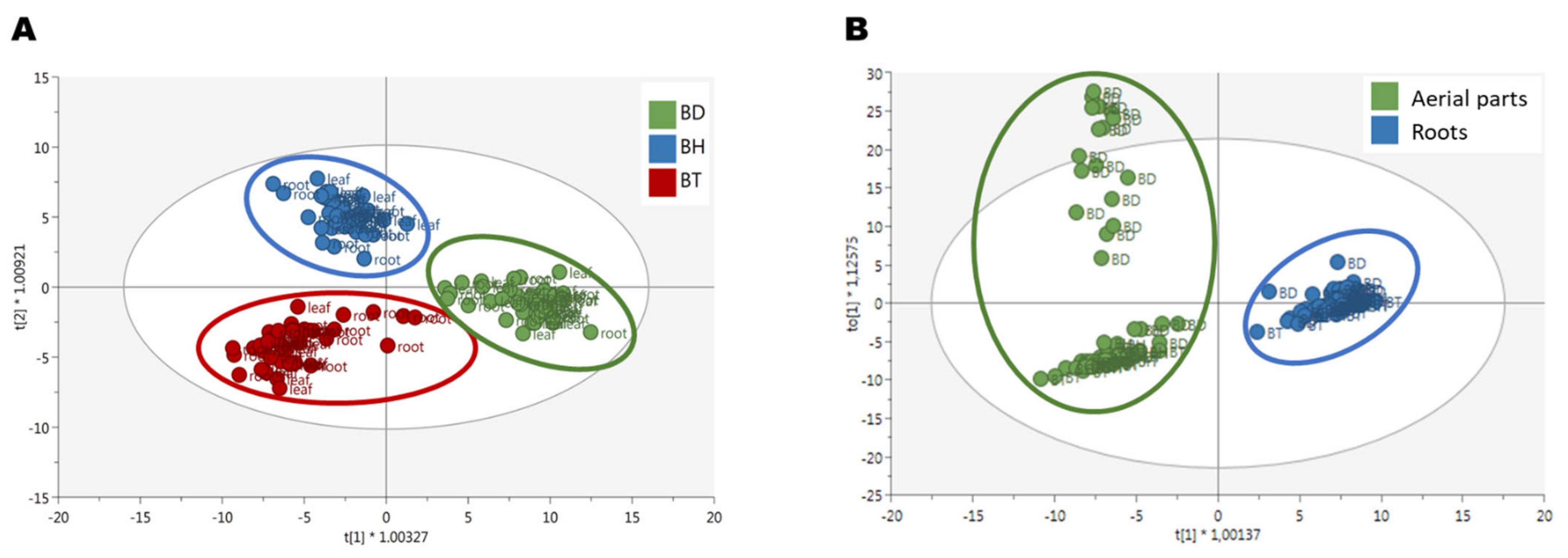
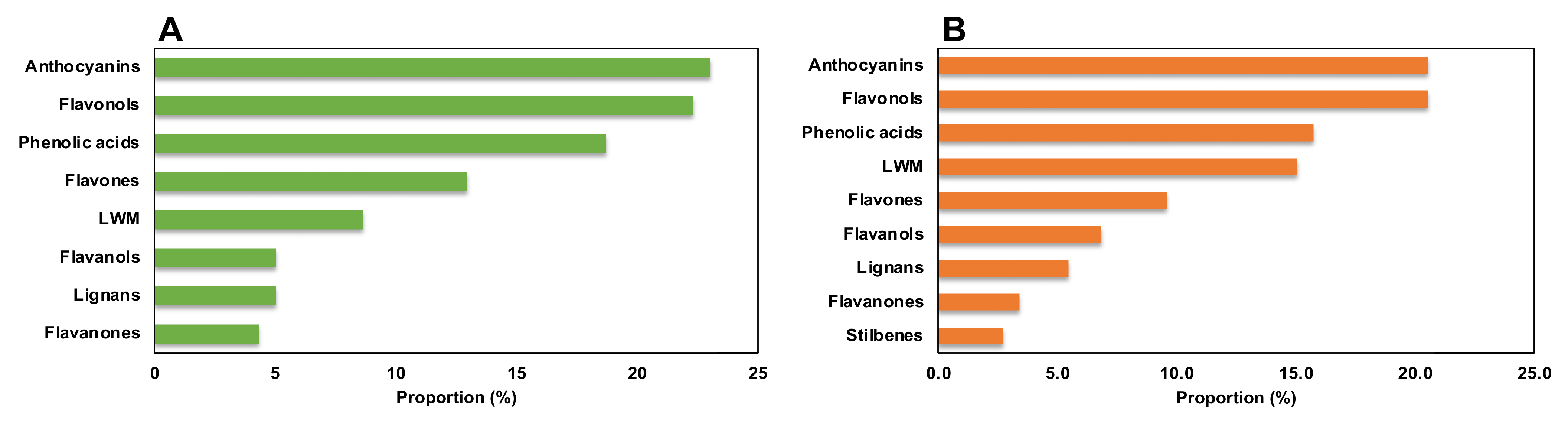
2.1. Determination of Phenolic Profiling by Untargeted Metabolomics
2.2. Machine Learning Prediction of the Biosynthesis of Phenolic Compounds
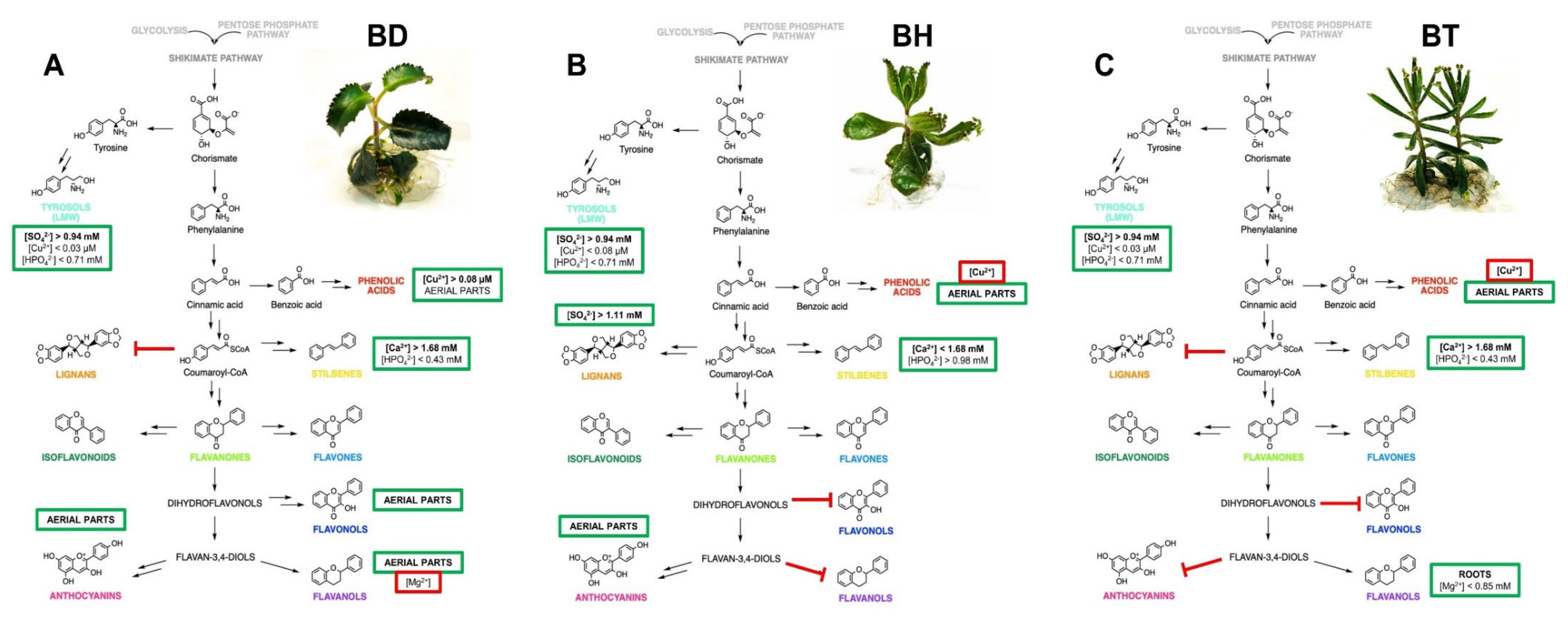
2.3. Proposed Mechanism of Phenolic Compound Biosynthesis of Bryophyllum Plants Cultured In Vitro
3. Discussion
4. Materials and Methods
4.1. Plant Material and Culture Conditions
4.2. Experimental Design
4.3. Sample Preparation and Extraction
4.4. Phenolic Profiling Using Untargeted Metabolomics
4.5. Statistical Analysis
4.6. Modeling Tools
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- García-Pérez, P.; Barreal, M.; Dios, L.R.-D.; Cameselle-Teijeiro, J.; Gallego, P. Bioactive Natural Products from the Genus Kalanchoe as Cancer Chemopreventive Agents: A Review. In Studies in Natural Products Chemistry; Rahman, A., Ed.; Elsevier BV: Amsterdam, The Netherlands, 2019; Volume 61, pp. 49–84. [Google Scholar]
- García-Pérez, P.; Lozano-Milo, E.; Landin, M.; Gallego, P.P. From Ethnomedicine to Plant Biotechnology and Machine Learning: The Valorization of the Medicinal Plant Bryophyllum sp. Pharmaceuticals 2020, 13, 444. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Milo, E.; García-Pérez, P.; Gallego, P.P. Narrative review of production of antioxidants and anticancer compounds from Bryophyllum spp. (Kalanchoe) using plant cell tissue culture. Longhua Chin. Med. 2020, 3, 18. [Google Scholar] [CrossRef]
- Katrucha, E.M.; Lopes, J.; Paim, M.; dos Santos, J.C.; Siebert, D.A.; Micke, G.A.; Vitali, L.; Alberton, M.D.; Tenfen, A. Phenolic profile by HPLC-ESI-MS/MS and enzymatic inhibitory effect of Bryophyllum delagoense. Nat. Prod. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Stefanowicz-Hajduk, J.; Asztemborska, M.; Krauze-Baranowska, M.; Godlewska, S.; Gucwa, M.; Moniuszko-Szajwaj, B.; Stochmal, A.; Ochocka, J.R. Identification of Flavonoids and Bufadienolides and Cytotoxic Effects of Kalanchoe daigremontiana Extracts on Human Cancer Cell Lines. Planta Medica 2020, 86, 239–246. [Google Scholar] [CrossRef]
- García-Pérez, P.; Ayuso, M.; Lozano-Milo, E.; Pereira, C.; Dias, M.I.; Ivanov, M.; Calhelha, R.C.; Soković, M.; Ferreira, I.C.F.R.; Barros, L.; et al. Phenolic profiling and in vitro bioactivities of three medicinal Bryophyllum plants. Ind. Crop. Prod. 2021, 162, 113241. [Google Scholar] [CrossRef]
- García-Pérez, P.; Losada-Barreiro, S.; Bravo-Díaz, C.; Gallego, P.P. Exploring the use of Bryophyllum as natural source of bioactive compounds with antioxidant activity to prevent lipid oxidation of fish oil-in-water emulsions. Plants 2020, 9, 1012. [Google Scholar] [CrossRef]
- Tohge, T.; Watanabe, M.; Hoefgen, R.; Fernie, A.R. Shikimate and Phenylalanine Biosynthesis in the Green Lineage. Front. Plant Sci. 2013, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Saito, K.; Yonekura-Sakakibara, K.; Nakabayashi, R.; Higashi, Y.; Yamazaki, M.; Tohge, T.; Fernie, A.R. The flavonoid biosynthetic pathway in Arabidopsis: Structural and genetic diversity. Plant Physiol. Biochem. 2013, 72, 21–34. [Google Scholar] [CrossRef] [Green Version]
- Bogucka-Kocka, A.; Zidorn, C.; Kasprzycka, M.; Szymczak, G.; Szewczyk, K. Phenolic acid content, antioxidant and cytotoxic activities of four Kalanchoë species. Saudi J. Biol. Sci. 2018, 25, 622–630. [Google Scholar] [CrossRef] [Green Version]
- Hasanpour, M.; Iranshahy, M.; Iranshahi, M. The application of metabolomics in investigating anti-diabetic activity of medicinal plants. Biomed. Pharmacother. 2020, 128, 110263. [Google Scholar] [CrossRef]
- Wolfender, J.; Queiroz, E.F.; Allard, P. Massive metabolite profiling of natural extracts for a rational prioritization of bioactive natural products: A paradigm shift in pharmacognosy. Food Front. 2020, 1, 105–106. [Google Scholar] [CrossRef] [Green Version]
- Marchev, A.S.; Georgiev, M.I. Plant In Vitro Systems as a Sustainable Source of Active Ingredients for Cosmeceutical Application. Molecules 2020, 25, 2006. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.I.; Sousa, M.J.; Alves, R.C.; Ferreira, I.C. Exploring plant tissue culture to improve the production of phenolic compounds: A review. Ind. Crop. Prod. 2016, 82, 9–22. [Google Scholar] [CrossRef] [Green Version]
- Espinosa-Leal, C.A.; Puente-Garza, C.A.; García-Lara, S. In vitro plant tissue culture: Means for production of biological active compounds. Planta 2018, 248, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Phillips, G.C.; Garda, M. Plant tissue culture media and practices: An overview. Vitr. Cell. Dev. Biol. Anim. 2019, 55, 242–257. [Google Scholar] [CrossRef]
- Da Silva, J.A.T.; Nezami-Alanagh, E.; Barreal, M.E.; Kher, M.M.; Wicaksono, A.; Gulyás, A.; Hidvégi, N.; Magyar-Tábori, K.; Mendler-Drienyovszki, N.; Márton, L.; et al. Shoot tip necrosis of in vitro plant cultures: A reappraisal of possible causes and solutions. Planta 2020, 252, 1–35. [Google Scholar] [CrossRef]
- García-Pérez, P.; Lozano-Milo, E.; Landín, M.; Gallego, P.P. Combining Medicinal Plant In Vitro Culture with Machine Learning Technologies for Maximizing the Production of Phenolic Compounds. Antioxidants 2020, 9, 210. [Google Scholar] [CrossRef] [Green Version]
- Gallego, P.P.; Gago, J.; Landín, M. Artificial neural networks technology to model and predict plant biology process. In Meth-odological Advances and Biomedical Applications; Suzuki, K., Ed.; IntechOpen: Rijeka, Croatia, 2011. [Google Scholar]
- Nezami-Alanagh, E.; Garoosi, G.-A.; Landin, M.; Gallego, P.P. Combining DOE with Neurofuzzy Logic for Healthy Mineral Nutrition of Pistachio Rootstocks in vitro Culture. Front. Plant Sci. 2018, 9, 1474. [Google Scholar] [CrossRef]
- Hesami, M.; Jones, A.M.P. Application of artificial intelligence models and optimization algorithms in plant cell and tissue culture. Appl. Microbiol. Biotechnol. 2020, 104, 9449–9485. [Google Scholar] [CrossRef]
- Landín, M.; Rowe, R.C.; York, P. Advantages of neurofuzzy logic against conventional experimental design and statistical analysis in studying and developing direct compression formulations. Eur. J. Pharm. Sci. 2009, 38, 325–331. [Google Scholar] [CrossRef]
- Gago, J.; Martínez-Núñez, L.; Landín, M.; Gallego, P.P. Artificial neural networks as an alternative to the traditional statistical methodology in plant research. J. Plant Physiol. 2010, 167, 23–27. [Google Scholar] [CrossRef]
- Niazian, M.; Nalousi, A.M. Artificial polyploidy induction for improvement of ornamental and medicinal plants. Plant Cell Tissue Organ Cult. 2020, 142, 447–469. [Google Scholar] [CrossRef]
- Ayuso, M.; Ramil-Rego, P.; Landin, M.; Gallego, P.P.; Barreal, M.E. Computer-Assisted Recovery of Threatened Plants: Keys for Breaking Seed Dormancy of Eryngium viviparum. Front. Plant Sci. 2017, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hameg, R.; Arteta, T.A.; Landin, M.; Gallego, P.P.; Barreal, M.E. Modeling and Optimizing Culture Medium Mineral Composition for in vitro Propagation of Actinidia arguta. Front. Plant Sci. 2020, 11, 11. [Google Scholar] [CrossRef]
- Nezami-Alanagh, E.; Garoosi, G.-A.; Landin, M.; Gallego, P.P. Computer-based tools provide new insight into the key factors that cause physiological disorders of pistachio rootstocks cultured in vitro. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Landin, M.; Rowe, R.C. Artificial neural networks technology to model, understand, and optimize drug formulations. In Formulation Tools for Pharmaceutical Development; Woodhead Publishing Limited: Cambridge, UK, 2013; pp. 7–37. ISBN 9781907568992. [Google Scholar]
- Gago, J.; Pérez-Tornero, O.; Landin, M.; Burgos, L.; Gallego, P.P. Improving knowledge of plant tissue culture and formulation bv neurofuzzy logic: A practical case of data mining using apricot databases. J. Plant Physiol. 2011, 168, 1858–1865. [Google Scholar] [CrossRef]
- García-Pérez, P.; Losada-Barreiro, S.; Gallego, P.P.; Bravo-Díaz, C. Cyclodextrin-elicited Bryophyllum suspension cultured cells: Enhancement of the production of bioactive compounds. Int. J. Mol. Sci. 2019, 20, 5180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and Human Health: The Role of Bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef]
- Efenberger-Szmechtyk, M.; Nowak, A.; Czyżowska, A. Plant extracts rich in polyphenols: Antibacterial agents and natural preservatives for meat and meat products. Crit. Rev. Food Sci. Nutr. 2021, 61, 149–178. [Google Scholar] [CrossRef]
- Chandran, H.; Meena, M.; Barupal, T.; Sharma, K. Plant tissue culture as a perpetual source for production of industrially important bioactive compounds. Biotechnol. Rep. 2020, 26, e00450. [Google Scholar] [CrossRef]
- Marchev, A.S.; Yordanova, Z.P.; Georgiev, M.I. Green (cell) factories for advanced production of plant secondary metabolites. Crit. Rev. Biotechnol. 2020, 40, 443–458. [Google Scholar] [CrossRef] [PubMed]
- García-Pérez, P.; Lozano-Milo, E.; Landin, M.; Gallego, P.P. Machine Learning Unmasked Nutritional Imbalances on the Medicinal Plant Bryophyllum sp. Cultured in vitro. Front. Plant Sci. 2020, 11, 576177. [Google Scholar] [CrossRef] [PubMed]
- Casanova, J.M.; Nascimento, L.B.D.S.; Casanova, L.M.; Leal-Costa, M.V.; Costa, S.S.; Tavares, E.S. Differential Distribution of Flavonoids and Phenolic Acids in Leaves of Kalanchoe delagoensis Ecklon & Zeyher (Crassulaceae). Microsc. Microanal. 2020, 26, 1061–1068. [Google Scholar] [CrossRef] [PubMed]
- Stefanowicz-Hajduk, J.; Hering, A.; Gucwa, M.; Hałasa, R.; Soluch, A.; Kowalczyk, M.; Stochmal, A.; Ochocka, R. Biological activities of leaf extracts from selected Kalanchoe species and their relationship with bufadienolides content. Pharm. Biol. 2020, 58, 732–740. [Google Scholar] [CrossRef]
- Gajula, S.N.R.; Nanjappan, S. Metabolomics: A Recent Advanced Omics Technology in Herbal Medicine Research; Elsevier BV: Amsterdam, The Netherlnds, 2020; pp. 97–117. [Google Scholar]
- García-Pérez, P.; Miras-Moreno, B.; Lucini, L.; Gallego, P.P. The metabolomics reveals intraspecies variability of bioactive compounds in elicited suspension cell cultures of three Bryophyllum species. Ind. Crop. Prod. 2021, 163, 113322. [Google Scholar] [CrossRef]
- Herrando-Moraira, S.; Vitales, D.; Nualart, N.; Gómez-Bellver, C.; Ibáñez, N.; Massó, S.; Cachón-Ferrero, P.; González-Gutiérrez, P.A.; Guillot, D.; Herrera, I.; et al. Global distribution patterns and niche modelling of the invasive Kalanchoe × houghtonii (Crassulaceae). Sci. Rep. 2020, 10, 1–18. [Google Scholar] [CrossRef] [Green Version]
- García-Pérez, P.; Lozano-Milo, E.; Landín, M.; Gallego, P.P. Machine Learning Technology Reveals the Concealed Interactions of Phytohormones on Medicinal Plant In Vitro Organogenesis. Biomolecules 2020, 10, 746. [Google Scholar] [CrossRef]
- Li, D.-D.; Ni, R.; Wang, P.-P.; Zhang, X.-S.; Wang, P.-Y.; Zhu, T.-T.; Sun, C.-J.; Liu, C.-J.; Lou, H.-X.; Cheng, A.-X. Molecular Basis for Chemical Evolution of Flavones to Flavonols and Anthocyanins in Land Plants. Plant Physiol. 2020, 184, 1731–1743. [Google Scholar] [CrossRef]
- Agati, G.; Brunetti, C.; Fini, A.; Gori, A.; Guidi, L.; Landi, M.; Sebastiani, F.; Tattini, M. Are Flavonoids Effective Antioxidants in Plants? Twenty Years of Our Investigation. Antioxidants 2020, 9, 1098. [Google Scholar] [CrossRef]
- Shah, A.; Smith, D.L. Flavonoids in Agriculture: Chemistry and Roles in, Biotic and Abiotic Stress Responses, and Microbial Associations. Agronomy 2020, 10, 1209. [Google Scholar] [CrossRef]
- García-Pérez, P.; Lozano-Milo, E.; Gallego, P.P.; Tojo, C.; Losada-Barreiro, S.; Bravo-Díaz, C. Plant antioxidants in food emul-sions. In Some New Aspects of Colloidal Systems in Foods; Milani, J., Ed.; IntechOpen: Rijeka, Croatia, 2019; pp. 11–29. [Google Scholar]
- Chernetskyy, M.; Woźniak, A.; Skalska-Kamińska, A.; Żuraw, B.; Blicharska, E.; Rejdak, R.; Donica, H.; Weryszko-Chmielewska, E. Structure of leaves and phenolic acids in Kalanchoë daigremontiana Raym.-Hamet & H. Perrier. Acta Sci. Pol. Hortorum Cultus 2018, 17, 137–155. [Google Scholar] [CrossRef]
- Shao, Q.; Rowe, R.C.; York, P. Comparison of neurofuzzy logic and neural networks in modelling experimental data of an immediate release tablet formulation. Eur. J. Pharm. Sci. 2006, 28, 394–404. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Hesami, M.; Naderi, R.; Tohidfar, M.; Yoosefzadeh-Najafabadi, M. Development of support vector machine-based model and comparative analysis with artificial neural network for modeling the plant tissue culture procedures: Effect of plant growth regulators on somatic embryogenesis of chrysanthemum, as a case study. Plant Methods 2020, 16, 1–15. [Google Scholar] [CrossRef]
- Nezami-Alanagh, E.; Garoosi, G.-A.; Maleki, S.; Landin, M.; Gallego, P.P. Predicting optimal in vitro culture medium for Pistacia vera micropropagation using neural networks models. Plant Cell Tissue Organ Cult. 2017, 129, 19–33. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Gallardo, A.; Bowker, M.A.; Wallenstein, M.; Quero, J.L.; Ochoa, V.; Gozalo, B.; García-Gómez, M.; Soliveres, S.; et al. Decoupling of soil nutrient cycles as a function of aridity in global drylands. Nat. Cell Biol. 2013, 502, 672–676. [Google Scholar] [CrossRef]
- Pereira, P.N.; Cushman, J.C. Exploring the Relationship between Crassulacean Acid Metabolism (CAM) and Mineral Nutrition with a Special Focus on Nitrogen. Int. J. Mol. Sci. 2019, 20, 4363. [Google Scholar] [CrossRef] [Green Version]
- Bertelli, M.; Kiani, A.K.; Paolacci, S.; Manara, E.; Kurti, D.; Dhuli, K.; Bushati, V.; Miertus, J.; Pangallo, D.; Baglivo, M.; et al. Hydroxytyrosol: A natural compound with promising pharmacological activities. J. Biotechnol. 2020, 309, 29–33. [Google Scholar] [CrossRef]
- Siatka, T.; Chlebekb, J.; Hostalkova, A. Copper(II) Sulfate Stimulates Scopoletin Production in Cell Suspension Cultures of Angelica archangelica. Nat. Prod. Commun. 2017, 12, 1779–1780. [Google Scholar] [CrossRef] [Green Version]
- Mosher, S.; Seybold, H.; Rodriguez, P.; Stahl, M.; Davies, K.A.; Dayaratne, S.; Morillo, S.A.; Wierzba, M.; Favery, B.; Keller, H.; et al. The tyrosine-sulfated peptide receptors PSKR1 and PSY1R modify the immunity of Arabidopsis to biotrophic and necrotrophic pathogens in an antagonistic manner. Plant J. 2013, 73, 469–482. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhu, J.; Shao, L.; Guo, M. Current advances in acteoside biosynthesis pathway elucidation and biosynthesis. Fitoterapia 2020, 142, 104495. [Google Scholar] [CrossRef]
- Chutia, R.; Abel, S.; Ziegler, J. Iron and Phosphate Deficiency Regulators Concertedly Control Coumarin Profiles in Arabidopsis thaliana Roots During Iron, Phosphate, and Combined Deficiencies. Front. Plant Sci. 2019, 10, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Yang, R.; Pan, Y.; Ren, B.; Chen, Q.; Li, X.; Xiong, X.; Tao, J.; Cheng, Q.; Ma, M. Beneficial behavior of nitric oxide in copper-treated medicinal plants. J. Hazard. Mater. 2016, 314, 140–154. [Google Scholar] [CrossRef]
- Sgherri, C.; Cosi, E.; Navari-Izzo, F. Phenols and antioxidative status of Raphanus sativus grown in copper excess. Physiol. Plant. 2003, 118, 21–28. [Google Scholar] [CrossRef]
- Kováčik, J.; Grúz, J.; Klejdus, B.; Štork, F.; Marchiosi, R.; Ferrarese-Filho, O. Lignification and related parameters in copper-exposed Matricaria chamomilla roots: Role of H2O2 and NO in this process. Plant Sci. 2010, 179, 383–389. [Google Scholar] [CrossRef]
- Fakhari, S.; Sharifi, M.; de Michele, R.; Ghanati, F.; Safaie, N.; Sadeghnezhad, E. Hydrogen sulfide directs metabolic flux towards the lignan biosynthesis in Linum album hairy roots. Plant Physiol. Biochem. 2019, 135, 359–371. [Google Scholar] [CrossRef]
- Causero, A.; Elsen, H.; Ballmann, G.; Escalona, A.; Harder, S. Calcium stilbene complexes: Structures and dual reactivity. Chem. Commun. 2017, 53, 10386–10389. [Google Scholar] [CrossRef]
- Aleynova, O.A.; Dubrovina, A.S.; Kiselev, K.V. Activation of stilbene synthesis in cell cultures of Vitis amurensis by calcium-dependent protein kinases VaCPK1 and VaCPK26. Plant Cell Tissue Organ Cult. 2017, 130, 141–152. [Google Scholar] [CrossRef]
- Martins, V.; Garcia, A.; Costa, C.; Sottomayor, M.; Gerós, H. Calcium- and hormone-driven regulation of secondary metabolism and cell wall enzymes in grape berry cells. J. Plant Physiol. 2018, 231, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Wei, K.; Wang, L. Tea Leaf Age, Shade and Characteristic Levels of l-Theanine, Caffeine, (-)-Epigallocatechin Gallate (EGCG), (-)-Epigallocatechin (EGC), (-)-Epicatechin (EC), and (-)-Epicatechin Gallate (ECG). In Processing and Impact on Active Components in Food; Hagiwara, K.A., Wright, A.D., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2015; pp. 591–596. ISBN 9780124047099. [Google Scholar]
- Jayaganesh, S.; Venkatesan, S. Impact of Magnesium Sulphate on Biochemical and Quality Constituents of Black Tea. Am. J. Food Technol. 2009, 5, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Yu, T.; Zhou, B.; Wei, J.; Fang, Y.; Lu, J.; Guo, L.; Chen, W.; Liu, Z.-P.; Luo, J. Mg(II)-Catechin nanoparticles delivering siRNA targeting EIF5A2 inhibit bladder cancer cell growth in vitro and in vivo. Biomaterials 2016, 81, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Walker, T.S.; Bais, H.P.; Grotewold, E.; Vivanco, J.M. Root Exudation and Rhizosphere Biology. Plant Physiol. 2003, 132, 44–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerra-García, A.; Golubov, J.; Mandujano, M.C. Invasion of Kalanchoe by clonal spread. Biol. Invasions 2014, 17, 1615–1622. [Google Scholar] [CrossRef]
- García-Pérez, P.; Losada-Barreiro, S.; Gallego, P.P.; Bravo-Díaz, C. Adsorption of gallic acid, propyl gallate and polyphenols from Bryophyllum extracts on activated carbon. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Arriola, N.D.A.; Chater, P.I.; Wilcox, M.; Lucini, L.; Rocchetti, G.; Dalmina, M.; Pearson, J.P.; Amboni, R.D.D.M.C. Encapsulation of stevia rebaudiana Bertoni aqueous crude extracts by ionic gelation—Effects of alginate blends and gelling solutions on the polyphenolic profile. Food Chem. 2019, 275, 123–134. [Google Scholar] [CrossRef] [Green Version]
- Rocchetti, G.; Pagnossa, J.P.; Blasi, F.; Cossignani, L.; Piccoli, R.H.; Zengin, G.; Montesano, D.; Cocconcelli, P.S.; Lucini, L. Phenolic profiling and in vitro bioactivity of Moringa oleifera leaves as affected by different extraction solvents. Food Res. Int. 2020, 127, 108712. [Google Scholar] [CrossRef]
- Rocchetti, G.; Michelini, S.; Pizzamiglio, V.; Masoero, F.; Lucini, L. A combined metabolomics and peptidomics approach to discriminate anomalous rind inclusion levels in Parmigiano Reggiano PDO grated hard cheese from different ripening stages. Food Res. Int. 2021, 149, 110654. [Google Scholar] [CrossRef]
- Rocchetti, G.; Lucini, L.; Rodriguez, J.M.L.; Barba, F.J.; Giuberti, G. Gluten-free flours from cereals, pseudocereals and legumes: Phenolic fingerprints and in vitro antioxidant properties. Food Chem. 2019, 271, 157–164. [Google Scholar] [CrossRef]
- Rothwell, J.A.; Pérez-Jiménez, J.; Neveu, V.; Medina-Remón, A.; M’Hiri, N.; García-Lobato, P.; Manach, C.; Knox, C.; Eisner, R.; Wishart, D.S.; et al. Phenol-Explorer 3.0: A major update of the Phenol-Explorer database to incorporate data on the effects of food processing on polyphenol content. Database 2013, 2013, bat070. [Google Scholar] [CrossRef]
- Lucini, L.; Rocchetti, G.; Kane, D.; Trevisan, M. Phenolic fingerprint allows discriminating processed tomato products and tracing different processing sites. Food Control. 2017, 73, 696–703. [Google Scholar] [CrossRef]
- Salek, R.M.; Steinbeck, C.; Viant, M.R.; Goodacre, R.; Dunn, W.B. The role of reporting standards for metabolite annotation and identification in metabolomic studies. GigaScience 2013, 2, 1–3. [Google Scholar] [CrossRef]
- Zhang, L.; Saber, F.R.; Rocchetti, G.; Zengin, G.; Hashem, M.M.; Lucini, L. UHPLC-QTOF-MS based metabolomics and biological activities of different parts of Eriobotrya japonica. Food Res. Int. 2021, 143, 110242. [Google Scholar] [CrossRef] [PubMed]
- Niedz, R.P.; Evens, T.J. A solution to the problem of ion confounding in experimental biology. Nat. Methods 2006, 3, 417. [Google Scholar] [CrossRef] [PubMed]
- Kavli, T.; Weyer, E. ASMOD (Adaptive Spline Modelling of Observation Data): Some theoretical and experimental results. In Proceedings of the IEE Colloquium on Advances in Neural Networks for Control and Systems, Berlin, Germany, 25–27 May 1994; pp. 3/1–3/7. [Google Scholar]
- Arteta, T.; Hameg, R.; Landin, M.; Gallego, P.P.; Barreal, M.E. Neural networks models as decision-making tool for in vitro proliferation of hardy kiwi. Eur. J. Hortic. Sci. 2018, 83, 259–265. [Google Scholar] [CrossRef]
- Colbourn, E.; Rowe, R. Encyclopaedia of Pharmaceutical Technology; Marcel Dekker: New York, NY, USA, 2005. [Google Scholar]
- Vapnik, V. Principles of Risk Minimization for Learning Theory. In Advances in Neural Information Processing Systems; Morgan Kaufmann: Burlington, MA, USA, 1992; Volume 4, pp. 831–838. [Google Scholar]
- Colbourn, E.A.; Rowe, R.C. Novel approaches to neural and evolutionary computing in pharmaceutical formulation: Challenges and new possibilities. Future Med. Chem. 2009, 1, 713–726. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Output | Submodel | Significant Inputs | Training set R2 | MSE | F Ratio | df1, df2 | f Critical (α = 0.05) |
|---|---|---|---|---|---|---|---|
| LMW | 1 | Organ × SO42− | 71.08 | 11.08 | 3.47 | 17, 24 | 2.07 |
| 2 | Genotype × Cu2+ | ||||||
| 3 | HPO42− | ||||||
| Phenolic acids | 1 | Organ | 72.12 | 5.52 | 8.02 | 10, 31 | 2.15 |
| 2 | Genotype × Cu2+ | ||||||
| Lignans | 1 | Genotype × SO42− × Organ | 73.32 | 5.19 | 6.64 | 12, 29 | 2.10 |
| Stilbenes | 1 | Ca2+ × Organ × Genotype | 94.94 | 0.29 | 7.51 | 29, 12 | 2.47 |
| 2 | Genotype × HPO42− × Organ | ||||||
| Flavones | - | - | 68.77 | 2.71 | 12.84 | 6, 35 | 2.37 |
| Flavonols | 1 | Organ × Genotype | 74.49 | 0.41 | 17.03 | 6, 35 | 2.37 |
| Anthocyanins | 1 | Organ × Genotype | 77.15 | 0.51 | 19.70 | 6, 35 | 2.37 |
| Flavanols | 1 | Mg2+ × Organ | 78.04 | 1.16 | 12.63 | 9, 32 | 2.19 |
| 2 | Organ × Genotype |
| Rules | Gen 1 | Organ 2 | Ca2+ 3 | Mg2+ 3 | SO42− 3 | HPO42− 3 | Cu2+ 3 | LMW | Phenolic Acids | Lignans | Stilbenes | Flavonols | Anthocyanins | Flavanols | MD 4 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | IF | A | LOW | THEN | LOW | 1.00 | |||||||||||
| 2 | A | MID LOW | LOW | 1.00 | |||||||||||||
| 3 | A | MID HIGH | HIGH | 1.00 | |||||||||||||
| 4 | A | HIGH | HIGH | 1.00 | |||||||||||||
| 5 | R | LOW | LOW | 1.00 | |||||||||||||
| 6 | R | MID LOW | LOW | 1.00 | |||||||||||||
| 7 | R | MID HIGH | HIGH | 1.00 | |||||||||||||
| 8 | R | HIGH | HIGH | 1.00 | |||||||||||||
| 9 | BD | LOW | HIGH | 1.00 | |||||||||||||
| 10 | BD | MID | LOW | 1.00 | |||||||||||||
| 11 | BD | HIGH | LOW | 1.00 | |||||||||||||
| 12 | BH | LOW | HIGH | 1.00 | |||||||||||||
| 13 | BH | MID | HIGH | 0.67 | |||||||||||||
| 14 | BH | HIGH | LOW | 1.00 | |||||||||||||
| 15 | BT | LOW | HIGH | 1.00 | |||||||||||||
| 16 | BT | MID | LOW | 0.59 | |||||||||||||
| 17 | BT | HIGH | LOW | 1.00 | |||||||||||||
| 18 | LOW | HIGH | 1.00 | ||||||||||||||
| 19 | HIGH | LOW | 1.00 | ||||||||||||||
| 20 | IF | A | THEN | HIGH | 0.99 | ||||||||||||
| 21 | R | LOW | 0.65 | ||||||||||||||
| 22 | BD | LOW | LOW | 0.62 | |||||||||||||
| 23 | BD | MID | LOW | 1.00 | |||||||||||||
| 24 | BD | HIGH | HIGH | 0.58 | |||||||||||||
| 25 | BH | LOW | LOW | 0.84 | |||||||||||||
| 26 | BH | MID | LOW | 0.52 | |||||||||||||
| 27 | BH | HIGH | LOW | 0.75 | |||||||||||||
| 28 | BT | LOW | LOW | 1.00 | |||||||||||||
| 29 | BT | MID | LOW | 1.00 | |||||||||||||
| 30 | BT | HIGH | LOW | 1.00 | |||||||||||||
| 31 | IF | BD | A | LOW | THEN | LOW | 0.60 | ||||||||||
| 32 | BD | R | LOW | LOW | 0.80 | ||||||||||||
| 33 | BD | A | HIGH | LOW | 0.88 | ||||||||||||
| 34 | BD | R | HIGH | LOW | 0.69 | ||||||||||||
| 35 | BH | A | LOW | LOW | 0.74 | ||||||||||||
| 36 | BH | R | LOW | LOW | 0.84 | ||||||||||||
| 37 | BH | A | HIGH | HIGH | 0.84 | ||||||||||||
| 38 | BH | R | HIGH | HIGH | 0.51 | ||||||||||||
| 39 | BT | A | LOW | LOW | 0.77 | ||||||||||||
| 40 | BT | R | LOW | LOW | 0.90 | ||||||||||||
| 41 | BT | A | HIGH | LOW | 0.80 | ||||||||||||
| 42 | BT | R | HIGH | LOW | 0.73 | ||||||||||||
| 43 | IF | BD | A | LOW | THEN | LOW | 1.00 | ||||||||||
| 44 | BH | A | LOW | HIGH | 1.00 | ||||||||||||
| 45 | BT | A | LOW | LOW | 1.00 | ||||||||||||
| 46 | BD | R | LOW | LOW | 1.00 | ||||||||||||
| 47 | BH | R | LOW | HIGH | 1.00 | ||||||||||||
| 48 | BT | R | LOW | LOW | 1.00 | ||||||||||||
| 49 | BD | A | HIGH | HIGH | 1.00 | ||||||||||||
| 50 | BH | A | HIGH | LOW | 1.00 | ||||||||||||
| 51 | BT | A | HIGH | HIGH | 1.00 | ||||||||||||
| 52 | BD | R | HIGH | HIGH | 0.65 | ||||||||||||
| 53 | BH | R | HIGH | LOW | 1.00 | ||||||||||||
| 54 | BT | R | HIGH | HIGH | 1.00 | ||||||||||||
| 55 | BD | A | LOW | HIGH | 1.00 | ||||||||||||
| 56 | BD | R | LOW | HIGH | 0.70 | ||||||||||||
| 57 | BD | A | MID | LOW | 1.00 | ||||||||||||
| 58 | BD | R | MID | LOW | 0.81 | ||||||||||||
| 59 | BD | A | HIGH | LOW | 1.00 | ||||||||||||
| 60 | BD | R | HIGH | LOW | 1.00 | ||||||||||||
| 61 | BH | A | LOW | LOW | 1.00 | ||||||||||||
| 62 | BH | R | LOW | LOW | 1.00 | ||||||||||||
| 63 | BH | A | MID | HIGH | 1.00 | ||||||||||||
| 64 | BH | R | MID | LOW | 0.52 | ||||||||||||
| 65 | BH | A | HIGH | HIGH | 1.00 | ||||||||||||
| 66 | BH | R | HIGH | HIGH | 1.00 | ||||||||||||
| 67 | BT | A | LOW | HIGH | 1.00 | ||||||||||||
| 68 | BT | R | LOW | HIGH | 1.00 | ||||||||||||
| 69 | BT | A | MID | LOW | 0.85 | ||||||||||||
| 70 | BT | R | MID | LOW | 0.93 | ||||||||||||
| 71 | BT | A | HIGH | LOW | 1.00 | ||||||||||||
| 72 | BT | R | HIGH | LOW | 1.00 | ||||||||||||
| 73 | IF | BD | A | THEN | HIGH | 0.76 | |||||||||||
| 74 | BD | R | LOW | 0.83 | |||||||||||||
| 75 | BH | A | LOW | 0.88 | |||||||||||||
| 76 | BH | R | LOW | 0.86 | |||||||||||||
| 77 | BT | A | LOW | 0.92 | |||||||||||||
| 78 | BT | R | LOW | 0.94 | |||||||||||||
| 79 | IF | BD | A | THEN | HIGH | 0.78 | |||||||||||
| 80 | BH | A | HIGH | 0.57 | |||||||||||||
| 81 | BT | A | LOW | 0.72 | |||||||||||||
| 82 | BD | R | LOW | 0.84 | |||||||||||||
| 83 | BH | R | LOW | 0.84 | |||||||||||||
| 84 | BT | R | LOW | 0.96 | |||||||||||||
| 85 | IF | A | LOW | THEN | LOW | 0.71 | |||||||||||
| 86 | R | LOW | HIGH | 0.74 | |||||||||||||
| 87 | A | HIGH | LOW | 0.76 | |||||||||||||
| 88 | R | HIGH | LOW | 0.86 | |||||||||||||
| 89 | BD | A | HIGH | 0.74 | |||||||||||||
| 90 | BD | R | LOW | 0.98 | |||||||||||||
| 91 | BH | A | LOW | 1.00 | |||||||||||||
| 92 | BH | R | LOW | 0.90 | |||||||||||||
| 93 | BT | A | LOW | 1.00 | |||||||||||||
| 94 | BT | R | HIGH | 0.76 |
| Salts (mg L−1) | MS Control | 1/2MSM | 1/4MSM | 1/8MSM | 1/2MSµ | 1/4MSµ | 1/8MSµ | |
|---|---|---|---|---|---|---|---|---|
| Macro-nutrients | KNO3 | 1900 | 950 | 475 | 237.5 | 1900 | 1900 | 1900 |
| NH4NO3 | 1650 | 825 | 412.5 | 206.3 | 1650 | 1650 | 1650 | |
| CaCl2 2H2O | 440 | 220 | 110 | 55 | 440 | 440 | 440 | |
| MgSO4 7H2O | 370 | 185 | 92.5 | 46.3 | 370 | 370 | 370 | |
| KH2PO4 | 170 | 85 | 42.5 | 21.3 | 170 | 170 | 170 | |
| Micronutrients | MnSO4 4H2O | 22.3 | 22.3 | 22.3 | 22.3 | 11.2 | 5.6 | 2.8 |
| ZnSO4 7H2O | 8.6 | 8.6 | 8.6 | 8.6 | 4.3 | 2.2 | 1.1 | |
| H3BO3 | 6.2 | 6.2 | 6.2 | 6.2 | 3.1 | 1.6 | 0.78 | |
| KI | 0.83 | 0.83 | 0.83 | 0.83 | 0.42 | 0.21 | 0.11 | |
| Na2MoO4 2H2O | 0.25 | 0.25 | 0.25 | 0.25 | 0.13 | 0.063 | 0.03 | |
| CuSO4 5H2O | 0.025 | 0.025 | 0.025 | 0.025 | 0.013 | 0.0063 | 0.0031 | |
| CoCl2 6H2O | 0.025 | 0.025 | 0.025 | 0.025 | 0.013 | 0.0063 | 0.0031 | |
| Iron source | Na2EDTA | 37.25 | 37.25 | 37.25 | 37.25 | 37.25 | 37.25 | 37.25 |
| FeSO4 7H2O | 27.85 | 27.85 | 27.85 | 27.85 | 27.85 | 27.85 | 27.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Pérez, P.; Zhang, L.; Miras-Moreno, B.; Lozano-Milo, E.; Landin, M.; Lucini, L.; Gallego, P.P. The Combination of Untargeted Metabolomics and Machine Learning Predicts the Biosynthesis of Phenolic Compounds in Bryophyllum Medicinal Plants (Genus Kalanchoe). Plants 2021, 10, 2430. https://doi.org/10.3390/plants10112430
García-Pérez P, Zhang L, Miras-Moreno B, Lozano-Milo E, Landin M, Lucini L, Gallego PP. The Combination of Untargeted Metabolomics and Machine Learning Predicts the Biosynthesis of Phenolic Compounds in Bryophyllum Medicinal Plants (Genus Kalanchoe). Plants. 2021; 10(11):2430. https://doi.org/10.3390/plants10112430
Chicago/Turabian StyleGarcía-Pérez, Pascual, Leilei Zhang, Begoña Miras-Moreno, Eva Lozano-Milo, Mariana Landin, Luigi Lucini, and Pedro P. Gallego. 2021. "The Combination of Untargeted Metabolomics and Machine Learning Predicts the Biosynthesis of Phenolic Compounds in Bryophyllum Medicinal Plants (Genus Kalanchoe)" Plants 10, no. 11: 2430. https://doi.org/10.3390/plants10112430
APA StyleGarcía-Pérez, P., Zhang, L., Miras-Moreno, B., Lozano-Milo, E., Landin, M., Lucini, L., & Gallego, P. P. (2021). The Combination of Untargeted Metabolomics and Machine Learning Predicts the Biosynthesis of Phenolic Compounds in Bryophyllum Medicinal Plants (Genus Kalanchoe). Plants, 10(11), 2430. https://doi.org/10.3390/plants10112430