
New Species of Chloroidium (Trebouxiophyceae, Chlorophyta) from East Asia


 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
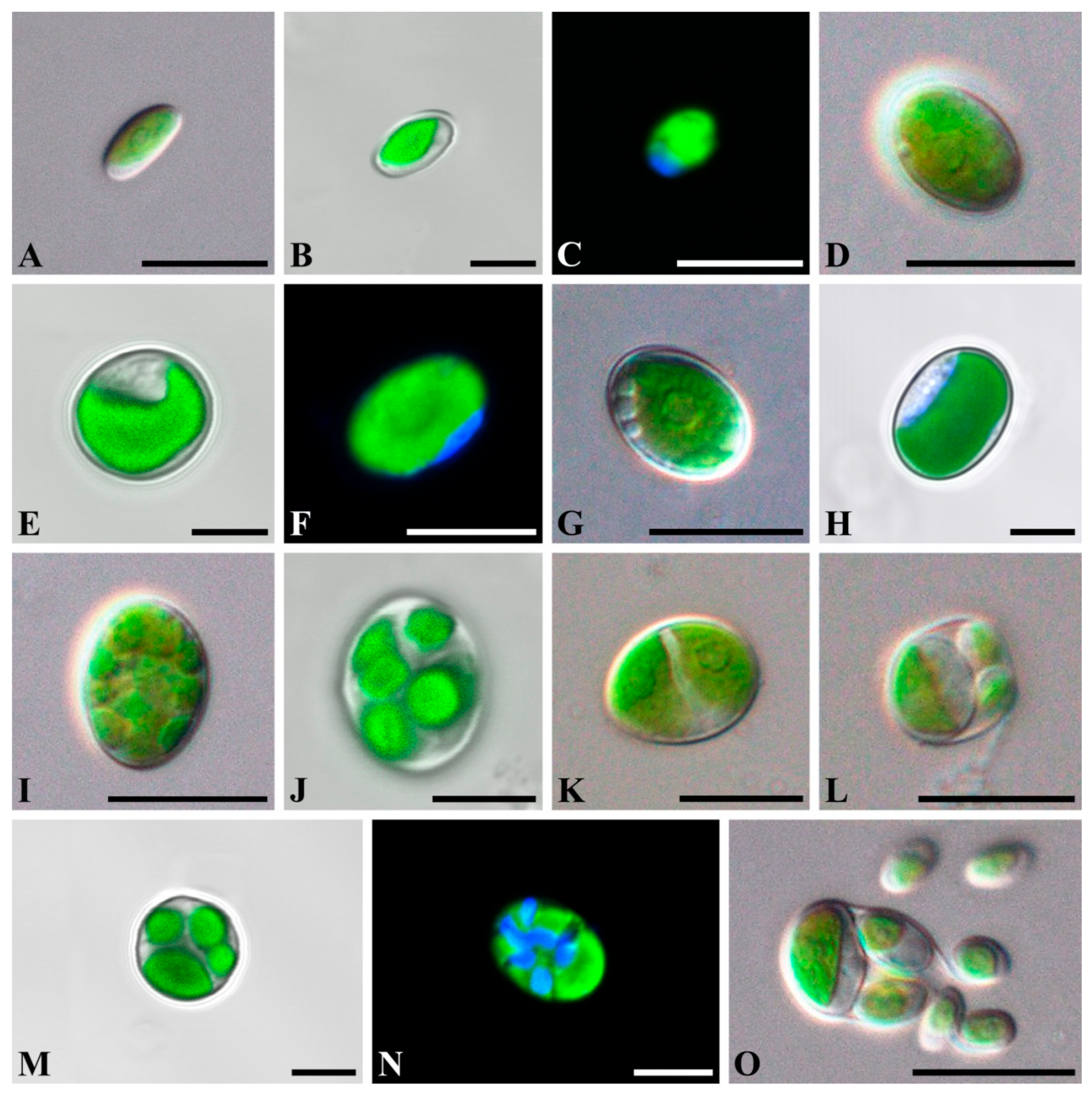
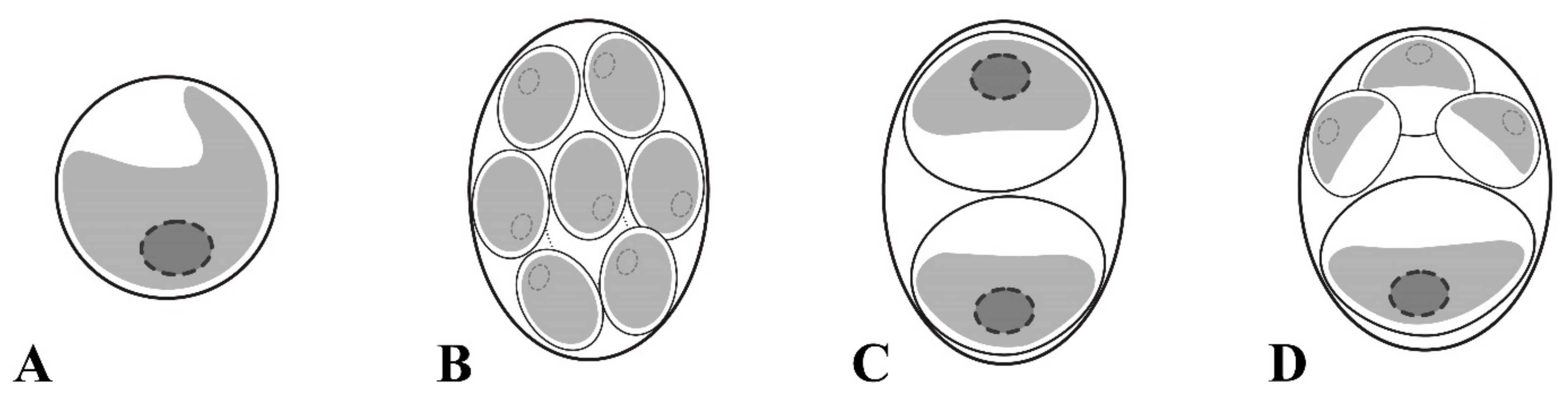
2.1. Taxonomic Treatment
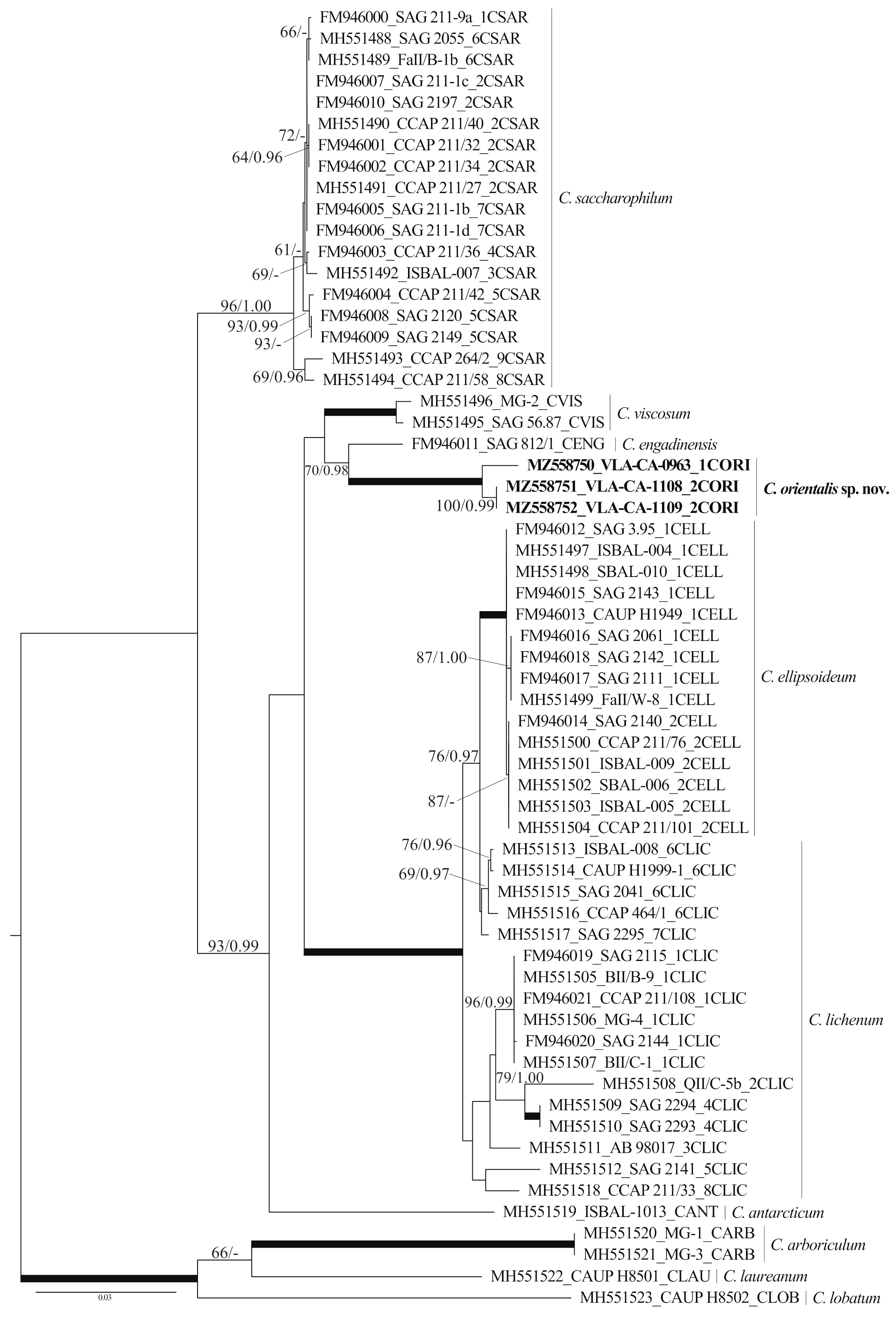
2.2. Phylogenetic Analyses
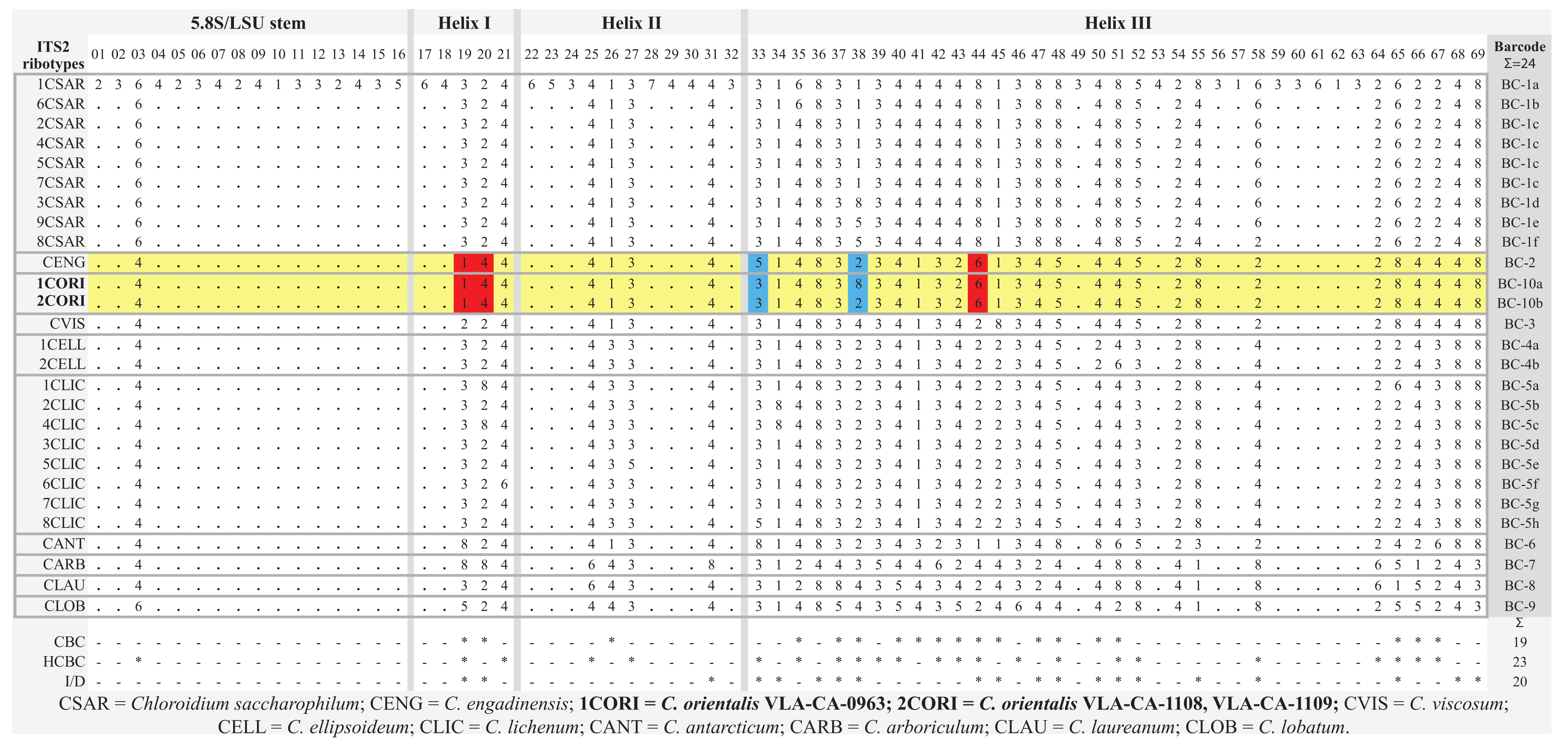
2.3. ITS2 Secondary Structure
3. Discussion
4. Materials and Methods
4.1. Strain Origin, Culture Conditions and Light Microscopy
4.2. DNA Extraction, Amplification and Sequencing
4.3. Phylogenetic Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huss, V.A.R.; Frank, C.; Hartmann, E.C.; Hirmer, M.; Kloboucek, A.; Seidel, B.M.; Wenzeler, P.; Kessler, E. Biochemical taxonomy and molecular phylogeny of the genus Chlorella sensu lato (Chlorophyta). J. Phycol. 1999, 35, 587–598. [Google Scholar] [CrossRef] [Green Version]
- Pröschold, T.; Leliaert, F. Systematics of the green algae: Conflict of classic and modern approaches. In Unravelling the Algae: The Past, Present, and Future of Algal Systematics; Brodie, J., Lewis, J., Eds.; CRC Press: Boca Raton, FL, USA, 2007; ISBN 978-0-8493-7989-5. [Google Scholar]
- Krienitz, L.; Bock, C. Present state of the systematics of planktonic coccoid green algae of inland waters. Hydrobiologia 2012, 698, 295–326. [Google Scholar] [CrossRef]
- Leliaert, F.; Smith, D.; Moreau, H.; Herron, M.; Verbruggen, H.; Delwiche, C.; De Clerck, O. Phylogeny and Molecular Evolution of the Green Algae. Crit. Rev. Plant Sci. 2012, 31, 1–46. [Google Scholar] [CrossRef] [Green Version]
- Fučíková, K.; Lewis, P.O.; Lewis, L. Widespread desert affiliation of trebouxiophycean algae (Trebouxiophyceae, Chlorophyta) including discovery of three new desert genera. Phycol. Res. 2014, 62, 294–305. [Google Scholar] [CrossRef]
- Lemieux, C.; Otis, C.; Turmel, M. Chloroplast phylogenomic analysis resolves deep-level relationships within the green algal class Trebouxiophyceae. BMC Evol. Biol. 2014, 14, 211. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Tan, H.; Liu, B.; Zhu, H.; Hu, Z.; Liu, G. Watanabeales ord. nov. and twelve novel species of Trebouxiophyceae (Chlorophyta). J. Phycol. 2021, 57, 1167–1186. [Google Scholar] [CrossRef]
- Darienko, T.; Gustavs, L.; Mudimu, O.; Menéndez, C.R.; Schumann, R.; Karsten, U.; Friedl, T.; Pröschold, T. Chloroidium, a common terrestrial coccoid green alga previously assigned toChlorella(Trebouxiophyceae, Chlorophyta). Eur. J. Phycol. 2010, 45, 79–95. [Google Scholar] [CrossRef] [Green Version]
- Darienko, T.; Lukešová, A.; Pröschold, T. The polyphasic approach revealed new species of Chloroidium (Trebouxiophyceae, Chlorophyta). Phytotaxa 2018, 372, 51–66. [Google Scholar] [CrossRef]
- Zhu, H.; Li, S.; Hu, Z.; Liu, G. Molecular characterization of eukaryotic algal communities in the tropical phyllosphere based on real-time sequencing of the 18S rDNA gene. BMC Plant Biol. 2018, 18, 365. [Google Scholar] [CrossRef]
- Darienko, T.; Pröschold, T. Reevaluation and discovery of new species of the rare genus Watanabea and establishment of Massjukichlorella gen. nov. (Trebouxiophyceae, Chlorophyta) using an integrative approach. J. Phycol. 2019, 55, 493–499. [Google Scholar] [CrossRef]
- Li, S.; Sun, H.; Hu, Y.; Liu, B.; Zhu, H.; Hu, Z.; Liu, G. Four New Members of Foliicolous Green Algae Within the Watanabea Clade (Trebouxiophyceae, Chlorophyta) from China. J. Eukaryot. Microbiol. 2020, 67, 369–382. [Google Scholar] [CrossRef]
- Darienko, T.; Gustavs, L.; Pröschold, T. Species concept and nomenclatural changes within the genera Elliptochloris and Pseudochlorella (Trebouxiophyceae) based on an integrative approach. J. Phycol. 2016, 52, 1125–1145. [Google Scholar] [CrossRef]
- Neustupa, J.; Němcová, Y.; Veselá, J.; Steinová, J.; Skaloud, P.; Kulichová, J. Parachloroidium gen. nov. (Trebouxiophyceae, Chlorophyta), a novel genus of coccoid green algae from subaerial corticolous biofilms. Phycologia 2013, 52, 411–421. [Google Scholar] [CrossRef] [Green Version]
- Kuzyakhmetov, G.G.; Dubovik, I.E. Metody Izucheniya Pochvennyh Vodorosley [Methods for Studying Soil Algae]; Izdatelstvo RIO BashGU: Ufa, Russia, 2001. (In Russian) [Google Scholar]
- Andersen, R.A. Algal Culturing Techniques; Elsevier Academic Press: Burlington, MA, USA, 2005; ISBN 0-12-088426-7. [Google Scholar]
- McFadden, G.I.; Melkonian, M. Use of Hepes buffer for microalgal culture media and fixation for electron microscopy. Phycologia 1986, 25, 551–557. [Google Scholar] [CrossRef]
- Echt, C.S.; Erdahl, L.A.; McCoy, T.J. Genetic segregation of random amplified polymorphic DNA in diploid cultivated alfalfa. Genome 1992, 35, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Kiselev, K.; Dubrovina, A.; Tyunin, A. The methylation status of plant genomic DNA influences PCR efficiency. J. Plant Physiol. 2015, 175, 59–67. [Google Scholar] [CrossRef]
- Lopez-Garcia, P.; Philippe, H.; Gail, F.; Moreira, D. Autochthonous eukaryotic diversity in hydrothermal sediment and experimental microcolonizers at the Mid-Atlantic Ridge. Proc. Natl. Acad. Sci. USA 2003, 100, 697–702. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Elwood, H.J.; Olsen, G.J.; Sogin, M.L. The small-subunit ribosomal RNA gene sequences from the hypotrichous ciliates Oxytricha nova and Stylonychia pustulata. Mol. Biol. Evol. 1985, 2, 399–410. [Google Scholar] [CrossRef] [Green Version]
- Marin, B.; Palm, A.; Klingberg, M.; Melkonian, M. Phylogeny and Taxonomic Revision of Plastid-Containing Euglenophytes based on SSU rDNA Sequence Comparisons and Synapomorphic Signatures in the SSU rRNA Secondary Structure. Protist 2003, 154, 99–145. [Google Scholar] [CrossRef]
- Goka, K.; Yokoyama, J.; Une, Y.; Kuroki, T.; Suzuki, K.; Nakahara, M.; Kobayashi, A.; Inaba, S.; Mizutani, T.; Hyatt, A.D. Amphibian chytridiomycosis in Japan: Distribution, haplotypes and possible route of entry into Japan. Mol. Ecol. 2009, 18, 4757–4774. [Google Scholar] [CrossRef]
- Bonfield, J.; Smith, K.F.; Staden, R. A new DNA sequence assembly program. Nucleic Acids Res. 1995, 23, 4992–4999. [Google Scholar] [CrossRef] [Green Version]
- Galtier, N.; Gouy, M.; Gautier, C. SEAVIEW and PHYLO_WIN: Two graphic tools for sequence alignment and molecular phylogeny. Bioinformatics 1996, 12, 543–548. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods), Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A Rapid Bootstrap Algorithm for the RAxML Web Servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [Green Version]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| № | Species | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | C. saccharophilum | 0.0166 | 0.0157 | 0.0139 | 0.0147 | 0.0127 | 0.0135 | 0.0149 | 0.0130 | 0.0165 | |
| 2 | C. laureanum | 0.2402 | 0.0162 | 0.0174 | 0.0168 | 0.0174 | 0.0191 | 0.0176 | 0.0165 | 0.0154 | |
| 3 | C. lobatum | 0.2443 | 0.2520 | 0.0173 | 0.0175 | 0.0165 | 0.0166 | 0.0162 | 0.0159 | 0.0181 | |
| 4 | C. viscosum | 0.1573 | 0.2555 | 0.2707 | 0.0148 | 0.9621 | 0.0112 | 0.0133 | 0.0132 | 0.0165 | |
| 5 | C. antarcticum | 0.1859 | 0.2778 | 0.2953 | 0.1725 | 0.0145 | 0.0142 | 0.0175 | 0.0167 | 0.0154 | |
| 6 | C. engadinensis | 0.1494 | 0.2639 | 0.2771 | 0.0834 | 0.1612 | 0.0118 | 0.0139 | 0.0135 | 0.0160 | |
| 7 | C. orientalis | 0.1797 | 0.2666 | 0.2927 | 0.1277 | 0.1879 | 0.1153 | 0.0146 | 0.0138 | 0.0159 | |
| 8 | C. ellipsoideum | 0.1630 | 0.2274 | 0.2703 | 0.1376 | 0.1751 | 0.1469 | 0.1582 | 0.0067 | 0.0169 | |
| 9 | C. lichenum | 0.1490 | 0.2250 | 0.2694 | 0.1387 | 0.1786 | 0.1443 | 0.1418 | 0.0505 | 0.0166 | |
| 10 | C. arboriculum | 0.2434 | 0.2234 | 0.2601 | 0.2718 | 0.2806 | 0.2720 | 0.2694 | 0.2617 | 0.2450 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gontcharov, A.A.; Nikulin, A.Y.; Nikulin, V.Y.; Bagmet, V.B.; Allaguvatova, R.Z.; Abdullin, S.R. New Species of Chloroidium (Trebouxiophyceae, Chlorophyta) from East Asia. Plants 2021, 10, 2560. https://doi.org/10.3390/plants10122560
Gontcharov AA, Nikulin AY, Nikulin VY, Bagmet VB, Allaguvatova RZ, Abdullin SR. New Species of Chloroidium (Trebouxiophyceae, Chlorophyta) from East Asia. Plants. 2021; 10(12):2560. https://doi.org/10.3390/plants10122560
Chicago/Turabian StyleGontcharov, Andrey A., Arthur Yu. Nikulin, Vyacheslav Yu. Nikulin, Veronika B. Bagmet, Rezeda Z. Allaguvatova, and Shamil R. Abdullin. 2021. "New Species of Chloroidium (Trebouxiophyceae, Chlorophyta) from East Asia" Plants 10, no. 12: 2560. https://doi.org/10.3390/plants10122560
APA StyleGontcharov, A. A., Nikulin, A. Y., Nikulin, V. Y., Bagmet, V. B., Allaguvatova, R. Z., & Abdullin, S. R. (2021). New Species of Chloroidium (Trebouxiophyceae, Chlorophyta) from East Asia. Plants, 10(12), 2560. https://doi.org/10.3390/plants10122560