
Bacterial Endophytes as a Promising Approach to Enhance the Growth and Accumulation of Bioactive Metabolites of Three Species of Chenopodium Sprouts


 and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Characterization of the Isolated Endophytic Bacteria
2.2. Selection of the Most Active Endophytic Bacteria
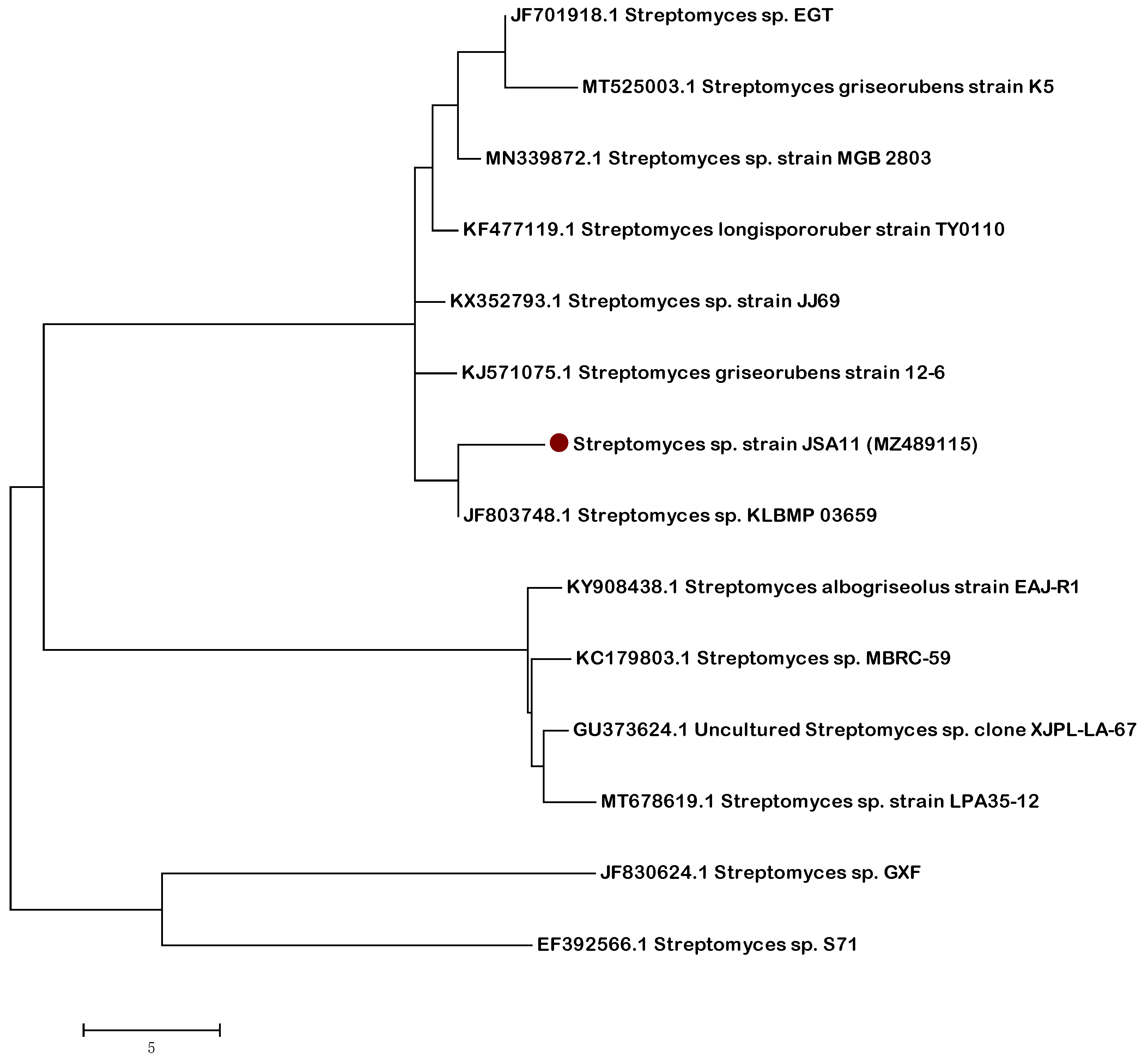
2.3. Molecular Characterization of the Most Active Isolate
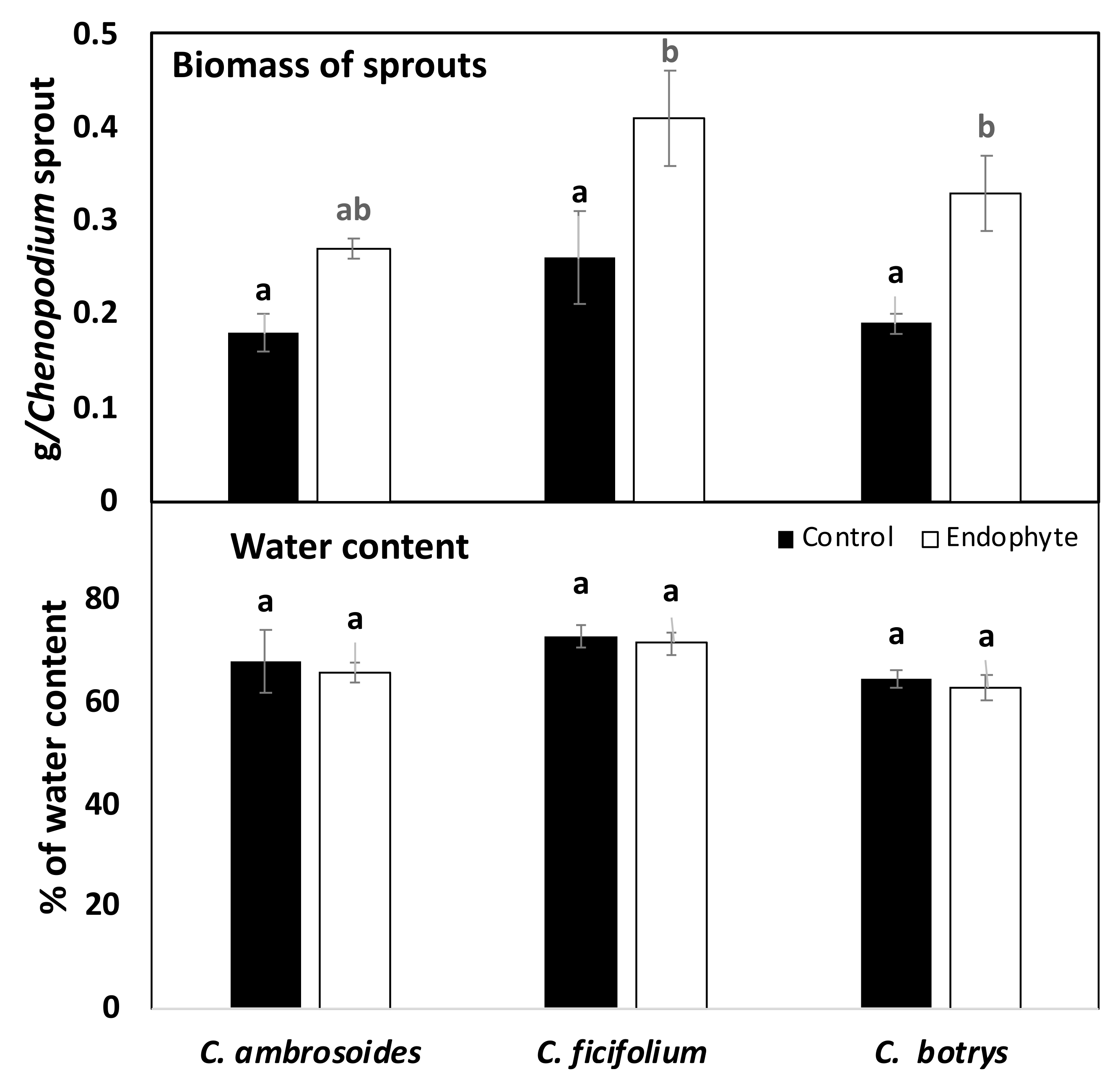
2.4. Bacterial Endophytes Promoted Photosynthesis and Biomass Production of Chenopodium Sprouts
2.5. Improved Minerals and Vitamins Contents by Endophytic Bacterial Treatment Contribute to Enhancing the Nutritive Value of Chenopodium Sprouts
2.6. Endophytic Bacterial Treatment Improved the Functional Food Value of Chenopodium Sprouts through Enhancing Their Bioactive Primary Metabolites Levels
2.7. Endophytic Bacteria Stimulated the Antioxidant Activities of Chenopodium Sprouts through Enhancing Their Phenolic Content
2.8. Endophytic Bacteria Enhanced the Chenopodium Biological Activities
2.8.1. Antibacterial Activities
2.8.2. Anti-Inflammatory Activities of Chenopodium Sprouts
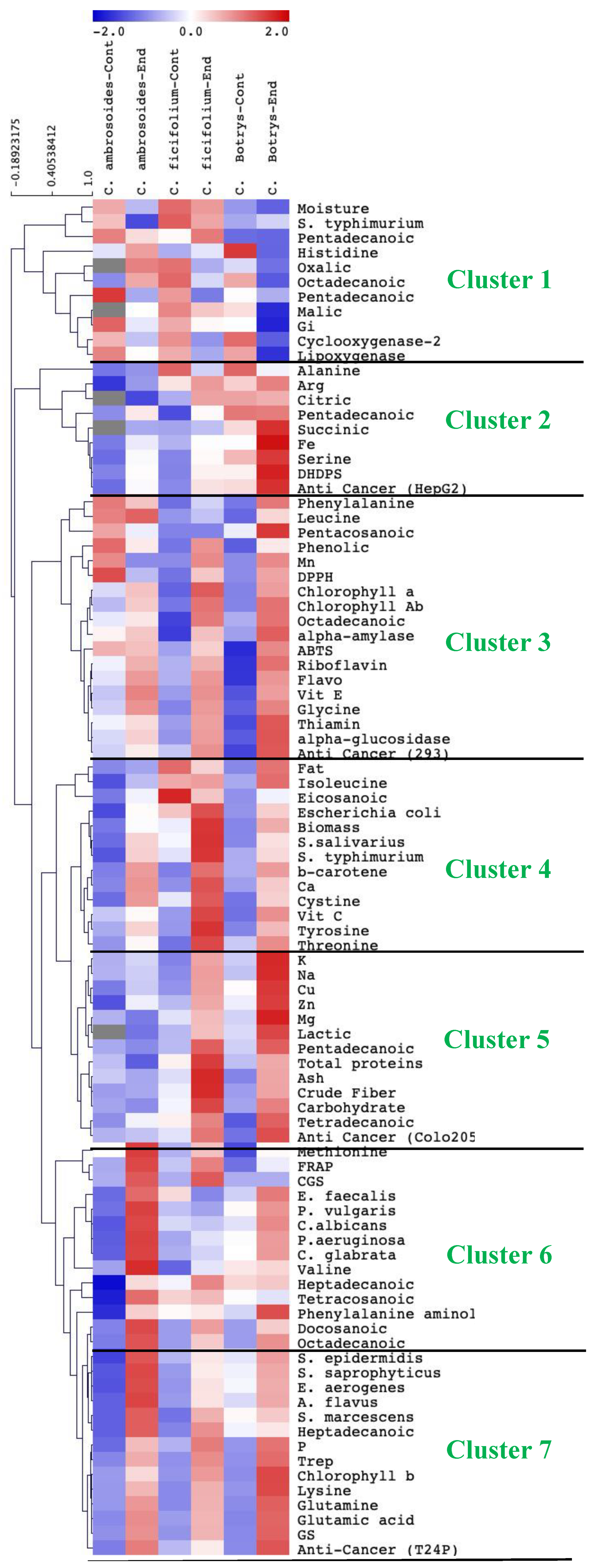
2.9. Species-Specific Response to Endophytic Bacterial Treatment
3. Material and Methods
3.1. Experimental Setup, Plant Materials, and Growth Conditions
3.1.1. Isolation and Characterization of Endophyte Isolates
3.1.2. Extraction of DNA
3.1.3. Seed Inoculation with Bacterial Endophyte and Growth of Chenopodium Sprouts
3.2. Determination of Photosynthetic Rate
3.3. Pigment Analysis
3.4. Determination of the Nutritional Value
3.4.1. Proximate Composition Analysis
3.4.2. Elemental Analysis
3.4.3. Amino Acids Levels and Metabolism
3.4.4. Organic Acid Analysis
3.4.5. Fatty Acids Analysis
3.4.6. Vitamin Level Analysis
3.4.7. Determination of Phenolics Levels and Their Biosynthetic Enzyme Activity
3.4.8. Phosphate Solubilization
3.5. Biological Activities
3.5.1. Antioxidant Activities
3.5.2. Antibacterial Activity
3.5.3. Determination of Lipoxygenase (LOX) and Cyclooxygenase (COX) Activities
3.6. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Manchali, S.; Murthy, K.N.C.; Patil, B.S. Crucial facts about health benefits of popular cruciferous vegetables. J. Funct. Foods 2012, 4, 94–106. [Google Scholar]
- Marton, M.; Mandoki, Z.S.; Csapo-Kiss, Z.S.; Csapo, J. The role of sprouts in human nutrition. A review. Acta Univ. Sapientiae 2010, 3, 81–117. [Google Scholar]
- Almuhayawi, M.S.; Hassan, A.H.A.; Abdel-Mawgoud, M.; Khamis, G.; Selim, S.; Al Jaouni, S.K.; AbdElgawad, H. Laser light as a promising approach to improve the nutritional value, antioxidant capacity and anti-inflammatory activity of flavonoid-rich buckwheat sprouts. Food Chem. 2020, 345, 128788. [Google Scholar] [CrossRef]
- Almuhayawi, M.S.; Al Jaouni, S.K.; Almuhayawi, S.M.; Selim, S.; Abdel-Mawgoud, M. Elevated CO2 improves the nutritive value, antibacterial, anti-inflammatory, antioxidant and hypocholestecolemic activities of lemongrass sprouts. Food Chem. 2021, 357, 129730. [Google Scholar] [CrossRef]
- Santoyo, G.; Moreno-Hagelsieb, G.; Del Carmen Orozco-Mosqueda, M.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Lodewyckx, C.; Vangronsveld, J.; Porteous, F.; Moore, E.R.B.; Taghavi, S.; Mezgeay, M.; Der Lelie, D.V. Endophytic bacteria and their potential applications. CRC Crit. Rev. Plant Sci. 2002, 21, 583–606. [Google Scholar] [CrossRef]
- Chaturvedi, H.; Singh, V.; Gupta, G. Potential of bacterial endophytes as plant growth promoting factors. J. Plant Pathol. Microbiol. 2016, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Jasim, B.; Joseph, A.A.; John, C.J.; Mathew, J.; Radhakrishnan, E.K. Isolation and characterization of plant growth promoting endophytic bacteria from the rhizome of Zingiber officinale. 3 Biotech 2014, 4, 197–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brader, G.; Compant, S.; Mitter, B.; Trognitz, F.; Sessitsch, A. Metabolic potential of endophytic bacteria. Curr. Opin. Biotechnol. 2014, 27, 30–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aeron, A.; Maheshwari, D.K.; Meena, V.S. Endophytic bacteria promote growth of the medicinal legume Clitoria ternatea L. by chemotactic activity. Arch. Microbiol. 2020, 202, 1049–1058. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Sun, K.; Chen, F.; Yuan, J.; Li, X.; Dai, C. Endophytic Pseudomonas induces metabolic flux changes that enhance medicinal sesquiterpenoid accumulation in Atractylodes lancea. Plant Physiol. Biochem. 2018, 130, 473–481. [Google Scholar] [CrossRef]
- Xu, M.; Bai, H.-Y.; Fu, W.-Q.; Sun, K.; Wang, H.-W.; Xu, D.-L.; Dai, C.-C.; Jia, Y. Endophytic bacteria promote the quality of Lyophyllum decastes by improving non-volatile taste components of mycelia. Food Chem. 2021, 336, 127672. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Estrella, D.; Bresciani, A.; Iametti, S.; Marengo, M.; Pagani, M.A.; Marti, A. Effect of sprouting on proteins and starch in quinoa (Chenopodium quinoa Willd.). Plant Foods Hum. Nutr. 2020, 75, 635–641. [Google Scholar] [CrossRef]
- Paśko, P.; Sajewicz, M.; Gorinstein, S.; Zachwieja, Z. Analysis of selected phenolic acids and flavonoids in Amaranthus cruentus and Chenopodium quinoa seeds and sprouts by HPLC. Acta Chromatogr. 2008, 20, 661–672. [Google Scholar] [CrossRef] [Green Version]
- Chao, Y.-Y.; Wang, W.-J.; Liu, Y.-T. Effect of Calcium on the Growth of Djulis (Chenopodium formosanum Koidz.) Sprouts. Agronomy 2021, 11, 82. [Google Scholar] [CrossRef]
- Kumar, R.; Mishra, A.K.; Dubey, N.K.; Tripathi, Y.B. Evaluation of Chenopodium ambrosioides oil as a potential source of antifungal, antiaflatoxigenic and antioxidant activity. Int. J. Food Microbiol. 2007, 115, 159–164. [Google Scholar] [CrossRef]
- Kokanova-Nedialkova, Z.; Nedialkov, P.; Nikolov, S. The genus Chenopodium: Phytochemistry, ethnopharmacology and pharmacology. Pharmacogn. Rev. 2009, 3, 280. [Google Scholar]
- Ullah, F.; Iqbal, N.; Ayaz, M.; Sadiq, A.; Ullah, I.; Ahmad, S.; Imran, M. DPPH, ABTS free radical scavenging, antibacterial and phytochemical evaluation of crude methanolic extract and subsequent fractions of Chenopodium botrys aerial parts. Pak. J. Pharm. Sci. 2017, 30, 761–766. [Google Scholar]
- Sera, B.; Stranák, V.; Serý, M.; Tichý, M.; Spatenka, P. Germination of Chenopodium album in response to microwave plasma treatment. Plasma Sci. Technol. 2008, 10, 506. [Google Scholar] [CrossRef]
- Khan, A.L.; Waqas, M.; Kang, S.-M.; Al-Harrasi, A.; Hussain, J.; Al-Rawahi, A.; Al-Khiziri, S.; Ullah, I.; Ali, L.; Jung, H.-Y. Bacterial endophyte Sphingomonas sp. LK11 produces gibberellins and IAA and promotes tomato plant growth. J. Microbiol. 2014, 52, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Almuhayawi, M.S.; Mohamed, M.S.M.; Abdel-Mawgoud, M.; Selim, S.; Al Jaouni, S.K.; AbdElgawad, H. Bioactive Potential of Several Actinobacteria Isolated from Microbiologically Barely Explored Desert Habitat, Saudi Arabia. Biology 2021, 10, 235. [Google Scholar] [CrossRef]
- Manfio, G.P. Towards minimal standards for the description of Streptomyces species. Biotekhnologiya 1995, 8, 228–237. [Google Scholar]
- Boroujeni, M.E.; Arijit, D.; Prashanthi, K.; Sandeep, S.; Sourav, B. Enzymatic screening and random amplified polymorphic DNA fingerprinting of soil streptomycetes isolated from Wayanad district in Kerala, India. J. Biol. Sci. 2012, 12, 43–50. [Google Scholar]
- Ser, H.-L.; Law, J.W.-F.; Chaiyakunapruk, N.; Jacob, S.A.; Palanisamy, U.D.; Chan, K.-G.; Goh, B.-H.; Lee, L.-H. Fermentation conditions that affect clavulanic acid production in Streptomyces clavuligerus: A systematic review. Front. Microbiol. 2016, 7, 522. [Google Scholar] [CrossRef]
- Briatia, X.; Azad, M.O.K.; Khanongnuch, C.; Woo, S.H.; Park, C.H. Effect of Endophytic Bacterium Inoculation on Total Polyphenol and Flavonoid Contents of Tartary Buckwheat Sprouts. Korean J. Crop Sci. 2018, 63, 57–63. [Google Scholar]
- Mandal, S.M.; Chakraborty, D.; Dey, S. Phenolic acids act as signaling molecules in plant-microbe symbioses. Plant Signal. Behav. 2010, 5, 359–368. [Google Scholar] [CrossRef] [Green Version]
- Abdelmohsen, U.R.; Grkovic, T.; Balasubramanian, S.; Kamel, M.S.; Quinn, R.J.; Hentschel, U. Elicitation of secondary metabolism in actinomycetes. Biotechnol. Adv. 2015, 33, 798–811. [Google Scholar] [CrossRef] [Green Version]
- Bianco, C.; Imperlini, E.; Calogero, R.; Senatore, B.; Pucci, P.; Defez, R. Indole-3-acetic acid regulates the central metabolic pathways in Escherichia coli. Microbiology 2006, 152, 2421–2431. [Google Scholar] [CrossRef] [Green Version]
- Ryan, R.P.; Monchy, S.; Cardinale, M.; Taghavi, S.; Crossman, L.; Avison, M.B.; Berg, G.; Van Der Lelie, D.; Dow, J.M. The versatility and adaptation of bacteria from the genus Stenotrophomonas. Nat. Rev. Microbiol. 2009, 7, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Lou, K.; Li, C. Growth and photosynthetic efficiency promotion of sugar beet (Beta vulgaris L.) by endophytic bacteria. Photosynth. Res. 2010, 105, 5–13. [Google Scholar] [CrossRef]
- Rho, H.; Kim, S.-H. Endophyte effects on photosynthesis and water use of plant hosts: A meta-analysis. In Functional Importance of the Plant Microbiome; Springer: Cham, Switzerland, 2017; pp. 43–69. [Google Scholar]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3-acetic acid in plant–microbe interactions. Antonie Van Leeuwenhoek 2014, 106, 85–125. [Google Scholar] [CrossRef]
- Saleem, M.; Asghar, H.N.; Zahir, Z.A.; Shahid, M. Impact of lead tolerant plant growth promoting rhizobacteria on growth, physiology, antioxidant activities, yield and lead content in sunflower in lead contaminated soil. Chemosphere 2018, 195, 606–614. [Google Scholar] [CrossRef]
- Kang, S.-M.; Radhakrishnan, R.; You, Y.-H.; Joo, G.-J.; Lee, I.-J.; Lee, K.-E.; Kim, J.-H. Phosphate solubilizing Bacillus megaterium mj1212 regulates endogenous plant carbohydrates and amino acids contents to promote mustard plant growth. Indian J. Microbiol. 2014, 54, 427–433. [Google Scholar] [CrossRef]
- Stefan, M.; Munteanu, N.; Stoleru, V.; Mihasan, M. Effects of inoculation with plant growth promoting rhizobacteria on photosynthesis, antioxidant status and yield of runner bean. Rom. Biotechnol. Lett. 2013, 18, 8132–8143. [Google Scholar]
- Lemoine, R.; La Camera, S.; Atanassova, R.; Dédaldéchamp, F.; Allario, T.; Pourtau, N.; Bonnemain, J.-L.; Laloi, M.; Coutos-Thévenot, P.; Maurousset, L. Source-to-sink transport of sugar and regulation by environmental factors. Front. Plant Sci. 2013, 4, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briatia, X.; Jomduang, S.; Park, C.H.; Lumyong, S.; Kanpiengjai, A.; Khanongnuch, C. Enhancing growth of buckwheat sprouts and microgreens by endophytic bacterium inoculation. Int. J. Agric. Biol. 2017, 19, 374–380. [Google Scholar] [CrossRef]
- Cipriano, M.A.P.; Freitas-Iório, R.D.P.; Dimitrov, M.R.; De Andrade, S.A.L.; Kuramae, E.E.; Silveira, A.P.D.D. Plant-Growth Endophytic Bacteria Improve Nutrient Use Efficiency and Modulate Foliar N-Metabolites in Sugarcane Seedling. Microorganisms 2021, 9, 479. [Google Scholar] [CrossRef] [PubMed]
- Ahemad, M.; Kibret, M. Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. J. King Saud Univ. 2014, 26, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Taurian, T.; Anzuay, M.S.; Angelini, J.G.; Tonelli, M.L.; Ludueña, L.; Pena, D.; Ibáñez, F.; Fabra, A. Phosphate-solubilizing peanut associated bacteria: Screening for plant growth-promoting activities. Plant Soil 2010, 329, 421–431. [Google Scholar] [CrossRef]
- Ren, X.-M.; Guo, S.-J.; Tian, W.; Chen, Y.; Han, H.; Chen, E.; Li, B.-L.; Li, Y.-Y.; Chen, Z.-J. Effects of plant growth-promoting bacteria (PGPB) inoculation on the growth, antioxidant activity, Cu uptake, and bacterial community structure of rape (Brassica napus L.) grown in Cu-contaminated agricultural soil. Front. Microbiol. 2019, 10, 1455. [Google Scholar] [CrossRef]
- Andov, L.A.; Karapandzova, M.; Stefkov, G.; Cvetkovikj, I.; Baceva, K.; Stafilov, T.; Kulevanova, S. The content of some biogenic elements in Chenopodium album L. and Chenopodium botrys L.(Amaranthaceae) from Macedonian flora. Maced. Pharm. Bull. 2016, 62, 499−500. [Google Scholar]
- Reyes-Becerril, M.; Angulo, C.; Sanchez, V.; Vázquez-Martínez, J.; López, M.G. Antioxidant, intestinal immune status and anti-inflammatory potential of Chenopodium ambrosioides L. in fish: In vitro and in vivo studies. Fish Shellfish Immunol. 2019, 86, 420–428. [Google Scholar] [CrossRef]
- Aborisade, A.B.; Adetutu, A.; Owoade, A.O. Phytochemical and proximate analysis of some medicinal leaves. Clin. Med. Res. 2017, 6, 209. [Google Scholar] [CrossRef] [Green Version]
- Whiteside, M.D.; Garcia, M.O.; Treseder, K.K. Amino acid uptake in arbuscular mycorrhizal plants. PLoS ONE 2012, 7, e47643. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Zhang, S.; Zhang, J.; Ma, P.; Duan, J.; Liang, Z. Effect and mechanism of endophytic bacteria on growth and secondary metabolite synthesis in Salvia miltiorrhiza hairy roots. Acta Physiol. Plant. 2014, 36, 1095–1105. [Google Scholar] [CrossRef]
- Barros, L.; Pereira, E.; Calhelha, R.C.; Dueñas, M.; Carvalho, A.M.; Santos-Buelga, C.; Ferreira, I.C.F.R. Bioactivity and chemical characterization in hydrophilic and lipophilic compounds of Chenopodium ambrosioides L. J. Funct. Foods 2013, 5, 1732–1740. [Google Scholar] [CrossRef]
- Saleh, A.M.; Abdel-Mawgoud, M.; Hassan, A.R.; Habeeb, T.H.; Yehia, R.S.; AbdElgawad, H. Global metabolic changes induced by arbuscular mycorrhizal fungi in oregano plants grown under ambient and elevated levels of atmospheric CO2. Plant Physiol. Biochem. 2020, 151, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Ajaib, M.; Hussain, T.; Farooq, S.; Ashiq, M. Analysis of antimicrobial and antioxidant activities of Chenopodium ambrosioides: An ethnomedicinal plant. J. Chem. 2016, 2016, 4827157. [Google Scholar] [CrossRef] [Green Version]
- Ozer, M.S.; Sarikurkcu, C.; Tepe, B. Phenolic composition, antioxidant and enzyme inhibitory activities of ethanol and water extracts of Chenopodium botrys. RSC Adv. 2016, 6, 64986–64992. [Google Scholar] [CrossRef]
- Chen, L.; Xu, M.; Zheng, Y.; Men, Y.; Sheng, J.; Shen, L. Growth promotion and induction of antioxidant system of tomato seedlings (Solanum lycopersicum L.) by endophyte TPs-04 under low night temperature. Sci. Hortic. 2014, 176, 143–150. [Google Scholar] [CrossRef]
- Brahim, M.A.S.; Fadli, M.; Hassani, L.; Boulay, B.; Markouk, M.; Bekkouche, K.; Abbad, A.; Ali, M.A.; Larhsini, M. Chenopodium ambrosioides var. ambrosioides used in Moroccan traditional medicine can enhance the antimicrobial activity of conventional antibiotics. Ind. Crops Prod. 2015, 71, 37–43. [Google Scholar] [CrossRef]
- Jesus, R.S.; Piana, M.; Freitas, R.B.; Brum, T.F.; Alves, C.F.S.; Belke, B.V.; Mossmann, N.J.; Cruz, R.C.; Santos, R.C.V.; Dalmolin, T.V. In vitro antimicrobial and antimycobacterial activity and HPLC–DAD screening of phenolics from Chenopodium ambrosioides L. Braz. J. Microbiol. 2018, 49, 296–302. [Google Scholar] [CrossRef]
- TrivellatoGrassi, L.; Malheiros, A.; Meyre-Silva, C.; Da Silva Buss, Z.; Monguilhott, E.D.; Fröde, T.S.; Da Silva, K.A.B.S.; De Souza, M.M. From popular use to pharmacological validation: A study of the anti-inflammatory, anti-nociceptive and healing effects of Chenopodium ambrosioides extract. J. Ethnopharmacol. 2013, 145, 127–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taechowisan, T.; Peberdy, J.F.; Lumyong, S. Isolation of endophytic actinomycetes from selected plants and their antifungal activity. World J. Microbiol. Biotechnol. 2003, 19, 381–385. [Google Scholar] [CrossRef]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indoleacetic acid. Plant Physiol. 1951, 26, 192. [Google Scholar] [CrossRef] [Green Version]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Williams, S.T. Genus Streptomyces waksman and henrici 1943. Bwergey’s Man. Syst. Bacteriol. 1989, 4, 2452–2492. [Google Scholar]
- Hong, K.; Gao, A.-H.; Xie, Q.-Y.; Gao, H.G.; Zhuang, L.; Lin, H.-P.; Yu, H.-P.; Li, J.; Yao, X.-S.; Goodfellow, M. Actinomycetes for marine drug discovery isolated from mangrove soils and plants in China. Mar. Drugs 2009, 7, 24–44. [Google Scholar] [CrossRef] [PubMed]
- Stackebrandt, E.; Goodfellow, M. Nucleic Acid Techniques in Bacterial Systematics; Wiley: Hoboken, NJ, USA, 1991; ISBN 0471929069. [Google Scholar]
- Reddy, G.S.; Aggarwal, R.K.; Matsumoto, G.I.; Shivaji, S. Arthrobacter flavus sp. nov., a psychrophilic bacterium isolated from a pond in McMurdo Dry Valley, Antarctica. Int. J. Syst. Evol. Microbiol. 2000, 50, 1553–1561. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+ C− content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.H.; Cheung, P.C.K. Nutritional evaluation of some subtropical red and green seaweeds: Part I—proximate composition, amino acid profiles and some physico-chemical properties. Food Chem. 2000, 71, 475–482. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Shiva, S.; Enninful, R.; Roth, M.R.; Tamura, P.; Jagadish, K.; Welti, R. An efficient modified method for plant leaf lipid extraction results in improved recovery of phosphatidic acid. Plant Methods 2018, 14, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.C.; Prosky, L.; Vries, J.W.D. Determination of total, soluble, and insoluble dietary fiber in foods—Enzymatic-gravimetric method, MES-TRIS buffer: Collaborative study. J. AOAC Int. 1992, 75, 395–416. [Google Scholar] [CrossRef]
- AbdElgawad, H.; Peshev, D.; Zinta, G.; Van den Ende, W.; Janssens, I.A.; Asard, H. Climate extreme effects on the chemical composition of temperate grassland species under ambient and elevated CO2: A comparison of fructan and non-fructan accumulators. PLoS ONE 2014, 9, e92044. [Google Scholar]
- Sinha, A.K.; Giblen, T.; AbdElgawad, H.; De Rop, M.; Asard, H.; Blust, R.; De Boeck, G. Regulation of amino acid metabolism as a defensive strategy in the brain of three freshwater teleosts in response to high environmental ammonia exposure. Aquat. Toxicol. 2013, 130–131, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Kumpaisal, R.; Hashimoto, T.; Yamada, Y. Purification and characterization of dihydrodipicolinate synthase from wheat suspension cultures. Plant Physiol. 1987, 85, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Ravanel, S.; Gakière, B.; Job, D.; Douce, R. Cystathionine γ-synthase from Arabidopsis thaliana: Purification and biochemical characterization of the recombinant enzyme overexpressed in Escherichia coli. Biochem. J. 1998, 331, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Hamad, I.; Abdelgawad, H.; Al Jaouni, S.; Zinta, G.; Asard, H.; Hassan, S.; Hegab, M.; Hagagy, N.; Selim, S. Metabolic analysis of various date palm fruit (Phoenix dactylifera L.) cultivars from Saudi Arabia to assess their nutritional quality. Molecules 2015, 20, 13620–13641. [Google Scholar] [CrossRef]
- Habeeb, T.H.; Abdel-Mawgoud, M.; Yehia, R.S.; Khalil, A.M.A.; Saleh, A.M.; AbdElgawad, H. Interactive Impact of Arbuscular Mycorrhizal Fungi and Elevated CO2 on Growth and Functional Food Value of Thymus vulgare. J. Fungi 2020, 6, 168. [Google Scholar] [CrossRef] [PubMed]
- Farfan-Vignolo, E.R.; Asard, H. Effect of elevated CO2 and temperature on the oxidative stress response to drought in Lolium perenne L. and Medicago sativa L. Plant Physiol. Biochem. 2012, 59, 55–62. [Google Scholar] [CrossRef] [PubMed]
- King, E.J. The colorimetric determination of phosphorus. Biochem. J. 1932, 26, 292–297. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Endo 1 | Endo 2 | Endo 3 | Endo 4 | Endo 5 | Endo 6 | |
|---|---|---|---|---|---|---|---|
| Colony | Aerial mycelium | + | − | + | + | − | + |
| Pigmentation | + | + | + | + | + | + | |
| Spore chain | Spiral | + | − | − | + | + | − |
| Rectiflexibles | − | + | + | − | − | − | |
| Verticillate | − | − | − | − | − | + | |
| Spore color | Yellow | + | − | − | − | − | − |
| Orange | − | + | + | + | − | + | |
| Red | − | − | − | − | + | − | |
| N source utilization | L-Cysteine | + | + | + | + | + | − |
| L-Phenylalanine | − | − | + | − | − | ||
| L-Histidine | − | − | + | + | + | − | |
| L-Lysine | + | + | + | − | − | + | |
| L-Asparagine | + | - | + | + | + | + | |
| L-Arginine | + | − | − | − | − | ||
| L-proline | + | + | + | + | − | ||
| L-Valine | − | − | + | − | − | + | |
| Tyrosine | + | + | − | + | + | ||
| C source utilization | D-fructose | − | − | + | + | + | − |
| D-glucose | + | + | − | + | − | − | |
| Sucrose | + | + | + | - | + | + | |
| Maltose | − | − | + | − | + | ||
| Raffinose | + | + | − | + | + | + | |
| Lactose | − | − | + | + | − | ||
| Galactose | + | + | − | + | + | − | |
| Meso-Inositol | + | + | + | − | + | + | |
| Cellulose | − | − | + | + | + | + | |
| Xylose | + | + | + | − | + | − | |
| Dextran | + | + | − | − | − | − | |
| Enzyme activity | Catalase | + | − | + | − | + | − |
| Peroxidase | + | − | + | + | + | + | |
| Starch hydrolysis | + | + | − | − | + | ||
| Gelatin liquefication | + | + | − | + | − | + | |
| Casein hydrolysis | − | − | + | + | + | + | |
| Lipolysis | + | + | + | + | − | + | |
| Citrate utilization | + | + | + | + | + | + | |
| H2S Production | − | + | − | + | + | + | |
| Nitrate reduction | + | + | + | − | − | − | |
| Urease | + | + | + | − | − | ||
| L-Asparaginase | − | + | + | + | + | − | |
| L-Glutaminase | + | + | + | + | + | + | |
| Biological activity | Antioxidant Activity (FRAP) | 42.4 ± 3.1 | 66.6 ± 4.0 | 21.8 ± 1.6 | 33.1 ± 1.5 | 24 ± 1.4 | 32.4 ± 2.3 |
| Antioxidant Activity DPPH (%) | 68.7 ± 2.8 | 73.7 ± 3.8 | 34.2 ± 2.1 | 20.8 ± 1.5 | 27.5 ± 1.4 | 48.2 ± 2.9 | |
| Antioxidant Activity (ABTS%) | 30.5 ± 1.0 | 59.6 ± 1.9 | 39.7 ± 1.3 | 34.8 ± 1.1 | 23.2 ± 0.8 | 53.6 ± 1.7 | |
| Phosphate Solubilization (mg/mL) | 5.7 ± 0.8 | 7.2 ± 0.2 | 4.5 ± 0.1 | 3.8 ± 1.5 | 2.7 ± 1.4 | 7.3 ± 1.0 | |
| Bioactive compounds production | Total flavonoids (mg/100 g bacteria) | 5 ± 1.19 | 8 ± 1.53 | 5.8 ± 0.84 | 5.3 ± 1.36 | 6.4 ± 0.86 | 7.5 ± 1.18 |
| Total Phenols (mg/100 g bacteria) | 36.8 ± 1.06 | 45.4 ± 1.31 | 23.7 ± 0.6 | 42.2 ± 1.2 | 23.7 ± 0.6 | 33.8 ± 0.9 | |
| Tocopherols (mg/g bacteria) | 0.3 ± 0.01 | 0.5 ± 0.01 | 0.2 ± 0.01 | 0.5 ± 0.01 | 0.3 ± 0.01 | 0.2 ± 0.01 | |
| Flavonoids (mg/100 g bacteria) | |||||||
| Quercetin | 1.63 ± 0.1 | ||||||
| Quercetrin | 1.41 ± 0.5 | ||||||
| Luteolin | 0.70 ± 0.1 | ||||||
| Apigenin | 4.11 ± 0.7 | ||||||
| Isoquercetrin | 10.2 ± 1.6 | ||||||
| Rutin | 1.27 ± 0.4 | ||||||
| Ellagic acid | 0.71 ± 0.1 | ||||||
| Velutin | 0.30 ± 0.0 | ||||||
| Naringenin | 1.12 ± 0.3 | ||||||
| Genistein | 0.95 ± 0.2 | ||||||
| Daidzein | 1.08 ± 0.1 | ||||||
| Fisetin | 0.74 ± 0.1 | ||||||
| O-hydroxydaidzein | 1.13 ± 0.1 | ||||||
| IAA-Me | 1.15 ± 0.21 | ||||||
| ABA | 0.29 ± 0.1 | ||||||
| GA | 0.16 ± 0.07 | ||||||
| Sidephore Catechol | 7.3 ± 0.3 | ||||||
| Sidephore Salicylate | 9.23 ± 0.42 |
| C. ambrosoides | C. ficifolium | C. botrys | ||||
|---|---|---|---|---|---|---|
| Control | Endo | Control | Endo | Control | Endo | |
| Photosynthetic Related Parameters (μmol CO2 m−2 s−1) | ||||||
| Photosynthesis | 10.1 ± 0.8 a | 11.5 ± 0.8 b | 11.3 ± 1.1 a | 13.1 ± 0.7 b | 9.7 ± 1 a | 12.8 ± 1.2 b |
| Respiration | 1.4 ± 5.4 a | 1.9 ± 0.0 ab | 1.2 ± 0.06 a | 2.0 ± 0.1 b | 1.1 ± 0.0 a | 1.9 ± 0.1 b |
| Pigments (mg/gFW) | ||||||
| Chl a | 2.07 ± 0.4 a | 2.42 ± 0.38 a | 1.65 ± 0.2 a | 2.78 ± 0.4 b | 1.76 ± 0.3 a | 2.55 ± 0.4 b |
| Chl b | 1.04 ± 0.1 a | 1.46 ± 0.2 a | 1.02 ± 0.1 a | 1.65 ± 0.3 ab | 1 ± 0.05 a | 1.89 ± 0.41 b |
| Chl a+b | 3.11 ± 0.5 a | 3.8 ± 0.3 a | 2.68 ± 0.4 a | 4.4 ± 0.41 a | 2.7 ± 0.3 a | 4.44 ± 0.8 b |
| Beta-carotene | 0.06 ± 0.01 a | 0.11 ± 0.02 b | 0.06 ± 0.0 a | 0.12 ± 0.02 b | 0.07 ± 0.01 a | 0.11 ± 0.01 ab |
| C. ambrosoides | C. ficifolium | C. botrys | ||||
|---|---|---|---|---|---|---|
| Control | Endo | Control | Endo | Control | Endo | |
| Minerals (mg/gDW) | ||||||
| K | 13.9 ± 0.4 a | 15.97 ± 1.2 ab | 11.4 ± 0.12 a | 23.2 ± 1 b | 15.5 ± 1 a | 29.8 ± 1.3 b |
| Na | 1.8 ± 0.04 a | 2.1 ± 0.3 b | 1.5 ± 0.01 a | 3.2 ± 0.1 b | 1.9 ± 0.1 a | 4.1 ± 0.1 b |
| Ca | 1.46 ± 0.3 a | 2.1 ± 0.12 b | 1.4 ± 0.17 a | 2.28 ± 0.1 b | 1.5 ± 0.2 a | 1.95 ± 0.02 a |
| Cu | 0.006 ± 0 a | 0.008 ± 0 ab | 0.007 ± 0 a | 0.011 ± 0 b | 0.01 ± 0 a | 0.013 ± 0 a |
| Fe | 0.13 ± 0.02 a | 0.19 ± 0 b | 0.16 ± 0.01 a | 0.2 ± 0.02 a | 0.2 ± 0.01 a | 0.33 ± 0.04 ab |
| P | 0.96 ± 0.1 a | 1.4 ± 0.05 ab | 1.13 ± 0.07 a | 1.62 ± 0.05 b | 1.06 ± 0.0 a | 1.64 ± 0.2 a |
| Zn | 0.09 ± 0 a | 0.12 ± 0.01 ab | 0.11 ± 0 a | 0.14 ± 0.01 a | 0.12 ± 0.0 a | 0.16 ± 0.01 a |
| Mn | 0.028 ± 0 a | 0.024 ± 0 a | 0.027 ± 0 a | 0.03 ± 0 a | 0.02 ± 0 a | 0.031 ± 0 ab |
| Mg | 2.8 ± 0.3 a | 2.3 ± 0.2 a | 3.18 ± 0.39 a | 3.9 ± 0.2 a | 3.1 ± 0.5 a | 5.18 ± 0.1 b |
| Vitamins (mg/gFW) | ||||||
| Vitamin C | 2.09 ± 0.07 a | 2.34 ± 0.42 b | 1.92 ± 0.06 a | 3.04 ± 0.61 b | 1.7 ± 0.72 a | 2.7 ± 0.8 b |
| Vitamin E | 4.71 ± 0.2 a | 6.69 ± 0.23 b | 4.25 ± 0.42 a | 6.54 ± 0.58 b | 3.46 ± 0.2 a | 6.4 ± 0.6 b |
| Thiamin | 0.49 ± 0.1 a | 0.53 ± 0.14 a | 0.42 ± 0.05 a | 0.58 ± 0.13 a | 0.35 ± 0.07 a | 0.64 ± 0.1 b |
| Riboflavin | 0.15 ± 0.03 a | 0.18 ± 0.05 a | 0.13 ± 0.03 a | 0.18 ± 0.02 a b | 0.09 ± 0.01 a | 0.2 ± 0.04 b |
| Proximate composition (mg/gFW) | ||||||
| Total proteins | 9.1 ± 1 a | 6.5 ± 4.8 b | 11.2 ± 1.8 a | 16 ± 3.7 b | 9.1 ± 1 a | 13.1 ± 3.2 b |
| Fat | 123 ± 5.4 a | 125 ± 15 a | 151.3 ± 6 a | 140. ± 6 b | 123 ± 5.4 a | 150.1 ± 16 b |
| Crude Fiber | 6.6 ± 0.6 a | 6.8 ± 1.8 a | 8.1 ± 0.7 a | 12.3 ± 1 b | 6.6 ± 0.6 a | 10.1 ± 0.8 b |
| Ash | 3.8 ± 0.5 a | 3.6 ± 0.4 a | 3.9 ± 0.5 a | 5.3 ± 0.8 b | 3.4 ± 0.5 a | 4.6 ± 0.7 b |
| Carbohydrate | 6.3 ± 0.3 a | 5.9 ± 0.2 a | 7.7 ± 0.4 a | 11.1 ± 0.8 b | 6.3 ± 0.3 a | 10.1 ± 2 b |
| C. ambrosoides | C. ficifolium | C. botrys | ||||
|---|---|---|---|---|---|---|
| Control | Endo | Control | Endo | Control | Endo | |
| Amino Acid Metabolism (µg/gFW) | ||||||
| Glutamic acid | 14.6 ± 1.1 a | 19.2 ± 1.8 b | 14.98 ± 1 a | 18.4 ± 2.7 b | 14.5 ± 1.4 a | 19.9 ± 1.3 b |
| Glutamine | 12.8 ± 0.9 a | 16.4 ± 1.1 b | 12.5 ± 0.9 a | 16.0 ± 1.2 b | 12.6 ± 1.3 a | 17.4 ± 1.3 b |
| Serine | 6.39 ± 0.3 a | 8.61 ± 1.1 b | 6.7 ± 0.5 a | 8.8 ± 0.8 b | 9.8 ± 2.4 a | 11.8 ± 1.9 c |
| Glycine | 8.3 ± 0.13 a | 9.6 ± 0.0 b | 7.6 ± 0.12 a | 9.5 ± 0.7 b | 7.3 ± 0.66 a | 9.7 ± 0.18 b |
| Arg | 13.6 ± 0.5 a | 16 ± 1.5 ab | 20.9 ± 3.5 a | 23.3 ± 2.3 b | 21.6 ± 3.8 a | 24.2 ± 1.8 b |
| Alanine | 2.4 ± 0.3 a | 2.48 ± 0.8 a | 3.5 ± 0.16 a | 2.7 ± 0.5 b | 3.4 ± 0.8 a | 2.85 ± 0.5 a |
| Histidine | 5.2 ± 0.48 a | 5.7 ± 0.2 ab | 4.98 ± 0.8 a | 5.2 ± 0.29 a | 6.1 ± 0.42 a | 4.6 ± 0.7 b |
| Valine | 5.3 ± 0.7 a | 6.95 ± 1 b | 5.05 ± 1.3 a | 5.72 ± 0.9 a | 5.9 ± 0.8 a | 6.0 ± 1.04 a |
| Methionine | 2.2 ± 0.19 a | 2.9 ± 0.3 ab | 2.01 ± 0.2 a | 3.5 ± 0.4 b | 1.65 ± 0.1 a | 3.31 ± 0.0 b |
| Cystine | 1.4 ± 0.03 a | 2.1 ± 0.18 b | 1.7 ± 0.06 a | 2.3 ± 0.2 b | 1.4 ± 0.12 a | 1.9 ± 0.1 ab |
| Isoleucine | 4.2 ± 0.2 a | 5.48 ± 0.3 a | 7.15 ± 1 a | 7.3 ± 1.5 a | 5.2 ± 0.79 a | 7.8 ± 0.9 b |
| Leucine | 13.8 ± 0.4 a | 14 ± 0.9 ab | 11.5 ± 1.4 a | 12.0 ± 1.1 a | 11.1 ± 0.7 a | 13.0 ± 1 ab |
| Tyrosine | 5.6 ± 0.7 a | 6.3 ± 0.5 ab | 5.47 ± 0.4 a | 7.2 ± 0.2 b | 5.3 ± 0.1 a | 6.4 ± 0.6 ab |
| Lysine | 13.5 ± 0.3 a | 21.1 ± 1.6 b | 12.8 ± 0.6 a | 21.3 ± 1.3 b | 13.3 ± 0.7 a | 25.8 ± 2.5 b |
| Threonine | 4.6 ± 0.3 a | 5.3 ± 0.6 ab | 4.4 ± 0.4 a | 6.3 ± 0.2 b | 4.9 ± 0.1 a | 5.9 ± 0.2 ab |
| Trep | 0.4 ± 0.04 a | 0.61 ± 0.01 | 0.4 ± 0.04 a | 0.6 ± 0.03 | 0.4 ± 0.04 a | 0.67 ± 0.04 |
| Amino Acid Biosynthesis Enzymes | ||||||
| GS (nmol γ-glutamyl hydroxamate/mg protein min−1) | 5.4 ± 0.4 a | 7.14 ± 0.5 b | 5.51 ± 0.4 a | 6.8 ± 0.7 ab | 5.44 ± 0.5 a | 7.4 ± 0.7 b |
| DHDPS (nmol o-ABA and L-2,3- dihydrodipicolinate adduct/mg protein min−1) | 2.09 ± 0.3 a | 2.4 ± 0.2 a | 2.09 ± 0.3 a | 3.5 ± 0.2 b | 2.5 ± 0.4 a | 3.1 ± 0.3 b |
| CGS (nmol L-cystathionine/mg protein min−1) | 0.012 ± 0 a | 0.033 ± 0 b | 0.013 ± 0 a | 0.039 ± 0 b | 0.014 ± 0 a | 0.37 ± 0 b |
| Organic Acid (mg/g FW) | ||||||
| Oxalic | 9.08 ± 0.8 a | 9.5 ± 0.4 a | 3.5 ± 0.5 a | 4.73 ± 4.7 b | 1.92 ± 0.3 a | 4.73 ± 0.5 b |
| Malic | 2.3 ± 0.31 a | 3.58 ± 0.2 ab | 2.92 ± 0.3 a | 2.67 ± 0.6 a | 2.1 ± 0.22 a | 4.27 ± 0.9 b |
| Succinic | 2.17 ± 0 a | 2.03 ± 0 a | 1.75 ± 0.3 a | 3.5 ± 0.3 a | 3.2 ± 1.2 a | 5.9 ± 0.9 b |
| Citric | 1.3 ± 0.04 a | 1.9 ± 0.09 a | 2.96 ± 0.29 a | 2.9 ± 0.2 a | 2.88 ± 0.5 a | 2.5 ± 0.5 ab |
| Lactic | 0.2 ± 0.1 a | 0.24 ± 0.1 a | 0.32 ± 0.09 a | 0.26 ± 0.07 a | 0.33 ± 0.05 a | 0.49 ± 0.05 ab |
| Fatty Acids (mg/g FW) | ||||||
| Tetradecanoic (C14:0) | 0.7 ± 0.1 a | 0.8 ± 0.1 ab | 0.8 ± 0.1 a | 0.98 ± 0.09 b | 0.66 ± 0.06 a | 1.02 ± 0.1 b |
| Pentadecanoic (C16:0) | 12 ± 2 a | 15.8 ± 1 ab | 11.6 ± 0.6 a | 15.5 ± 0.7 ab | 18 ± 1.6 a | 18.1 ± 1.3 a |
| Eicosanoic (C20:0) | 0.8 ± 0.03 a | 0.96 ± 0.1 ab | 1.1 ± 0.2 a | 1.02 ± 0.1 a | 0.88 ± 0.09 a | 0.96 ± 0.1 a |
| Docosanoic (C22:0) | 0.8 ± 0.1 a | 1.45 ± 0.2 b | 0.91 ± 0.1 a | 1.29 ± 0.1 ab | 0.91 ± 0.1 a | 1.2 ± 0.1 ab |
| Octadecanoic (C18:0) | 6.9 ± 0.7 a | 10.2 ± 1.8 b | 7.2 ± 0.8 a | 8.9 ± 0.9 ab | 7.2 ± 1.2 a | 9.5 ± 1.2 b |
| Pentacosanoic (C24:0) | 0.14 ± 0.0 a | 0.13 ± 0.02 a | 0.12 ± 0.01 a | 0.12 ± 0.01 a | 0.13 ± 0.01 a | 0.15 ± 0.1 a |
| Pentadecanoic (C16:1) | 1.42 ± 0.1 a | 1.3 ± 0.09 a | 1.4 ± 0.23 a | 2.3 ± 0.2 b | 1.55 ± 0.3 a | 2.5 ± 0.1 b |
| Pentadecanoic (C16:1) | 0.89 ± 0.0 a | 0.74 ± 0.09 a | 0.69 ± 0.03 a | 0.9 ± 0.14 b | 0.44 ± 0.1 a | 0.43 ± 0.06 a |
| Pentadecanoic (C16:3) | 1.0 ± 0.5 a | 0.59 ± 0.2 a | 0.9 ± 0.4 a | 0.52 ± 0.2 a | 0.73 ± 0.3 a | 0.6 ± 0.2 a |
| Octadecanoic (C18:1) | 3.73 ± 0. a | 3.96 ± 0.8 a | 2.97 ± 0.4 a | 4.45 ± 0.6 b | 3.46 ± 0.6 a | 4.48 ± 0.7 b |
| Octadecanoic (C18:2) | 21.5 ± 0.9 a | 24.2 ± 1.4 a | 25.1 ± 1.9 a | 22.5 ± 0.8 a | 24.2 ± 1 a | 20.8 ± 0.5 a |
| Heptadecanoic (C18:3) | 12.1 ± 3. a | 25.64 ± 2.3 b | 23.3 ± 3.3 a | 29.68 ± 6 b | 25.81 ± 4.1 a | 26.52 ± 0.4 a |
| Heptadecanoic (C18:4) | 1.0 ± 0.06 a | 1.27 ± 0.14 a | 1.04 ± 0.3 a | 1.2 ± 0.08 a | 1.13 ± 0.2 a | 1.16 ± 0.09 a |
| Tetracosanoic (C20:3) | 0.11 ± 0.01 a | 0.26 ± 0.0 b | 0.22 ± 0.07 a | 0.23 ± 0.04 a | 0.2 ± 0.06 a | 0.19 ± 0.04 a |
| C. ambrosoides | C. ficifolium | C. botrys | ||||||
|---|---|---|---|---|---|---|---|---|
| Control | Endophyte | Control | Endophyte | Control | Endophyte | Extraction Solvent * | Cipro-Floxacin | |
| Antioxidant Activities | ||||||||
| FRAP (mg gallic acid/gFW) | 16.2 ± 1.2 a | 37.5 ± 1 c | 18.3 ± 0.6 b | 33.06 ± 3 c | 11.5 ± 0.1 a | 21.4 ± 6 c | 0.5 ± 0 | - |
| ABTS (µmol trolox/gFW) | 2.0 ± 0.1 a | 1.9 ± 0.2 b | 1.37 ± 0.1 a | 1.9 ± 0.01 b | 0.93 ± 0.1 a | 2.2 ± 0.3 b | 0.7 ± 0 | - |
| DPPH% | 18.8 ± 0.7 a | 13.7 ± 1 a | 12.2 ± 1 a | 16.37 ± 3 b | 12.7 ± 1 a | 17.06 ± 3 b | 0.1 ± 0 | - |
| C. ambrosoides | C. ficifolium | C. botrys | ||||||
|---|---|---|---|---|---|---|---|---|
| Control | Endo | Control | Endo | Control | Endo | Ex. Solvent | Cipro-Floxacin | |
| Antibacterial Activities (Zone Inhibition, mm) | ||||||||
| S. saprophyticus ATCC 19701 | 14.3 ± 0.4 a | 25.5 ± 1 b | 16.5 ± 0.3 a | 20 ± 2.3 a | 19 ± 0.1 a | 22.5 ± 0.6 b | 1.5 ± 0.1 | 33.4 ± 2.3 |
| S. epidermidis ATCC 12228 | 10.9 ± 0.3 a | 22.0 ± 1 b | 14.5 ± 0.9 a | 17.6 ± 2 ab | 16. ± 0.3 b | 19.92 ± 1 ab | 0.9 ± 0 | 23.6 ± 5 |
| E. faecalis ATCC 10541 | 15.0 ± 0.3 a | 20 ± 0.9 b | 17.8 ± 0.4 a | 15 ± 1.7 ab | 16 ± 0.1 a | 19.1 ± 1 ab | 1.1 ± 0 | 21.5 ± 4.0 |
| S. salivarius ATCC25975 | 13.2 ± 0.8 a | 17 ± 0.4 ab | 16.2 ± 0.3 a | 20 ± 0.4 a | 13 ± 0.4 a | 17.09 ± 1 ab | 0.89 ± 0 | 30.6 ± 2.4 |
| E. coli ATCC 29998 | 14.52 ± 1 a | 18.3 ± 1 ab | 19.5 ± 0.6 a | 21 ± 0.7 a | 16 ± 1.0 a | 19.29 ± 1 ab | 1.2 ± 0 | 25.9 ± 0.7 |
| S.typhimurium ATCC14028 | 16.2 ± 0.3 a | 20.6 ± 1 ab | 19.1 ± 0 a | 23 ± 0.6 b | 18 ± 0.2 a | 20.5 ± 1 ab | 0.5 ± 0 | 22.1 ± 0.6 |
| P. aeruginosa ATCC10145 | 16.9 ± 0.5 a | 33 ± 1.6 b | 20.2 ± 0.7 a | 22 ± 1 a | 24 ± 0.2 a | 29 ± 1 b | 1 ± 0 | 22.8 ± 1 a |
| P. vulgaris ATCC8427 | 16. ± 0.4 a | 26.3 ± 1 b | 18.7 ± 0.4 a | 17 ± 0.5 a | 20 ± 0.1 a | 23.9 ± 0.8 a | 1.9 ± 0.1 | 27.1 ± 1.5 |
| E.r aerogenes ATCC 13048 | 15.4 ± 0.3 a | 28.6 ± 1 b | 17.8 ± 0.4 a | 22 ± 0.4 b | 21 ± 0.2 a | 25 ± 0.7 a | 0.7 ± 0.0 | 24 ± 0.7 |
| S. marcescens ATCC99006 | 1.9 ± 0.06 a | 7.3 ± 0.6 b | 2.2 ± 0.05 a | 5.9 ± 0.5 b | 4 ± 0.05 a | 5.5 ± 0.3 a | 0.9 ± 0.1 | 8.1 ± 0.5 |
| S. typhimurium ATCC14028 | 16 ± 0.5 a | 10 ± 0.5 ab | 19.2 ± 0.4 a | 17 ± 0.4 a | 12 ± 0.1 a | 13.89 ± 1 a | 1.1 ± 0 | 22 ± 0.2 |
| C. albicans ATCC90028 | 6.9 ± 0.2 a | 9.8 ± 0.5 b | 8.0 ± 0.19 a | 7.9 ± 0.9 a | 8.1 ± 0.1 a | 9.3 ± 0.4 a | 0.07 ± 0 | 8.9 ± 0.4 |
| C. glabrata ATCC90030 | 2.3 ± 0.07 a | 4.8 ± 0.2 b | 2.9 ± 0.14 a | 3.1 ± 0.1 a | 3 ± 0.03 a | 4.18 ± 0.1 a | 0.2 ± 0 | 4.0 ± 0.01 |
| A. flavus ATCC9170 | 22 ± 0.5 a | 43 ± 1.1 b | 25.7 ± 0.6 a | 33.2 ± 5 b | 29 ± 0.3 a | 36 ± 1 b | 20.5 ± 1 | 33.29 ± 5 b |
| C. ambrosoides | C. ficifolium | C. botrys | ||||
|---|---|---|---|---|---|---|
| Anti-Inflammatory | Control | Endo | Control | Endo | Control | Endo |
| Cyclooxygenase-2 (µg/mL) | 1.1 ± 0.2 a | 0.73 ± 0.0 ab | 1.2 ± 0.1 a | 0.6 ± 0.04 b | 1.3 ± 1 a | 0.45 ± 0 c |
| Lipoxygenase (µg/mL) | 7.2 ± 0.1 a | 5.7 ± 0.5 ab | 6.7 ± 4 a | 4.6 ± 0.6 b | 6.8 ± 0.7 a | 3.1 ± 0.6 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almuhayawi, M.S.; Abdel-Mawgoud, M.; Al Jaouni, S.K.; Almuhayawi, S.M.; Alruhaili, M.H.; Selim, S.; AbdElgawad, H. Bacterial Endophytes as a Promising Approach to Enhance the Growth and Accumulation of Bioactive Metabolites of Three Species of Chenopodium Sprouts. Plants 2021, 10, 2745. https://doi.org/10.3390/plants10122745
Almuhayawi MS, Abdel-Mawgoud M, Al Jaouni SK, Almuhayawi SM, Alruhaili MH, Selim S, AbdElgawad H. Bacterial Endophytes as a Promising Approach to Enhance the Growth and Accumulation of Bioactive Metabolites of Three Species of Chenopodium Sprouts. Plants. 2021; 10(12):2745. https://doi.org/10.3390/plants10122745
Chicago/Turabian StyleAlmuhayawi, Mohammed S., Mohamed Abdel-Mawgoud, Soad K. Al Jaouni, Saad M. Almuhayawi, Mohammed H. Alruhaili, Samy Selim, and Hamada AbdElgawad. 2021. "Bacterial Endophytes as a Promising Approach to Enhance the Growth and Accumulation of Bioactive Metabolites of Three Species of Chenopodium Sprouts" Plants 10, no. 12: 2745. https://doi.org/10.3390/plants10122745