Biomass Functions and Carbon Content Variabilities of Natural and Planted Pinus koraiensis in Northeast China




Abstract
:1. Introduction
2. Materials and Methods
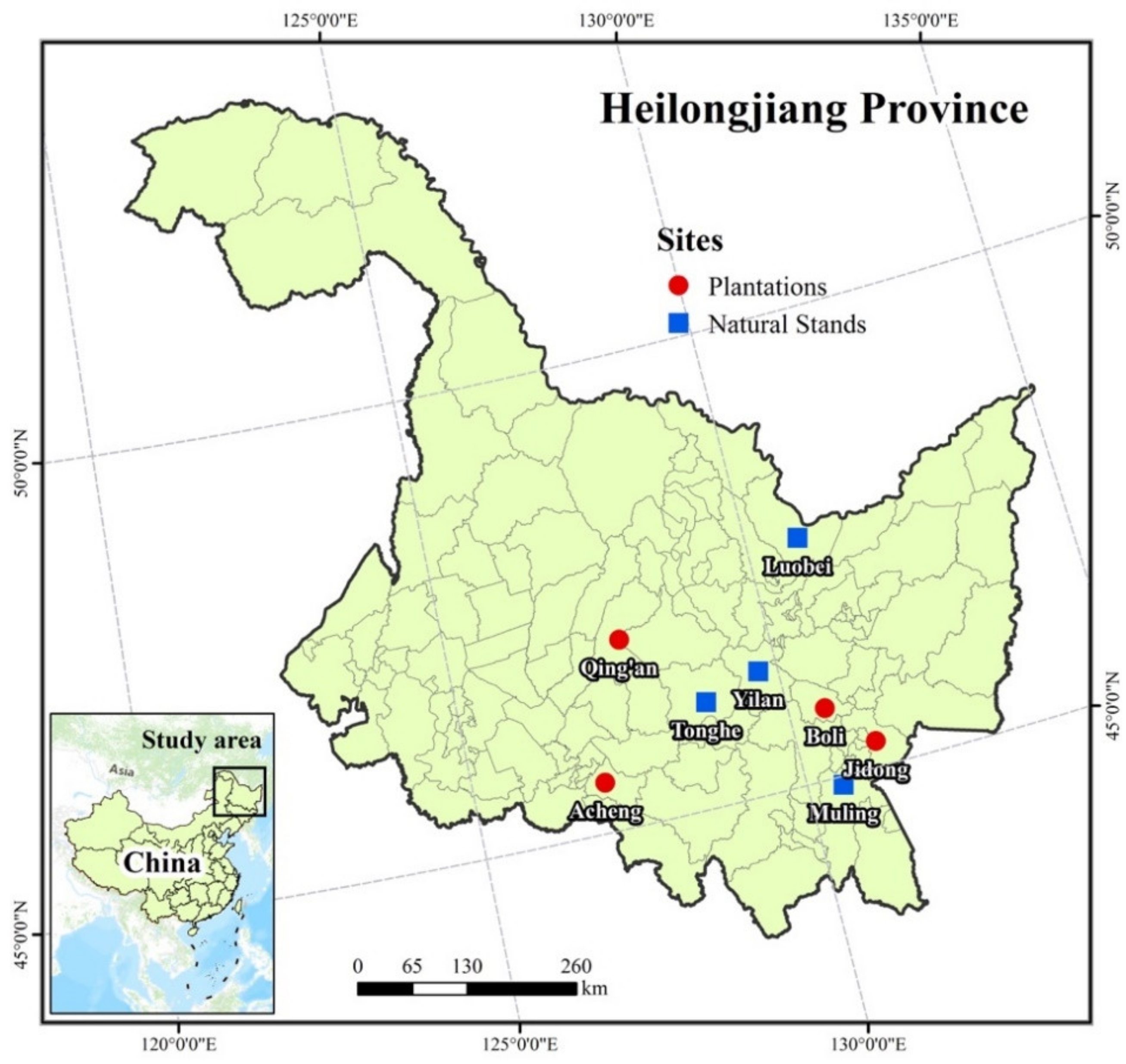
2.1. Data Collection
2.2. Statistical Analysis
2.2.1. Base Model
2.2.2. Population Average (Generalized) Model
2.2.3. Origin-Based Dummy Variable Model
2.2.4. Heteroscedasticity Neutralization
2.2.5. Model Validation
2.2.6. Variation of Carbon Content
3. Results
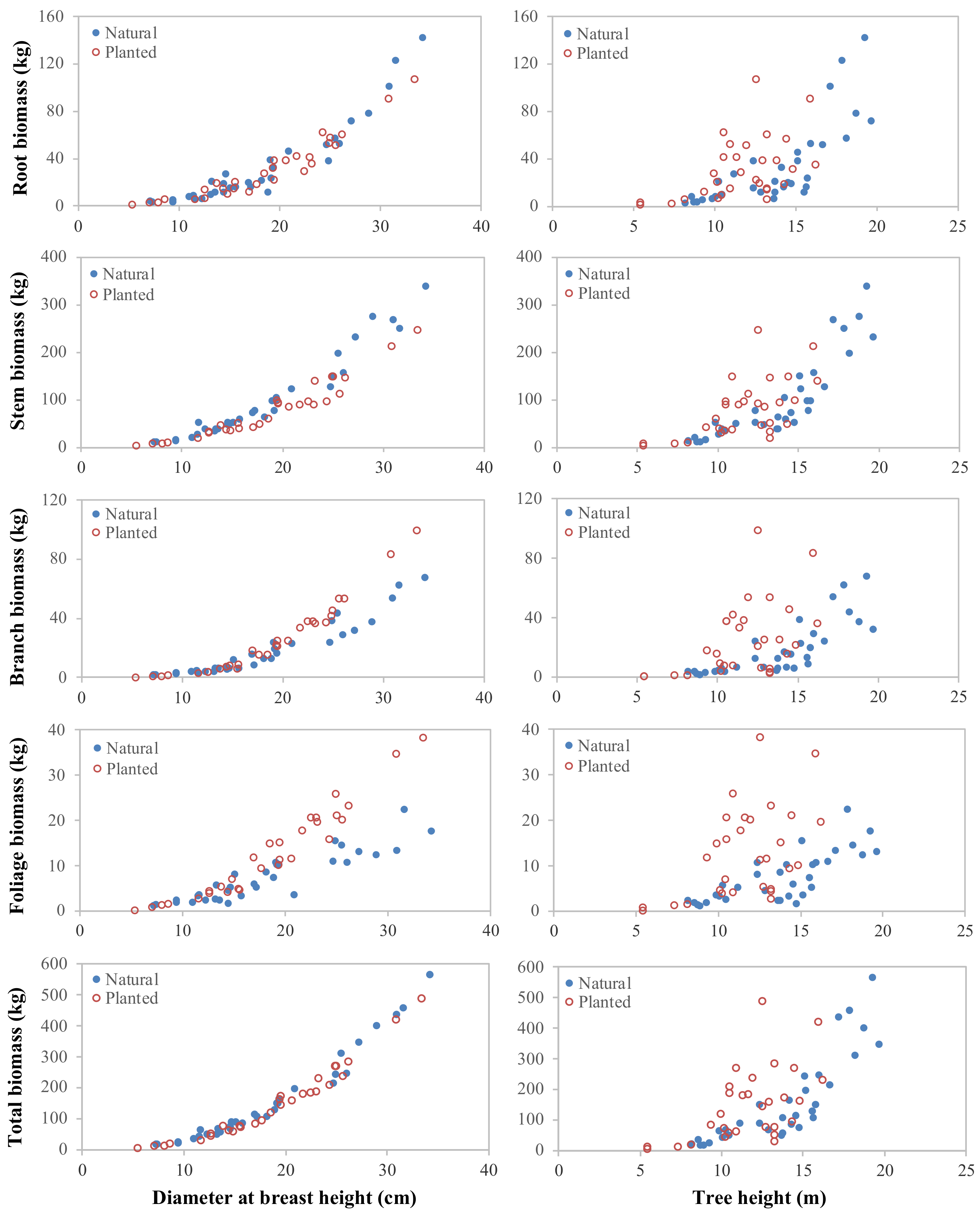
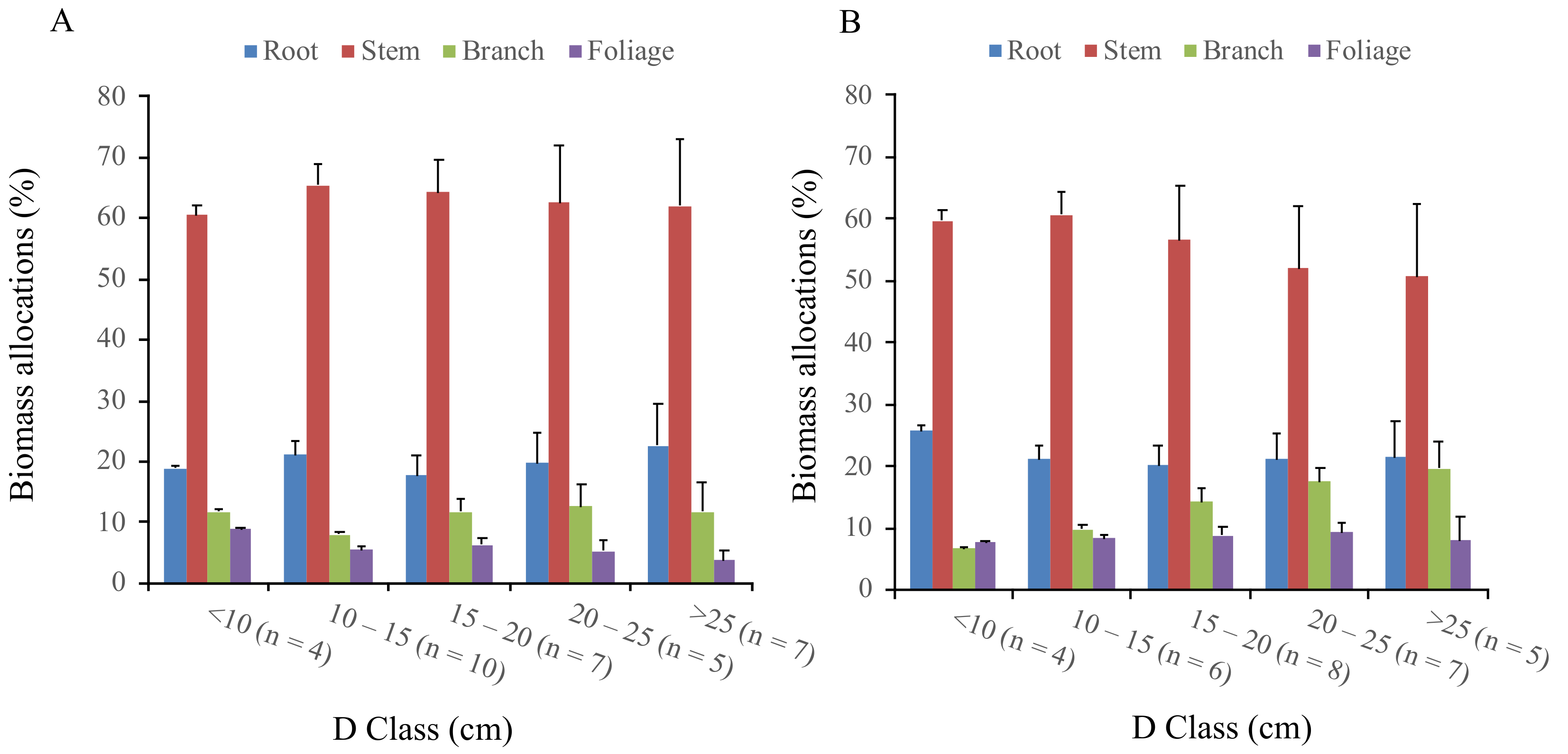
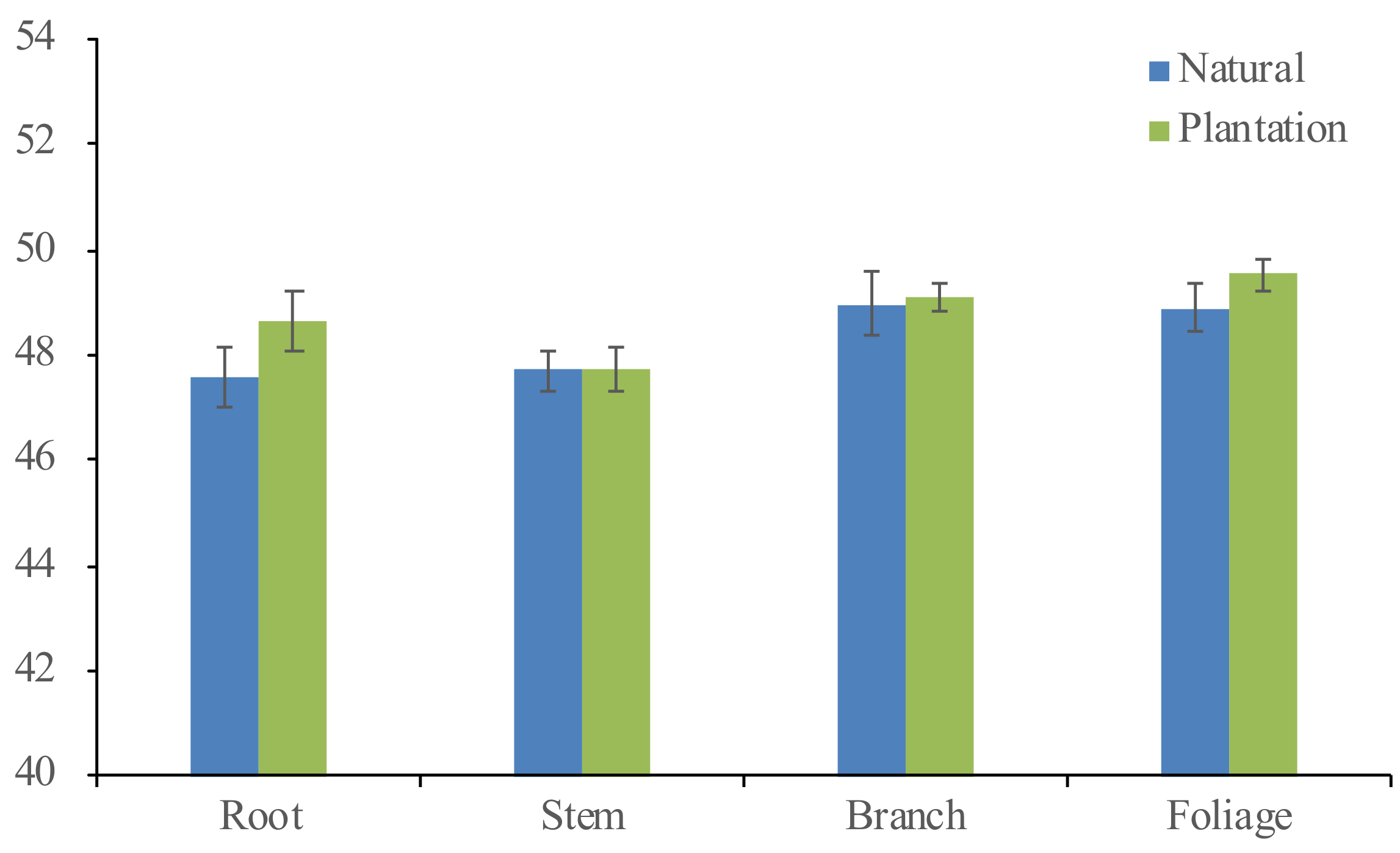
3.1. Biomass Partitioning between the Planted and Naturally Regenerated Korean Pine
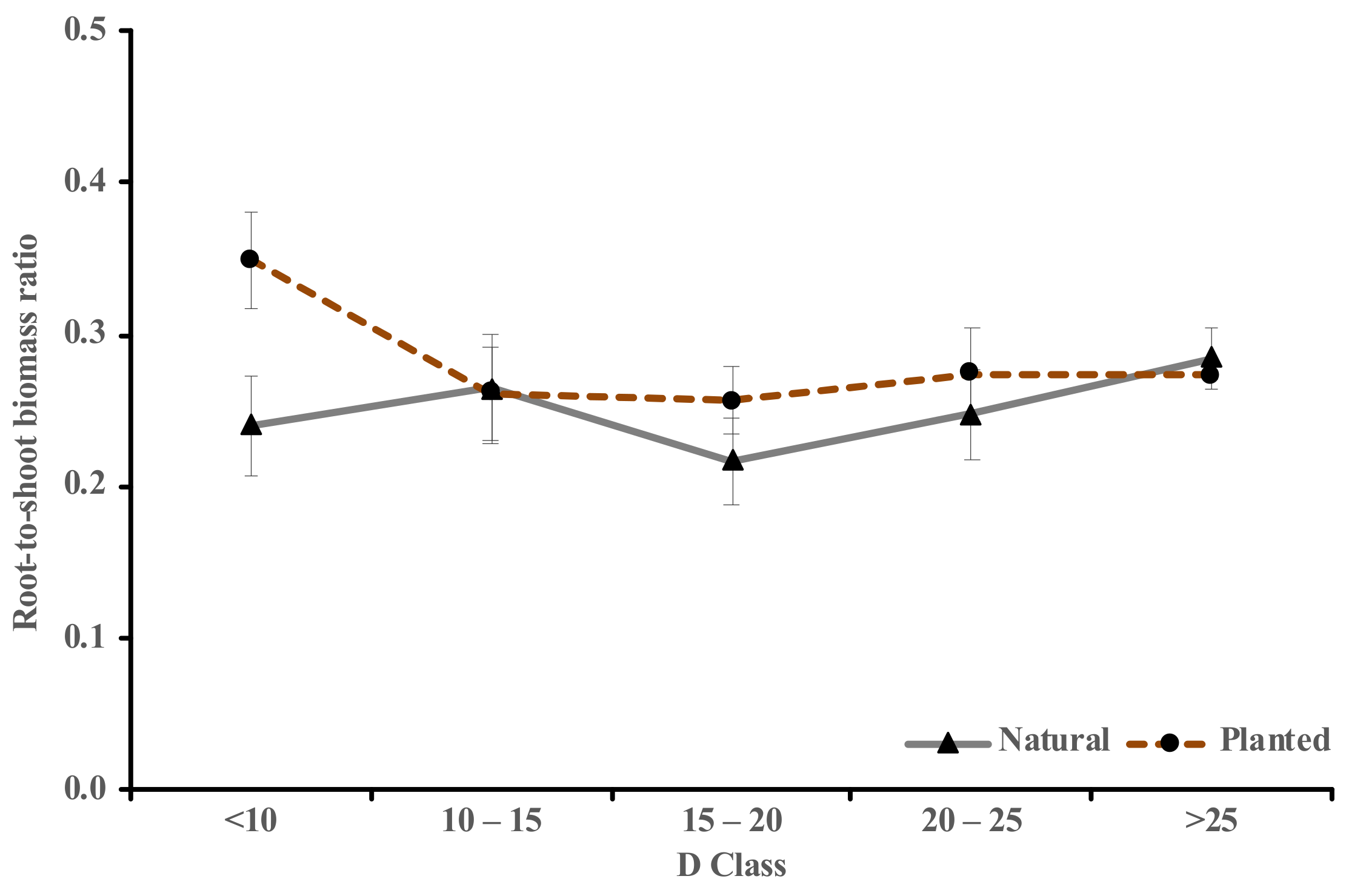
3.2. RS Ratio and Carbon Content Variations of the Two Origins
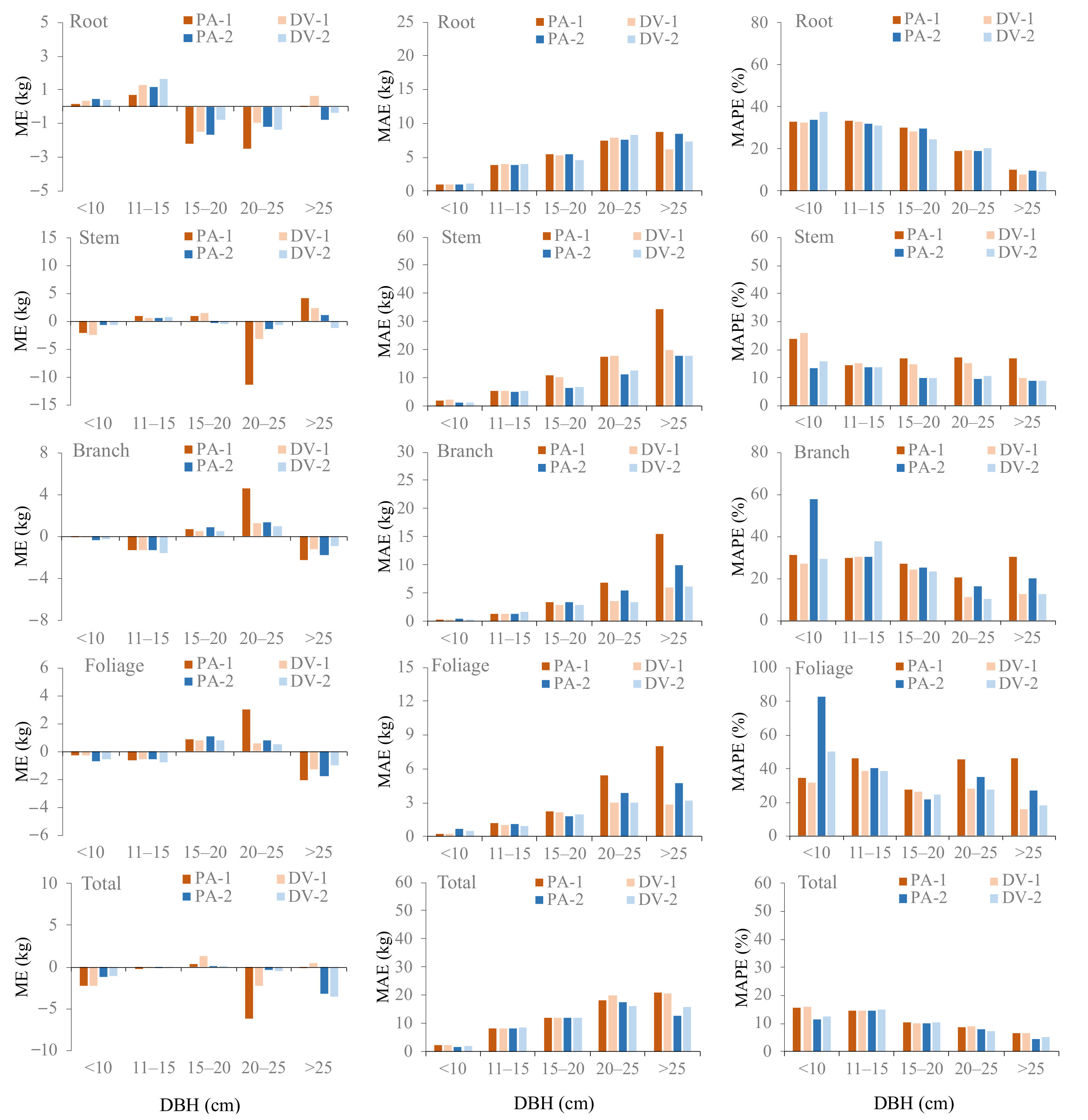
3.3. The PA and DV Additive Biomass Model Systems
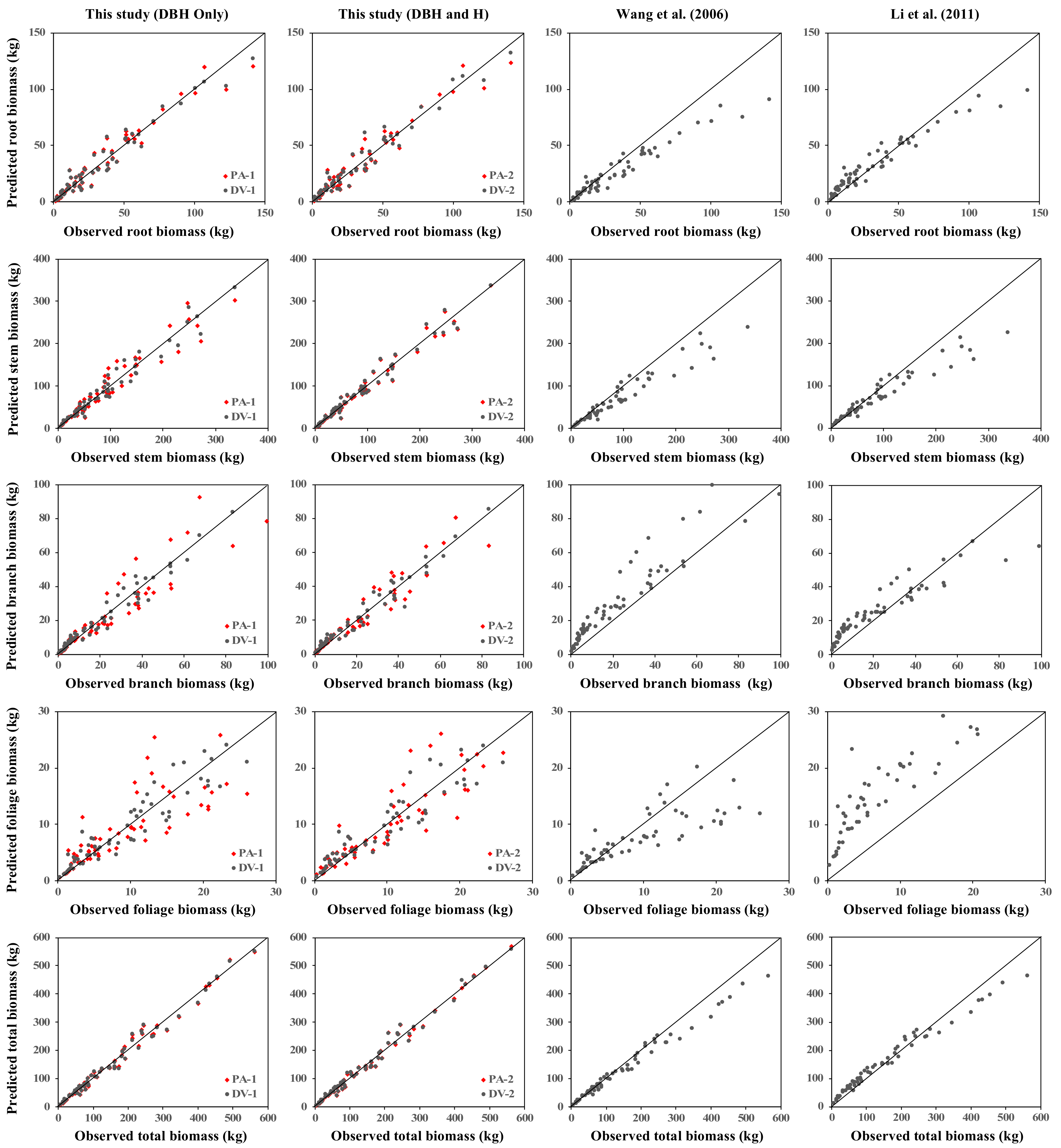
3.4. Comparison with the Established Pinus Koraiensis Biomass Models
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Erb, K.H.; Kastner, T.; Plutzar, C.; Bais, A.L.S.; Carvalhais, N.; Fetzel, T.; Gingrich, S.; Haberl, H.; Lauk, C.; Niedertscheider, M.; et al. Unexpectedly large impact of forest management and grazing on global vegetation biomass. Nature 2018, 553, 73–76. [Google Scholar] [CrossRef]
- Köhl, M.; Lasco, R.; Cifuentes, M.; Jonsson, Ö.; Korhonen, K.T.; Mundhenk, P.; de Jesus Navar, J.; Stinson, G. Changes in forest production, biomass and carbon: Results from the 2015 UN FAO Global Forest Resource Assessment. For. Ecol. Manag. 2015, 352, 21–34. [Google Scholar] [CrossRef] [Green Version]
- Poorter, L.; van der Sande, M.T.; Thompson, J.; Arets, E.J.M.M.; Alarcón, A.; Álvarez-Sánchez, J.; Ascarrunz, N.; Balvanera, P.; Barajas-Guzmán, G.; Boit, A.; et al. Diversity enhances carbon storage in tropical forests. Glob. Ecol. Biogeogr. 2015, 24, 1314–1328. [Google Scholar] [CrossRef]
- Hernandez, J.O.; Maldia, L.S.J.; Park, B.B. The Impacts of Windstorms on Forests in Tropical, Subtropical, and Temperate Zones: Where Are We Now and How Should Research Move Forward? Plants 2020, 9, 1709. [Google Scholar] [CrossRef]
- Thompson, L.G. Climate change: The evidence and our options. Behav. Anal. 2010, 33, 153–170. [Google Scholar] [CrossRef] [PubMed]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.C.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef] [PubMed]
- Kusmana, C.; Hidayat, T.; Tiryana, T.; Rusdiana, O. Istomo Allometric models for above- and below-ground biomass of Sonneratia spp. Glob. Ecol. Conserv. 2018, 15, e00417. [Google Scholar] [CrossRef]
- Bi, H.; Turner, J.; Lambert, M.J. Additive biomass equations for native eucalypt forest tress of temperate Australia. Trees 2004, 18, 467–479. [Google Scholar] [CrossRef]
- Zeng, W.S. Using nonlinear mixed model and dummy variable model approaches to develop origin-based individual tree biomass equations. Trees Struct. Funct. 2015, 29, 275–283. [Google Scholar] [CrossRef]
- Usoltsev, V.; Danilin, I.; Tsogt, Z.; Osmirko, A.A.; Tsepordey, I.S.; Chasovskikh, V.P. Aboveground biomass of mongolian larch (Larix sibiricledeb) forests in the eurasian region. Geogr. Environ. Sustain. 2019, 12, 117–132. [Google Scholar] [CrossRef] [Green Version]
- Cunia, T.; Briggs, R.D. Forcing additivity of biomass tables: Some empirical results. Can. J. For. Res. 1984, 14, 376–384. [Google Scholar] [CrossRef]
- Parresol, B.R. Assessing tree and stand biomass: A review with examples and critical comparisons. For. Sci. 1999, 45, 573–593. [Google Scholar] [CrossRef]
- Affleck, D.L.R.; Diéguez-Aranda, U. Additive nonlinear biomass equations: A likelihood-based approach. For. Sci. 2016, 62, 129–140. [Google Scholar] [CrossRef]
- Dong, L.; Zhang, L.; Li, F. A three-step proportional weighting system of nonlinear biomass equations. For. Sci. 2015, 61, 35–45. [Google Scholar] [CrossRef]
- Parresol, B.R. Additivity of nonlinear biomass equations. Can. J. For. Res. 2001, 31, 865–878. [Google Scholar] [CrossRef]
- Zhao, D.; Westfall, J.; Coulston, J.W.; Lynch, T.B.; Bullock, B.P.; Montes, C.R. Additive biomass equations for slash pine trees: Comparing three modeling approaches. Can. J. For. Res. 2019, 49, 27–40. [Google Scholar] [CrossRef]
- Li, H.; Zhao, P. Improving the accuracy of tree-level aboveground biomass equations with height classification at a large regional scale. For. Ecol. Manag. 2013, 289, 153–163. [Google Scholar] [CrossRef]
- Menéndez-Miguélez, M.; Canga, E.; Barrio-Anta, M.; Majada, J.; Álvarez-Álvarez, P. A three level system for estimating the biomass of Castanea sativa Mill. coppice stands in north-west Spain. For. Ecol. Manag. 2013, 291, 417–426. [Google Scholar] [CrossRef]
- Zhao, D.; Kane, M.; Markewitz, D.; Teskey, R.; Clutter, M. Additive tree biomass equations for midrotation loblolly pine plantations. For. Sci. 2015, 61, 613–623. [Google Scholar] [CrossRef] [Green Version]
- Widagdo, F.R.A.; Li, F.; Zhang, L.; Dong, L. Aggregated biomass model systems and carbon concentration variations for tree carbon quantification of natural mongolian oak in northeast China. Forests 2020, 11, 397. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Soalleiro, R.; Eimil-Fraga, C.; Gómez-García, E.; García-Villabrille, J.D.; Rojo-Alboreca, A.; Muñoz, F.; Oliveira, N.; Sixto, H.; Pérez-Cruzado, C. Exploring the factors affecting carbon and nutrient concentrations in tree biomass components in natural forests, forest plantations and short rotation forestry. For. Ecosyst. 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.; Widagdo, F.R.A.; Xie, L.; Li, F. Biomass and volume modeling along with carbon concentration variations of short-rotation poplar plantations. Forests 2020, 11, 780. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, D.; Liu, G.; Yang, C.; Teskey, R.O. Additive tree biomass equations for Betula platyphylla Suk. plantations in Northeast China. Ann. For. Sci. 2018, 75. [Google Scholar] [CrossRef] [Green Version]
- Widagdo, F.R.A.; Xie, L.; Dong, L.; Li, F. Origin-based biomass allometric equations, biomass partitioning, and carbon concentration variations of planted and natural Larix gmelinii in northeast China. Glob. Ecol. Conserv. 2020, 23, e01111. [Google Scholar] [CrossRef]
- FAO. Global Forest Resources Assessment 2015: How are the World’s Forest Changing? FAO Inter Departmental Working: Rome, Italy, 2015; ISBN 9789251088210. [Google Scholar]
- Licata, J.A.; Gyenge, J.E.; Fernández, M.E.; Schlichter, T.M.; Bond, B.J. Increased water use by ponderosa pine plantations in northwestern Patagonia, Argentina compared with native forest vegetation. For. Ecol. Manag. 2008, 255, 753–764. [Google Scholar] [CrossRef]
- Guo, Q.; Ren, H. Productivity as related to diversity and age in planted versus natural forests. Glob. Ecol. Biogeogr. 2014, 23, 1461–1471. [Google Scholar] [CrossRef]
- Zarafshar, M.; Bazot, S.; Matinizadeh, M.; Bordbar, S.K.; Rousta, M.J.; Kooch, Y.; Enayati, K.; Abbasi, A.; Negahdarsaber, M. Do tree plantations or cultivated fields have the same ability to maintain soil quality as natural forests? Appl. Soil Ecol. 2020, 151, 103536. [Google Scholar] [CrossRef]
- Erdle, T.; Pollard, J. Are plantations changing the tree species composition of New Brunswick’s forest? For. Chron. 2002, 78, 812–821. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.Y.; Zeng, W.S.; Tang, S.Z.; Sharma, R.P.; Li, H.K. Using linear mixed model and dummy variable model approaches to construct compatible single-tree biomass equations at different scales—A case study for Masson pine in Southern China. J. For. Sci. 2012, 58, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Schuler, J.; Bragg, D.C.; Mcelligott, K. Biomass Estimates of Small Diameter Planted and Natural-Origin Loblolly Pines Show Major Departures from the National Biomass Estimator Equations. For. Sci. 2017, 63, 319–330. [Google Scholar] [CrossRef] [Green Version]
- Thomas, P.; Farjon, A. IUCN Red List Threat.Species 2013 e.T42373A2975987. Pinus koraiensis. 2013. Available online: https://www.iucnredlist.org/species/42373/2975987 (accessed on 7 December 2020).
- Kaviriri, D.K.; Zhang, Q.; Zhang, X.; Jiang, L.; Zhang, J.; Wang, J.; Khasa, D.P.; You, X.; Zhao, X. Phenotypic variability and genetic diversity in a Pinus koraiensis clonal trial in northeastern china. Genes 2020, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Adelina, N.M.; Wang, H.; Zhang, L.; Zhao, Y. Comparative analysis of volatile profiles in two grafted pine nuts by headspace-SPME/GC-MS and electronic nose as responses to different roasting conditions. Food Res. Int. 2020, 110026. [Google Scholar] [CrossRef]
- State Forestry and Grassland Administration. The Ninth Forest Resources Survey Report (2014–2018); China Forestry Press: Beijing, China, 2019. [Google Scholar]
- Kaviriri, D.K.; Li, Y.; Zhang, D.; Li, H.; Fan, Z.; Wang, J.; Wang, L.; Wang, Q.; Wang, D.; Chiang, V.L.; et al. Clonal variations in cone, seed and nut traits in a Pinus koraiensis seed orchard in Northeast China. J. For. Res. 2020, 32. [Google Scholar] [CrossRef]
- Wang, C. Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests. For. Ecol. Manag. 2006, 222, 9–16. [Google Scholar] [CrossRef]
- Li, X.; Yi, M.J.; Son, Y.; Park, P.S.; Lee, K.H.; Son, Y.M.; Kim, R.H.; Jeong, M.J. Biomass and Carbon Storage in an Age-Sequence of Korean Pine (Pinus koraiensis) Plantation Forests in Central Korea. J. Plant Biol. 2011, 54, 33–42. [Google Scholar] [CrossRef]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future köppen-geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, L.; Zhang, L.; Li, F. Allometry and partitioning of individual tree biomass and carbon of Abies nephrolepis Maxim in northeast China. Scand. J. For. Res. 2016, 31, 399–411. [Google Scholar] [CrossRef]
- Xie, L.; Li, F.; Zhang, L.; Widagdo, F.R.A.; Dong, L. A Bayesian Approach to Estimating Seemingly Unrelated Regression for Tree Biomass Model Systems. Forests 2020, 11, 1302. [Google Scholar] [CrossRef]
- Bi, H.; Murphy, S.; Volkova, L.; Weston, C.; Fairman, T.; Li, Y.; Law, R.; Norris, J.; Lei, X.; Caccamo, G. Additive biomass equations based on complete weighing of sample trees for open eucalypt forest species in south-eastern Australia. For. Ecol. Manag. 2015, 349, 106–121. [Google Scholar] [CrossRef]
- Usoltsev, V.A.; Reza, S.S.O.; Stepanovich, T.I.; Amirhossein, A.; Meng, Z.; Anees, S.A.; Petrovich, C.V. Are There Differences in the Response of Natural Stand and Plantation Biomass to Changes in Temperature and Precipitation? A Case for Two-Needled Pines in Eurasia. J. Resour. Ecol. 2020, 11, 331. [Google Scholar] [CrossRef]
- Harvey, A.C. Estimating Regression Models with Multiplicative Heteroscedasticity. Econometrica 1977, 44. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/ETS 9.3 User’s Guide; SAS Institute Inc: Cary, NC, USA, 2011. [Google Scholar]
- Balboa-Murias, M.Á.; Rodríguez-Soalleiro, R.; Merino, A.; Álvarez-González, J.G. Temporal variations and distribution of carbon stocks in aboveground biomass of radiata pine and maritime pine pure stands under different silvicultural alternatives. For. Ecol. Manag. 2006, 237, 29–38. [Google Scholar] [CrossRef]
- Barrio-Anta, M.; Balboa-Murias, M.Á.; Castedo-Dorado, F.; Diéguez-Aranda, U.; Álvarez-González, J.G. An ecoregional model for estimating volume, biomass and carbon pools in maritime pine stands in Galicia (Northwestern Spain). For. Ecol. Manag. 2006, 223, 24–34. [Google Scholar] [CrossRef]
- Bates, D.M.; Watts, D.G. Nonlinear Regression Analysis and Its Applications; John Wiley & Sons, Ltd: New York, NY, USA, 1988. [Google Scholar]
- Wang, X.W.; Weng, Y.H.; Liu, G.F.; Krasowski, M.J.; Yang, C.P. Variations in carbon concentration, sequestration and partitioning among Betula platyphylla provenances. For. Ecol. Manag. 2015, 358, 344–352. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, C.; Wang, X.; Quan, X. Carbon concentration variability of 10 Chinese temperate tree species. For. Ecol. Manag. 2009, 258, 722–727. [Google Scholar] [CrossRef]
- Ruiz-Peinado, R.; Montero, G.; del Rio, M. Biomass models to estimate carbon stocks for hardwood tree species. For. Syst. 2012, 21, 42–52. [Google Scholar] [CrossRef]
- Mokany, K.; Raison, R.J.; Prokushkin, A.S. Critical analysis of root: Shoot ratios in terrestrial biomes. Glob. Chang. Biol. 2006, 12, 84–96. [Google Scholar] [CrossRef]
- Wu, J.; Hong, J.; Wang, X.; Sun, J.; Lu, X.; Fan, J.; Cai, Y. Biomass partitioning and its relationship with the environmental factors at the alpine steppe in northern Tibet. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Jin, X.; Pukkala, T.; Li, F.; Dong, L. Optimal management of Korean pine plantations in multifunctional forestry. J. For. Res. 2017, 28, 1027–1037. [Google Scholar] [CrossRef]
- Eisa, A.; Malenica, K.; Schwenkert, S.; Bölter, B. High Light Acclimation Induces Chloroplast Precursor Phosphorylation and Reduces Import Efficien. Plants 2020, 9, 24. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, R.T. Distinct responses to light in plants. Plants 2020, 9, 894. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, Ü. A review of light interception in plant stands from leaf to canopy in different plant functional types and in species with varying shade tolerance. Ecol. Res. 2010, 25, 693–714. [Google Scholar] [CrossRef]
- Bloom, A.J.; Chapin, F.S.; Mooney, H.A. Resource limitation in plants—An economic analogy. Annu. Rev. Ecol. Syst. 1985, 16, 363–392. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Návar, J. Root stock biomass and productivity assessments of reforested pine stands in northern Mexico. For. Ecol. Manag. 2015, 338, 139–147. [Google Scholar] [CrossRef]
- Cairns, M.A.; Brown, S.; Helmer, E.H.; Baumgardner, G.A. Root biomass allocation in the world’s upland forests. Oecologia 1997, 111, 1–11. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, C.; Xia, F.; Zhao, X.; Wu, L.; von Gadow, K. Biomass structure and allometry of Abies nephrolepis (Maxim) in northeast China. Silva Fenn. 2011, 45, 211–226. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Wang, X.; Zhang, X.; Booth, T.H.; Lu, F. Root: Shoot ratios across China’s forests: Forest type and climatic effects. For. Ecol. Manag. 2012, 269, 19–25. [Google Scholar] [CrossRef]
- Hermans, C.; Hammond, J.P.; White, P.J.; Verbruggen, N. How do plants respond to nutrient shortage by biomass allocation? Trends Plant. Sci. 2006, 11, 610–617. [Google Scholar] [CrossRef]
- Qi, Y.; Wei, W.; Chen, C.; Chen, L. Plant root-shoot biomass allocation over diverse biomes: A global synthesis. Glob. Ecol. Conserv. 2019, 18, e00606. [Google Scholar] [CrossRef]
- Elias, M.; Potvin, C. Assessing inter- and intra-specific variation in trunk carbon concentration for 32 neotropical tree species. Can. J. For. Res. 2003, 33, 1039–1045. [Google Scholar] [CrossRef]
- Zou, W.T.; Zeng, W.S.; Zhang, L.J.; Zeng, M. Modeling crown biomass for four pine species in China. Forests 2015, 6, 433. [Google Scholar] [CrossRef] [Green Version]
- Weisheng, Z. Comparison of three allometric equations for biomass modeling. Cent. South. For. Invent. Plan. 2014, 33, 1–3. (In Chinese) [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Naturally Regenerated (N = 33) | Planted (N = 30) | Total (N = 63) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Mean | Range | S.D. | Mean | Range | S.D. | Mean | Range | S.D. | |
| Tree DBH (cm) | 18.2 | 7.3–34.4 | 7.3 | 18.4 | 5.4–33.4 | 6.9 | 18.3 | 5.4–34.4 | 7.0 |
| Total height (m) | 13.7 | 8.2–19.7 | 3.3 | 11.5 | 5.4–16.2 | 2.7 | 12.7 | 5.4–19.7 | 3.2 |
| Root biomass (kg) | 32.7 | 2.3–141.4 | 35.0 | 31.3 | 1.6–107.0 | 26.0 | 32.0 | 1.6–141.4 | 30.8 |
| Stem biomass (kg) | 97.3 | 8.3–336.9 | 87.9 | 79.1 | 3.4–246.5 | 60.0 | 88.6 | 3.4–336.9 | 75.8 |
| Branch biomass (kg) | 17.7 | 1.4–67.4 | 18.1 | 25.2 | 0.3–99.3 | 24.6 | 21.3 | 0.3–99.3 | 21.6 |
| Foliage biomass (kg) | 7.3 | 1.1–22.4 | 5.4 | 12.9 | 0.3–38.3 | 10.0 | 9.9 | 0.3–38.3 | 8.3 |
| Total biomass (kg) | 155.0 | 14.0–563.1 | 144.1 | 148.6 | 5.6–491.2 | 118.8 | 152.0 | 5.6–563.1 | 131.6 |
| Equations | Components | Parameters | Estimates | S.E. |
|---|---|---|---|---|
| PA-1 (One-variable) | Root | −4.10368 | 0.1765 | |
| 2.52639 | 0.0548 | |||
| Stem | −2.29469 | 0.2002 | ||
| 2.26941 | 0.0644 | |||
| Branch | −5.40818 | 0.3448 | ||
| 2.79699 | 0.1066 | |||
| Foliage | −3.73742 | 0.4135 | ||
| 2.01881 | 0.1325 | |||
| PA-2 (Two-variables) | Root | −4.47742 | 0.2138 | |
| 2.51914 | 0.0720 | |||
| 0.14399 | 0.1051 | |||
| Stem | −3.36539 | 0.2068 | ||
| 1.94692 | 0.0661 | |||
| 0.77835 | 0.0914 | |||
| Branch | −4.14573 | 0.3243 | ||
| 3.13166 | 0.1041 | |||
| −0.86857 | 0.1251 | |||
| Foliage | −1.96307 | 0.3978 | ||
| 2.56451 | 0.1310 | |||
| −1.31504 | 0.1849 |
| Equations | Origins | Components | Parameters | Estimates | S.E. | |
|---|---|---|---|---|---|---|
| DV-1 (One-variable) | Natural | Root | −4.58785 | 0.3204 | ||
| 2.67952 | 0.0994 | |||||
| Stem | −2.25782 | 0.2153 | ||||
| 2.28230 | 0.0652 | |||||
| Branch | −4.95561 | 0.3411 | ||||
| 2.60134 | 0.1051 | |||||
| Foliage | −3.18675 | 0.4247 | ||||
| 1.75082 | 0.1385 | |||||
| Planted | Root | −3.81245 | 0.3607 | |||
| 2.41863 | 0.1154 | |||||
| Stem | −2.23070 | 0.2973 | ||||
| 2.20883 | 0.0918 | |||||
| Branch | −5.82831 | 0.3084 | ||||
| 2.99239 | 0.0953 | |||||
| Foliage | −4.26419 | 0.3503 | ||||
| 2.27914 | 0.1114 | |||||
| DV-2 (Two-variables) | Natural | Root | −4.24899 | 0.5111 | ||
| 3.02592 | 0.2160 | |||||
| −0.51898 | 0.3727 | |||||
| Stem | −3.40236 | 0.3582 | ||||
| 1.85305 | 0.1258 | |||||
| 0.90319 | 0.2300 | |||||
| Branch | −3.63392 | 0.5838 | ||||
| 3.07384 | 0.2351 | |||||
| −1.01454 | 0.4008 | |||||
| Foliage | −1.53751 | 0.8336 | ||||
| 2.34938 | 0.4170 | |||||
| −1.27838 | 0.6761 | |||||
| Planted | Root | −3.64574 | 0.3971 | |||
| 2.47972 | 0.1166 | |||||
| −0.14226 | 0.2108 | |||||
| Stem | −3.33091 | 0.4193 | ||||
| 2.05227 | 0.0976 | |||||
| 0.62892 | 0.1616 | |||||
| Branch | −5.77974 | 0.4322 | ||||
| 2.94660 | 0.1052 | |||||
| 0.03947 | 0.1565 | |||||
| Foliage | −3.95709 | 0.6063 | ||||
| 2.25431 | 0.1504 | |||||
| −0.08953 | 0.2446 |
| Models | Weighting Functions | R2 | RMSE | ME | MAE | MAPE | L-Value | Pr > |L| | F-Value | Pr > |F| |
|---|---|---|---|---|---|---|---|---|---|---|
| Root | ||||||||||
| PA-1 | D3.5046 | 0.948 | 7.02 | −0.82 | 5.41 | 25.76 | ||||
| DV-1 | D2.2508 | 0.956 | 6.51 | −0.08 | 5.03 | 24.79 | 50.06 | <0.0001 | 11.04 | 0.0015 |
| PA-2 | D2.6015 | 0.952 | 6.81 | −0.45 | 5.40 | 25.38 | ||||
| DV-2 | D3.0141 | 0.958 | 6.44 | −0.05 | 5.16 | 24.55 | 41.56 | <0.0001 | 5.92 | <0.0001 |
| Stem | ||||||||||
| PA-1 | D2.1503 | 0.936 | 19.25 | −1.03 | 13.72 | 17.28 | ||||
| DV-1 | D1.1556 | 0.963 | 14.75 | 0.14 | 11.01 | 15.50 | 165.33 | <0.0001 | 44.65 | <0.0001 |
| PA-2 | D1.4699 | 0.976 | 11.89 | −0.01 | 8.36 | 11.08 | ||||
| DV-2 | D1.4078 | 0.977 | 11.68 | −0.32 | 8.76 | 11.57 | 18.07 | <0.0001 | 2.47 | 0.1209 |
| Branch | ||||||||||
| PA-1 | D1.7870 | 0.884 | 7.37 | 0.28 | 5.22 | 27.69 | ||||
| DV-1 | D1.8359 | 0.972 | 3.65 | −0.21 | 2.86 | 21.90 | 428.88 | <0.0001 | 192.59 | <0.0001 |
| PA-2 | D1.7992 | 0.943 | 5.18 | −0.23 | 4.03 | 28.09 | ||||
| DV-2 | D1.5032 | 0.975 | 3.48 | −0.31 | 2.88 | 23.65 | 245.76 | <0.0001 | 50.50 | <0.0001 |
| Foliage | ||||||||||
| PA-1 | D1.5015 | 0.699 | 4.58 | 0.20 | 3.28 | 39.59 | ||||
| DV-1 | D1.9542 | 0.927 | 2.72 | −0.08 | 1.87 | 28.65 | 426.82 | <0.0001 | 190.86 | <0.0001 |
| PA-2 | D1.6854 | 0.848 | 3.26 | −0.09 | 2.35 | 37.42 | ||||
| DV-2 | D1.6308 | 0.930 | 2.25 | −0.14 | 1.93 | 31.04 | 231.83 | <0.0001 | 46.41 | <0.0001 |
| Total | ||||||||||
| PA-1 | D1.9378 | 0.985 | 16.76 | −1.36 | 12.46 | 11.11 | ||||
| DV-1 | D1.5307 | 0.985 | 16.86 | −0.23 | 12.70 | 11.18 | 17.47 | 0.0007 | 0.82 | 0.5168 |
| PA-2 | D1.5013 | 0.988 | 15.01 | −0.79 | 10.74 | 10.01 | ||||
| DV-2 | D1.3057 | 0.989 | 15.58 | −0.81 | 11.14 | 10.33 | 11.10 | 0.0299 | 0.32 | 0.9239 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Widagdo, F.R.A.; Dong, L.; Li, F. Biomass Functions and Carbon Content Variabilities of Natural and Planted Pinus koraiensis in Northeast China. Plants 2021, 10, 201. https://doi.org/10.3390/plants10020201
Widagdo FRA, Dong L, Li F. Biomass Functions and Carbon Content Variabilities of Natural and Planted Pinus koraiensis in Northeast China. Plants. 2021; 10(2):201. https://doi.org/10.3390/plants10020201
Chicago/Turabian StyleWidagdo, Faris Rafi Almay, Lihu Dong, and Fengri Li. 2021. "Biomass Functions and Carbon Content Variabilities of Natural and Planted Pinus koraiensis in Northeast China" Plants 10, no. 2: 201. https://doi.org/10.3390/plants10020201
APA StyleWidagdo, F. R. A., Dong, L., & Li, F. (2021). Biomass Functions and Carbon Content Variabilities of Natural and Planted Pinus koraiensis in Northeast China. Plants, 10(2), 201. https://doi.org/10.3390/plants10020201