
Morphological, Physiological and Molecular Markers for Salt-Stressed Plants


Abstract
:1. Introduction
2. Detection of Salinity Stress
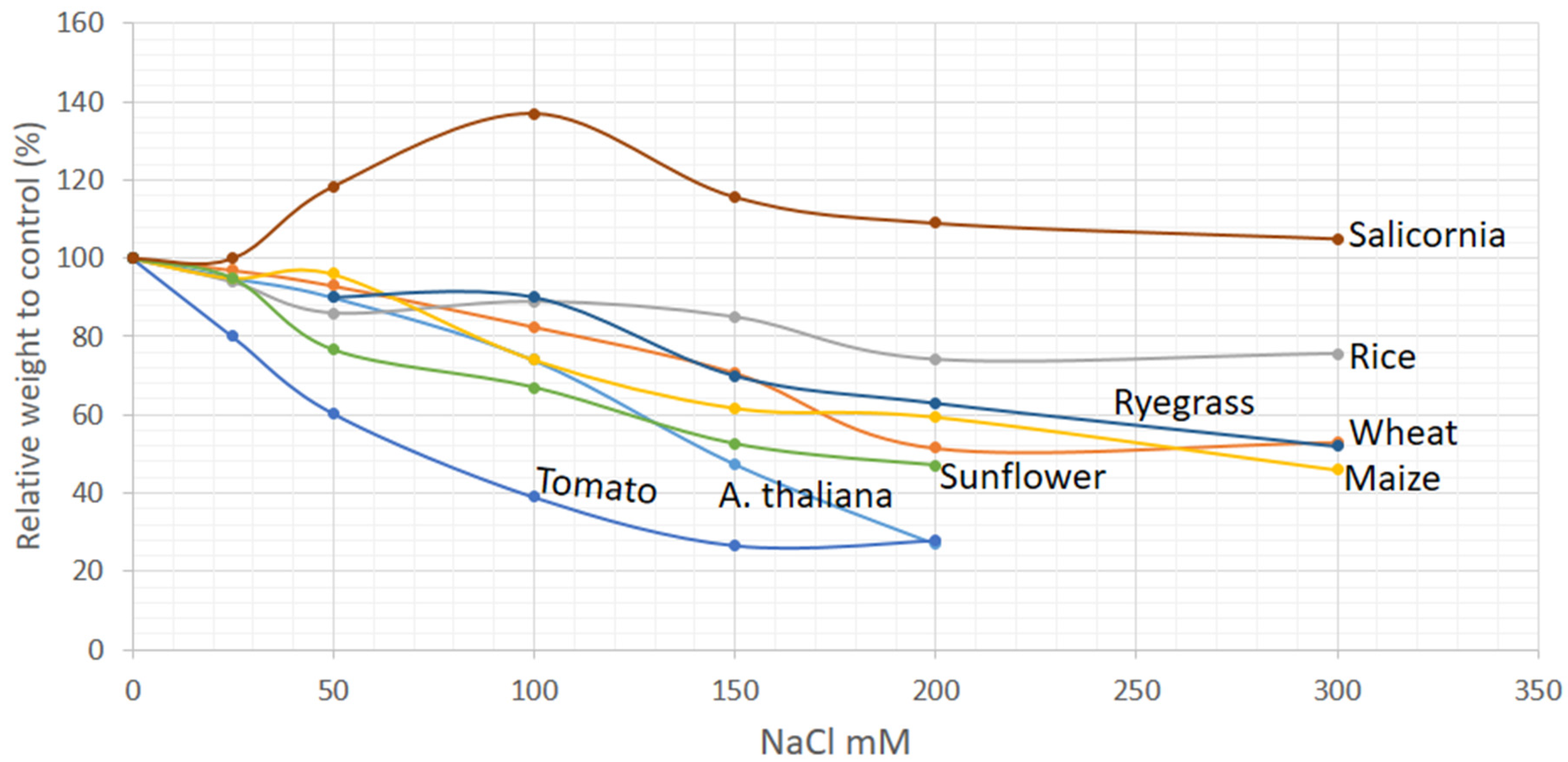
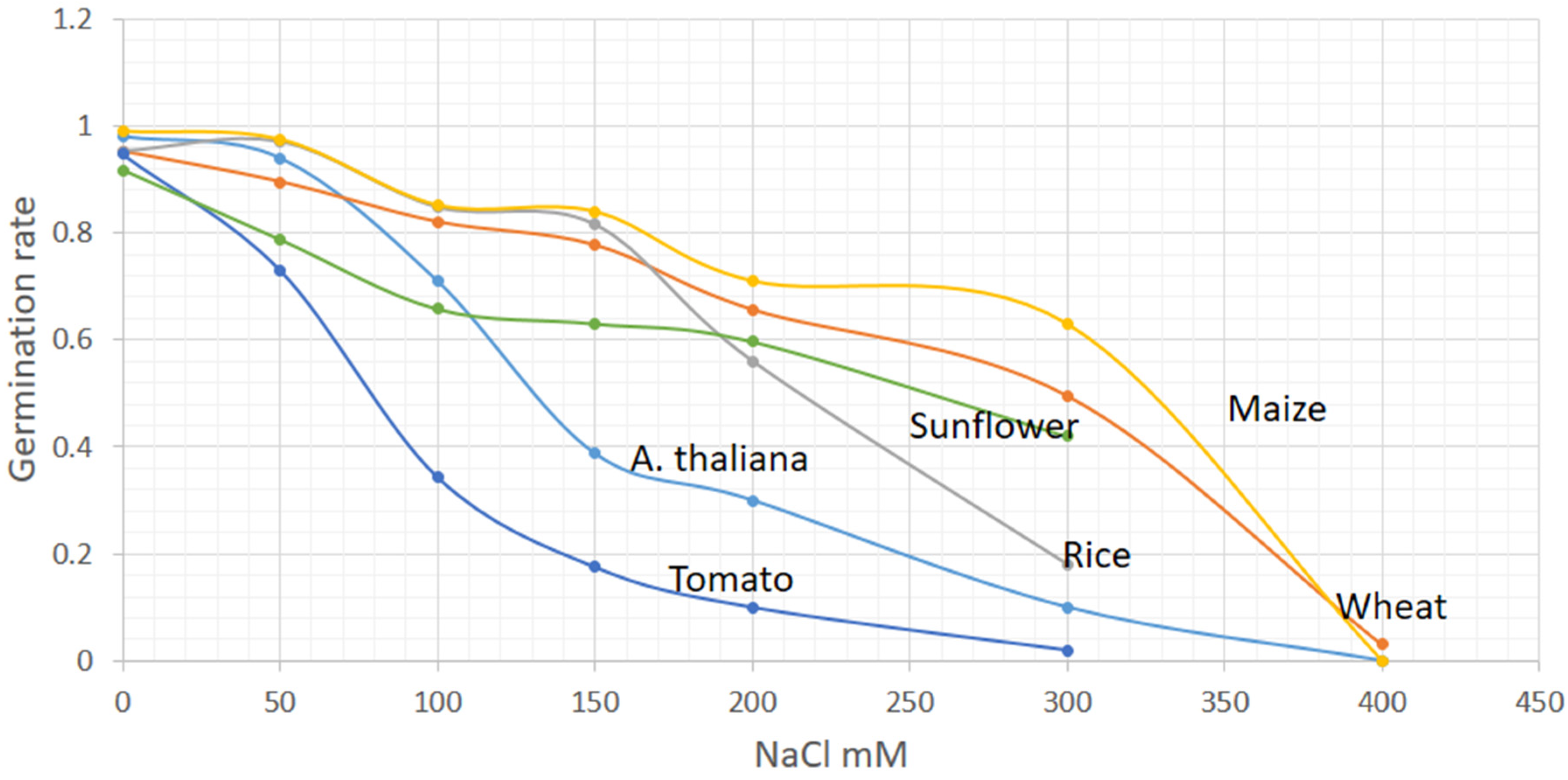
2.1. Morphological Markers
2.2. Physiological Markers
2.3. Oxidative Stress Markers
2.4. Molecular Stress Markers
3. Evaluation of Salinity Stress in Plants by Different Stress Markers
4. Prediction and Identification of Stressed Plants Using Deep Learning Approaches
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flowers, T.J.; Yeo, A.R. Effects of salinity on plant growth and crops yields. In Environmental Stress in Plants; Cherry, J.H., Ed.; NATO ASI Series; Springer: Berlin, Germany, 1989; Volume G19, pp. 101–119. [Google Scholar]
- Ali, Y.; Aslam, Z.; Ashraf, M.Y.; Tahir, G.R. Effect of salinity on chlorophyll concentration, leaf area, yield and yield components of rice genotypes grown under saline environment. Int. J. Environ. Sci. Technol. 2004. [Google Scholar] [CrossRef] [Green Version]
- Yermiyahu, U.; Ben-Gal, A.; Keren, R.; Reid, R.J. Combined effect of salinity and excess boron on plant growth and yield. Plant Soil 2008. [Google Scholar] [CrossRef]
- Ivushkin, K.; Bartholomeus, H.; Bregt, A.K.; Pulatov, A.; Kempen, B.; de Sousa, L. Global mapping of soil salinity change. Remote Sens. Environ. 2019. [Google Scholar] [CrossRef]
- Khatoon, A.; Qureshi, M.S.; Hussain, M.K. Effect of Salinity on Some Yield Parameters of Sunflower (Helianthus annuus L.). Int. J. Agric. Biol. 2000, 2, 382–384. [Google Scholar]
- Abd El-Kader, A.A.; Mohamedin, A.A.M.; Ahmed, M.K.A. Growth and Yield of Sunflower as Affected by Different Salt Affected Soils. Int. J. Agric. Biol. 2006, 8, 583–587. [Google Scholar]
- Chamekh, Z.; Ayadi, S.; Karmous, C.; Trifa, Y.; Amara, H.; Boudabbous, K.; Yousfi, S.; Serret, M.D.; Araus, J.L. Comparative effect of salinity on growth, grain yield, water use efficiency, δ13C and δ15N of landraces and improved durum wheat varieties. Plant Sci. 2016. [Google Scholar] [CrossRef]
- Elkhatib, H.A.; Elkhatib, E.A.; Khalaf Allah, A.M.; El-Sharkawy, A.M. Yield Response of Salt-Stressed Potato to Potassium Fertilization: A Preliminary Mathematical Model. J. Plant Nutr. 2004. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genomics 2014. [Google Scholar] [CrossRef]
- Ondrasek, G.; Rengel, Z.; Veres, S. Soil Salinisation and Salt Stress in Crop Production. In Abiotic Stress in Plants—Mechanisms and Adaptations; Shanker, A.K., Venkateswarlu, B., Eds.; InTech: Rijeka, Croatia, 2011; pp. 171–190. ISBN 978-953-307-394-1. [Google Scholar] [CrossRef] [Green Version]
- Sytar, O.; Brestic, M.; Zivcak, M.; Olsovska, K.; Kovar, M.; Shao, H.; He, X. Applying hyperspectral imaging to explore natural plant diversity towards improving salt stress tolerance. Sci. Total Environ. 2017. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Mbarki, S.; Sytar, O.; Cerda, A.; Zivcak, M.; Rastogi, A.; He, X.; Zoghlami, A.; Abdelly, C.; Brestic, M. Strategies to mitigate the salt stress effects on photosynthetic apparatus and productivity of crop plants. In Salinity Responses and Tolerance in Plants, Volume 1: Targeting Sensory, Transport and Signaling Mechanisms; Springer International Publishing: Cham, Switzerland, 2018; pp. 85–136. ISBN 9783319756714. [Google Scholar]
- Samuel, A.L. Some Studies in Machine Learning. IBM J. Res. Dev. 1959, 3, 210–229. [Google Scholar] [CrossRef]
- Lecun, Y.; Bengio, Y.; Hinton, G. Deep learning. Nature 2015, 521, 436–444. [Google Scholar] [CrossRef]
- Zhang, L.; Tan, J.; Han, D.; Zhu, H. From machine learning to deep learning: Progress in machine intelligence for rational drug discovery. Drug Discov. Today 2017, 22, 1680–1685. [Google Scholar] [CrossRef]
- Kamilaris, A.; Prenafeta-Boldú, F.X. Deep learning in agriculture: A survey. Comput. Electron. Agric. 2018, 147, 70–90. [Google Scholar] [CrossRef] [Green Version]
- Liakos, K.G.; Busato, P.; Moshou, D.; Pearson, S.; Bochtis, D. Machine learning in agriculture: A review. Sensors (Switzerland) 2018, 18, 2674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.; Zhan, Y.; Wang, Q.; Yang, X.; Yu, C.; Wang, H.; Tang, Z.Y.; Jiang, D.; Peng, C.; He, Y. Hyperspectral imaging combined with machine learning as a tool to obtain high-throughput plant salt-stress phenotyping. Plant J. 2020. [Google Scholar] [CrossRef] [PubMed]
- Behmann, J.; Steinrücken, J.; Plümer, L. Detection of early plant stress responses in hyperspectral images. ISPRS J. Photogramm. Remote Sens. 2014. [Google Scholar] [CrossRef]
- Chen, D.; Neumann, K.; Friedel, S.; Kilian, B.; Chen, M.; Altmann, T.; Klukas, C. Dissecting the phenotypic components of crop plant growthand drought responses based on high-throughput image analysis w open. Plant Cell 2014. [Google Scholar] [CrossRef] [Green Version]
- Kersting, K.; Xu, Z.; Wahabzada, M.; Bauckhage, C.; Thurau, C.; Roemer, C.; Ballvora, A.; Rascher, U.; Leon, J.; Pluemer, L. Pre-symptomatic prediction of plant drought stress using Dirichlet-aggregation regression on hyperspectral images. In Proceedings of the Twenty-Sixth AAAI Conference on Artificial Intelligence, Toronto, ON, Canada, 22–26 July 2012; AAAI Publications: Toronto, ON, Canada, 2012; pp. 302–308. [Google Scholar]
- Gong, H.J.; Randall, D.P.; Flowers, T.J. Silicon deposition in the root reduces sodium uptake in rice (Oryza sativa L.) seedlings by reducing bypass flow. Plant Cell Environ. 2006. [Google Scholar] [CrossRef]
- Turan, M.A.; Elkarim, A.H.A.; Taban, N.; Taban, S. Effect of salt stress on growth, stomatal resistance, proline and chlorophyll concentrations on maize plant. Afr. J. Agric. Res. 2009, 4, 893–897. [Google Scholar]
- Li, N.; Wang, X.; Ma, B.; Du, C.; Zheng, L.; Wang, Y. Expression of a Na+/H+ antiporter RtNHX1 from a recretohalophyte Reaumuria trigyna improved salt tolerance of transgenic Arabidopsis thaliana. J. Plant Physiol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Tornero, O.; Tallón, C.I.; Porras, I.; Navarro, J.M. Physiological and growth changes in micropropagated Citrus macrophylla explants due to salinity. J. Plant Physiol. 2009. [Google Scholar] [CrossRef] [PubMed]
- Ghaleb, W.S.; Sawwan, J.S.; Akash, M.W.; Al-Abdallat, A.M. In vitro response of two citrus rootstocks to salt stress. Int. J. Fruit Sci. 2010. [Google Scholar] [CrossRef]
- Abbas, G.; Saqib, M.; Akhtar, J.; Basra, S.M.A. Salinity tolerance potential of two acacia species at early seedling stage. Pak. J. Agric. Sci. 2013, 50, 683–688. [Google Scholar]
- Al-Abdoulhadi, I.A.; Dinar, H.A.; Ebert, G.; Büttner, C. Effect of salinity on leaf growth, leaf injury and biomass production in date palm (Phoenix dactylifera L.) cultivars. Indian J. Sci. Technol. 2011. [Google Scholar] [CrossRef]
- Hemida, K.A.; Ali, R.M.; Ibrahim, W.M.; Sayed, M.A. Ameliorative role of some antioxidant compounds on physiological parameters and antioxidants response of wheat (Triticum aestivum L.) seedlings under salinity stress. Life Sci. J. 2014, 11, 324–342. [Google Scholar]
- Umnajkitikorn, K.; Faiyue, B. Enhancing Antioxidant Properties of Germinated Thai rice (Oryza sativa L.) cv. Kum Doi Saket with Salinity. Rice Res. Open Access 2013. [Google Scholar] [CrossRef]
- Ghars, M.A.; Parre, E.; Debez, A.; Bordenave, M.; Richard, L.; Leport, L.; Bouchereau, A.; Savouré, A.; Abdelly, C. Comparative salt tolerance analysis between Arabidopsis thaliana and Thellungiella halophila, with special emphasis on K+/Na+ selectivity and proline accumulation. J. Plant Physiol. 2008. [Google Scholar] [CrossRef]
- Sultana, N.; Ikeda, T.; Itoh, R. Effect of NaCl salinity on photosynthesis and dry matter accumulation in developing rice grains. Environ. Exp. Bot. 1999. [Google Scholar] [CrossRef]
- Gautam, S.; Singh, P.K. Salicylic acid-induced salinity tolerance in corn grown under NaCl stress. Acta Physiol. Plant. 2009. [Google Scholar] [CrossRef]
- Shiyab, S.M.; Shatnawi, M.A.; Shibli, R.A.; Al Smeirat, N.G.; Ayad, J.; Akash, M.W. Growth, nutrient acquisition, and physiological responses of hydroponic grown tomato to sodium chloride salt induced stress. J. Plant Nutr. 2013. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Yuan, Y.Z.; Ou, J.Q.; Lin, Q.H.; Zhang, C.F. Glutamine synthetase and glutamate dehydrogenase contribute differentially to proline accumulation in leaves of wheat (Triticum aestivum) seedlings exposed to different salinity. J. Plant Physiol. 2007. [Google Scholar] [CrossRef] [PubMed]
- Aghaleh, M.; Niknam, V.; Ebrahimzadeh, H.; Razavi, K. Salt stress effects on growth, pigments, proteins and lipid peroxidation in Salicornia persica and S. europaea. Biol. Plant. 2009. [Google Scholar] [CrossRef]
- Lv, S.; Jiang, P.; Chen, X.; Fan, P.; Wang, X.; Li, Y. Multiple compartmentalization of sodium conferred salt tolerance in Salicornia europaea. Plant Physiol. Biochem. 2012. [Google Scholar] [CrossRef] [PubMed]
- Gul, B.; Ansari, R.; Khan, M.A. Salt tolerance of Salicornia Utahensis from the great basin desert. Pak. J. Bot. 2009, 41, 2925–2932. [Google Scholar]
- Ventura, Y.; Sagi, M. Halophyte crop cultivation: The case for salicornia and sarcocornia. Environ. Exp. Bot. 2013. [Google Scholar] [CrossRef]
- Singh, A.; Jha, S.K.; Bagri, J.; Pandey, G.K. ABA inducible rice protein phosphatase 2C confers ABA insensitivity and abiotic stress tolerance in Arabidopsis. PLoS ONE 2015. [Google Scholar] [CrossRef] [Green Version]
- Renault, H.; Roussel, V.; El Amrani, A.; Arzel, M.; Renault, D.; Bouchereau, A.; Deleu, C. The Arabidopsis pop2-1 mutant reveals the involvement of GABA transaminase in salt stress tolerance. BMC Plant Biol. 2010. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.P.; Jha, P.N. A halotolerant bacterium Bacillus licheniformis HSW-16 augments induced systemic tolerance to salt stress in wheat plant (Triticum aestivum). Front. Plant Sci. 2016. [Google Scholar] [CrossRef]
- El-Hendawy, S.E.; Hu, Y.; Yakout, G.M.; Awad, A.M.; Hafiz, S.E.; Schmidhalter, U. Evaluating salt tolerance of wheat genotypes using multiple parameters. Eur. J. Agron. 2005. [Google Scholar] [CrossRef]
- Azooz, M.M.; Ismail, A.M.; Elhamd, M.F.A. Growth, lipid peroxidation and antioxidant enzyme activities as a selection criterion for the salt tolerance of maize cultivars grown under salinity stress. Int. J. Agric. Biol. 2009, 11, 21–26. [Google Scholar]
- Sultana, R.; Uddin, M.N.; Rahim, M.A.; Fakir, M.S.A. Effect of salt stress on germination and seedling growth in four maize genotypes. Pak. J. Biol. Sci. 2016, 4, 359–360. [Google Scholar] [CrossRef] [Green Version]
- Saboora, A.; Kiarostami, K.; Behroozbayati, F.; Hajihashemi, S. Salinity (NaCl) tolerance of wheat genotypes at germination and early seedling growth. Pak. J. Biol. Sci. 2006. [Google Scholar] [CrossRef]
- Al-aghabary, K.; Zhu, Z.; Shi, Q. Influence of silicon supply on chlorophyll content, chlorophyll fluorescence, and antioxidative enzyme activities in tomato plants under salt stress. J. Plant Nutr. 2004. [Google Scholar] [CrossRef]
- Debouba, M.; Maâroufi-Dghimi, H.; Suzuki, A.; Ghorbel, M.H.; Gouia, H. Changes in growth and activity of enzymes involved in nitrate reduction and ammonium assimilation in tomato seedlings in response to NaCl stress. Ann. Bot. 2007. [Google Scholar] [CrossRef] [Green Version]
- Delgado, I.C.; Sánchez-Raya, A.J. Effects of sodium chloride and mineral nutrients on initial stages of development of sunflower life. Commun. Soil Sci. Plant Anal. 2007. [Google Scholar] [CrossRef]
- Burgos, P.A.; Roldan, M.; Donaire, J.P.; Agui, I. Effect of sodium chloride on growth, ion content, and hydrogen ion extrusion activity of sunflower and jojoba roots. J. Plant Nutr. 1993. [Google Scholar] [CrossRef]
- Shahbaz, M.; Ashraf, M.; Akram, N.A.; Hanif, A.; Hameed, S.; Joham, S.; Rehman, R. Salt-induced modulation in growth, photosynthetic capacity, proline content and ion accumulation in sunflower (Helianthus annuus L.). Acta Physiol. Plant. 2011. [Google Scholar] [CrossRef]
- Tang, J.; Camberato, J.J.; Yu, X.; Luo, N.; Bian, S.; Jiang, Y. Growth response, carbohydrate and ion accumulation of diverse perennial ryegrass accessions to increasing salinity. Sci. Hortic. (Amst.) 2013. [Google Scholar] [CrossRef]
- Chu, T.N.; Tran, B.T.H.; Van Bui, L.; Hoang, M.T.T. Plant growth-promoting rhizobacterium Pseudomonas PS01 induces salt tolerance in Arabidopsis thaliana. BMC Res. Notes 2019. [Google Scholar] [CrossRef] [Green Version]
- Butt, H.I.; Yang, Z.; Gong, Q.; Chen, E.; Wang, X.; Zhao, G.; Ge, X.; Zhang, X.; Li, F. GaMYB85, an R2R3 MYB gene, in transgenic Arabidopsis plays an important role in drought tolerance. BMC Plant Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Galpaz, N.; Reymond, M. Natural variation in Arabidopsis thaliana revealed a genetic network controlling germination under salt stress. PLoS ONE 2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Raza, Q.; Xu, L.; He, X.; Huang, Y.; Yi, J.; Zhang, D.; Shao, H.B.; Ma, H.; Ali, Z. GmWRKY49, a salt-responsive nuclear protein, improved root length and governed better salinity tolerance in transgenic Arabidopsis. Front. Plant Sci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Ferdosizadeh, L.; Ahmad Sadat-Noori, S.; Zare, N.; Saghafi, S. Assessment of Diode Laser Pretreatments on Germination and Yield of Wheat (Triticum aestivum L.) under Salinity Stress. World J. Agric. Res. 2013. [Google Scholar] [CrossRef]
- Begum, F.; Karmoker, J.L.; Fattah, Q.A.; Maniruzzaman, A.F.M. The effect of salinity on germination and its correlation with K+, Na+, Cl− accumulation in germinating seeds of Triticum aestivum L. cv. akbar. Plant Cell Physiol. 1992, 33, 1009–1014. [Google Scholar]
- Djanaguiraman, M.; Senthil, A.; Ramadass, R. Assessment of rice genotypes for salinity tolerance at germination and seedling stage. Assessment 2003, 90, 506–510. [Google Scholar]
- Aliu, S.; Rusinovci, I.; Fetahu, S.; Gashi, B.; Simeonovska, E.; Rozman, L. The effect of salt stress on the germination of maize (Zea mays L.) seeds and photosynthetic pigments. Acta Agric. Slov. 2015. [Google Scholar] [CrossRef]
- Mohammad, M.; Jun, Z.; Guoying, W. Impact of salinity stress on seed germination indices of maize (Zea mays L.) genotypes. Kragujev. J. Sci. 2014. [Google Scholar] [CrossRef]
- Fellner, M.; Sawhney, V.K. Seed germination in a tomato male-sterile mutant is resistant to osmotic, salt and low-temperature stresses. Theor. Appl. Genet. 2001. [Google Scholar] [CrossRef]
- Torres-Schumann, S.; Godoy, J.A.; Pintor-Toro, J.A.; Moreno, F.J.; Rodrigo, R.M.; Garcia-Herdugo, G. NaCl Effects on Tomato Seed Germination, Cell Activity and Ion Allocation. J. Plant Physiol. 1989. [Google Scholar] [CrossRef]
- Bozcuk, S. Effects of kinetin and salinity on germination of tomato, barley and cotton seeds. Ann. Bot. 1981. [Google Scholar] [CrossRef]
- El Midaoui, M.; Benbella, M.; Talouizete, A. Effect of sodium chloride on sunflower (helianthus annuus l.) seed germination. Helia 2002. [Google Scholar] [CrossRef]
- Khan, M.A.; Gul, B. Halophyte seed germination. In Ecophysiology of High Salinity Tolerant Plants; Khan, M.A., Weber, D.J., Eds.; Springer Academic Publishers: Dordrecht, The Netherlands, 2006; pp. 11–31. [Google Scholar] [CrossRef]
- Zörb, C.; Schmitt, S.; Neeb, A.; Karl, S.; Linder, M.; Schubert, S. The biochemical reaction of maize (Zea mays L.) to salt stress is characterized by a mitigation of symptoms and not by a specific adaptation. Plant Sci. 2004. [Google Scholar] [CrossRef]
- Julkowska, M.M.; Koevoets, I.T.; Mol, S.; Hoefsloot, H.; Feron, R.; Tester, M.A.; Keurentjes, J.J.B.; Korte, A.; Haring, M.A.; De Boer, G.J.; et al. Genetic components of root architecture remodeling in response to salt stress. Plant Cell 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, J.Y.; Lee, H.J.; Seo, P.J.; Jung, J.H.; Ahn, J.H.; Park, C.M. The Arabidopsis floral repressor BFT delays flowering by competing with FT for FD binding under high salinity. Mol. Plant 2014. [Google Scholar] [CrossRef] [Green Version]
- Ryu, J.Y.; Park, C.M.; Seo, P.J. The floral repressor BROTHER OF FT AND TFL1 (BFT) modulates flowering initiation under High salinity in Arabidopsis. Mol. Cells 2011. [Google Scholar] [CrossRef] [Green Version]
- Park, H.J.; Kim, W.Y.; Yun, D.J. A role for GIGANTEA: Keeping the balance between flowering and salinity stress tolerance. Plant Signal. Behav. 2013. [Google Scholar] [CrossRef] [Green Version]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. Changes in plant response to NaCl during development of rice (Oryza sativa L.) varieties differing in salinity resistance. J. Exp. Bot. 1995. [Google Scholar] [CrossRef]
- Khatun, S.; Rizzo, C.A.; Flowers, T.J. Genotypic variation in the effect of salinity on fertility in rice. Plant Soil 1995. [Google Scholar] [CrossRef]
- Khorasani, S.; Mostafavi, K.; Heidarian, A. Response of Maize (Zea mays L.) hybrids and inbred lines to salinity stress under field condition. Tech. J. Eng. Appl. Sci. 2011, 2, 1–7. [Google Scholar]
- Castillo, E.G.; Tuong, T.P.; Ismail, A.M.; Inubushi, K. Response to salinity in rice: Comparative effects of osmotic and ionic stresses. Plant Prod. Sci. 2007. [Google Scholar] [CrossRef] [Green Version]
- Romdhane, L.; Dal Cortivo, C.; Vamerali, T.; Radhouane, L. Effects of drought and salinity on maize phenology, morphology and productivity in a semi-arid environment. Ital. J. Agrometeorol. 2016. [Google Scholar] [CrossRef]
- Lefèvre, I.; Gratia, E.; Lutts, S. Discrimination between the ionic and osmotic components of salt stress in relation to free polyamine level in rice (Oryza sativa). Plant Sci. 2001. [Google Scholar] [CrossRef]
- Jedmowski, C.; Ashoub, A.; Brüggemann, W. Reactions of Egyptian landraces of Hordeum vulgare and Sorghum bicolor to drought stress, evaluated by the OJIP fluorescence transient analysis. Acta Physiol. Plant. 2013. [Google Scholar] [CrossRef]
- Turan, M.A.; Katkat, V.; Taban, S. Variations in proline, chlorophyll and mineral elements contents of wheat plants grown under salinity stress. J. Agron. 2007. [Google Scholar] [CrossRef] [Green Version]
- Datta, J.; Nag, S.; Banerjee, A.; Mondai, N. Impact of salt stress on five varieties of Wheat (Triticum aestivum L.) cultivars under laboratory condition. J. Appl. Sci. Environ. Manag. 2010. [Google Scholar] [CrossRef] [Green Version]
- Vaidyanathan, H.; Sivakumar, P.; Chakrabarty, R.; Thomas, G. Scavenging of reactive oxygen species in NaCl-stressed rice (Oryza sativa L.)—Differential response in salt-tolerant and sensitive varieties. Plant Sci. 2003. [Google Scholar] [CrossRef]
- Shibli, R.A.; Kushad, M.; Yousef, G.G.; Lila, M.A. Physiological and biochemical responses of tomato microshoots to induced salinity stress with associated ethylene accumulation. Plant Growth Regul. 2007. [Google Scholar] [CrossRef]
- Choi, J.Y.; Seo, Y.S.; Kim, S.J.; Kim, W.T.; Shin, J.S. Constitutive expression of CaXTH3, a hot pepper xyloglucan endotransglucosylase/hydrolase, enhanced tolerance to salt and drought stresses without phenotypic defects in tomato plants (Solanum lycopersicum cv. Dotaerang). Plant Cell Rep. 2011. [Google Scholar] [CrossRef]
- Poór, P.; Gémes, K.; Horváth, F.; Szepesi, Á.; Simon, M.L.; Tari, I. Salicylic acid treatment via the rooting medium interferes with stomatal response, CO2 fixation rate and carbohydrate metabolism in tomato, and decreases harmful effects of subsequent salt stress. Plant Biol. 2011. [Google Scholar] [CrossRef]
- Navarro, J.M.; Martínez, V.; Carvajal, M. Ammonium, bicarbonate and calcium effects on tomato plants grown under saline conditions. Plant Sci. 2000. [Google Scholar] [CrossRef]
- Ellouzi, H.; Ben Hamed, K.; Cela, J.; Munné-Bosch, S.; Abdelly, C. Early effects of salt stress on the physiological and oxidative status of Cakile maritima (halophyte) and Arabidopsis thaliana (glycophyte). Physiol. Plant. 2011. [Google Scholar] [CrossRef] [PubMed]
- M’rah, S.; Ouerghi, Z.; Eymery, F.; Rey, P.; Hajji, M.; Grignon, C.; Lachaâl, M. Efficiency of biochemical protection against toxic effects of accumulated salt differentiates Thellungiella halophila from Arabidopsis thaliana. J. Plant Physiol. 2007. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Zhao, Y.; Jin, D.; Zhang, L.; Bi, X.; Chen, H.; Xu, Q.; Ma, C.; Li, G. Salicylic acid-altering Arabidopsis mutants response to salt stress. Plant Soil 2012. [Google Scholar] [CrossRef]
- Wang, H.; Liang, X.; Wan, Q.; Wang, X.; Bi, Y. Ethylene and nitric oxide are involved in maintaining ion homeostasis in Arabidopsis callus under salt stress. Planta 2009. [Google Scholar] [CrossRef]
- Shi, H.; Lee, B.H.; Wu, S.J.; Zhu, J.K. Overexpression of a plasma membrane Na+/H+ antiporter gene improves salt tolerance in Arabidopsis thaliana. Nat. Biotechnol. 2003. [Google Scholar] [CrossRef]
- Huang, X.Y.; Chao, D.Y.; Gao, J.P.; Zhu, M.Z.; Shi, M.; Lin, H.X. A previously unknown zinc finger protein, DST, regulates drought and salt tolerance in rice via stomatal aperture control. Genes Dev. 2009. [Google Scholar] [CrossRef] [Green Version]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. NaCl-induced senescence in leaves of rice (Oryza sativa L.) cultivars differing in salinity resistance. Ann. Bot. 1996. [Google Scholar] [CrossRef]
- Yoon, B.S.; Jin, C.; Un, P.S.; Cho, D.H. Change in Photosynthesis, Proline Content, and Osmotic Potential of Corn Seedling under High-Saline Condition. Korean J. Crop Sci. 2005, 50, 28–31. [Google Scholar]
- Yang, X.; Lu, C. Photosynthesis is improved by exogenous glycinebetaine in salt-stressed maize plants. Physiol. Plant. 2005. [Google Scholar] [CrossRef]
- Estrada, B.; Aroca, R.; Barea, J.M.; Ruiz-Lozano, J.M. Native arbuscular mycorrhizal fungi isolated from a saline habitat improved maize antioxidant systems and plant tolerance to salinity. Plant Sci. 2013. [Google Scholar] [CrossRef] [PubMed]
- Katerji, N.; van Hoorn, J.W.; Hamdy, A.; Karam, F.; Mastrorilli, M. Effect of salinity on emergence and on water stress and early seedling growth of sunflower and maize. Agric. Water Manag. 1994. [Google Scholar] [CrossRef]
- Almansouri, M.; Kinet, J.M.; Lutts, S. Effect of salt and osmotic stresses on germination in durum wheat (Triticum durum Desf.). Plant Soil 2001. [Google Scholar] [CrossRef]
- Zheng, Y.H.; Li, X.; Li, Y.G.; Miao, B.H.; Xu, H.; Simmons, M.; Yang, X.H. Contrasting responses of salinity-stressed salt-tolerant and intolerant winter wheat (Triticum aestivum L.) cultivars to ozone pollution. Plant Physiol. Biochem. 2012. [Google Scholar] [CrossRef]
- Kafi, M. The effects of salinity and light on photosynthesis, respiration and chlorophyll fluorescence in salt-tolerant and salt-sensitive wheat (Triticum aestivum L.) cultivars. J. Agric. Sci. Technol. 2009, 11, 535–547. [Google Scholar]
- Brini, F.; Amara, I.; Feki, K.; Hanin, M.; Khoudi, H.; Masmoudi, K. Physiological and molecular analyses of seedlings of two Tunisian durum wheat (Triticum turgidum L. subsp. Durum [Desf.]) varieties showing contrasting tolerance to salt stress. Acta Physiol. Plant. 2009. [Google Scholar] [CrossRef]
- Khan, N.A. NaCl-inhibited chlorophyll synthesis and associated changes in ethylene evolution and antioxidative enzyme activities in wheat. Biol. Plant. 2003. [Google Scholar] [CrossRef]
- Heidari, A.; Toorchi, M.; Bandehagh, A.; Shakiba, M. Effect of NaCl Stress on Growth, Water Relations, Organic and Inorganic Osmolytes Accumulation in Sunflower (Helianthus annuus L.) Lines. Univers. J. Environ. Res. Technol. 2011, 1, 351–362. [Google Scholar]
- Liu, J.; Shi, D.C. Photosynthesis, chlorophyll fluorescence, inorganic ion and organic acid accumulations of sunflower in responses to salt and salt-alkaline mixed stress. Photosynthetica 2010. [Google Scholar] [CrossRef]
- Barros, C.V.S.D.; Melo, Y.L.; Souza, M. de F.; Silva, D.V.; de Macedo, C.E.C. Sensitivity and biochemical mechanisms of sunflower genotypes exposed to saline and water stress. Acta Physiol. Plant. 2019. [Google Scholar] [CrossRef]
- Rivelli, A.R.; de Maria, S.; Pizza, S.; Gherbin, P. Growth and physiological response of hydroponically-grown sunflower as affected by salinity and magnesium levels. J. Plant Nutr. 2010. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, J.; Huang, B. Cytokinin-mitigation of salt-induced leaf senescence in perennial ryegrass involving the activation of antioxidant systems and ionic balance. Environ. Exp. Bot. 2016. [Google Scholar] [CrossRef]
- Ji, Z.; Camberato, J.J.; Zhang, C.; Jiang, Y. Effects of 6-benzyladenine, g-aminobutyric acid, and nitric oxide on plant growth, photochemical efficiency, and ion accumulation of perennial ryegrass cultivars to salinity stress. HortScience 2019. [Google Scholar] [CrossRef] [Green Version]
- Katschnig, D.; Broekman, R.; Rozema, J. Salt tolerance in the halophyte salicornia dolichostachya moss: Growth, morphology and physiology. Environ. Exp. Bot. 2013. [Google Scholar] [CrossRef]
- Park, K.W.; An, J.Y.; Lee, H.J.; Son, D.; Sohn, Y.G.; Kim, C.G.; Lee, J.J. The growth and accumulation of osmotic solutes of the halophyte common glasswort (Salicornia europaea) under salinity conditions. J. Aquat. Plant Manag. 2013, 51, 103–108. [Google Scholar]
- Santhanakrishnan, D.; Perumal, R.K.; Kanth, S.V.; Jonnalagadda, R.R.; Bangaru, C. Studies on the physiological and biochemical characteristics of Salicornia brachiata: Influence of saline stress due to soaking wastewater of tannery. Desalin. Water Treat. 2014. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Gémes, K.; Kim, Y.J.; Park, K.Y.; Moschou, P.N.; Andronis, E.; Valassaki, C.; Roussis, A.; Roubelakis-Angelakis, K.A. An NADPH-oxidase/polyamine oxidase feedback loop controls oxidative burst under salinity. Plant Physiol. 2016. [Google Scholar] [CrossRef] [Green Version]
- Li, Y. Physiological Responses of Tomato Seedlings (Lycopersicon Esculentum) to Salt Stress. Mod. Appl. Sci. 2009. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.E.; Mao, J.J.; Sun, L.Q.; Huang, B.; Ding, C.B.; Gu, Y.; Liao, J.Q.; Hu, C.; Zhang, Z.W.; Yuan, S.; et al. Exogenous melatonin enhances salt stress tolerance in maize seedlings by improving antioxidant and photosynthetic capacity. Physiol. Plant. 2018. [Google Scholar] [CrossRef] [PubMed]
- Esfandiari, E.; Gohari, G. Response of ROS-Scavenging systems to salinity stress in two different wheat (Triticum aestivum L.) cultivars. Not. Bot. Horti Agrobot. Cluj-Napoca 2017. [Google Scholar] [CrossRef] [Green Version]
- Jain, P.; Bhatla, S.C. Signaling role of phospholipid hydroperoxide glutathione peroxidase (PHGPX) accompanying sensing of NaCl stress in etiolated sunflower seedling cotyledons. Plant Signal. Behav. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Mun, B.G.; Khan, A.L.; Waqas, M.; Kim, H.H.; Shahzad, R.; Imran, M.; Yun, B.W.; Lee, I.J. Regulation of reactive oxygen and nitrogen species by salicylic acid in rice plants under salinity stress conditions. PLoS ONE 2018. [Google Scholar] [CrossRef] [Green Version]
- Hu, T.; Li, H.Y.; Zhang, X.Z.; Luo, H.J.; Fu, J.M. Toxic effect of NaCl on ion metabolism, antioxidative enzymes and gene expression of perennial ryegrass. Ecotoxicol. Environ. Saf. 2011. [Google Scholar] [CrossRef]
- Parida, A.K.; Jha, B. Antioxidative defense potential to salinity in the euhalophyte Salicornia brachiata. J. Plant Growth Regul. 2010. [Google Scholar] [CrossRef]
- Guan, Q.; Takano, T.; Liu, S. Genetic transformation and analysis of rice OsAPx2 gene in Medicago sativa. PLoS ONE 2012. [Google Scholar] [CrossRef]
- Poór, P.; Kovács, J.; Borbély, P.; Takács, Z.; Szepesi, Á.; Tari, I. Salt stress-induced production of reactive oxygen- and nitrogen species and cell death in the ethylene receptor mutant Never ripe and wild type tomato roots. Plant Physiol. Biochem. 2015. [Google Scholar] [CrossRef] [Green Version]
- Poór, P.; Kovács, J.; Szopkó, D.; Tari, I. Ethylene signaling in salt stress- and salicylic acid-induced programmed cell death in tomato suspension cells. Protoplasma 2013. [Google Scholar] [CrossRef] [Green Version]
- Kataria, S.; Baghel, L.; Guruprasad, K.N. Pre-treatment of seeds with static magnetic field improves germination and early growth characteristics under salt stress in maize and soybean. Biocatal. Agric. Biotechnol. 2017. [Google Scholar] [CrossRef]
- Hanqing, F.; Yifeng, W.; Hongyu, L.; Rongfang, W.; Kun, S.; Lingyun, J. Salt stress-induced expression of rice AOX1a is mediated through an accumulation of hydrogen peroxide. Biologia (Bratisl) 2010. [Google Scholar] [CrossRef]
- Li, J.; Jia, H.; Wang, J.; Cao, Q.; Wen, Z. Hydrogen sulfide is involved in maintaining ion homeostasis via regulating plasma membrane Na+/H+ antiporter system in the hydrogen peroxide-dependent manner in salt-stress Arabidopsis thaliana root. Protoplasma 2014. [Google Scholar] [CrossRef]
- Liu, S.G.; Zhu, D.Z.; Chen, G.H.; Gao, X.Q.; Zhang, X.S. Disrupted actin dynamics trigger an increment in the reactive oxygen species levels in the Arabidopsis root under salt stress. Plant Cell Rep. 2012. [Google Scholar] [CrossRef]
- Hu, L.; Hu, T.; Zhang, X.; Pang, H.; Fu, J. Exogenous glycine betaine Ameliorates the adverse effect of salt stress on perennial ryegrass. J. Am. Soc. Hortic. Sci. 2012. [Google Scholar] [CrossRef] [Green Version]
- Hu, T.; Chen, K.; Hu, L.; Amombo, E.; Fu, J. H2O2 and Ca2+-based signaling and associated ion accumulation, antioxidant systems and secondary metabolism orchestrate the response to NaCl stress in perennial ryegrass. Sci. Rep. 2016. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Z.B.; Guo, J.L.; Zhu, A.J.; Zhang, L.; Zhang, M.M. Exogenous jasmonic acid can enhance tolerance of wheat seedlings to salt stress. Ecotoxicol. Environ. Saf. 2014. [Google Scholar] [CrossRef]
- Sairam, R.K.; Srivastava, G.C.; Agarwal, S.; Meena, R.C. Differences in antioxidant activity in response to salinity stress in tolerant and susceptible wheat genotypes. Biol. Plant. 2005. [Google Scholar] [CrossRef]
- Yamane, K.; Mitsuya, S.; Kawasaki, M.; Taniguchi, M.; Miyake, H. Antioxidant capacity and damages caused by salinity stress in apical and basal regions of rice leaf. Plant Prod. Sci. 2009. [Google Scholar] [CrossRef] [Green Version]
- AbdElgawad, H.; Zinta, G.; Hegab, M.M.; Pandey, R.; Asard, H.; Abuelsoud, W. High salinity induces different oxidative stress and antioxidant responses in maize seedlings organs. Front. Plant Sci. 2016. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Yu, H.; Ouyang, B.; Shi, C.; Demidchik, V.; Hao, Z.; Yu, M.; Shabala, S. NADPH oxidases and the evolution of plant salinity tolerance. Plant Cell Environ. 2020. [Google Scholar] [CrossRef]
- Qi, Y.C.; Liu, W.Q.; Qiu, L.Y.; Zhang, S.M.; Ma, L.; Zhang, H. Overexpression of glutathione S-transferase gene increases salt tolerance of Arabidopsis. Russ. J. Plant Physiol. 2010. [Google Scholar] [CrossRef]
- Li, G.; Peng, X.; Wei, L.; Kang, G. Salicylic acid increases the contents of glutathione and ascorbate and temporally regulates the related gene expression in salt-stressed wheat seedlings. Gene 2013. [Google Scholar] [CrossRef]
- Badran, E.G.; Abogadallah, G.M.; Nada, R.M.; Nemat Alla, M.M. Role of glycine in improving the ionic and ROS homeostasis during NaCl stress in wheat. Protoplasma 2015. [Google Scholar] [CrossRef]
- Kaur, H.; Bhatla, S.C. Melatonin and nitric oxide modulate glutathione content and glutathione reductase activity in sunflower seedling cotyledons accompanying salt stress. Nitric Oxide Biol. Chem. 2016. [Google Scholar] [CrossRef]
- Mansour, M.M.F. Plasma membrane permeability as an indicator of salt tolerance in plants. Biol. Plant. 2013, 57, 1–10. [Google Scholar] [CrossRef]
- Singam, K.; Juntawong, N.; Cha-Um, S.; Kirdmanee, C. Salt stress induced ion accumulation, ion homeostasis, membrane injury and sugar contents in salt-sensitive rice (Oryza sativa L. spp. indica) roots under isoosmotic conditions. Afr. J. Biotechnol. 2011. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Q.; Wu, J.; Zheng, X.; Zheng, S.; Sun, X.; Qiu, Q.; Lu, T. Gene Knockout Study Reveals That Cytosolic Ascorbate Peroxidase 2(OsAPX2) Plays a Critical Role in Growth and Reproduction in Rice under Drought, Salt and Cold Stresses. PLoS ONE 2013. [Google Scholar] [CrossRef]
- Yarmolinsky, D.; Brychkova, G.; Kurmanbayeva, A.; Bekturova, A.; Ventura, Y.; Khozin-Goldberg, I.; Eppel, A.; Fluhr, R.; Sagi, M. Impairment in sulfite reductase leads to early leaf senescence in tomato plants. Plant Physiol. 2014. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, S.; Brychkova, G.; Yarmolinsky, D.; Soltabayeva, A.; Samani, T.; Sagi, M. Aldehyde oxidase 4 plays a critical role in delaying silique senescence by catalyzing aldehyde detoxification. Plant Physiol. 2017. [Google Scholar] [CrossRef] [Green Version]
- Gapińska, M.; Skłodowska, M.; Gabara, B. Effect of short- and long-term salinity on the activities of antioxidative enzymes and lipid peroxidation in tomato roots. Acta Physiol. Plant. 2008. [Google Scholar] [CrossRef]
- Gong, B.; Wen, D.; VandenLangenberg, K.; Wei, M.; Yang, F.; Shi, Q.; Wang, X. Comparative effects of NaCl and NaHCO3 stress on photosynthetic parameters, nutrient metabolism, and the antioxidant system in tomato leaves. Sci. Hortic. (Amst.) 2013. [Google Scholar] [CrossRef]
- Torabi, S.; Niknam, V. Effects of Iso-osmotic Concentrations of NaCl and Mannitol on some Metabolic Activity in Calluses of Two Salicornia species. Vitr. Cell. Dev. Biol. Plant 2011. [Google Scholar] [CrossRef]
- Wang, Y.; Ying, Y.; Chen, J.; Wang, X. Transgenic Arabidopsis overexpressing Mn-SOD enhanced salt-tolerance. Plant Sci. 2004. [Google Scholar] [CrossRef]
- El-Hendawy, S.; Al-Suhaibani, N.; Hassan, W.; Tahir, M.; Schmidhalter, U. Hyperspectral reflectance sensing to assess the growth and photosynthetic properties of wheat cultivars exposed to different irrigation rates in an irrigated arid region. PLoS ONE 2017. [Google Scholar] [CrossRef]
- Awlia, M.; Alshareef, N.; Saber, N.; Korte, A.; Oakey, H.; Panzarová, K.; Trtílek, M.; Negrão, S.; Tester, M.; Julkowska, M.M. Genetic mapping of the early responses to salt stress in Arabidopsis thaliana. bioRxiv Plant Biol. 2020. [Google Scholar] [CrossRef]
- Gong, Q.; Li, P.; Ma, S.; Indu Rupassara, S.; Bohnert, H.J. Salinity stress adaptation competence in the extremophile Thellungiella halophila in comparison with its relative Arabidopsis thaliana. Plant J. 2005. [Google Scholar] [CrossRef]
- Goyal, E.; Amit, S.K.; Singh, R.S.; Mahato, A.K.; Chand, S.; Kanika, K. Transcriptome profiling of the salt-stress response in Triticum aestivum cv. Kharchia Local. Sci. Rep. 2016. [Google Scholar] [CrossRef] [Green Version]
- Liu, J. The Arabidopsis thaliana SOS2 gene encodes a protein kinase that is required for salt tolerance. Proc. Natl. Acad. Sci. USA 2000. [Google Scholar] [CrossRef]
- Shabala, S.; Wu, H.; Bose, J. Salt stress sensing and early signalling events in plant roots: Current knowledge and hypothesis. Plant Sci. 2015, 241, 109–119. [Google Scholar] [CrossRef]
- Sagor, G.H.M.; Zhang, S.; Kojima, S.; Simm, S.; Berberich, T.; Kusano, T. Reducing cytoplasmic polyamine oxidase activity in Arabidopsis increases salt and drought tolerance by reducing reactive oxygen species production and increasing defense gene expression. Front. Plant Sci. 2016. [Google Scholar] [CrossRef] [Green Version]
- Formentin, E.; Sudiro, C.; Perin, G.; Riccadonna, S.; Barizza, E.; Baldoni, E.; Lavezzo, E.; Stevanato, P.; Sacchi, G.A.; Fontana, P.; et al. Transcriptome and cell physiological analyses in different rice cultivars provide new insights into adaptive and salinity stress responses. Front. Plant Sci. 2018. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Mao, B.; Ou, S.; Wang, W.; Liu, L.; Wu, Y.; Chu, C.; Wang, X. OsbZIP71, a bZIP transcription factor, confers salinity and drought tolerance in rice. Plant Mol. Biol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Liu, Y.; Zhou, G.; Li, Q.; Yang, C.; Peng, S. Overexpression of CsCLCc, a Chloride Channel Gene from Poncirus trifoliata, Enhances Salt Tolerance in Arabidopsis. Plant Mol. Biol. Rep. 2013. [Google Scholar] [CrossRef]
- Seki, M.; Narusaka, M.; Ishida, J.; Nanjo, T.; Fujita, M.; Oono, Y.; Kamiya, A.; Nakajima, M.; Enju, A.; Sakurai, T.; et al. Monitoring the expression profiles of 7000 Arabidopsis genes under drought, cold and high-salinity stresses using a full-length cDNA microarray. Plant J. 2002. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Guo, H.; Xie, Y.; Zhao, L.; Gu, J.; Zhao, S.; Li, J.; Liu, L. RNAseq analysis reveals pathways and candidate genes associated with salinity tolerance in a spaceflight-induced wheat mutant. Sci. Rep. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Hu, T.; Fu, J. Identification of genes associated with adaptation to NaCl toxicity in perennial ryegrass (Lolium perenne L.). Ecotoxicol. Environ. Saf. 2012. [Google Scholar] [CrossRef]
- Jiang, Y.; Deyholos, M.K. Comprehensive transcriptional profiling of NaCl-stressed Arabidopsis roots reveals novel classes of responsive genes. BMC Plant Biol. 2006. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Yang, B.; Harris, N.S.; Deyholos, M.K. Comparative proteomic analysis of NaCl stress-responsive proteins in Arabidopsis roots. J. Exp. Bot. 2007. [Google Scholar] [CrossRef] [Green Version]
- Chookhampaeng, S.; Pattanagul, W.; Theerakulpisut, P. Effects of salinity on growth, activity of antioxidant enzymes and sucrose content in tomato (Lycopersicon esculentum Mill.) at the reproductive stage. Sci. Asia 2008. [Google Scholar] [CrossRef]
- Hu, L.; Li, H.; Pang, H.; Fu, J. Responses of antioxidant gene, protein and enzymes to salinity stress in two genotypes of perennial ryegrass (Lolium perenne) differing in salt tolerance. J. Plant Physiol. 2012. [Google Scholar] [CrossRef]
- Zarza, X.; Atanasov, K.E.; Marco, F.; Arbona, V.; Carrasco, P.; Kopka, J.; Fotopoulos, V.; Munnik, T.; Gómez-Cadenas, A.; Tiburcio, A.F.; et al. Polyamine oxidase 5 loss-of-function mutations in Arabidopsis thaliana trigger metabolic and transcriptional reprogramming and promote salt stress tolerance. Plant Cell Environ. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nounjan, N.; Nghia, P.T.; Theerakulpisut, P. Exogenous proline and trehalose promote recovery of rice seedlings from salt-stress and differentially modulate antioxidant enzymes and expression of related genes. J. Plant Physiol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Rivero, R.M.; Mestre, T.C.; Mittler, R.; Rubio, F.; Garcia-Sanchez, F.; Martinez, V. The combined effect of salinity and heat reveals a specific physiological, biochemical and molecular response in tomato plants. Plant Cell Environ. 2014. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhou, E.; Gao, L.; Mao, X.; Zhou, R.; Jia, J. Isolation, expression analysis and chromosomal location of P5CR gene in common wheat (Triticum aestivum L.). S. Afr. J. Bot. 2008. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Guo, H.; Zhang, X.; Fu, J. Expression profiles of Pr5CS1 and Pr5CS2 genes and proline accumulation under salinity stress in perennial ryegrass (Lolium perenne L.). Plant Breed. 2014. [Google Scholar] [CrossRef]
- Cheng, L.; Li, X.; Huang, X.; Ma, T.; Liang, Y.; Ma, X.; Peng, X.; Jia, J.; Chen, S.; Chen, Y.; et al. Overexpression of sheepgrass R1-MYB transcription factor LcMYB1 confers salt tolerance in transgenic Arabidopsis. Plant Physiol. Biochem. 2013. [Google Scholar] [CrossRef]
- Thomas, S.; Krishna, G.K.; Yadav, P.; Pal, M. Cloning and abiotic stress responsive expression analysis of arginine decarboxylase genes in contrasting rice genotypes. Indian J. Genet. Plant Breed. 2019. [Google Scholar] [CrossRef]
- Fan, P.; Nie, L.; Jiang, P.; Feng, J.; Lv, S.; Chen, X.; Bao, H.; Guo, J.; Tai, F.; Wang, J.; et al. Transcriptome analysis of salicornia europaea under saline conditions revealed the adaptive primary metabolic pathways as early events to facilitate salt adaptation. PLoS ONE 2013. [Google Scholar] [CrossRef]
- Balazadeh, S.; Siddiqui, H.; Allu, A.D.; Matallana-Ramirez, L.P.; Caldana, C.; Mehrnia, M.; Zanor, M.I.; Köhler, B.; Mueller-Roeber, B. A gene regulatory network controlled by the NAC transcription factor ANAC092/AtNAC2/ORE1 during salt-promoted senescence. Plant J. 2010. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Lv, B.; Li, J.; Luo, L.; Lu, S.; Zhang, X.; Ma, H.; Ming, F. The NAC family transcription factor OsNAP confers abiotic stress response through the ABA pathway. Plant Cell Physiol. 2014. [Google Scholar] [CrossRef] [Green Version]
- Balazadeh, S.; Wu, A.; Mueller-Roeber, B. Salt-triggered expression of the ANAC092-dependent senescence regulon in Arabidopsis thaliana. Plant Signal. Behav. 2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allu, A.D.; Soja, A.M.; Wu, A.; Szymanski, J.; Balazadeh, S. Salt stress and senescence: Identification of cross-talk regulatory components. J. Exp. Bot. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Zhang, M.; Guo, R.; Shi, D.; Liu, B.; Lin, X.; Yang, C. Effects of salt stress on ion balance and nitrogen metabolism of old and young leaves in rice (Oryza sativa L.). BMC Plant Biol. 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey-Serres, J.; Mittler, R. The roles of reactive oxygen species in plant cells. Plant Physiol. 2006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vestergaard, C.L.; Flyvbjerg, H.; Møller, I.M. Intracellular signaling by diffusion: Can waves of hydrogen peroxide transmit intracellular information in plant cells? Front. Plant Sci. 2012. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Villasante, C.; Burén, S.; Blázquez-Castro, A.; Barón-Sola, Á.; Hernández, L.E. Fluorescent in vivo imaging of reactive oxygen species and redox potential in plants. Free Radic. Biol. Med. 2018, 122, 202–220. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010. [Google Scholar] [CrossRef]
- Jiang, K.; Schwarzer, C.; Lally, E.; Zhang, S.; Ruzin, S.; Machen, T.; Remington, S.J.; Feldman, L. Expression and characterization of a redox-sensing green fluorescent protein (reduction-oxidation-sensitive green fluorescent protein) in Arabidopsis. Plant Physiol. 2006. [Google Scholar] [CrossRef] [Green Version]
- Meyer, A.J.; Brach, T.; Marty, L.; Kreye, S.; Rouhier, N.; Jacquot, J.P.; Hell, R. Redox-sensitive GFP in Arabidopsis thaliana is a quantitative biosensor for the redox potential of the cellular glutathione redox buffer. Plant J. 2007. [Google Scholar] [CrossRef]
- Morgan, M.J.; Lehmann, M.; Schwarzländer, M.; Baxter, C.J.; Sienkiewicz-Porzucek, A.; Williams, T.C.R.; Schauer, N.; Fernie, A.R.; Fricker, M.D.; Ratcliffe, R.G.; et al. Decrease in manganese superoxide dismutase leads to reduced root growth and affects tricarboxylic acid cycle flux and mitochondrial redox homeostasis. Plant Physiol. 2008. [Google Scholar] [CrossRef] [Green Version]
- Shao, N.; Krieger-Liszkay, A.; Schroda, M.; Beck, C.F. A reporter system for the individual detection of hydrogen peroxide and singlet oxygen: Its use for the assay of reactive oxygen species produced in vivo. Plant J. 2007. [Google Scholar] [CrossRef] [PubMed]
- Puniran-Hartley, N.; Hartley, J.; Shabala, L.; Shabala, S. Salinity-induced accumulation of organic osmolytes in barley and wheat leaves correlates with increased oxidative stress tolerance: In planta evidence for cross-tolerance. Plant Physiol. Biochem. 2014. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Zhang, P.; Zhu, R.; Fu, J.; Su, J.; Zheng, J.; Wang, Z.; Wang, D.; Gong, Q. Autophagy is rapidly induced by salt stress and is required for salt tolerance in Arabidopsis. Front. Plant Sci. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, W.; Bilsborrow, P.E.; Hooley, P.; Fincham, D.A.; Lombi, E.; Forster, B.P. Salinity induced differences in growth, ion distribution and partitioning in barley between the cultivar Maythorpe and its derived mutant Golden Promise. Plant Soil 2003. [Google Scholar] [CrossRef]
- Hu, Y.; Burucs, Z.; Schmidhalter, U. Effect of foliar fertilization application on the growth and mineral nutrient content of maize seedlings under drought and salinity. Soil Sci. Plant Nutr. 2008. [Google Scholar] [CrossRef] [Green Version]
- Ebrahimi, R.; Bhatla, S.C. Effect of sodium chloride levels on growth, water status, uptake, transport, and accumulation pattern of sodium and chloride ions in young sunflower plants. Commun. Soil Sci. Plant Anal. 2011. [Google Scholar] [CrossRef]
- Vieira Santos, C.L.; Campos, A.; Azevedo, H.; Caldeira, G. In situ and in vitro senescence induced by KCI stress: Nutritional imbalance, lipid peroxidation and antioxidant metabolism. J. Exp. Bot. 2001. [Google Scholar] [CrossRef]
- Hajlaoui, H.; Ayeb, N.E.; Garrec, J.P.; Denden, M. Differential effects of salt stress on osmotic adjustment and solutes allocation on the basis of root and leaf tissue senescence of two silage maize (Zea mays L.) varieties. Ind. Crops Prod. 2010. [Google Scholar] [CrossRef]
- Cuin, T.A.; Parsons, D.; Shabala, S. Wheat cultivars can be screened for NaCl salinity tolerance by measuring leaf chlorophyll content and shoot sap potassium. Funct. Plant Biol. 2010. [Google Scholar] [CrossRef]
- Wimmer, M.A.; Mühling, K.H.; Läuchli, A.; Brown, P.H.; Goldbach, H.E. The interaction between salinity and boron toxicity affects the subcellular distribution of ions and proteins in wheat leaves. Plant Cell Environ. 2003. [Google Scholar] [CrossRef]
- Singh, A.K.; Ganapathysubramanian, B.; Sarkar, S.; Singh, A. Deep Learning for Plant Stress Phenotyping: Trends and Future Perspectives. Trends Plant Sci. 2018, 23, 883–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douarre, C.; Schielein, R.; Frindel, C.; Gerth, S.; Rousseau, D. Deep learning based root-soil segmentation from X-ray tomography. bioRxiv 2016. [Google Scholar] [CrossRef]
- Akar, Ö.; Güngör, O. Classification of multispectral images using Random Forest algorithm. J. Geod. Geoinf. 2012. [Google Scholar] [CrossRef] [Green Version]
- Ho Tong Minh, D.; Ienco, D.; Gaetano, R.; Lalande, N.; Ndikumana, E.; Osman, F.; Maurel, P. Deep Recurrent Neural Networks for Winter Vegetation Quality Mapping via Multitemporal SAR Sentinel-1. IEEE Geosci. Remote Sens. Lett. 2018. [Google Scholar] [CrossRef]
- Feng, X.; Yu, C.; Chen, Y.; Peng, J.; Ye, L.; Shen, T.; Wen, H.; He, Y. Non-destructive determination of shikimic acid concentration in transgenic maize exhibiting glyphosate tolerance using chlorophyll fluorescence and hyperspectral imaging. Front. Plant Sci. 2018. [Google Scholar] [CrossRef]
- Biju, S.; Fuentes, S.; Gupta, D. The use of infrared thermal imaging as a non-destructive screening tool for identifying drought-tolerant lentil genotypes. Plant Physiol. Biochem. 2018. [Google Scholar] [CrossRef]
- Chen, D.; Shi, R.; Pape, J.M.; Neumann, K.; Arend, D.; Graner, A.; Chen, M.; Klukas, C. Predicting plant biomass accumulation from image-derived parameters. Gigascience 2018. [Google Scholar] [CrossRef] [Green Version]
- Alhnaity, B.; Pearson, S.; Leontidis, G.; Kollias, S. Using deep learning to predict plant growth and yield in greenhouse environments. arXiv 2019, arXiv:1907.00624. [Google Scholar]
- Altangerel, N.; Ariunbold, G.O.; Gorman, C.; Alkahtani, M.H.; Borrego, E.J.; Bohlmeyer, D.; Hemmer, P.; Kolomiets, M.V.; Yuan, J.S.; Scully, M.O. In vivo diagnostics of early abiotic plant stress response via Raman spectroscopy. Proc. Natl. Acad. Sci. USA 2017. [Google Scholar] [CrossRef] [Green Version]
- Gao, G.; Tester, M.A.; Julkowska, M.M. The Use of High-Throughput Phenotyping for Assessment of Heat Stress-Induced Changes in Arabidopsis. Plant Phenomics 2020. [Google Scholar] [CrossRef]
- Chiu, W.L.; Niwa, Y.; Zeng, W.; Hirano, T.; Kobayashi, H.; Sheen, J. Engineered GFP as a vital reporter in plants. Curr. Biol. 1996. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Physiological Parameters | Salt Sensitive Plants | Less Salt Sensitive Plants | Halophyte | |||||
|---|---|---|---|---|---|---|---|---|
| Tomato | A.Thaliana | Rice | Maize | Wheat | Sunflower | Ryegrass | Salicornia | |
| Na content increase | 50–100 mM; 1 d | 100 mM; 4 h–1 d | 100 mM; 6 h–7 d | 100 mM; 21 d | 100 mM; 3 d | 100–150 mM; 1 d | 100–250 mM; 4 d | 100–200 mM; 10–42 d |
| K content decrease | 50–100 mM; 1 d | 100 mM; 4 h | 100 mM; 7 d | - | 75–100 mM; 7–42 d | 100 mM; 1 d | 100–250 mM; 4 d | 100–200 mM; 10–42 d |
| RWC * decrease | 100 mM; 2 h | 100–200 mM; 1–15 d | 100 mM; 4–12 h | 50 mM; 7 d | 75–100 mM; 3–42 d | 50–100 mM; 2–54 d | 200–250 mM; 12–14 d | 100–200 mM; 10–42 d |
| Chlorophyll decrease | 75–100 mM; 3 d | 100–200 mM; 4–18 d | 30–150 mM; 6 h–3 d | 50 mM; 21 d | 50–100 mM; 3–42 d | 100 mM; 40–60 d | 250 mM; 14 d | 85–250 mM; 14–30 d |
| Electrolyte leakage increase | 50–100 mM; 1 d | 100 mM; 6 h–15 d | 100 mM; 6 h | 100 mM; 8 d | 100 mM; 30 d | 100 mM; 2 d | 100–250 mM; 12–14 d | - |
| Stomatal conductance decrease | 100 mM; 7–12 d | 100 mM; 15 d | 100 mM; 30 d | 60 mM; 12 h | 100 mM; 7–21 d | 50–100 mM; 5–35 d | - | - |
| Water potential | 100 mM; 2 h | 100 mM; 4 d | 100 mM; 48 h | 60 mM; 20 d | 100 mM; 5–70 d | 25–100 mM; 1–54 d | - | 150–400 mM; 24–42 d |
| Proline increase | - | 100 mM; 14–15 d | - | 50 mM; 1 d | 50–100 mM; 7–42 d | 100–150 mM; 7–15 d | - | 85–600 mM; 30–45 d |
| Glycine betaine increase | - | - | - | 50 mM; 21 d | - | 100 mM; 7 d | - | 200–500 mM; 15 d |
| Reference | [83,84,85,86] | [87,88,89,90,91] | [92,93] | [94,95,96,97] | [98,99,100,101,102] | [103,104,105,106] | [107,108] | [109,110,111] |
| Groups/Class | Genes | Inductions of Genes under Salinity Conditions | Plant Species | Reference | ||
|---|---|---|---|---|---|---|
| NaCl Concentrations | Time | |||||
| Stress sensors | Ca sensor kinase | CIPK11 CIPK21 | 150 mM | 24 h | A. thaliana | [151] |
| CBL2 | 250 mM | 24 h | T. aestivum | [152] | ||
| Protein kinase | SOS2 SOS3 | 100–200 mM | 3–6 h | A. thaliana | [153,154,155] | |
| Ion balance regulators | Na transporters | NHX1 | 100 mM–250 mM | 3 h–21 d | Salicornia, T. aestivum, O. sativa | [38,152,156,157] |
| Chloride anion channel | CLC | 150–200 mM | 3–9 d | O. sativa, A. thaliana | [157,158] | |
| Cation channel | ERD4 GLRX | 200 mM–300 mM | 24 h–5 d | A. thaliana; T. aestivum | [159,160] | |
| Transferases | GST | 150 mM–255 mM | 6–48 h | T. aestivum, A. thaliana, R | [152,161,162,163] | |
| ROS regulators | ROS scavengers | APX | 50–255 mM | 48 h–15 d | T. aestivum, O. sativa, Z. mays, S.lycopersicum, Ryegrass | [30,45,161,164,165] |
| CAT | 25–255 mM | 48 h–7 d | T. aestivum, Z. mays, S. lycopersicum, O. sativa, Ryegrass | [30,45,157,161,164,165] | ||
| SOD | 50–250 mM | 4–75 d | T. aestivum, S. lycopersicum, Ryegrass | [30,164,165] | ||
| Redox enzymes | GR | 100–150 mM | 24 h–7 d | A. thaliana, T. aestivum | [30,162] | |
| LOX | 300 mM | 5 d | T. aestivum | [160] | ||
| PAO | 50–250 mM | 3 h–5 d | T. aestivum, A. thaliana | [160,166] | ||
| Cell protectors | Proline synthesis | P5CR | 100 mM–250 mM | 2 h–6 d | O. sativa, T. aestivum, S. lycopersicum | [167,168,169] |
| P5CS | 100 mM–255 mM | 2 h–6 d | A. thaliana, O. sativa, S. lycopersicum, Ryegrass | [161,167,168,170,171] | ||
| Polyamine biosynthesis | ADC | 100–250 mM | 5–7 d | T. aestivum, O. sativa | [160,172] | |
| Senescence regulators | Transcriptional factors | NAC | 150 mM–200 mM | 12 h–24 h | A. thaliana, Salicornia | [162,173] |
| NAP | 150–200 mM | 5 h–6 d | A. thaliana, O. sativa | [174,175] | ||
| ANAC092 | 150 mM | 6–24 h | A. thaliana | [151,176] | ||
| WRKY53 | 150 mM | 4 d | A. thaliana | [177] | ||
| WRKY25 | 150 mM | 6 h | A. thaliana | [162] | ||
| Protease | SAG12 | 150 mM | 4 d | A. thaliana | [176,177] | |
| Nitrogen remobilization | GDH2 | 100–150 mM | 48 h–6 d | O. sativa, A. thaliana | [163,178] | |
| Parameters | Plants | References |
|---|---|---|
| Higher accumulation of Na+ in old leaves than young leaves | Wheat, Barley, Rice, Maize, Sunflower | [93,133,134,187,189,190,191,192,193] |
| Higher accumulation of Cl− in old leaves than young leaves | Wheat and Barley, Sunflower | [187,189,191] |
| Higher photochemical efficiency decrease in old leaves than young leaves | Wheat and Barley, Rice | [93,187] |
| Higher decrease of total chlorophyll content in old leaves than young leaves | Wheat, Barley, Sunflower | [93,192,194] |
| Higher soluble protein decrease in old leavesthan young leaves | Rice, Wheat | [93,195] |
| Higher increase in MDA content in old leaves than young leaves than young leaves | Rice, Maize | [93,133,134] |
| Higher electrolyte leakage in older leaves than young leaves | Rice, Maize | [93,134] |
| Higher ROS reduction and H2O2 generation in older leaves than young leaves | Rice, Maize | [133,134] |
| Higher increase in catalase activity in older leaves compare to young | Rice, Maize | [133,134] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soltabayeva, A.; Ongaltay, A.; Omondi, J.O.; Srivastava, S. Morphological, Physiological and Molecular Markers for Salt-Stressed Plants. Plants 2021, 10, 243. https://doi.org/10.3390/plants10020243
Soltabayeva A, Ongaltay A, Omondi JO, Srivastava S. Morphological, Physiological and Molecular Markers for Salt-Stressed Plants. Plants. 2021; 10(2):243. https://doi.org/10.3390/plants10020243
Chicago/Turabian StyleSoltabayeva, Aigerim, Assel Ongaltay, John Okoth Omondi, and Sudhakar Srivastava. 2021. "Morphological, Physiological and Molecular Markers for Salt-Stressed Plants" Plants 10, no. 2: 243. https://doi.org/10.3390/plants10020243
APA StyleSoltabayeva, A., Ongaltay, A., Omondi, J. O., & Srivastava, S. (2021). Morphological, Physiological and Molecular Markers for Salt-Stressed Plants. Plants, 10(2), 243. https://doi.org/10.3390/plants10020243