
Telomere Length in Norway Spruce during Somatic Embryogenesis and Cryopreservation


Abstract
:1. Introduction
2. Results
2.1. SE Initiation
2.2. Recovery from Cryopreservation
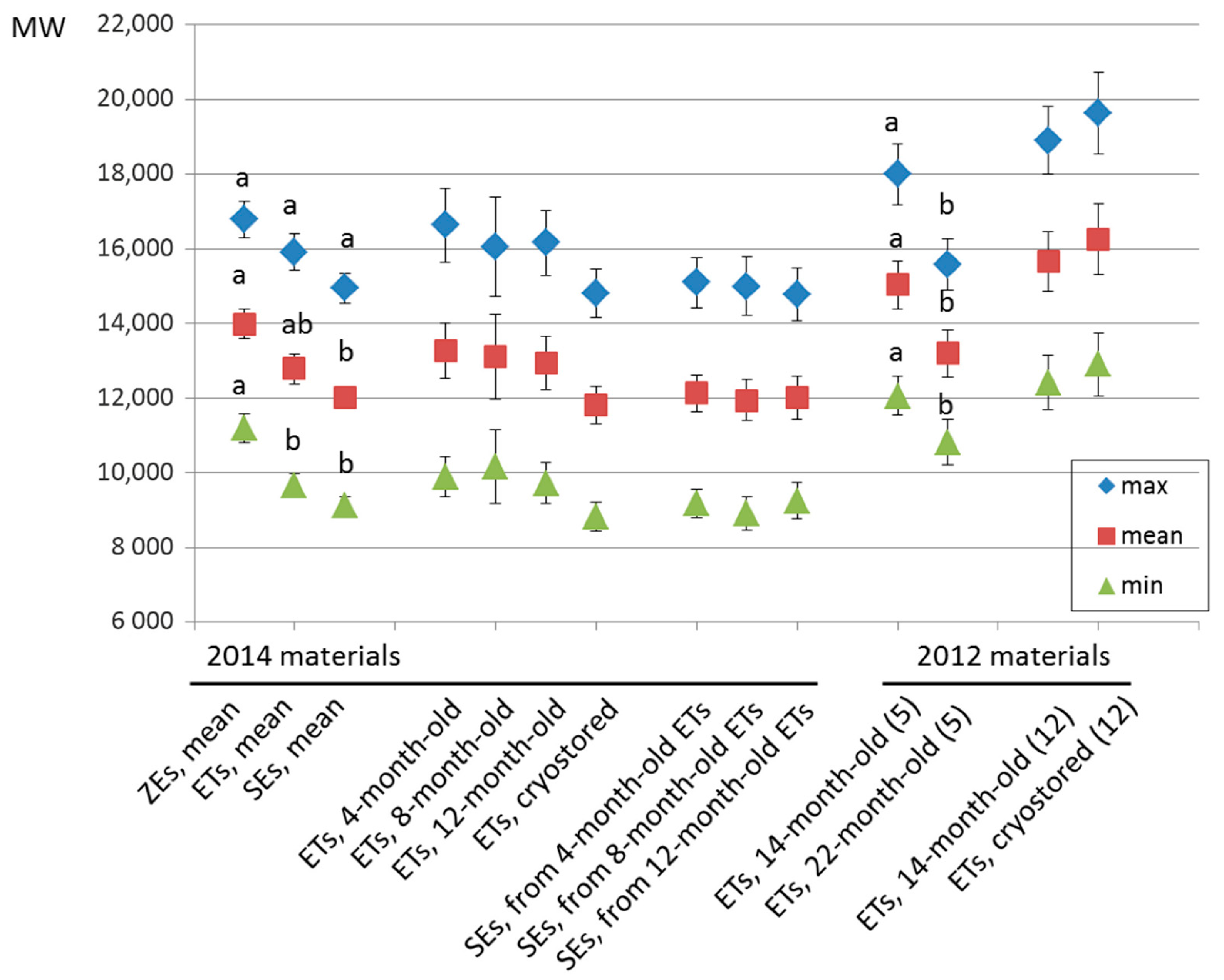
2.3. Embryo Production Capacity
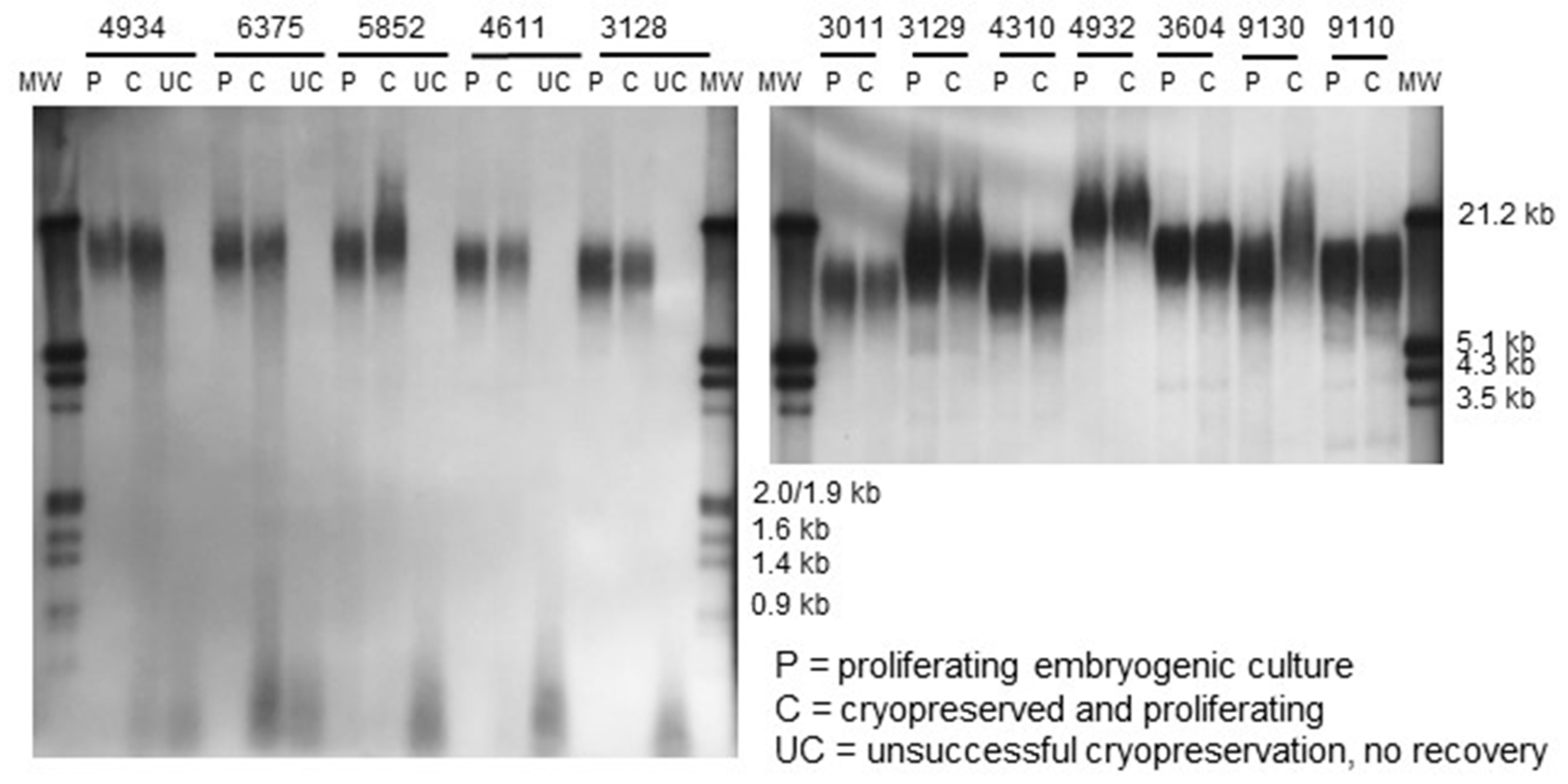
2.4. Telomere Length in Embryogenic Materials
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Maintenance and Cryopreservation of Embryogenic Lines
4.3. Evaluation of Somatic Embryo Production Capacity
4.4. DNA Extraction and Southern Blot Analysis of Telomeric Repeats
4.5. Experimental Design and Statistical Analyses
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year and Cross | Number of Explants | Initiation % |
|---|---|---|
| 2014 | ||
| E18 × E2273 | 200 | 71.5 |
| E18 × E436 | 201 | 67.2 |
| E18 × E5535 | 200 | 50.5 |
| E207 × E1373 | 202 | 72.8 |
| E207 × E252 | 200 | 30.0 |
| E2105 × E2283 | 200 | 57.0 |
| E2105 × E3354 | 67 | 53.7 |
| E212 × E54 | 200 | 63.5 |
| E46 × E3222 | 400 | 61.0 |
| E462 × E1369 | 200 | 36.5 |
| E9 × E1361 | 200 | 77.0 |
| E162 × E81 | 200 | 51.0 |
| E212 × E1518 | 11 | 54.6 |
| E81 × E3224 | 132 | 68.9 |
| E162 × E2149 | 202 | 37.6 |
| E1551 × E2229 | 99 | 67.7 |
| E799 × E1366 | 46 | 93,48 |
| 2012 | ||
| E2515 × K805 | 41 | 95.1 |
| E2853 × E231 | 143 | 95.8 |
| E2853 × E330 | 135 | 68.9 |
| E318 × E231 | 72 | 69.4 |
| E318 × K805 | 329 | 61.4 |
| E329 × K805 | 22 | 100.0 |
| K264 × E231 | 68 | 89.7 |
| K264 × E330 | 89 | 85.4 |
| Norway Spruce Genotypes Used in | ||||
|---|---|---|---|---|
| Year and Cross | Embryo Production Experiments | Telomere Length Measurements | ||
| 2014 | Proliferating ETs of 4, 8, or 12 months in age; cryostored proliferating ETs, 4 + 2 months in age | Proliferating ETs of 4, 8, or 12 months in age; cryostored proliferating ETs, 4 + 2 months in age | Mature somatic embryos originating in ETs of 4, 8, or 12 months in age | Immature zygotic embryos, excised at time of SE initiation |
| E18 × E2273 | yes | |||
| E18 × E436 | 653 | |||
| E18 × E5535 | 872 | |||
| E207 × E1373 | yes | |||
| E207 × E252 | 1206 | 1206 | 1206 | |
| 1305 | ||||
| E2105 × E2283 | 1548 | 1548 | 1548 | yes |
| E2105 × E3354 | 1606 | |||
| 1607 | ||||
| E212 × E54 | 2181 | |||
| E46 × E3222 | 5820 | 5820 | 5820 | |
| 2833 | ||||
| E462 × E1369 | 3022 | 3022 | 3022 | yes |
| E9 × E1361 | 4027 | 4031 | 4031 | yes |
| 4031 | ||||
| E162 × E81 | 243 | 243 | 243 | yes |
| 259 | 259 | 259 | ||
| E212 × E1518 | yes | |||
| E81 × E3224 | 3620 | yes | ||
| E162 × E2149 | 30 | |||
| E1551 × E2229 | 4623 | 4623 | 4623 | |
| 4639 | 4639 | 4639 | ||
| E799 × E1366 | 5101 | 5101 | 5101 | |
| 5109 | 5109 | 5109 | ||
| Norway Spruce Genotypes Used in | |||
|---|---|---|---|
| Year and Cross | Telomere Length Measurements | ||
| 2012 | Proliferating ETs of 14 months in age; Cryostored proliferating ETs, 14 + 1 months in age | Cryostored ETs showing no recovery | Proliferating ETs of 22 months in age |
| E2515 × K805 | 3011 | ||
| E2853 × E231 | 3128 | 3128 | |
| 3129 | |||
| E2853 × E330 | 4310 | 4310 | |
| E318 × E231 | 4932 | 4934 | 4934 |
| 4934 | |||
| E318 × K805 | 6375 | 6375 | 6375 |
| E329 × K805 | 9130 | 9130 | |
| K264 × E231 | 4611 | 4611 | 4611 |
| 9110 | |||
| K264 × E330 | 3604 | 5852 | |
| 5852 | |||
References
- McKnight, T.D.; Riha, K.; Shippen, D.E. Telomeres, telomerase, and stability of the plant genome. Plant Mol. Biol. 2002, 48, 331–337. [Google Scholar] [CrossRef]
- Watson, J.M.; Riha, K. Telomeres, ageing, and plants: From weeds to Methuselah—A mini-review. Gerontology 2011, 57, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Chatelain, M.; Drobniak, S.M.; Szulkin, M. The association between stressors and telomeres in non-human vertebrates: A meta-analysis. Ecol. Lett. 2020, 23, 381–398. [Google Scholar] [CrossRef] [PubMed]
- Passos, J.; von Zglinicki, T. Mitochondria, telomeres and cell senescence. Exp. Gerontol. 2005, 40, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Flanary, B.E.; Kletetschka, G. Analysis of telomere length and telomerase activity in tree species of various life-spans, and with age in the bristlecone pine Pinus longaeva. Biogerontology 2005, 6, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Qiao, N.; Song, H.; Hua, X.; Du, J.; Hai, L.; Fenglan, L. Comparative analysis of telomeric restriction fragment lengths in different tissues of Ginkgo biloba trees of different age. J. Plant Res. 2007, 120, 523–528. [Google Scholar] [CrossRef]
- Song, H.; Liu, D.; Chen, X.; Ying, Z.; Zhang, B.; Li, F.; Lu, H. Change of season-specific telomere lengths in Ginkgo biloba L. Mol. Biol. Rep. 2010, 37, 819–824. [Google Scholar] [CrossRef]
- Moriguchi, R.; Kato, K.; Kanahama, K.; Kanayama, Y.; Kikuchi, H. Analysis of telomere lengths in apple and cherry trees. Acta Hortic. 2007, 738, 389–395. [Google Scholar] [CrossRef]
- Aronen, T.; Ryynänen, L. Variation in telomeric repeats of Scots pine (Pinus sylvestris L.). Tree Genet. Genomes 2012, 8, 267–275. [Google Scholar] [CrossRef]
- Aronen, T.; Ryynänen, L. Silver birch telomeres shorten in tissue culture. Tree Genet. Genomes 2014, 10, 67–74. [Google Scholar] [CrossRef]
- Mu, Y.; Ren, L.-F.; Xun, Z.-L.; Zhang, D.-D.; Song, H.; Lu, H.; Li, F.-L.; Liu, D. Sex- and season-dependent differences in telomere length and telomerase activity in the leaves of ash and willow. SpringerPlus 2014, 3, 163. [Google Scholar] [CrossRef] [Green Version]
- Rani, V.; Raina, S.N. Genetic fidelity of organized meristem-derived micropropagated plants: A critical reapprasail. Vitr. Cell. Dev. Biol. Plant 2000, 36, 319–330. [Google Scholar] [CrossRef]
- Rescalvo-Morales, A.; Monja-Mio, K.M.; Robert, M.L.; Sanchez-Teyer, L.F. Telomere length in Agave tequilana Weber plants during the in vitro to ex vitro transition. Plant Cell Tissue Organ Cult. PCTOC 2019, 136, 133–140. [Google Scholar] [CrossRef]
- Kilian, A.; Stiff, C.; Kleinhofs, A. Barley telomeres shorten during differentiation but grow in callus culture. Proc. Natl. Acad. Sci. USA 1995, 92, 9555–9559. [Google Scholar] [CrossRef] [Green Version]
- Riha, K.; Fajkus, J.; Siroky, J.; Vystok, B. Developmental control of telomere lengths and telomerase activity in plants. Plant Cell 1998, 10, 1691–1698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fajkus, J.; Fulneckova, J.; Hulanova, M.; Berkova, K.; Riha, K.; Matyasek, R. Plant cells express telomerase activity upon transfer to callus culture, without extensively changing telomere length. Mol. Gen. Genet. 1998, 260, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Sutton, B. Commercial delivery of genetic improvement to conifer plantations using somatic embryogenesis. Ann. For. Sci. 2002, 59, 657–661. [Google Scholar] [CrossRef]
- Högberg, K.A.; Varis, S. Vegetative propagation of Norway spruce: Experiences and present situation in Sweden and Finland, In Vegetative Propagation of Forest Trees; Park, Y.-S., Bonga, J.M., Moon, H.-K., Eds.; Korea Forest Research Institute: Seoul, Korea, 2016; pp. 538–550. [Google Scholar]
- Bonga, J. Conifer clonal propagation in tree improvement programs. In Vegetative Propagation of Forest Trees; Park, Y., Bonga, J., Moon, H., Eds.; National Institute of Forest Science: Seoul, Korea, 2016; pp. 3–31. [Google Scholar]
- Adams, G.; Kunze, H.; McCartney, A.; Millican, S.; Park, Y. An industrial perspective on the use of advanced reforestation stock technologies. In Vegetative Propagation of Forest Trees; Park, Y., Bonga, J., Moon, H., Eds.; National Institute of Forest Science: Seoul, Korea, 2016; pp. 323–334. [Google Scholar]
- Find, J. Towards industrial production of tree varieties through somatic embryogenesis and other vegetative propagation technologies: Nordmanns fir (Abies nordamanniana (Steven) Spach)—From research laboratory to production. In Vegetative Propagation of Forest Trees; Park, Y., Bonga, J., Moon, H., Eds.; National Institute of Forest Science: Seoul, Korea, 2016; pp. 528–537. [Google Scholar]
- Lelu-Walter, M.A.; Thompson, D.; Harvengt, L.; Sanchez, L.; Toribio, M.; Pâques, L.E. Somatic embryogenesis in forestry with a focus on Europe: State-of-the-art, benefits, challenges and future direction. Tree Genet. Genomes 2013, 9, 883–899. [Google Scholar] [CrossRef]
- Hakman, I.; von Arnold, S. Plantlet regeneration through somatic embryogenesis in Picea abies (Norway spruce). J. Plant Physiol. 1985, 121, 149–158. [Google Scholar] [CrossRef]
- Klimaszewska, K.; Lachance, D.; Pelletier, G.; Lelu, M.-A.; Séguin, A. Regeneration of transgenic Picea glauca, P. mariana and P. abies after cultivation of embryogenic tissue with Agrobacterium tumefaciens. Vitr. Cell. Dev. Biol. Plant. 2001, 37, 748–755. [Google Scholar] [CrossRef]
- Vagner, M.; Fischerova, L.; Spackova, J.; Vondrakova, Z. Somatic embryogenesis in Norway spruce. In Protocols for Somatic Embryogenesis in Woody Plants; Jain, S.M., Gupta, P., Eds.; Forestry Sciences 77; Springer: Dordrecht, The Netherlands, 2005; pp. 141–155. [Google Scholar]
- Varis, S. Norway spruce Picea abies (L.) Karst. In Step Wise Protocols for Somatic Embryogenesis of Important Woody Plants; Jain, S.M., Gupta, P., Eds.; Forestry Sciences 84; Springer: Dordrecht, The Netherlands, 2018; pp. 255–268. [Google Scholar] [CrossRef]
- Norgaard, J.V.; Duran, V.; Johnsen, Ø.; Krogstrup, P.; Baldursson, S.; von Arnold, S. Variations in cryotolerance of embryogenic Picea abies cell lines and the association to genetic, morphological, and physiological factors. Can. J. For. Res. 1993, 23, 2560–2567. [Google Scholar] [CrossRef]
- Varis, S.; Ahola, S.; Jaakola, L.; Aronen, T. Reliable and practical methods for cryopreservation of embryogenic cultures and cold storage of somatic embryos of Norway spruce. Cryobiology 2017, 76, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Egertsdotter, U.; Iftikhar, A.; Clapham, D. Automation and Scale Up of Somatic Embryogenesis for Commercial Plant Production, With Emphasis on Conifers. Front. Plant Sci. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Välimäki, S.; Paavilainen, L.; Tikkinen, M.; Salonen, F.; Varis, S.; Aronen, T. Production of Norway spruce embryos in a temporary immersion system (TIS). Vitr. Cell. Dev. Biol. Plant 2020. [Google Scholar] [CrossRef]
- Tikkinen, M.; Varis, S.; Välimäki, S.; Nikkanen, T.; Aronen, T. Somatic embryogenesis of Norway spruce in Finland—Seven years from start to first commercial pilots. In Proceedings of the 5th international Conference of the IUFRO Unit 2.09.02 on “Clonal Trees in the Bioeconomy Age: Opportunities and Challenges”, Coimbra, Portugal, 10–15 September 2018; pp. 166–172. [Google Scholar]
- Miguel, C.M.; Rupps, A.; Raschke, J.; Rodrigues, A.; Trontin, J.-F. Impact of molecular studies on somatic embryogenesis development for implementation in conifer multi-varietal forestry. In Vegetative Propagation of Forest Trees; Park, Y., Bonga, J., Moon, H., Eds.; National Institute of Forest Science: Seoul, Korea, 2016; pp. 373–421. [Google Scholar]
- Aronen, T.; Pehkonen, T.; Ryynänen, L. Enhancement of somatic embryogenesis from immature zygotic embryos of Pinus Sylvestris. Scand. J. For. Res. 2009, 24, 372–383. [Google Scholar] [CrossRef]
- Filonova, L.H.; Bozhkov, P.V.; von Arnold, S. Developmental pathway of somatic embryogenesis in Picea abies as revealed by time-laps tracking. J. Exp. Bot. 2000, 51, 249–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breton, D.; Harvengt, L.; Trontin, J.-F.; Bouvet, A.; Favre, J.-M. Long-term subculture randomly affects morphology and subsequent maturation of early somatic embryos in maritime pine. Plant Cell Tiss. Organ. Cult. PCTOC 2006. [Google Scholar] [CrossRef]
- Klimaszewska, K.; Noceda, C.; Pelletier, G.; Label, P.; Rodriguez, R.; Lelu-Walter, M.A. Biological characterization of young and aged embryogenic cultures of Pinus pinaster (Ait.). Vitr. Cell. Dev. Biol. Plant 2009, 45, 20–33. [Google Scholar] [CrossRef]
- Chalupka, W. Reproductive development. In Biology and Ecology of Norway Spruce; Tjoelker, M.G., Boratynski, A., Bugala, E., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 97–106. [Google Scholar]
- Lüttge, U.; Hertel, B. Diurnal and annual rhythms in trees. Trees 2009, 23, 683–700. [Google Scholar] [CrossRef]
- Gyllenstrand, N.; Clapham, D.; Källman, T.; Lagercrantz, U. A Norway Spruce Flowering Locus T Homolog is Implicated in Control of Growth Rhythm in Conifers. Plant Physiol. 2007, 144, 248–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benson, E. Cryopreservation theory. In Plant Cryopreservation. A Practical Guide; Reed, B.M., Ed.; Springer: New York, NY, USA, 2008; pp. 15–32. [Google Scholar]
- Litvay, D.J.; Verma, D.C.; Johnson, M.A. Influence of loblolly pine (Pinus taeda L.) culture medium and its components on growth and somatic embryogenesis of wild carrot (Daucus carota L.). Plant Cell Rep. 1985, 4, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Lelu-Walter, M.A.; Bernier-Cardou, M.; Klimaszewska, K. Clonal plant production from self- and cross-pollinated seed families of Pinus sylvestris (L.) through somatic embryogenesis. Plant Cell Tissue Organ. Cult. PCTOC 2008, 92, 31–45. [Google Scholar] [CrossRef]
- Lodhi, M.A.; Ye, G.-N.; Weeden, N.F.; Reisch, B.I. A simple and efficient method for DNA extraction from grapevine cultivars and Vitis species. Plant Mol. Biol. Rep. 1994, 12, 6–13. [Google Scholar] [CrossRef]
- Valjakka, M.; Aronen, T.; Kangasjärvi, J.; Vapaavuori, E.; Häggman, H. Genetic transformation of silver birch (Betula pendula) by particle bombardment. Tree Physiol. 2000, 20, 607–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aronen, T.; Tiimonen, H.; Tsai, C.-J.; Jokipii, S.; Chen, X.; Chiang, V.; Häggman, H. Altered lignin in transgenic silver birch (Betula pendula) expressing PtCOMT gene. In Sustainable Forestry, Wood Products & Biotechnology; Espinel, S., Barredo, Y., Ritter, E., Eds.; DFE-AFA Press: Vitoria-Gasteiz, Spain, 2003; pp. 149–161. [Google Scholar]
- Cox, A.V.; Bennett, S.T.; Parokonny, A.S.; Kenton, A.; Callimassia, M.A.; Bennett, M.D. Comparison of plant telomere locations using a PCR generated synthetic probe. Ann. Bot. 1993, 72, 239–247. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aronen, T.; Virta, S.; Varis, S. Telomere Length in Norway Spruce during Somatic Embryogenesis and Cryopreservation. Plants 2021, 10, 416. https://doi.org/10.3390/plants10020416
Aronen T, Virta S, Varis S. Telomere Length in Norway Spruce during Somatic Embryogenesis and Cryopreservation. Plants. 2021; 10(2):416. https://doi.org/10.3390/plants10020416
Chicago/Turabian StyleAronen, Tuija, Susanna Virta, and Saila Varis. 2021. "Telomere Length in Norway Spruce during Somatic Embryogenesis and Cryopreservation" Plants 10, no. 2: 416. https://doi.org/10.3390/plants10020416
APA StyleAronen, T., Virta, S., & Varis, S. (2021). Telomere Length in Norway Spruce during Somatic Embryogenesis and Cryopreservation. Plants, 10(2), 416. https://doi.org/10.3390/plants10020416