
A Plant Based Modified Biostimulant (Copper Chlorophyllin), Mediates Defense Response in Arabidopsis thaliana under Salinity Stress



Abstract
:1. Introduction
2. Results
2.1. Pretreatment with Cu-chl Reduces H2O2
2.2. Cu-chl Protects Arabidopsis thaliana Seedlings from Herbicidal Damage by Reducing H2O2
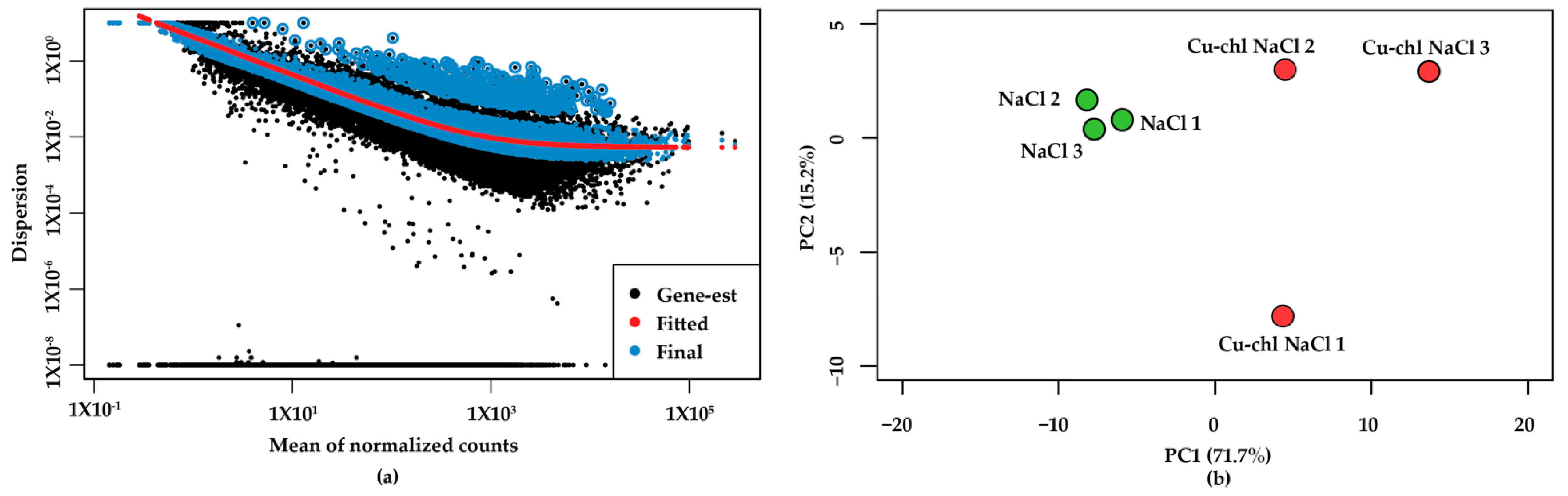
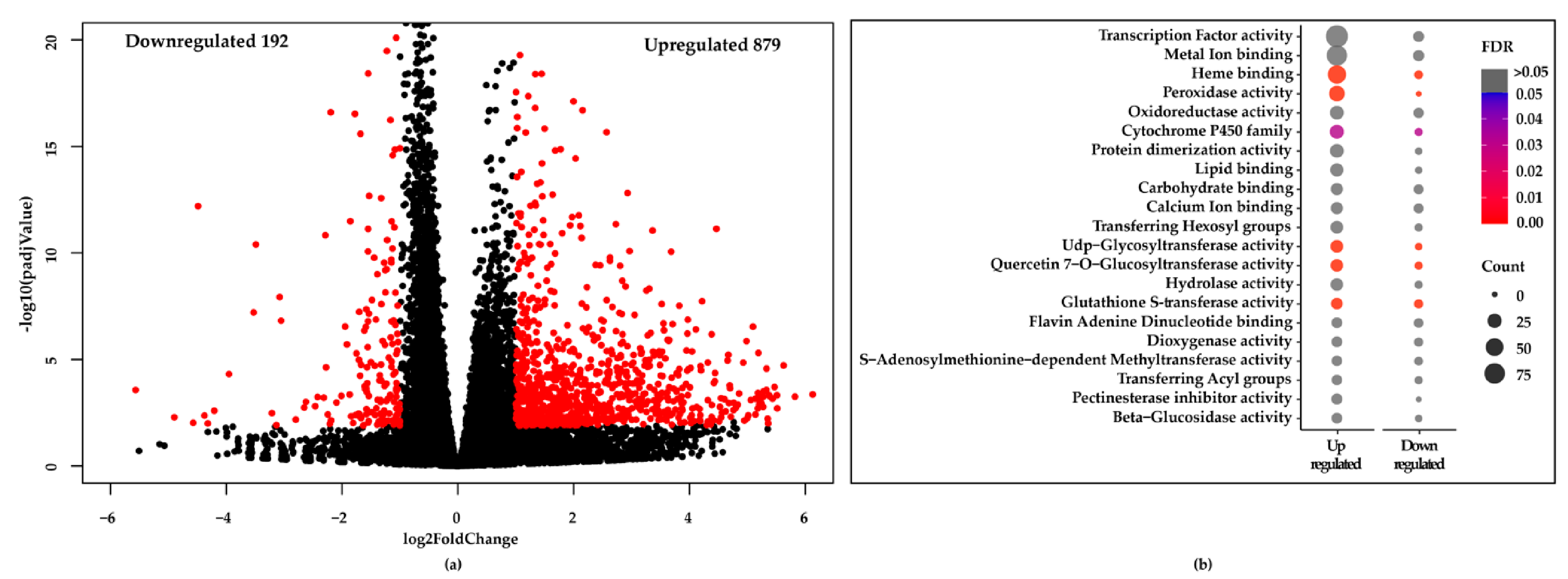
2.3. RNA-Seq Reveals the Molecular Mechanism of Cu-chl
2.4. RNA-Seq Validation by Real-Time RT-PCR (qPCR)
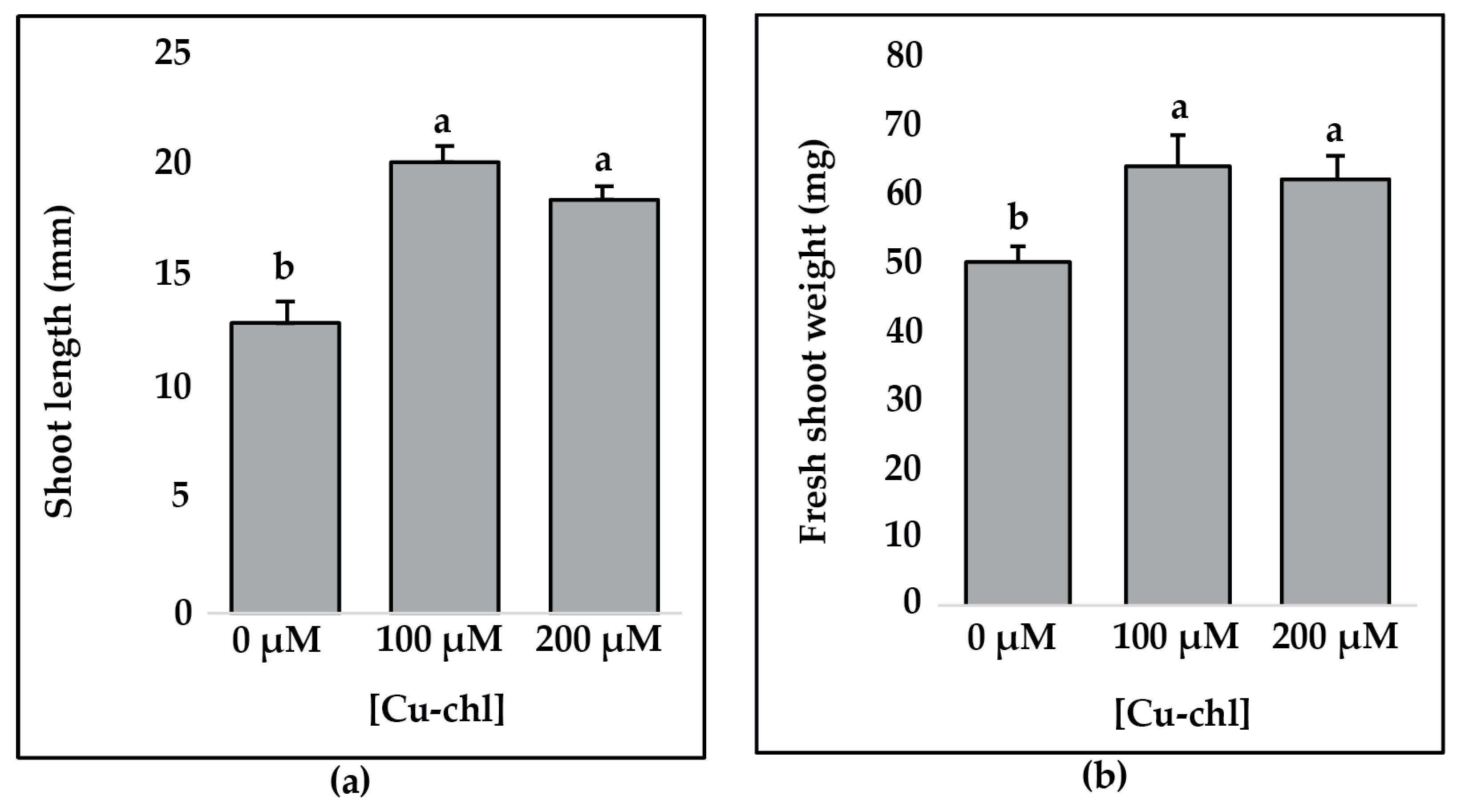
2.5. Effect of Cu-chl on Arabidopsis thaliana Growth
3. Discussion
4. Materials and Methods
4.1. Seed Sterilization and Stratification
4.2. H2O2 Accumulation Measurement via DAB (3,3′-Diaminobenzidin) and Amplex® Red Assay under Salt Stress
4.3. H2O2 Accumulation Measurement under Herbicide Stress
4.4. Cu-chl and Salt Stress Application for RNA-Seq and qPCR
4.5. Effect of Cu-chl on Seedling Growth
4.6. RNA Sequencing and Analysis
4.7. cDNA Synthesis and qPCR
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Den Herder, G.; Van Isterdael, G.; Beeckman, T.; De Smet, I. The Roots of a New Green Revolution. Trends Plant Sci. 2010, 15, 600–607. [Google Scholar] [CrossRef]
- Mahajan, S.; Tuteja, N. Cold, Salinity and Drought Stresses: An Overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Von Braun, J. The World Food Situtation: New Driving Forces and Required Actions; International Food Policy Research Institute: Washington, DC, USA, 2007. [Google Scholar]
- Fahad, S.; Hussain, S.; Matloob, A.; Khan, F.A.; Khaliq, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.; Ullah, N.; et al. Phytohormones and Plant Responses to Salinity Stress: A Review. Plant Growth Regul. 2015, 75, 391–404. [Google Scholar] [CrossRef]
- Shabala, S. Learning from Halophytes: Physiological Basis and Strategies to Improve Abiotic Stress Tolerance in Crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under Drought and Salt Stress: Regulation Mechanisms from Whole Plant to Cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Qadir, M.; Quillérou, E.; Nangia, V.; Murtaza, G.; Singh, M.; Thomas, R.J.; Drechsel, P.; Noble, A.D. Economics of Salt-Induced Land Degradation and Restoration. Nat. Resour. Forum 2014, 38, 282–295. [Google Scholar] [CrossRef]
- Riyazuddin, R.; Verma, R.; Singh, K.; Nisha, N.; Keisham, M.; Bhati, K.K.; Kim, S.T.; Gupta, R. Ethylene: A Master Regulator of Salinity Stress Tolerance in Plants. Biomolecules 2020, 10, 959. [Google Scholar] [CrossRef] [PubMed]
- Shahid, S.A.; Zaman, M.; Heng, L. Soil salinity: Historical perspectives and a world overview of the problem. In Guideline for Salinity Assessment, Mitigation and Adaptation Using Nuclear and Related Techniques; Springer: Berlin/Heidelberg, Germany, 2018; pp. 43–53. [Google Scholar]
- Jamil, A.; Riaz, S.; Ashraf, M.; Foolad, M.R. Gene Expression Profiling of Plants under Salt Stress. Crit. Rev. Plant Sci. 2011, 30, 435–458. [Google Scholar] [CrossRef]
- Pitman, M.G.; Läuchli, A. Global impact of salinity and agricultural ecosystems. In Salinity: Environment-Plants-Molecules; Springer: Berlin/Heidelberg, Germany, 2002; pp. 3–20. [Google Scholar]
- Ashraf, M. Biotechnological Approach of Improving Plant Salt Tolerance Using Antioxidants as Markers. Biotechnol. Adv. 2009, 27, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P.; Cirillo, C.; De Micco, V.; Arena, C.; De Pascale, S.; Rouphael, Y. Morpho-Anatomical, Physiological and Biochemical Adaptive Responses to Saline Water of Bougainvillea Spectabilis Willd. Trained to Different Canopy Shapes. Agric. Water Manag. 2019, 212, 12–22. [Google Scholar] [CrossRef]
- Dell’Aversana, E.; Hessini, K.; Ferchichi, S.; Fusco, G.M.; Woodrow, P.; Ciarmiello, L.F.; Abdelly, C.; Carillo, P. Salinity Duration Differently Modulates Physiological Parameters and Metabolites Profile in Roots of Two Contrasting Barley Genotypes. Plants 2021, 10, 307. [Google Scholar] [CrossRef]
- Fricke, W. Rapid and Tissue-Specific Accumulation of Solutes in the Growth Zone of Barley Leaves in Response to Salinity. Planta 2004, 219, 515–525. [Google Scholar] [CrossRef]
- Fichman, Y.; Miller, G.; Mittler, R. Whole-Plant Live Imaging of Reactive Oxygen Species. Mol. Plant 2019, 12, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
- Inze, D. Oxidative Stress in Plants; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Li, J.-Y.; Jiang, A.-L.; Zhang, W. Salt Stress-Induced Programmed Cell Death in Rice Root Tip Cells. J. Integr. Plant Biol. 2007, 49, 481–486. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive Oxygen Species and Antioxidant Machinery in Abiotic Stress Tolerance in Crop Plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Veal, E.A.; Day, A.M.; Morgan, B.A. Hydrogen Peroxide Sensing and Signaling. Mol. Cell 2007, 26, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Ullah, F.; Zhou, D.-X.; Yi, M.; Zhao, Y. Mechanisms of ROS Regulation of Plant Development and Stress Responses. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Decros, G.; Baldet, P.; Beauvoit, B.; Stevens, R.; Flandin, A.; Colombié, S.; Gibon, Y.; Pétriacq, P. Get the Balance Right: ROS Homeostasis and Redox Signalling in Fruit. Front. Plant Sci. 2019, 10, 1091. [Google Scholar] [CrossRef] [PubMed]
- Tsukagoshi, H.; Busch, W.; Benfey, P.N. Transcriptional Regulation of ROS Controls Transition from Proliferation to Differentiation in the Root. Cell 2010, 143, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Baek, D.; Cha, J.-Y.; Kang, S.; Park, B.; Lee, H.-J.; Hong, H.; Chun, H.J.; Kim, D.H.; Kim, M.C.; Lee, S.Y.; et al. The Arabidopsis a Zinc Finger Domain Protein ARS1 Is Essential for Seed Germination and ROS Homeostasis in Response to ABA and Oxidative Stress. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef]
- Kollist, H.; Zandalinas, S.I.; Sengupta, S.; Nuhkat, M.; Kangasjärvi, J.; Mittler, R. Rapid Responses to Abiotic Stress: Priming the Landscape for the Signal Transduction Network. Trends Plant Sci. 2019, 24, 25–37. [Google Scholar] [CrossRef]
- Neill, S.J.; Desikan, R.; Clarke, A.; Hurst, R.D.; Hancock, J.T. Hydrogen Peroxide and Nitric Oxide as Signalling Molecules in Plants. J. Exp. Bot. 2002, 53, 1237–1247. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, England, 2015. [Google Scholar]
- Noctor, G.; Foyer, C.H. Ascorbate and Glutathione: Keeping Active Oxygen under Control. Annu. Rev. Plant Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J. Al; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant Biostimulants: Definition, Concept, Main Categories and Regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Li, L.; Wang, Y.; Shen, W. Roles of Hydrogen Sulfide and Nitric Oxide in the Alleviation of Cadmium-Induced Oxidative Damage in Alfalfa Seedling Roots. Biometals 2012, 25, 617–631. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Colla, G. Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural Uses of Plant Biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef]
- Ong, T.; Whong, W.-Z.; Stewart, J.; Brockman, H.E. Chlorophyllin: A Potent Antimutagen against Environmental and Dietary Complex Mixtures. Mutat. Res. Lett. 1986, 173, 111–115. [Google Scholar] [CrossRef]
- Sato, M.; Fujimoto, I.; Sakai, T.; Aimoto, T.; Kimura, R.; Murata, T. Effect of Sodium Copper Chlorophyllin on Lipid Peroxidation. IX.: On the Antioxidative Components in Commercial Preparations of Sodium Copper Chlorohyllin. Chem. Pharm. Bull. 1986, 34, 2428–2434. [Google Scholar] [CrossRef]
- Kamat, J.P.; Boloor, K.K.; Devasagayam, T.P.A. Chlorophyllin as an Effective Antioxidant against Membrane Damage in Vitro and Ex Vivo. Biochim. Et Biophys. Acta (Bba)-Mol. Cell Biol. Lipids 2000, 1487, 113–127. [Google Scholar] [CrossRef]
- Zhang, X.; Goatley, M.; Conner, J.; Wilkins, M.; Teshler, I.; Liu, J.; Fefer, M.; Ckurshumova, W. Copper Chlorophyllin Impacts on Growth and Drought Stress Tolerance of Tomato Plants. HortScience 2019, 54, 2195–2201. [Google Scholar] [CrossRef]
- Song, N.H.; Le Yin, X.; Chen, G.F.; Yang, H. Biological Responses of Wheat (Triticum Aestivum) Plants to the Herbicide Chlorotoluron in Soils. Chemosphere 2007, 68, 1779–1787. [Google Scholar] [CrossRef] [PubMed]
- Scarpeci, T.E.; Valle, E.M. Rearrangement of Carbon Metabolism in Arabidopsis Thaliana Subjected to Oxidative Stress Condition: An Emergency Survival Strategy. Plant Growth Regul. 2008, 54, 133–142. [Google Scholar] [CrossRef]
- Chen, R.; Sun, S.; Wang, C.; Li, Y.; Liang, Y.; An, F.; Li, C.; Dong, H.; Yang, X.; Zhang, J.; et al. The Arabidopsis PARAQUAT RESISTANT2 Gene Encodes an S-Nitrosoglutathione Reductase That Is a Key Regulator of Cell Death. Cell Res. 2009, 19, 1377–1387. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44. [Google Scholar]
- Chapman, J.M.; Muhlemann, J.K.; Gayomba, S.R.; Muday, G.K. RBOH-Dependent ROS Synthesis and ROS Scavenging by Plant Specialized Metabolites to Modulate Plant Development and Stress Responses. Chem. Res. Toxicol. 2019, 32, 370–396. [Google Scholar] [CrossRef]
- Llorente, F.; López-Cobollo, R.M.; Catalá, R.; Mart\’\inez-Zapater, J.M.; Salinas, J. A Novel Cold-Inducible Gene from Arabidopsis, RCI3, Encodes a Peroxidase That Constitutes a Component for Stress Tolerance. Plant J. 2002, 32, 13–24. [Google Scholar] [CrossRef]
- Cai, S.; Lashbrook, C.C. Stamen Abscission Zone Transcriptome Profiling Reveals New Candidates for Abscission Control: Enhanced Retention of Floral Organs in Transgenic Plants Overexpressing Arabidopsis ZINC FINGER PROTEIN2. Plant Physiol. 2008, 146, 1305–1321. [Google Scholar] [CrossRef]
- Mentewab, A.; Cardoza, V.; Stewart, C.N., Jr. Genomic Analysis of the Response of Arabidopsis Thaliana to Trinitrotoluene as Revealed by CDNA Microarrays. Plant Sci. 2005, 168, 1409–1424. [Google Scholar] [CrossRef]
- Cominelli, E.; Sala, T.; Calvi, D.; Gusmaroli, G.; Tonelli, C. Over-Expression of the Arabidopsis AtMYB41 Gene Alters Cell Expansion and Leaf Surface Permeability. Plant J. 2008, 53, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Taylor, G.J.; Deyholos, M.K. Transcriptomic Responses to Aluminum Stress in Roots of Arabidopsis Thaliana. Mol. Genet. Genom. 2008, 279, 339. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.G.; Pyo, Y.J.; Cho, J.W.; Cho, M.H. Comparative Proteome Analysis of Differentially Expressed Proteins Induced by K+ Deficiency in Arabidopsis Thaliana. Proteomics 2004, 4, 3549–3559. [Google Scholar] [CrossRef]
- Irshad, M.; Canut, H.; Borderies, G.; Pont-Lezica, R.; Jamet, E. A New Picture of Cell Wall Protein Dynamics in Elongating Cells of Arabidopsis Thaliana: Confirmed Actors and Newcomers. Bmc Plant Biol. 2008, 8, 94. [Google Scholar] [CrossRef]
- Moseyko, N.; Zhu, T.; Chang, H.-S.; Wang, X.; Feldman, L.J. Transcription Profiling of the Early Gravitropic Response in Arabidopsis Using High-Density Oligonucleotide Probe Microarrays. Plant Physiol. 2002, 130, 720–728. [Google Scholar] [CrossRef]
- Hammond, J.P.; Bennett, M.J.; Bowen, H.C.; Broadley, M.R.; Eastwood, D.C.; May, S.T.; Rahn, C.; Swarup, R.; Woolaway, K.E.; White, P.J. Changes in Gene Expression in Arabidopsis Shoots during Phosphate Starvation and the Potential for Developing Smart Plants. Plant Physiol. 2003, 132, 578–596. [Google Scholar] [CrossRef]
- Nikiforova, V.; Freitag, J.; Kempa, S.; Adamik, M.; Hesse, H.; Hoefgen, R. Transcriptome Analysis of Sulfur Depletion in Arabidopsis Thaliana: Interlacing of Biosynthetic Pathways Provides Response Specificity. Plant J. 2003, 33, 633–650. [Google Scholar] [CrossRef]
- Arthikala, M.-K.; Quinto, C. RbohA Coordinates Lateral Root Emergence in Common Bean. Commun. Integr. Biol. 2018, 11, 1–5. [Google Scholar] [CrossRef]
- Zhang, T.; Lai, J.; Li, P.; Sun, W.; Diao, Z.; Wang, J.; Zheng, S. Expression Analysis of RbohA and RbohD Genes in Brassica. Campestris under Different Treatments. Chin. J. Ecol. 2019, 38, 173. [Google Scholar]
- Arthikala, M.-K.; Sánchez-López, R.; Nava, N.; Santana, O.; Cárdenas, L.; Quinto, C. RbohB, a Phaseolus Vulgaris NADPH Oxidase Gene, Enhances Symbiosome Number, Bacteroid Size, and Nitrogen Fixation in Nodules and Impairs Mycorrhizal Colonization. New Phytol. 2014, 202, 886–900. [Google Scholar] [CrossRef]
- Montiel, J.; Arthikala, M.-K.; Quinto, C. Phaseolus Vulgaris RbohB Functions in Lateral Root Development. Plant Signal. Behav. 2013, 8, e22694. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Köster, P.; Schlücking, K.; Balcerowicz, D.; Hashimoto, K.; Kuchitsu, K.; Vissenberg, K.; Kudla, J. CBL1-CIPK26-Mediated Phosphorylation Enhances Activity of the NADPH Oxidase RBOHC, but Is Dispensable for Root Hair Growth. FEBS Lett. 2018, 592, 2582–2593. [Google Scholar] [CrossRef]
- Zhang, T.G.; Li, Q.L.; Diao, Z.H.; Li, P.; Wang, J.; Zheng, S. Effects of Salt and Drought Stresses on Antioxidant System and RbohC and RbohF Genes Expression in Brassica Campestris. Ying Yong Sheng Tai Xue Bao= J. Appl. Ecol. 2019, 30, 969–978. [Google Scholar]
- Orman-Ligeza, B.; Parizot, B.; De Rycke, R.; Fernandez, A.; Himschoot, E.; Van Breusegem, F.; Bennett, M.J.; Périlleux, C.; Beeckman, T.; Draye, X. RBOH-Mediated ROS Production Facilitates Lateral Root Emergence in Arabidopsis. Development 2016, 143, 3328–3339. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Yan, J.; Yu, X.; Liang, Y.; Fang, L.; Scheller, H.V.; Zhang, A. The NADPH-Oxidase AtRbohI Plays a Positive Role in Drought-Stress Response in Arabidopsis Thaliana. Biochem. Biophys. Res. Commun. 2017, 491, 834–839. [Google Scholar] [CrossRef]
- Brentner, L.B.; Mukherji, S.T.; Merchie, K.M.; Yoon, J.M.; Schnoor, J.L.; Van Aken, B. Expression of Glutathione S-Transferases in Poplar Trees (Populus Trichocarpa) Exposed to 2, 4, 6-Trinitrotoluene (TNT). Chemosphere 2008, 73, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Mezzari, M.P.; Walters, K.; Jelínkova, M.; Shih, M.-C.; Just, C.L.; Schnoor, J.L. Gene Expression and Microscopic Analysis of Arabidopsis Exposed to Chloroacetanilide Herbicides and Explosive Compounds. A Phytoremediation Approach. Plant Physiol. 2005, 138, 858–869. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Yi, H. Differential Expression of Arabidopsis Defense-Related Genes in Response to Sulfur Dioxide. Chemosphere 2012, 87, 718–724. [Google Scholar] [CrossRef]
- Cicero, L.L.; Madesis, P.; Tsaftaris, A.; Piero, A.R. Lo Tobacco Plants Over-Expressing the Sweet Orange Tau Glutathione Transferases (CsGSTUs) Acquire Tolerance to the Diphenyl Ether Herbicide Fluorodifen and to Salt and Drought Stresses. Phytochemistry 2015, 116, 69–77. [Google Scholar] [CrossRef]
- Yang, G.; Xu, Z.; Peng, S.; Sun, Y.; Jia, C.; Zhai, M. In Planta Characterization of a Tau Class Glutathione S-Transferase Gene from Juglans Regia (JrGSTTau1) Involved in Chilling Tolerance. Plant Cell Rep. 2016, 35, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Dixon, D.P.; McEwen, A.G.; Lapthorn, A.J.; Edwards, R. Forced Evolution of a Herbicide Detoxifying Glutathione Transferase. J. Biol. Chem. 2003, 278, 23930–23935. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Sahoo, A.; Devendran, R.; Jain, M. Over-Expression of a Rice Tau Class Glutathione s-Transferase Gene Improves Tolerance to Salinity and Oxidative Stresses in Arabidopsis. PLoS ONE 2014, 9, e92900. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, N.; Liu, Z.; Liu, S.; Liu, C.; Lin, J.; Yang, H.; Li, S.; Yukawa, Y. The AtGSTU7 Gene Influences Glutathione-Dependent Seed Germination under ABA and Osmotic Stress in Arabidopsis. Biochem. Biophys. Res. Commun. 2020, 528, 538–544. [Google Scholar] [CrossRef]
- Sarry, J.-E.; Kuhn, L.; Ducruix, C.; Lafaye, A.; Junot, C.; Hugouvieux, V.; Jourdain, A.; Bastien, O.; Fievet, J.B.; Vailhen, D.; et al. The Early Responses of Arabidopsis Thaliana Cells to Cadmium Exposure Explored by Protein and Metabolite Profiling Analyses. Proteomics 2006, 6, 2180–2198. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Ramaswamy, N.K.; Suprasanna, P.; D’souza, S.F. Genome-Wide Analysis of Thiourea-Modulated Salinity Stress-Responsive Transcripts in Seeds of Brassica Juncea: Identification of Signalling and Effector Components of Stress Tolerance. Ann. Bot. 2010, 106, 663–674. [Google Scholar] [CrossRef]
- Chan, C.; Lam, H.-M. A Putative Lambda Class Glutathione S-Transferase Enhances Plant Survival under Salinity Stress. Plant Cell Physiol. 2014, 55, 570–579. [Google Scholar] [CrossRef]
- Mengiste, T.; Chen, X.; Salmeron, J.; Dietrich, R. The Botrytis Susceptible1 Gene Encodes an R2R3MYB Transcription Factor Protein That Is Required for Biotic and Abiotic Stress Responses in Arabidopsis. Plant Cell 2003, 15, 2551–2565. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Liu, N.; Virlouvet, L.; Riethoven, J.-J.; Fromm, M.; Avramova, Z. Four Distinct Types of Dehydration Stress Memory Genes in Arabidopsis Thaliana. BMC Plant Biol. 2013, 13, 229. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, R.; Ju, Q.; Li, W.; Tran, L.-S.P.; Xu, J. The R2R3-MYB Transcription Factor MYB49 Regulates Cadmium Accumulation. Plant Physiol. 2019, 180, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.-K.; Dubos, C.; Dodd, I.C.; Holroyd, G.H.; Hetherington, A.M.; Campbell, M.M. AtMYB61, an R2R3-MYB Transcription Factor Controlling Stomatal Aperture in Arabidopsis Thaliana. Curr. Biol. 2005, 15, 1201–1206. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Andreasson, E.; Maslak, A.; Mock, H.P.; Mattsson, O.; Mundy, J. Arabidopsis MYB68 in Development and Responses to Environmental Cues. Plant Sci. 2004, 167, 1099–1107. [Google Scholar] [CrossRef]
- Lotkowska, M.E.; Tohge, T.; Fernie, A.R.; Xue, G.-P.; Balazadeh, S.; Mueller-Roeber, B. The Arabidopsis Transcription Factor MYB112 Promotes Anthocyanin Formation during Salinity and under High Light Stress. Plant Physiol. 2015, 169, 1862–1880. [Google Scholar] [CrossRef]
- Liu, S.; Yang, R.; Liu, M.; Zhang, S.; Yan, K.; Yang, G.; Huang, J.; Zheng, C.; Wu, C. AtPLATZ2 Negatively Regulates Salt Tolerance in Arabidopsis Seedlings by Directly Suppressing the Expression of the CBL4/SOS3 and CBL10/SCaBP8 Genes. J. Exp. Bot. 2020, 71, 5589–5602. [Google Scholar] [CrossRef]
- Horiuchi, J.; Badri, D.V.; Kimball, B.A.; Negre, F.; Dudareva, N.; Paschke, M.W.; Vivanco, J.M. The Floral Volatile, Methyl Benzoate, from Snapdragon (Antirrhinum Majus) Triggers Phytotoxic Effects in Arabidopsis Thaliana. Planta 2007, 226, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ueda, H.; Ito, T.; Inoue, R.; Masuda, Y.; Nagashima, Y.; Kozuka, T.; Kusaba, M. Genetic Interaction Among Phytochrome, Ethylene and Abscisic Acid Signaling During Dark-Induced Senescence in Arabidopsis Thaliana. Front. Plant Sci. 2020, 11, 564. [Google Scholar] [CrossRef]
- Zang, D.; Wang, J.; Zhang, X.; Liu, Z.; Wang, Y. Arabidopsis Heat Shock Transcription Factor HSFA7b Positively Mediates Salt Stress Tolerance by Binding to an E-Box-like Motif to Regulate Gene Expression. J. Exp. Bot. 2019, 70, 5355–5374. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Song, Y.; Li, S.; Zhang, L.; Zou, C.; Yu, D. The Role of WRKY Transcription Factors in Plant Abiotic Stresses. Biochim. Et Biophys. Acta (Bba)-Gene Regul. Mech. 2012, 1819, 120–128. [Google Scholar] [CrossRef]
- Song, Y.; Chen, L.; Zhang, L.; Yu, D. Overexpression of OsWRKY72 Gene Interferes in the Abscisic Acid Signal and Auxin Transport Pathway of Arabidopsis. J. Biosci. 2010, 35, 459–471. [Google Scholar] [CrossRef]
- Zhang, J.; Peng, Y.; Guo, Z. Constitutive Expression of Pathogen-Inducible OsWRKY31 Enhances Disease Resistance and Affects Root Growth and Auxin Response in Transgenic Rice Plants. Cell Res. 2008, 18, 508–521. [Google Scholar] [CrossRef]
- Guo, P.; Li, Z.; Huang, P.; Li, B.; Fang, S.; Chu, J.; Guo, H. A Tripartite Amplification Loop Involving the Transcription Factor WRKY75, Salicylic Acid, and Reactive Oxygen Species Accelerates Leaf Senescence. Plant Cell 2017, 29, 2854–2870. [Google Scholar] [CrossRef]
- Yang, Y.; Liang, T.; Zhang, L.; Shao, K.; Gu, X.; Shang, R.; Shi, N.; Li, X.; Zhang, P.; Liu, H. UVR8 Interacts with WRKY36 to Regulate HY5 Transcription and Hypocotyl Elongation in Arabidopsis. Nat. Plants 2018, 4, 98–107. [Google Scholar] [CrossRef]
- Zhang, L.; Qin, L.-N.; Zeng, Z.-R.; Wu, C.-Z.; Gong, Y.-Y.; Liu, L.-H.; Cao, F.-Q. Molecular Identification of a Root Apical Cell-Specific and Stress-Responsive Enhancer from an Arabidopsis Enhancer Trap Line. Plant Methods 2019, 15, 1–11. [Google Scholar] [CrossRef]
- Qiu, Y.; Yu, D. Over-Expression of the Stress-Induced OsWRKY45 Enhances Disease Resistance and Drought Tolerance in Arabidopsis. Environ. Exp. Bot. 2009, 65, 35–47. [Google Scholar] [CrossRef]
- Coego, A.; Brizuela, E.; Castillejo, P.; Ru\’\iz, S.; Koncz, C.; del Pozo, J.C.; Pineiro, M.; Jarillo, J.A.; Paz-Ares, J.; León, J.; et al. The TRANSPLANTA Collection of A Rabidopsis Lines: A Resource for Functional Analysis of Transcription Factors Based on Their Conditional Overexpression. Plant J. 2014, 77, 944–953. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Hong, S.H.; Kim, Y.W.; Lee, I.H.; Jun, J.H.; Phee, B.-K.; Rupak, T.; Jeong, H.; Lee, Y.; Hong, B.S.; et al. Gene Regulatory Cascade of Senescence-Associated NAC Transcription Factors Activated by Ethylene-Insensitive2-Mediated Leaf Senescence Signalling in Arabidopsis. J. Exp. Bot. 2014, 65, 4023–4036. [Google Scholar] [CrossRef] [PubMed]
- Joseph, M.P.; Papdi, C.; Kozma-Bognár, L.; Nagy, I.; López-Carbonell, M.; Rigó, G.; Koncz, C.; Szabados, L. The Arabidopsis ZINC FINGER PROTEIN3 Interferes with Abscisic Acid and Light Signaling in Seed Germination and Plant Development. Plant Physiol. 2014, 165, 1203–1220. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Ohyama, Y.; Kobayashi, Y.; Ito, H.; Iuchi, S.; Fujita, M.; Zhao, C.-R.; Tanveer, T.; Ganesan, M.; Kobayashi, M.; et al. STOP2 Activates Transcription of Several Genes for Al-and Low PH-Tolerance That Are Regulated by STOP1 in Arabidopsis. Mol. Plant 2014, 7, 311–322. [Google Scholar] [CrossRef]
- Huang, L.; Jiang, Q.; Wu, J.; An, L.; Zhou, Z.; Wong, C.; Wu, M.; Yu, H.; Gan, Y. Zinc Finger Protein 5 (ZFP5) Associates with Ethylene Signaling to Regulate the Phosphate and Potassium Deficiency-Induced Root Hair Development in Arabidopsis. Plant Mol. Biol. 2020, 102, 143–158. [Google Scholar] [CrossRef]
- Vergnolle, C.; Vaultier, M.-N.; Taconnat, L.; Renou, J.-P.; Kader, J.-C.; Zachowski, A.; Ruelland, E. The Cold-Induced Early Activation of Phospholipase C and D Pathways Determines the Response of Two Distinct Clusters of Genes in Arabidopsis Cell Suspensions. Plant Physiol. 2005, 139, 1217–1233. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Chung, M.-S.; Ju, H.-W.; Na, H.-S.; Lee, D.J.; Cheong, H.-S.; Kim, C.S. Physiological Characterization of the Arabidopsis Thaliana Oxidation-Related Zinc Finger 1, a Plasma Membrane Protein Involved in Oxidative Stress. J. Plant Res. 2011, 124, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.-F.; Tsai, M.-Y.; Lu, C.-A.; Wu, S.-J.; Yeh, C.-H. The Roles of Arabidopsis HSFA2, HSFA4a, and HSFA7a in the Heat Shock Response and Cytosolic Protein Response. Bot. Stud. 2018, 59, 15. [Google Scholar] [CrossRef]
- Lariguet, P.; Ranocha, P.; De Meyer, M.; Barbier, O.; Penel, C.; Dunand, C. Identification of a Hydrogen Peroxide Signalling Pathway in the Control of Light-Dependent Germination in Arabidopsis. Planta 2013, 238, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.-G.; Shan, J.-X.; Shi, M.; Gao, J.-P.; Lin, H.-X. DCA1 Acts as a Transcriptional Co-Activator of DST and Contributes to Drought and Salt Tolerance in Rice. Plos Genet 2015, 11, e1005617. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Sun, Y.; Zhao, R.; Shan, Z.; Gai, J.; Li, Y. Overexpression of Peroxidase Gene GsPRX9 Confers Salt Tolerance in Soybean. Int. J. Mol. Sci. 2019, 20, 3745. [Google Scholar] [CrossRef]
- Kumar, S.; Jaggi, M.; Sinha, A.K. Ectopic Overexpression of Vacuolar and Apoplastic Catharanthus Roseus Peroxidases Confers Differential Tolerance to Salt and Dehydration Stress in Transgenic Tobacco. Protoplasma 2012, 249, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Gullner, G.; Komives, T.; Király, L.; Schröder, P. Glutathione S-Transferase Enzymes in Plant-Pathogen Interactions. Front. Plant Sci. 2018, 9, 1836. [Google Scholar] [CrossRef] [PubMed]
- Ezaki, B.; Suzuki, M.; Motoda, H.; Kawamura, M.; Nakashima, S.; Matsumoto, H. Mechanism of Gene Expression of Arabidopsis Glutathione S-Transferase, AtGST1, and AtGST11 in Response to Aluminum Stress. Plant Physiol. 2004, 134, 1672–1682. [Google Scholar] [CrossRef]
- Roxas, V.P.; Lodhi, S.A.; Garrett, D.K.; Mahan, J.R.; Allen, R.D. Stress Tolerance in Transgenic Tobacco Seedlings That Overexpress Glutathione S-Transferase/Glutathione Peroxidase. Plant Cell Physiol. 2000, 41, 1229–1234. [Google Scholar] [CrossRef] [PubMed]
- Kunieda, T.; Fujiwara, T.; Amano, T.; Shioi, Y. Molecular Cloning and Characterization of a Senescence-Induced Tau-Class Glutathione S-Transferase from Barley Leaves. Plant Cell Physiol. 2005, 46, 1540–1548. [Google Scholar] [CrossRef]
- Yousefirad, S.; Soltanloo, H.; Ramezanpour, S.S.; Zaynali Nezhad, K.; Shariati, V. The RNA-Seq Transcriptomic Analysis Reveals Genes Mediating Salt Tolerance through Rapid Triggering of Ion Transporters in a Mutant Barley. PLoS ONE 2020, 15, e0229513. [Google Scholar] [CrossRef]
- Passardi, F.; Cosio, C.; Penel, C.; Dunand, C. Peroxidases Have More Functions than a Swiss Army Knife. Plant Cell Rep. 2005, 24, 255–265. [Google Scholar] [CrossRef]
- Yan, J.; Su, P.; Li, W.; Xiao, G.; Zhao, Y.; Ma, X.; Wang, H.; Nevo, E.; Kong, L. Genome-Wide and Evolutionary Analysis of the Class III Peroxidase Gene Family in Wheat and Aegilops Tauschii Reveals That Some Members Are Involved in Stress Responses. Bmc Genom. 2019, 20, 666. [Google Scholar] [CrossRef] [PubMed]
- Liszkay, A.; Kenk, B.; Schopfer, P. Evidence for the Involvement of Cell Wall Peroxidase in the Generation of Hydroxyl Radicals Mediating Extension Growth. Planta 2003, 217, 658–667. [Google Scholar] [CrossRef] [PubMed]
- Passardi, F.; Penel, C.; Dunand, C. Performing the Paradoxical: How Plant Peroxidases Modify the Cell Wall. Trends Plant Sci. 2004, 9, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 1 February 2021).
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-Based Genome Alignment and Genotyping with HISAT2 and HISAT-Genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Kovaka, S.; Zimin, A.V.; Pertea, G.M.; Razaghi, R.; Salzberg, S.L.; Pertea, M. Transcriptome Assembly from Long-Read RNA-Seq Alignments with StringTie2. Genome Biol. 2019, 20, 1–13. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Akers, M.D. Exploring, Analysing and Interpeting Data with Minitab 18; Compass Publishing: Seoul, Korea, 2018. [Google Scholar]
- Das, K.; Roychoudhury, A. Reactive Oxygen Species (ROS) and Response of Antioxidants as ROS-Scavengers during Environmental Stress in Plants. Front. Environ. Sci. 2014, 2, 1–13. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function | Gene ID | Gene Name | log2Fold Change | Previously Reported | References |
|---|---|---|---|---|---|
| H2O2 detoxification | Peroxidases | ||||
| Class III peroxidases | |||||
| AT1G05260 | AtPrx3 | 1.93 | Cold inducible tolerance, Stamen abscission | [44,45] | |
| AT1G14550 | AtPrx5 | 3.50 | |||
| AT1G30870 | AtPrx7 | 5.33 | TNT treatment | [46] | |
| AT1G49570 | AtPrx10 | 3.90 | |||
| AT1G68850 | AtPrx11 | 1.88 | Cuticle metabolism regulation in response to abiotic stress | [47] | |
| AT2G18980 | AtPrx16 | 2.75 | |||
| AT2G37130 | AtPrx21 | 1.43 | Stamen abscission, aluminum stress | [45,48] | |
| AT2G38380 | AtPrx22 | 2.02 | potassium deficiency | [49] | |
| AT2G38390 | AtPrx23 | 2.52 | |||
| AT2G39040 | AtPrx24 | 3.42 | |||
| AT3G01190 | AtPrx27 | 4.11 | Aluminum stress, TNT treatment | [46,48] | |
| AT3G03670 | AtPrx28 | 2.87 | |||
| AT3G21770 | AtPrx30 | 1.42 | Cell elongation, Stamen abscission, Monolignin polymerization | [45,50,51] | |
| AT3G32980 | AtPrx32 | 1.69 | Cell elongation | [50] | |
| AT4G26010 | AtPrx44 | 1.65 | |||
| AT4G30170 | AtPrx45 | 2.35 | Cell elongation, aluminum stress, TNT treatment, Stamen abscission | [45,48,50,52] | |
| AT4G37520 | AtPrx50 | 1.34 | Low oxygen response, phosphate starvation, Stamen abscission | [45,53,54] | |
| AT5G06730 | AtPrx54 | 2.07 | |||
| AT5G14130 | AtPrx55 | 2.82 | |||
| AT5G15180 | AtPrx56 | 1.42 | Aluminum stress | [48] | |
| AT5G17820 | AtPrx57 | 4.38 | Arsenic stress, TNT treatment, cell elongation | [46,50,55] | |
| AT5G19890 | AtPrx59 | 4.22 | Aluminum stress, Mechanical stimulus | [48,51] | |
| AT5G24070 | AtPrx61 | 3.46 | |||
| AT5G64100 | AtPrx69 | 2.67 | Phosphate starvation, sulphur deficiency | [52,53] | |
| AT5G66390 | AtPrx72 | 1.33 | Cell elongation | [50] | |
| AT5G67400 | AtPrx73 | 2.56 | Aluminum stress | [48] | |
| H2O2 detoxification and signaling | Other peroxidases | ||||
| AT1G60740 | Trx | 4.66 | |||
| AT5G07390 | RbohA | 2.94 | Lateral root emergence, salinity and cold stress | [54,55] | |
| AT1G09090 | RbohB | 3.26 | Nitrogen fixation, lateral root emergence | [56,57] | |
| AT5G51060 | RbohC | 1.99 | Lateral root emergence, salinity and cold stress | [58,59] | |
| AT4G25090 | RbohG | 2.96 | Lateral root emergence | [60] | |
| AT4G11230 | RbohI | 1.12 | Drought stress | [61] | |
| AT3G01420 | Dox1 | 2.93 | |||
| AT1G65970 | Tpx2 | 1.27 | |||
| H2O2 detoxification | Glutathione S-transferase | ||||
| AT2G29490 | GSTU1 | 2.79 | Herbicide treatment, phytoremediation, oxidative stress response (SO2), salinity, drought and cold stress | [62,63,64,65,66] | |
| AT2G29480 | GSTU2 | 2.63 | Herbicide treatment, salinity and drought stress | [65,67] | |
| AT2G29470 | GSTU3 | 2.64 | Oxidative stress response (SO2) | [64] | |
| AT2G29460 | GSTU4 | 1.76 | Oxidative stress response (SO2), salinity | [64,68] | |
| AT2G29420 | GSTU7 | 1.37 | Seed germination, ABA response and osmotic stress | [69] | |
| AT3G09270 | GSTU8 | 1.40 | Cadmium treatment | [70] | |
| AT1G69920 | GSTU12 | 1.68 | Salinity stress | [71] | |
| AT1G27140 | GSTU14 | 4.32 | |||
| AT1G78340 | GSTU22 | 2.89 | |||
| AT1G17170 | GSTU24 | 1.61 | TNT treatment, herbicide treatment, phytoremediation, oxidative stress response (SO2) | [62,63,64] | |
| AT5G02780 | GSTL1 | 1.27 | Increased tolerance to salinity stress | [72] | |
| Gene ID | Gene Name | log2Fold Change | Previously Reported | References |
|---|---|---|---|---|
| MYB containing domain | ||||
| AT5G49620 | MYB 78 | 4.44 | Abiotic and biotic stress | [73] |
| AT1G74080 | MYB122 | 3.18 | Dehydration stress | [74] |
| AT1G79180 | MYB63 | 2.52 | Dehydration stress | [74] |
| AT5G54230 | MYB49 | 2.39 | Cadmium accumulation | [75] |
| AT1G09540 | MYB61 | 2.17 | Stomatal aperture | [76] |
| AT5G65790 | MYB68 | 1.85 | High temperature | [77] |
| AT1G48000 | MYB112 | 1.75 | Salinity and high light stress | [78] |
| AT4G34990 | MYB32 | 1.35 | Salinity stress | [79] |
| AT3G49690 | MYB84 | 1.27 | High temperature | [77] |
| Basic helix-loop-helix DNA binding superfamily protein | ||||
| AT4G21340 | bHLH | 4.04 | Response to phytotoxicity | [80] |
| AT1G02340 | bHLH | 2.57 | Dark induced senescence | [81] |
| AT4G29930 | bHLH | 1.67 | Dehydration stress | [74] |
| AT1G10585 | bHLH | 1.46 | Dehydration stress | [74] |
| AT5G51780 | bHLH | 1.11 | Salinity stress | [82] |
| WRKY DNA binding protein | ||||
| AT1G68150 | AtWRKY09 | 3.97 | Abiotic stresses | [83] |
| AT5G15130 | AtWRKY72 | 2.90 | Abscisic acid signal | [84] |
| AT4G22070 | AtWRKY31 | 2.70 | Root growth, pathogen attack | [85] |
| AT5G13080 | AtWRKY75 | 2.65 | Leaf senescence | [86] |
| AT1G69810 | AtWRKY36 | 1.61 | UV responsive | [87] |
| AT1G30650 | AtWRKY14 | 1.31 | Abiotic stresses | [88] |
| AT3G01970 | AtWRKY45 | 1.16 | Dehydration stress tolerance | [89] |
| NAC containing domain | ||||
| AT3G18400 | ANAC058 | 1.91 | ABA mediated germination | [90] |
| AT1G01010 | ANAC001 | 1.67 | Dehydration stress | [74] |
| AT3G29035 | ANAC003 | 1.34 | Leaf senescence | [91] |
| Zinc finger protein | ||||
| AT1G67030 | AtZFP67 | 3.98 | ABA repressor | [92] |
| AT5G22890 | AtSTOP2 (C2HC ZFP) | 3.56 | Aluminum and low pH | [93] |
| AT5G57520 | AtZFP2 | 3.51 | Salinity stress | [82] |
| AT1G10480 | AtZFP5 | 3.31 | Phosphate and potassium deficiency | [94] |
| AT1G68360 | AtGIS3 (C2HC ZFP) | 1.59 | Cold stress | [95] |
| AT2G28200 | C2H2 ZFP | 1.12 | Dehydration stress | [74] |
| AT2G19810 | AtOZF1(CCCH ZFP) | 1.08 | Hydrogen peroxide, abscisic acid and salinity responsive | [96] |
| Heat shock family protein | ||||
| AT3G51910 | AtHSFA7A 2 | 2.12 | Heat shock response | [97] |
| AT2G26150 | AtHSFA2 | 1.94 | Heat shock response | [97] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.T.; Ckurshumova, W.; Fefer, M.; Liu, J.; Uddin, W.; Rosa, C. A Plant Based Modified Biostimulant (Copper Chlorophyllin), Mediates Defense Response in Arabidopsis thaliana under Salinity Stress. Plants 2021, 10, 625. https://doi.org/10.3390/plants10040625
Islam MT, Ckurshumova W, Fefer M, Liu J, Uddin W, Rosa C. A Plant Based Modified Biostimulant (Copper Chlorophyllin), Mediates Defense Response in Arabidopsis thaliana under Salinity Stress. Plants. 2021; 10(4):625. https://doi.org/10.3390/plants10040625
Chicago/Turabian StyleIslam, Md Tariqul, Wenzi Ckurshumova, Michael Fefer, Jun Liu, Wakar Uddin, and Cristina Rosa. 2021. "A Plant Based Modified Biostimulant (Copper Chlorophyllin), Mediates Defense Response in Arabidopsis thaliana under Salinity Stress" Plants 10, no. 4: 625. https://doi.org/10.3390/plants10040625
APA StyleIslam, M. T., Ckurshumova, W., Fefer, M., Liu, J., Uddin, W., & Rosa, C. (2021). A Plant Based Modified Biostimulant (Copper Chlorophyllin), Mediates Defense Response in Arabidopsis thaliana under Salinity Stress. Plants, 10(4), 625. https://doi.org/10.3390/plants10040625