
Characterization of the Genetic Diversity Present in a Diverse Sesame Landrace Collection Based on Phenotypic Traits and EST-SSR Markers Coupled With an HRM Analysis

 , , ,
, , , 

Abstract
1. Introduction
2. Results
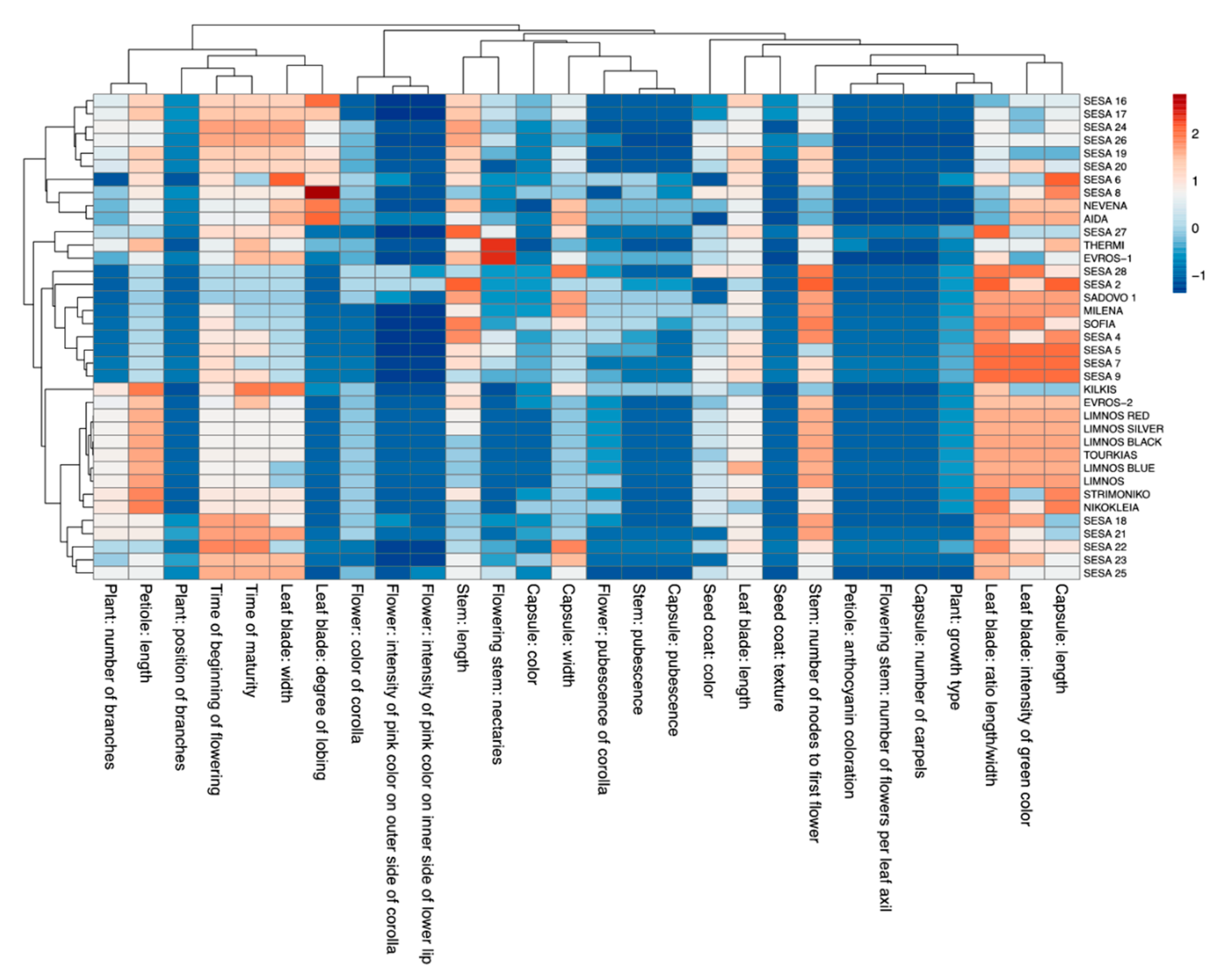
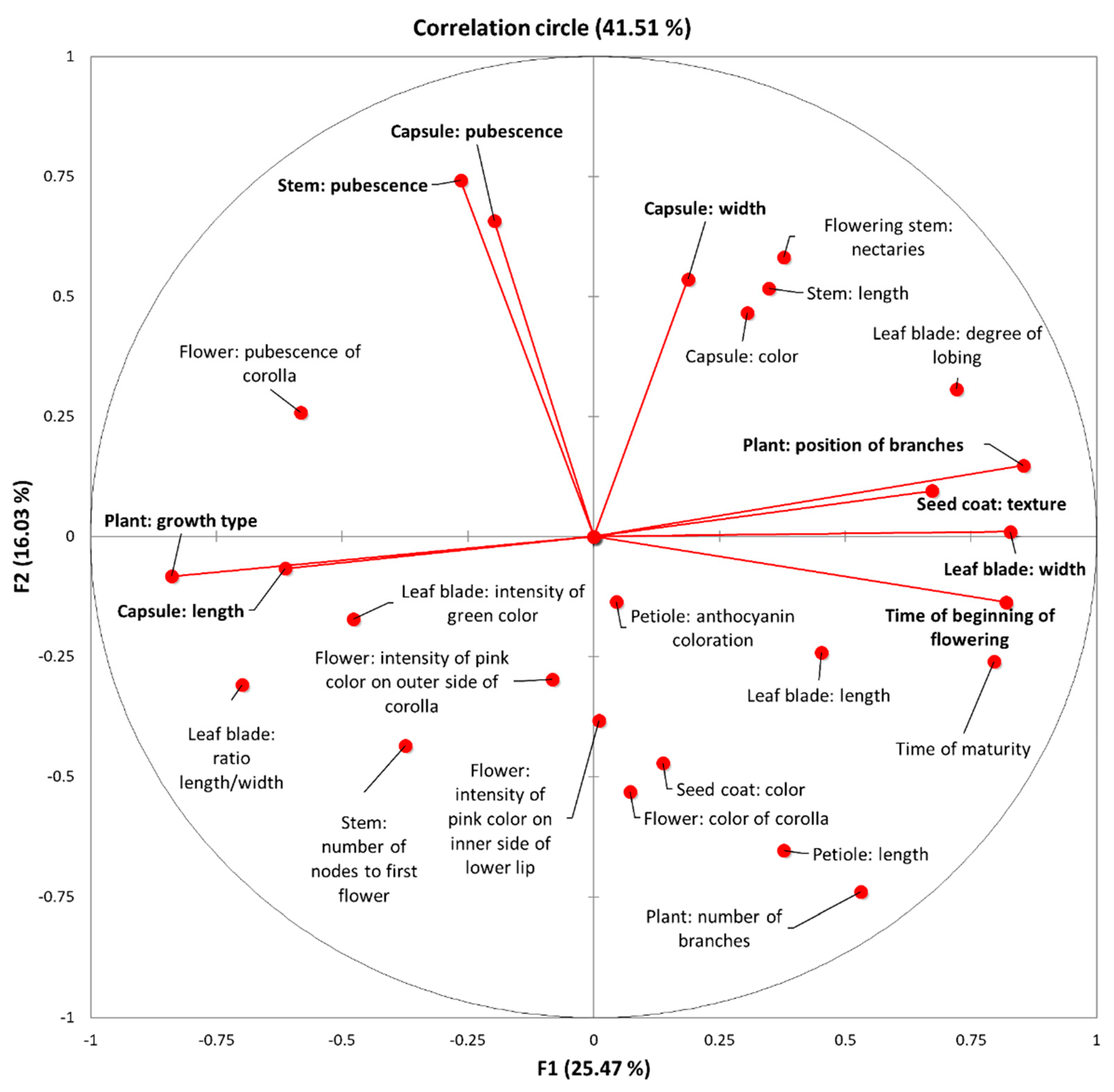
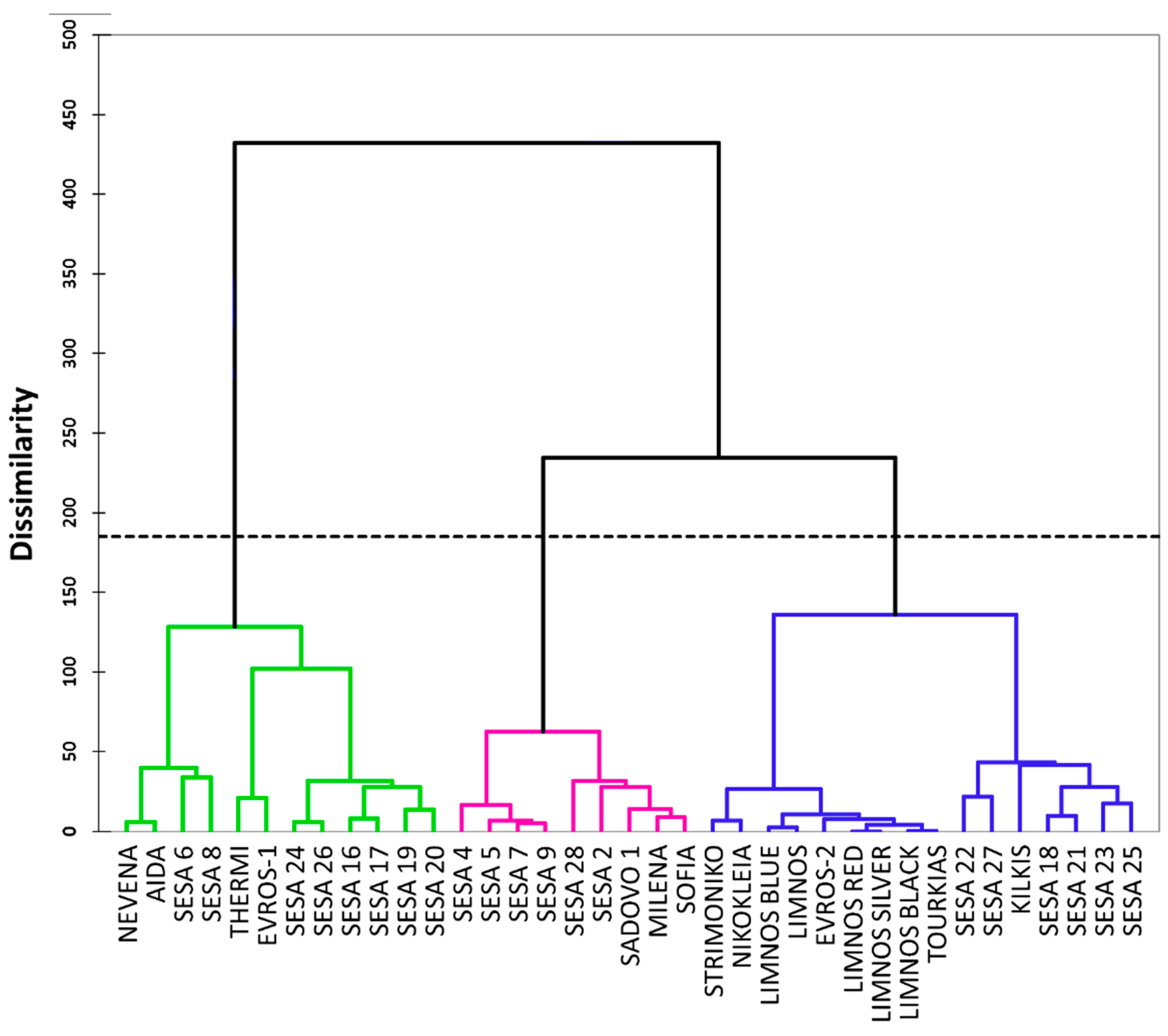
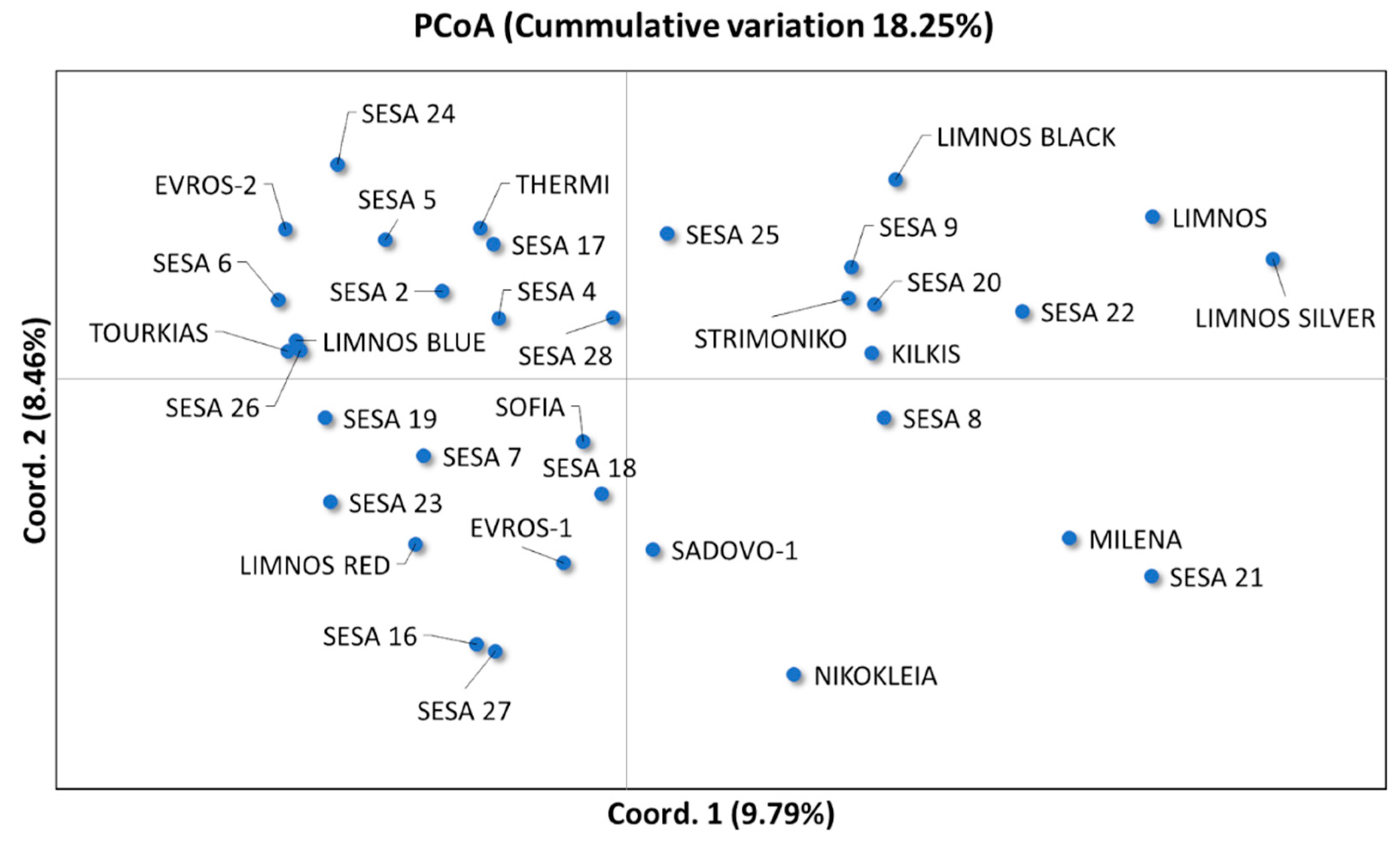
2.1. Morpho-Physiological and Agronomical Diversity of the Sesame Landraces
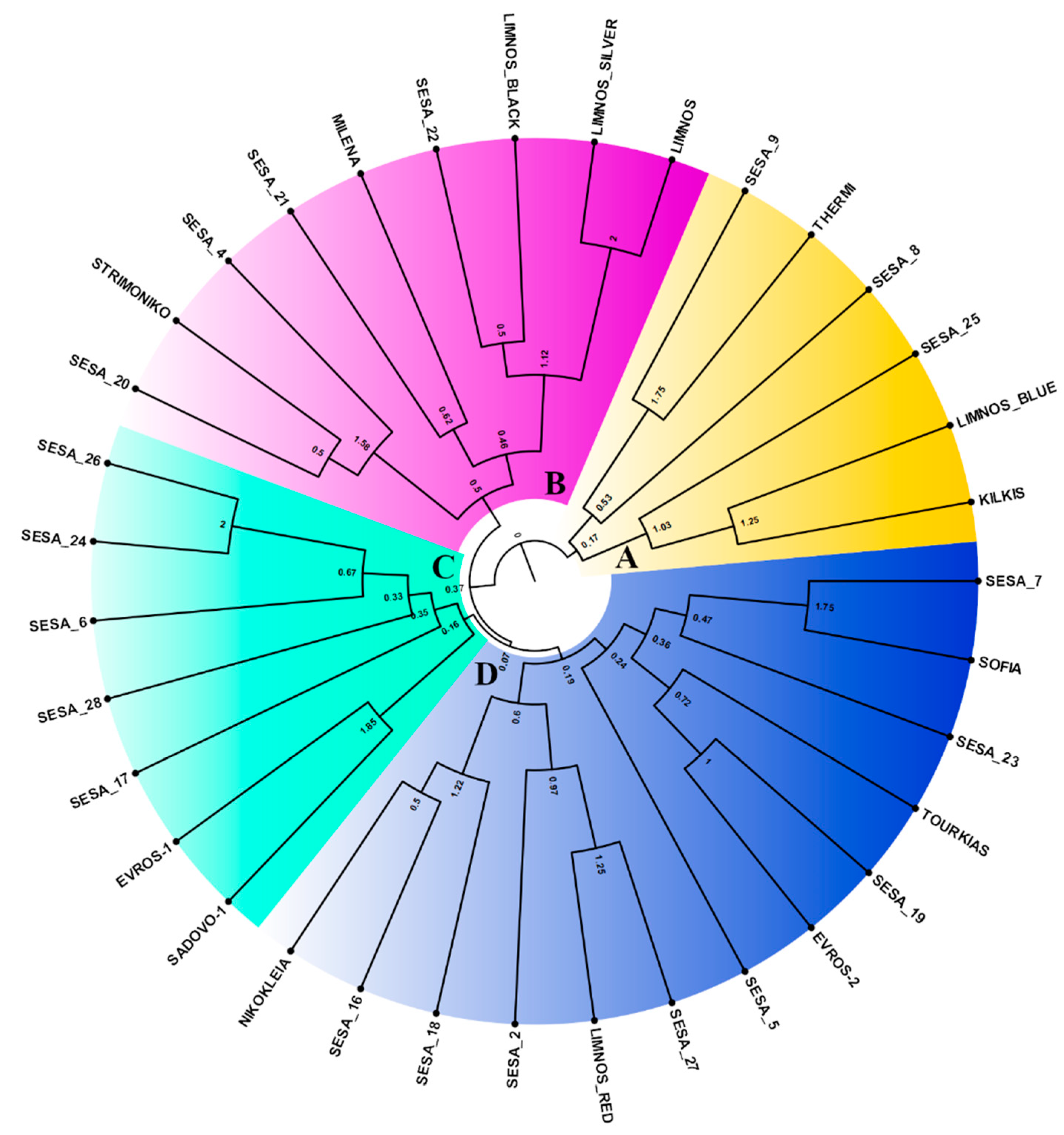
2.2. Genetic Diversity of the Sesame Landraces Based on the EST-SSR Marker Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Morpho-Physiological Analysis of Sesame Landraces
4.3. DNA Isolation, PCR Amplification and HRM Analysis
4.4. EST-SSR Genotyping and Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT Production—Crops. Available online: http://www.fao.org/faostat/en/#compare (accessed on 12 July 2020).
- Prasad, M.N.N.; Sanjay, K.R.; Prasad, S.D. A Review on Nutritional and Nutraceutical Properties of Sesame. J. Nutr. Food Sci. 2012, 2, 127. [Google Scholar] [CrossRef]
- Costa, F.T.; Neto, S.M.; Bloch, C.; Franco, O.L. Susceptibility of Human Pathogenic Bacteria to Antimicrobial Peptides from Sesame Kernels. Curr. Microbiol. 2007, 55, 162–166. [Google Scholar] [CrossRef]
- Anilakumar, K.R.; Pal, A.; Khanum, F.; Bawa, A.S. Nutritional, medicinal and industrial uses of sesame (Sesamum indicum L.) seeds—An overview. Agric. Conspec. Sci. 2010, 75, 159–168. [Google Scholar]
- Elleuch, M.; Besbes, S.; Roiseux, O.; Blecker, C.; Attia, H. Quality characteristics of sesame seeds and by-products. Food Chem. 2007, 103, 641–650. [Google Scholar] [CrossRef]
- Wei, X.; Zhu, X.; Yu, J.; Wang, L.; Zhang, Y.; Li, D.; Zhou, R.; Zhang, X. Identification of Sesame Genomic Variations from Genome Comparison of Landrace and Variety. Front. Plant. Sci. 2016, 7, 1169. [Google Scholar] [CrossRef]
- Dossa, K.; Diouf, D.; Wang, L.; Wei, X.; Zhang, Y.; Niang, M.; Fonceka, D.; Yu, J.; Mmadi, M.A.; Yehouessi, L.W.; et al. The emerging oilseed crop Sesamum indicum enters the “Omics” era. Front. Plant Sci. 2017, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Dossa, K.; Mmadi, M.A.; Zhou, R.; Zhang, T.; Su, R.; Zhang, Y.; Wang, L.; You, J.; Zhang, X. Depicting the Core Transcriptome Modulating Multiple Abiotic Stresses Responses in Sesame (Sesamum indicum L.). Int. J. Mol. Sci. 2019, 20, 3930. [Google Scholar] [CrossRef]
- Ganopoulos, I.; Mylona, P.; Mellidou, I.; Kalivas, A.; Bosmali, I.; Kontzidou, S.; Osathanunkul, M.; Madesis, P. Microsatellite genotyping and molecular screening of pea (Pisum sativum L.) germplasm with high-resolution melting analysis for resistance to powdery mildew. Plant. Gene 2018, 15, 1–5. [Google Scholar] [CrossRef]
- Furat, S.; Uzun, B. The use of agro-morphological characters for the assessment of genetic diversity in sesame (Sesamum indicum L.). Plant Omics 2010, 3, 85–91. [Google Scholar]
- Zhang, Y.; Zhang, X.; Che, Z.; Wang, L.; Wei, W.; Li, D. Genetic diversity assessment of sesame core collection in China by phenotype and molecular markers and extraction of a mini-core collection. BMC Genet. 2012, 13, 102. [Google Scholar] [CrossRef]
- Wei, X.; Liu, K.; Zhang, Y.; Feng, Q.; Wang, L.; Zhao, Y.; Li, D.; Zhao, Q.; Zhu, X.; Zhu, X.; et al. Genetic discovery for oil production and quality in sesame. Nat. Commun. 2015, 6, 8609. [Google Scholar] [CrossRef]
- Iqbal, A. Genetic estimates and diversity study in Sesame (Sesamum indicum L.). IOSR J. Agric. Veter. Sci. 2016, 9, 1–5. [Google Scholar] [CrossRef]
- Pérez, P.F.; López, J.F. Morphological and phenological description of 38 sweet chestnut cultivars (Castanea sativa Miller) in a contemporary collection. Span. J. Agric. Res. 2009, 7, 829. [Google Scholar] [CrossRef]
- De Oliveira, E.J.; Dias, N.L.P.; Dantas, J.L.L. Selection of morpho-agronomic descriptors for characterization of papaya cultivars. Euphytica 2011, 185, 253–265. [Google Scholar] [CrossRef]
- Mehmood, A.; Jaskani, M.J.; Khan, I.A.; Ahmad, S.; Ahmad, R.; Luo, S.; Ahmad, N.M. Genetic diversity of Pakistani guava (Psidium guajava L.) germplasm and its implications for conservation and breeding. Sci. Hortic. 2014, 172, 221–232. [Google Scholar] [CrossRef]
- Ganopoulos, I.; Tourvas, N.; Xanthopoulou, A.; Aravanopoulos, F.A.; Avramidou, E.; Zambounis, A.; Tsaftaris, A.; Madesis, P.; Sotiropoulos, T.; Koutinas, N. Phenotypic and molecular characterization of apple (Malus × domestica Borkh) genetic resources in Greece. Sci. Agricola 2018, 75, 509–518. [Google Scholar] [CrossRef]
- Ramprasad, E.; Senthilvel, S.; Jatoth, J.L.; Yamini, K.N.; Dangi, K.S.; Ranganatha, A.R.G.; Varaprasad, K.S. An insight into morphological and molecular diversity in Indian sesame cultivars. Indian J. Genet. Plant. Breed. 2017, 77, 271. [Google Scholar] [CrossRef]
- Bhattacharjee, M.; Iqbal, A.; Singha, S.; Nath, D.; Prakash, S.; Dasgupta, T. Genetic diversity in Sesamum indicum L. Bangladesh J. Bot. 2019, 48, 497–506. [Google Scholar] [CrossRef]
- Pandey, P.; Ramegowda, V.; Senthil-Kumar, M. Shared and unique responses of plants to multiple individual stresses and stress combinations: Physiological and molecular mechanisms. Front. Plant. Sci. 2015, 6, 723. [Google Scholar] [CrossRef]
- Bhat, K.V.; Babrekar, P.P.; Lakhanpaul, S. Study of genetic diversity in Indian and exotic sesame (Sesamum indicum L.) germplasm using random amplified polymorphic DNA (RAPD) markers. Euphytica 1999, 110, 21–34. [Google Scholar] [CrossRef]
- Ercan, A.G.; Taskin, K.M.; Turgut, K. Analysis of genetic diversity in Turkish sesame (Sesamum indicum L.) populations using RAPD markers. Genet. Resour. Crop. Evol. 2004, 51, 599–607. [Google Scholar] [CrossRef]
- Arriel, N.H.C.; Di Mauro, A.O.; Arriel, E.F.; Unêda-Trevisoli, S.H.; Costa, M.M.; Bárbaro, I.M.; Muniz, F.R.S. Genetic divergence in sesame based on morphological and agronomic traits. Crop. Breed. Appl. Biotechnol. 2007, 7, 253–261. [Google Scholar] [CrossRef]
- Salazar, B.; Laurentín, H.; Dávila, M.; Castillo, M.A. Reliability of the rapd technique for germplasm analysis of sesame (Sesamum indicum L) from Venezuela. Interciencia 2006, 31, 456–460. [Google Scholar]
- Abdellatef, E.; Sirelkhatem, R.; Mohamed Ahmed, M.M.; Radwan, K.H.; Khalafalla, M.M. Study of genetic diversity in Sudanese sesame (Sesamum indicum L.) germplasm using random amplified polymorphic DNA (RAPD) markers. Afr. J. Biotechnol. 2008, 7, 4423–4427. [Google Scholar]
- Uzun, B.; Lee, D.; Donini, P.; Çaǧirgan, M.L. Identification of a molecular marker linked to the closed capsule mutant trait in sesame using AFLP. Plant. Breed. 2003, 122, 95–97. [Google Scholar] [CrossRef]
- Laurentin, E.H.; Karlovsky, P. Genetic relationship and diversity in a sesame (Sesamum indicum L.) germplasm collection using amplified fragment length polymorphism (AFLP). BMC Genet. 2006, 7, 10. [Google Scholar] [CrossRef]
- Laurentin, H.; Karlovsky, P. AFLP fingerprinting of sesame (Sesamum indicum L.) cultivars: Identification, genetic relationship and comparison of AFLP informativeness parameters. Genet. Resour. Crop. Evol. 2007, 54, 1437–1446. [Google Scholar] [CrossRef]
- Kim, D.H.; Zur, G.; Danin-Poleg, Y.; Lee, S.W.; Shim, K.B.; Kang, C.W.; Kashi, Y. Genetic relationships of sesame germplasm collection as revealed by inter-simple sequence repeats. Plant. Breed. 2002, 121, 259–262. [Google Scholar] [CrossRef]
- Parsaeian, M.; Mirlohi, A.; Saeidi, G. Study of genetic variation in sesame (Sesamum indicum L.) using agro-morphological traits and ISSR markers. Russ. J. Genet. 2011, 47, 314–321. [Google Scholar] [CrossRef]
- Dixit, A.; Jin, M.-H.; Chung, J.-W.; Yu, J.-W.; Chung, H.-K.; Ma, K.-H.; Park, Y.-J.; Cho, E.-G. Development of polymorphic microsatellite markers in sesame (Sesamum indicum L.). Mol. Ecol. Notes 2005, 5, 736–738. [Google Scholar] [CrossRef]
- Sudhakara, V.; Kola, R.; Yepuri, V.; Surapaneni, M. Genetic Diversity and DNA Fingerprinting in Sesame (Sesamum indicum L.). Cultiv. Angrau. 2012, 6, 98–101. [Google Scholar]
- Uncu, A. Özgür; Gultekin, V.; Allmer, J.; Frary, A.; Doganlar, S. Genomic Simple Sequence Repeat Markers Reveal Patterns of Genetic Relatedness and Diversity in Sesame. Plant. Genome 2015, 8. [Google Scholar] [CrossRef] [PubMed]
- Dossa, K.; Wei, X.; Zhang, Y.; Fonceka, D.; Yang, W.; Diouf, D.; Liao, B.; Cisse, N.; Zhang, X. Analysis of Genetic Diversity and Population Structure of Sesame Accessions from Africa and Asia as Major Centers of Its Cultivation. Genes 2016, 7, 14. [Google Scholar] [CrossRef]
- Sehr, E.M.; Okello-Anyanga, W.; Hasel-Hohl, K.; Burg, A.; Gaubitzer, S.; Rubaihayo, P.R.; Okori, P.; Vollmann, J.; Gibson, P.; Fluch, S. Assessment of genetic diversity amongst Ugandan sesame (Sesamum indicum L.) landraces based on agromorphological traits and genetic markers. J. Crop. Sci. Biotechnol. 2016, 19, 117–124. [Google Scholar] [CrossRef]
- Suh, M.C.; Kim, M.J.; Hur, C.-G.; Bae, J.M.; Park, Y.I.; Chung, C.-H.; Kang, C.-W.; Ohlrogge, J.B. Comparative analysis of expressed sequence tags from Sesamum indicum and Arabidopsis thaliana developing seeds. Plant. Mol. Biol. 2003, 52, 1107–1123. [Google Scholar] [CrossRef]
- Wei, L.-B.; Zhang, H.-Y.; Zheng, Y.-Z.; Guo, W.-Z.; Zhang, T.-Z. Developing EST-Derived Microsatellites in Sesame (Sesamum indicum L.). Acta Agron. Sin. 2008, 34, 2077–2084. [Google Scholar] [CrossRef]
- Wei, W.; Qi, X.; Wang, L.; Zhang, Y.; Hua, W.; Li, D.; Lv, H.; Zhang, X. Characterization of the sesame (Sesamum indicum L.) global transcriptome using Illumina paired-end sequencing and development of EST-SSR markers. BMC Genom. 2011, 12, 451. [Google Scholar] [CrossRef]
- Ke, T.; Dong, C.; Mao, H.; Zhao, Y.; Chen, H.; Liu, H.; Dong, X.; Tong, C.; Liu, S. Analysis of expression sequence tags from a full-length-enriched cDNA library of developing sesame seeds (Sesamum indicum). BMC Plant. Biol. 2011, 11, 180. [Google Scholar] [CrossRef]
- Wu, K.; Yang, M.; Liu, H.; Tao, Y.; Mei, J.; Zhao, Y. Genetic analysis and molecular characterization of Chinese sesame (Sesamum indicum L.) cultivars using Insertion-Deletion (InDel) and Simple Sequence Repeat (SSR) markers. BMC Genet. 2014, 15, 35. [Google Scholar] [CrossRef] [PubMed]
- Badri, J.; Yepuri, V.; Ghanta, A.; Siva, S.; Siddiq, E.A. Development of microsatellite markers in sesame (Sesamum indicum L.). Turk. J. Agric. For. 2014, 38, 603–614. [Google Scholar] [CrossRef]
- Nandakumar, N.; Singh, A.; Sharma, R.; Mohapatra, T.; Prabhu, K.; Zaman, F. Molecular fingerprinting of hybrids and assessment of genetic purity of hybrid seeds in rice using microsatellite markers. Euphytica 2004, 136, 257–264. [Google Scholar] [CrossRef]
- Kaur, G.; Joshi, A.; Jain, D. SSR-Marker assisted evaluation of Genetic Diversity in Mungbean (Vigna radiata (L.) Wilcezk) genotypes. Braz. Arch. Biol. Technol. 2018, 61. [Google Scholar] [CrossRef]
- Zhang, Y.-X.; Zhang, X.-R.; Hua, W.; Wang, L.-H.; Che, Z. Analysis of genetic diversity among indigenous landraces from sesame (Sesamum indicum L.) core collection in China as revealed by SRAP and SSR markers. Genes Genom. 2010, 32, 207–215. [Google Scholar] [CrossRef]
- Thomson, M.J.; Septiningsih, E.M.; Suwardjo, F.; Santoso, T.J.; Silitonga, T.S.; McCouch, S.R. Genetic diversity analysis of traditional and improved Indonesian rice (Oryza sativa L.) germplasm using microsatellite markers. Theor. Appl. Genet. 2007, 114, 559–568. [Google Scholar] [CrossRef]
- Bhat, K.; Kumari, R.; Pathak, N.; Rai, A. Value addition in sesame: A perspective on bioactive components for enhancing utility and profitability. Pharmacogn. Rev. 2014, 8, 147–155. [Google Scholar] [CrossRef]
- UPOV International Union for the Protection of New Varieties of Plants (Sesame). 2013. Available online: https://www.upov.int/portal/index.html.en (accessed on 15 September 2018).
- Kaiser, H.F. The Application of Electronic Computers to Factor Analysis. Educ. Psychol. Meas. 1960, 20, 141–151. [Google Scholar] [CrossRef]
- Pandey, S.K.; Das, A.; Rai, P.; Dasgupta, T. Morphological and genetic diversity assessment of sesame (Sesamum indicum L.) accessions differing in origin. Physiol. Mol. Biol. Plants 2015, 21, 519–529. [Google Scholar] [CrossRef]
- Prasad, R.; Gangopadhyay, G. Phenomic analyses of Indian and exotic accessions of Sesame (Sesamum indicum L.). J. Plant Breed. Crop Sci. 2011, 3, 335–351. [Google Scholar]
- Jeyaraj, J.; Beevy, S.S. A Comparative Study on the Reproductive Success of Two Species of Sesamum L. (Pedaliaceae). Adv. Zool. Bot. 2020, 8, 144–153. [Google Scholar] [CrossRef]
- Tabatabaei, I.; Pazouki, L.; Bihamta, M.R.; Mansoori, S.; Javaran, M.J.; Niinemets, Ü. Genetic variation among iranian sesame (Sesamum indicum L.) accessions vis-à-vis exotic genotypes on the basis of morpho-physiological traits and RAPD markers. Aust. J. Crop Sci. 2011, 5, 1396–1407. [Google Scholar]
- Tahmasebi, A.; Ashrafi-Dehkordi, E.; Shahriari, A.G.; Mazloomi, S.M.; Ebrahimie, E. Integrative meta-analysis of transcriptomic responses to abiotic stress in cotton. Prog. Biophys. Mol. Biol. 2019, 146, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.-I.; Park, J.-H.; Lee, C.-W.; Ra, W.-H.; Chung, J.-W.; Lee, J.-R.; Ma, K.-H.; Lee, S.-Y.; Lee, K.-S.; Lee, M.-C.; et al. Evaluation of the genetic diversity and population structure of sesame (Sesamum indicum L.) using microsatellite markers. Genes Genom. 2011, 33, 187–195. [Google Scholar] [CrossRef]
- Dossa, K.; Li, D.; Wang, L.; Zheng, X.; Liu, A.; Yu, J.; Wei, X.; Zhou, R.; Foncéka, D.; Diouf, D.; et al. Transcriptomic, biochemical and physio-anatomical investigations shed more light on responses to drought stress in two contrasting sesame genotypes. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Olusola, T.O.; Opeyemi, A.O.; Oluwabukola, O.A.; Abiodun, S.; Omowumi, A.A.; Oyejide, S.O.; Oduoye, O.T.; Oluwasanya, O.A.; Arikawe, O.O.; Sunday, A.; et al. Genetic variation via SSR polymorphic information content and ecological distribution of Nigerian sesame. Afr. J. Biotechnol. 2020, 19, 165–170. [Google Scholar] [CrossRef]
- Woldesenbet, D.T.; Kassahun, T.; Endashaw, B. Genetic diversity of sesame germplasm collection (Sesamum indicum L.): Implication for conservation, improvement and use. Int. J. Biotechnol. Mol. Biol. Res. 2015, 6, 7–18. [Google Scholar]
- Pham, T.D.; Geleta, M.; Bui, T.M.; Bui, T.C.; Merker, A.; Carlsson, A.S. Comparative analysis of genetic diversity of sesame (Sesamum indicum L.) from Vietnam and Cambodia using agro-morphological and molecular markers. Hereditas 2011, 148, 28–35. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, 566–570. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemistry 1987, 19, 11–15. [Google Scholar]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research - an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | 2016 | 2017 | ||||
|---|---|---|---|---|---|---|
| Tmin (°C) | Tmax (°C) | Rainfall (mm) | Tmin (°C) | Tmax (°C) | Rainfall (mm) | |
| May | 9 | 32 | 75.2 | 11 | 30 | 5.8 |
| June | 14 | 38 | 13.3 | 14 | 39 | 13.2 |
| July | 17 | 36 | 0.7 | 16 | 40 | 78.3 |
| August | 18 | 36 | 56.6 | 14 | 38 | 12.9 |
| September | 9 | 31 | 80 | 12 | 35 | 2.7 |
| Mean | 13.4 | 34.6 | 13.4 | 36.4 | - | |
| Total | - | - | 225.8 | - | - | 158.9 |
| F1 | F2 | F3 | F4 | F5 | F6 | F7 | F8 | |
|---|---|---|---|---|---|---|---|---|
| Eigenvalue | 7.13 | 4.49 | 3.87 | 1.82 | 1.39 | 1.19 | 1.14 | 1.04 |
| Variability (%) | 25.48 | 16.03 | 13.83 | 6.48 | 4.98 | 4.26 | 4.08 | 3.71 |
| Cumulative (%) | 25.48 | 41.51 | 55.34 | 61.82 | 66.8 | 71.06 | 75.14 | 78.85 |
| Locus | I | h | uh | PIC | HRM Profiles |
|---|---|---|---|---|---|
| ZM_2 | 0.315 ± 0.053 | 0.181 ± 0.039 | 0.186 ± 0.041 | 0.87 | 7 |
| ZM_10 | 0.305 ± 0.09 | 0.186 ± 0.069 | 0.192 ± 0.072 | 0.65 | 10 |
| ZM_11 | 0.276 ± 0.046 | 0.154 ± 0.033 | 0.158 ± 0.034 | 0.85 | 11 |
| ZM_21 | 0.29 ± 0.052 | 0.168 ± 0.037 | 0.17 ± 0.038 | 0.84 | 8 |
| ZM_22 | 0.345 ± 0.072 | 0.21 ± 0.06 | 0.21 ± 0.06 | 0.78 | 8 |
| ZM_34 | 0.339 ± 0.05 | 0.197 ± 0.038 | 0.203 ± 0.039 | 0.85 | 7 |
| ZM_47 | 0.268 ± 0.045 | 0.148 ± 0.032 | 0.153 ± 0.033 | 0.89 | 11 |
| Landraces | Country of Origin | Landraces | Country of Origin |
|---|---|---|---|
| LIMNOS SILVER | Greece (GR) | SESA 2 | Iraq (IRQ) |
| LIMNOS BLUE | Greece (GR) | SESA 5 | Iraq (IRQ) |
| LIMNOS RED | Greece (GR) | SESA 9 | Iraq (IRQ) |
| LIMNOS BLACK | Greece (GR) | SESA 4 | Korea (PRK) |
| LIMNOS | Greece (GR) | SESA 6 | Korea (PRK) |
| EVROS-1 | Greece (GR) | SESA 7 | Tajikistan (TJK) |
| EVROS-2 | Greece (GR) | SESA 8 | Nepal (NPL) |
| KILKIS | Greece (GR) | SESA 16 | Yemen (YEM) |
| THERMI | Greece (GR) | SESA 17 | Yemen (YEM) |
| STRIMONIKO | Greece (GR) | SESA 18 | Yemen (YEM) |
| NIKOKLEIA | Cyprus (CY) | SESA 19 | Yemen (YEM) |
| SADOVO-1 | Bulgaria (BG) | SESA 20 | Yemen (YEM) |
| SOFIA | Bulgaria (BG) | SESA 21 | Yemen (YEM) |
| NEVENA | Bulgaria (BG) | SESA 22 | Yemen (YEM) |
| AIDA | Bulgaria (BG) | SESA 23 | Yemen (YEM) |
| MILENA | Italy (ΙΤ) | SESA 24 | Yemen (YEM) |
| TOURKIAS | Turkey (TR) | SESA 25 | Yemen (YEM) |
| SESA 26 | Yemen (YEM) | ||
| SESA 27 | Yemen (YEM) | ||
| SESA 28 | Yemen (YEM) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stavridou, E.; Lagiotis, G.; Kalaitzidou, P.; Grigoriadis, I.; Bosmali, I.; Tsaliki, E.; Tsiotsiou, S.; Kalivas, A.; Ganopoulos, I.; Madesis, P. Characterization of the Genetic Diversity Present in a Diverse Sesame Landrace Collection Based on Phenotypic Traits and EST-SSR Markers Coupled With an HRM Analysis. Plants 2021, 10, 656. https://doi.org/10.3390/plants10040656
Stavridou E, Lagiotis G, Kalaitzidou P, Grigoriadis I, Bosmali I, Tsaliki E, Tsiotsiou S, Kalivas A, Ganopoulos I, Madesis P. Characterization of the Genetic Diversity Present in a Diverse Sesame Landrace Collection Based on Phenotypic Traits and EST-SSR Markers Coupled With an HRM Analysis. Plants. 2021; 10(4):656. https://doi.org/10.3390/plants10040656
Chicago/Turabian StyleStavridou, Evangelia, Georgios Lagiotis, Parthena Kalaitzidou, Ioannis Grigoriadis, Irini Bosmali, Eleni Tsaliki, Stiliani Tsiotsiou, Apostolos Kalivas, Ioannis Ganopoulos, and Panagiotis Madesis. 2021. "Characterization of the Genetic Diversity Present in a Diverse Sesame Landrace Collection Based on Phenotypic Traits and EST-SSR Markers Coupled With an HRM Analysis" Plants 10, no. 4: 656. https://doi.org/10.3390/plants10040656
APA StyleStavridou, E., Lagiotis, G., Kalaitzidou, P., Grigoriadis, I., Bosmali, I., Tsaliki, E., Tsiotsiou, S., Kalivas, A., Ganopoulos, I., & Madesis, P. (2021). Characterization of the Genetic Diversity Present in a Diverse Sesame Landrace Collection Based on Phenotypic Traits and EST-SSR Markers Coupled With an HRM Analysis. Plants, 10(4), 656. https://doi.org/10.3390/plants10040656