
Pulse Root Ideotype for Water Stress in Temperate Cropping System


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
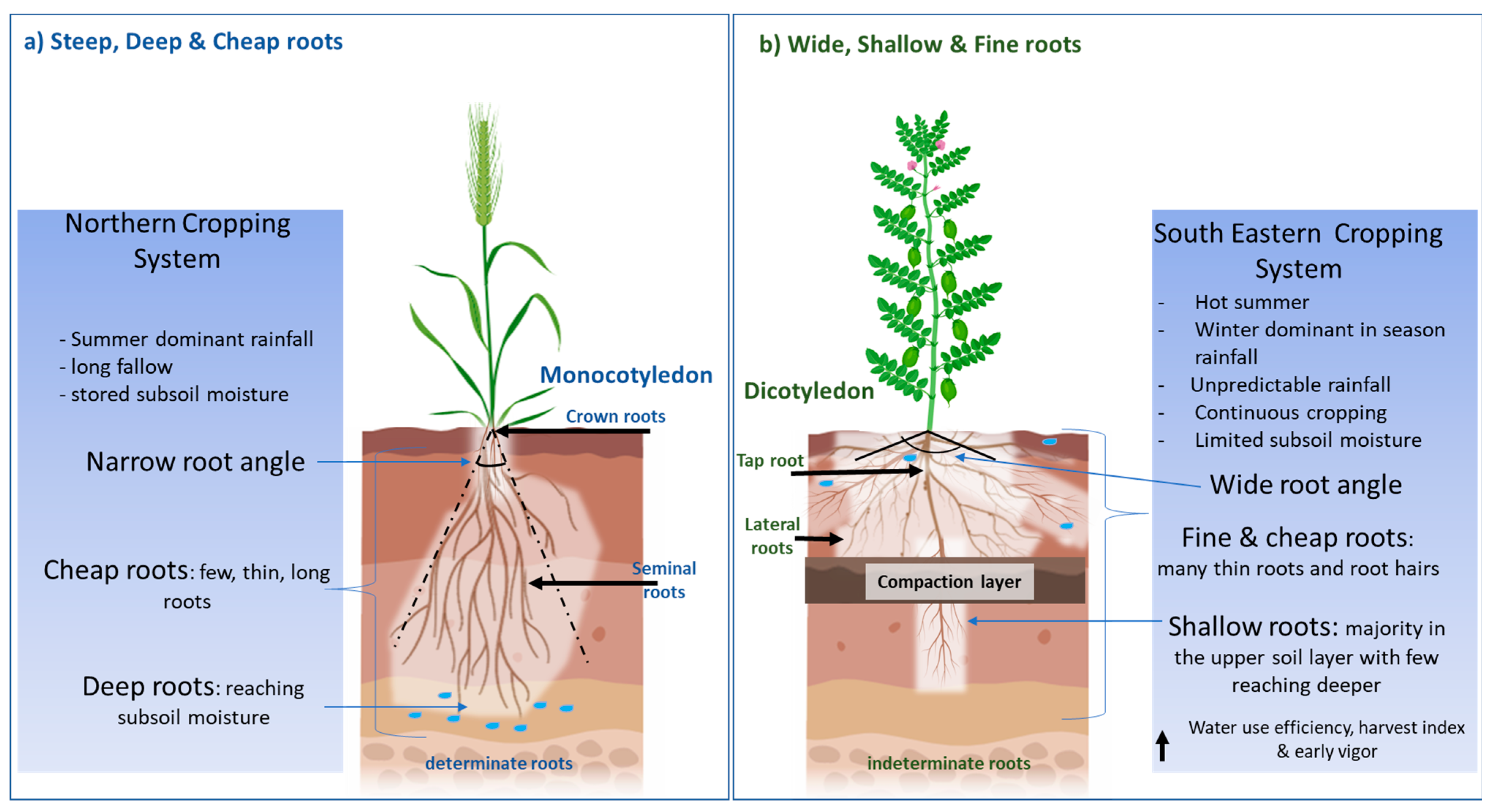
2. Ideotype Context and Traits
2.1. Soil, Climate and Constraints
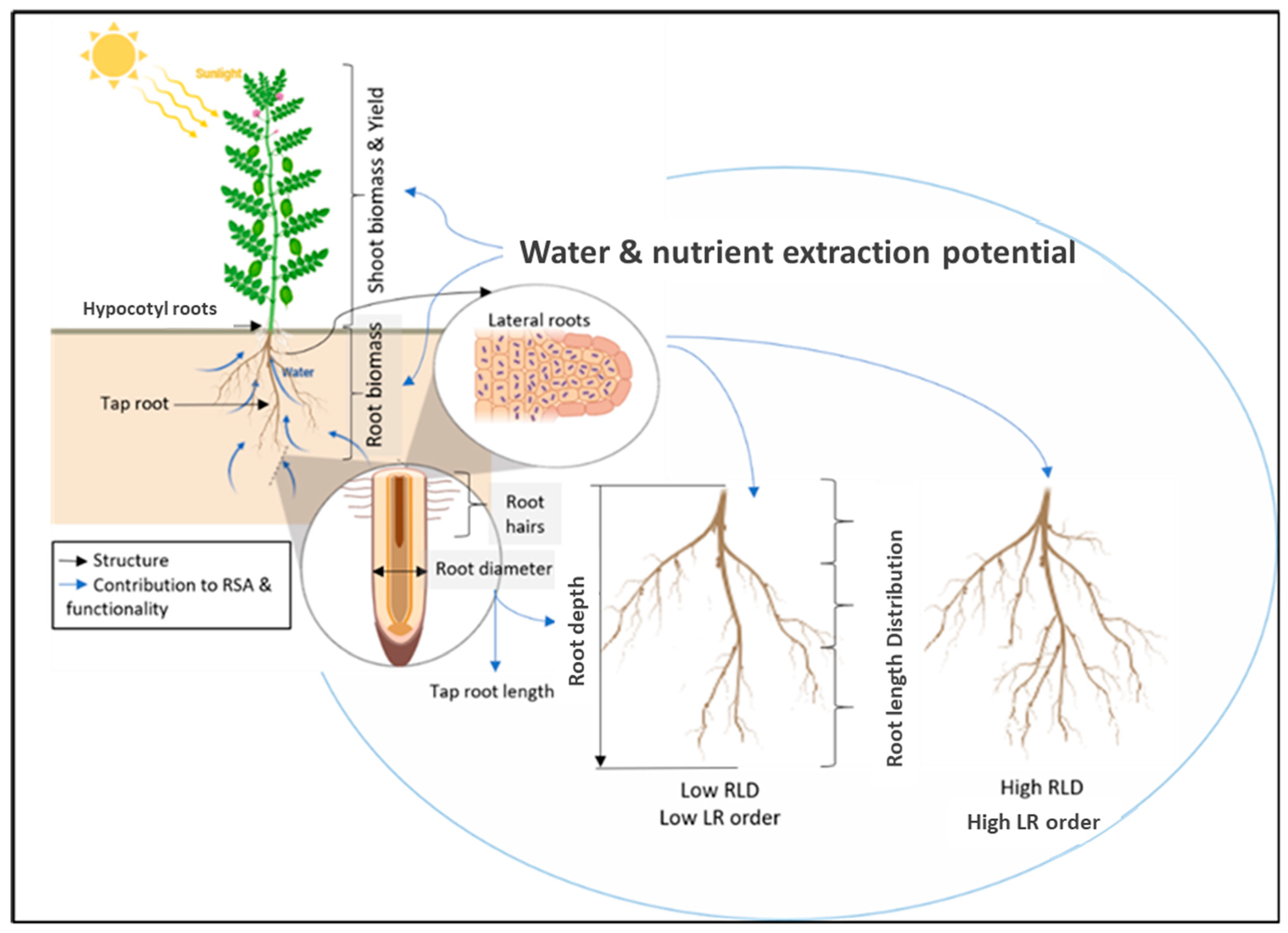
2.2. A New Root Ideotype—Wide, Shallow and Fine
3. Morphological Traits and Their Responses to Water Stress
3.1. Root System Differences between Cereals and Pulses
3.2. Taproot Length and Rooting Depth
3.3. Lateral Root Number Intensity and Density in the Soil Profile
3.4. Root Diameter
Xylem Vessel Diameter
4. Physiological Traits and Their Response to Water Stress
4.1. Vertical Stratification
4.1.1. Rooting Density
4.1.2. Root Mass and Root:Shoot Ratio
4.2. Surface Area
4.3. Root Angle
4.4. Water Movement in Pulses
4.4.1. Water Use Efficiency (WUE) in Pulses
4.4.2. Water Movements in Pulse Roots
4.4.3. Pulse Root Water Extraction Potential
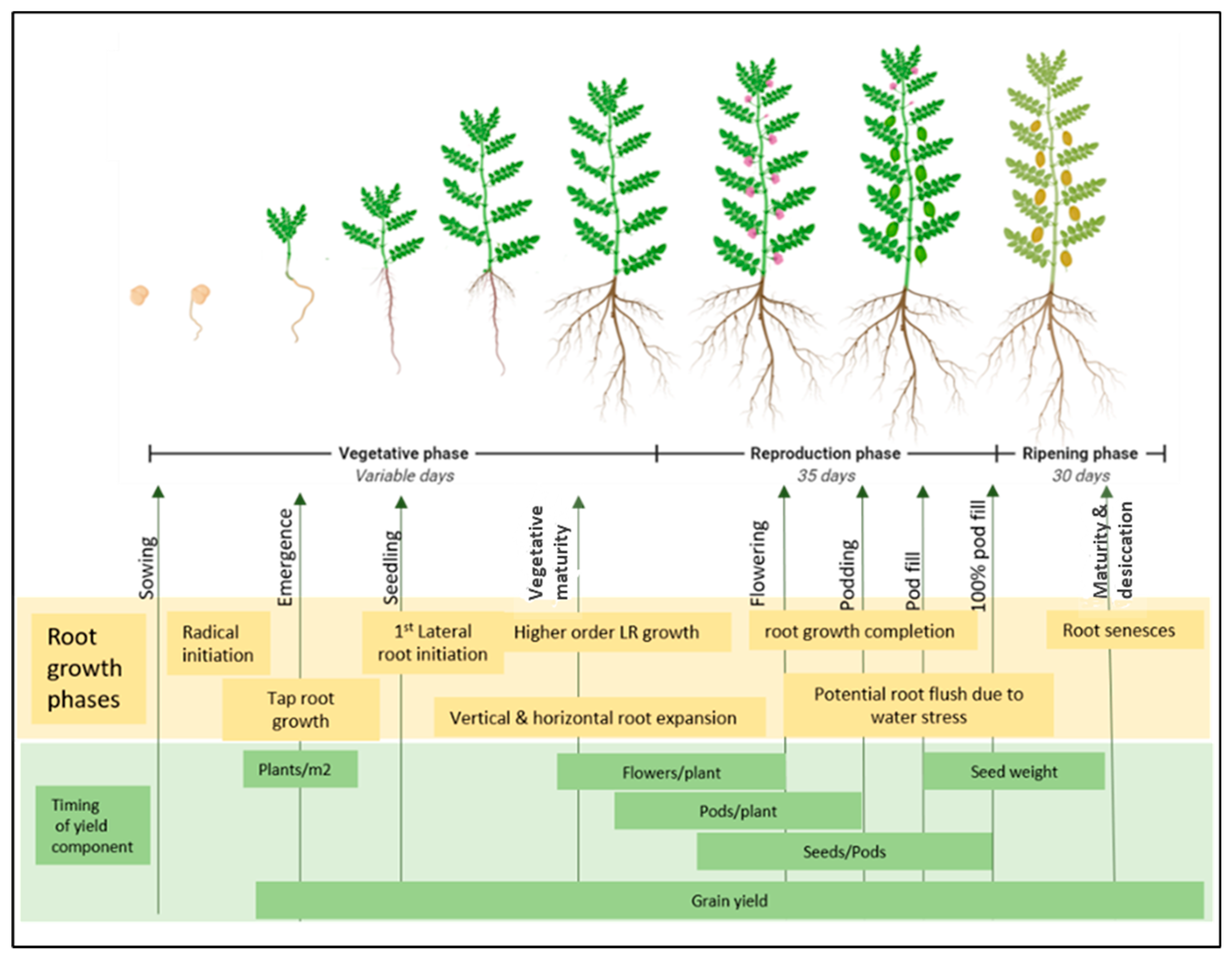
5. Phenological Traits and Their Response to Water Stress
Root Phenology
6. Research Priorities
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Campbell, C.A.; Zentner, R.P.; Basnyat, P.; Wang, H.; Selles, F.; McConkey, B.G.; Gan, Y.T.; Cutforth, H.W. Water use efficiency and water and nitrate distribution in soil in the semiarid prairie: Effect of crop type over 21 years. Can. J. Plant Sci. 2007, 87, 815–827. [Google Scholar] [CrossRef]
- Gan, Y.; Campbell, C.; Liu, L.; Basnyat, P.; McDonald, C. Water use and distribution profile under pulse and oilseed crops in semiarid northern high latitude areas. Agric. Water Manag. 2009, 96, 337–348. [Google Scholar] [CrossRef]
- Pulse Australia. Pulse Australia. Available online: http://www.pulseaus.com.au (accessed on 1 October 2019).
- Drew, E.; Herridge, D.; Ballard, R.; O’Hara, G.; Deaker, R.; Denton, M.; Yates, R.; Gemell, G.; Hartley, E.; Phillips, L.; et al. Inoculating Legumes: Apracticle Guide; Grains Research and Development Organisation. 2012. Available online: https://grdc.com.au/resources-and-publications/all-publications/bookshop/2015/07/inoculating-legumes (accessed on 1 October 2019).
- Sreeman, S.M.; Vijayaraghavareddy, P.; Sreevathsa, R.; Rajendrareddy, S.; Arakesh, S.; Bharti, P.; Dharmappa, P.; Soolanayakanahally, R. Introgression of physiological traits for a comprehensive improvement of drought adaptation in crop plants. Front. Chem. 2018, 6, 92. [Google Scholar] [CrossRef]
- Kebede, A.; Manjit, S.K.; Endashaw, B. Advances in mechanisms of drought tolerance in crops, with emphasis on barley. In Advances in Agronomy; Donald, L.S., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 156, pp. 266–300. [Google Scholar]
- Lynch, J.P. Steep, cheap and deep: An ideotype to optimize water and N acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Lilley, J.M.; Kirkegaard, J.A. Seasonal variation in the value of subsoil water to wheat: Simulation studies in southern New South Wales. Aust. J. Agric. Res. 2007, 58, 1115–1128. [Google Scholar] [CrossRef]
- Carroll, C.; Halpin, M.; Burger, P.; Bell, K.; Sallaway, M.M.; Yule, D.F. The effect of crop type, crop rotation, and tillage practice on runoff and soil loss on a Vertisol in central Queensland. Soil Res. 1997, 35, 925. [Google Scholar] [CrossRef]
- Adcock, D.; McNeill, A.M.; McDonald, G.K.; Armstrong, R.D. Subsoil constraints to crop production on neutral and alkaline soils in south-eastern Australia: A review of current knowledge and management strategies. Aust. J. Exp. Agric. 2007, 47, 1245–1261. [Google Scholar] [CrossRef]
- Lynch, J.P.; Chimungu, J.G.; Brown, K.M. Root anatomical phenes associated with water acquisition from drying soil: Targets for crop improvement. J. Exp. Bot. 2014, 65, 6155–6166. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Gan, Y.; Bueckert, R.A.; Van Rees, K. Rooting systems of oilseed and pulse crops. II: Vertical distribution patterns across the soil profile. Field Crop. Res. 2011, 122, 248–255. [Google Scholar] [CrossRef]
- Lynch, J. Root architecture and plant productivity. Plant Physiol. 1995, 109, 7–13. [Google Scholar] [CrossRef]
- Ramamoorthy, P.; Lakshmanan, K.; Upadhyaya, H.D.; Vadez, V.; Varshney, R.K. Root traits confer grain yield advantages under terminal drought in chickpea (Cicer arietinum L.). Field Crop. Res. 2017, 201, 146–161. [Google Scholar] [CrossRef] [PubMed]
- Kashiwagi, J.; Krishnamurthy, L.; Purushothaman, R.; Upadhyaya, H.; Gaur, P.; Gowda, C.; Ito, O.; Varshney, R. Scope for improvement of yield under drought through the root traits in chickpea (Cicer arietinum L.). Field Crop. Res. 2015, 170, 47–54. [Google Scholar] [CrossRef]
- Gorim, L.Y.; Vandenberg, A. Evaluation of wild lentil species as genetic resources to improve drought tolerance in cultivated lentil. Front. Plant Sci. 2017, 8, 1129. [Google Scholar] [CrossRef]
- Idrissi, O.; Houasli, C.; Udupa, S.M.; De Keyser, E.; Van Damme, P.; De Riek, J. Genetic variability for root and shoot traits in a lentil (Lens culinaris Medik.) recombinant inbred line population and their association with drought tolerance. Euphytica 2015, 204, 693–709. [Google Scholar] [CrossRef]
- Redden, R.J.; Berger, J.D. History and orgin of chickpea. In Chickpea Breeding and Management; Yadav, S.S., Ed.; CAB International: Wallingford, UK, 2007; pp. 1–13. [Google Scholar]
- Nuttall, J.G.; Armstrong, R.D.; Connor, D.J.; Matassa, V.J. Interrelationships between edaphic factors potentially limiting cereal growth on alkaline soils in north-western Victoria. Soil Res. 2003, 41, 277–292. [Google Scholar] [CrossRef]
- Sadras, V.O.; Hayman, P.T.; Rodriguez, D.; Monjardino, M.; Bielich, M.; Unkovich, M.; Mudge, B.; Wang, E. Interactions between water and nitrogen in Australian cropping systems: Physiological, agronomic, economic, breeding and modelling perspectives. Crop. Pasture Sci. 2016, 67, 1019–1053. [Google Scholar] [CrossRef]
- Verburg, K.; McBeath, T.M.; Armstrong, R.D.; Tavakkoli, E.; Wilhelm, N.; Mclaughlin, M.J.; Haling, R.E.; Richardson, A.E.; Mason, S.D.; Kirkegaard, J.A.; et al. Soil water dynamics as a function of soil and climate to inform phosphorus placement strategies. In Proceedings of the Soil Science Australia and New Zealand Scoiety of Soil Science Joint Conference, Cains, Australia, 29 November 2020. [Google Scholar]
- Reid, C.D.; Strain, B.R. Effects of CO2 enrichment on whole-plant carbon budget of seedlings of Fagus grandifolia and Acer saccharum in low irradiance. Oecologia 1994, 98, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Gutschick, V.P. Photosynthesis, Growth Rate, and Biomass Allocation. In Ecology in Agriculture; Elsevier: Amsterdam, The Netherlands, 1997; pp. 39–78. [Google Scholar]
- Urry, L.A.; Meyers, N.; Cain, M.L.; Wasserma, S.A.; Minorsk, P.V.; Reece, J.B. Campbell Biology: Australian and New Zealand Edition, 11th ed.; Pearson: Melbourne, VIC, Australia, 2017. [Google Scholar]
- Adu, M.O.; Asare, P.A.; Yawson, D.O.; Dzidzienyo, D.K.; Nyadanu, D.; Asare-Bediako, E.; Afutu, E.; Tachie-Menson, J.W.; Amoah, M.N. Identifying key contributing root system traits to genetic diversity in field-grown cowpea (Vigna unguiculata L. Walp.) genotypes. Field Crop. Res. 2019, 232, 106–118. [Google Scholar] [CrossRef]
- Gregory, P.J. Root growth of chickpea, faba bean, lentil, and pea and effects of water and salt stresses. In Nitrogen Fixation: From Molecules to Crop Productivity; Springer Science and Business Media LLC: Berlin, Germany, 1988; Volume 5, pp. 857–867. [Google Scholar]
- Manschadi, A.M.; Christopher, J.; Devoil, P.; Hammer, G.L. The role of root architectural traits in adaptation of wheat to water-limited environments. Funct. Plant Biol. 2006, 33, 823–837. [Google Scholar] [CrossRef]
- Lopes, M.S.; Reynolds, M.P. Partitioning of assimilates to deeper roots is associated with cooler canopies and increased yield under drought in wheat. Funct. Plant Biol. 2010, 37, 147–156. [Google Scholar] [CrossRef]
- Bengough, A.G.; McKenzie, B.M.; Hallett, P.D.; Valentine, T.A. Root elongation, water stress, and mechanical impedance: A review of limiting stresses and beneficial root tip traits. J. Exp. Bot. 2011, 62, 59–68. [Google Scholar] [CrossRef]
- Lucas, M.; Schlüter, S.; Vogel, H.-J.; Vetterlein, D. Roots compact the surrounding soil depending on the structures they encounter. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Han, E.; Kautz, T.; Perkons, U.; Uteau, D.; Peth, S.; Huang, N.; Horn, R.; Köpke, U. Root growth dynamics inside and outside of soil biopores as affected by crop sequence determined with the profile wall method. Biol. Fertil. Soils 2015, 51, 847–856. [Google Scholar] [CrossRef]
- Reynolds, M.; Ortiz-Monasterio, A.M. Application of Physiological in Wheat Breeding; CIMMYT: Mexico city, Mexico, 2001; Volume 31, pp. 162–171. [Google Scholar]
- Le Marié, C.A.; York, L.M.; Strigens, A.; Malosetti, M.; Camp, K.-H.; Giuliani, S.; Lynch, J.P.; Hund, A. Shovelomics root traits assessed on the EURoot maize panel are highly heritable across environments but show low genotype-by-nitrogen interaction. Euphytica 2019, 215, 173. [Google Scholar] [CrossRef]
- Sarker, A.; Erskine, W.; Singh, M. Variation in shoot and root characteristics and their association with drought tolerance in lentil landraces. Genet. Resour. Crop. Evol. 2005, 52, 89–97. [Google Scholar] [CrossRef]
- Futsaether, C.M.; Oxaal, U. A growth chamber for idealized studies of seedling root growth dynamics and structure. Plant Soil 2002, 246, 221–230. [Google Scholar] [CrossRef]
- Chen, Y.; Ghanem, M.E.; Siddique, K.H. Characterising root trait variability in chickpea (Cicer arietinum L.) germplasm. J. Exp. Bot. 2016, 68, 1987–1999. [Google Scholar] [CrossRef]
- Belachew, K.Y.; Nagel, K.A.; Fiorani, F.; Stoddard, F.L. Diversity in root growth responses to moisture deficit in young faba bean (Vicia faba L.) plants. PeerJ 2018, 6, e4401. [Google Scholar] [CrossRef] [PubMed]
- Zarebanadkouki, M.; Kim, Y.X.; Carminati, A. Where do roots take up water? Neutron radiography of water flow into the roots of transpiring plants growing in soil. New Phytol. 2013, 199, 1034–1044. [Google Scholar] [CrossRef]
- Popova, L.; Van Dusschoten, D.; Nagel, K.A.; Fiorani, F.; Mazzolai, B. Plant root tortuosity: An indicator of root path formation in soil with different composition and density. Ann. Bot. 2016, 118, 685–698. [Google Scholar] [CrossRef]
- Clark, L.; Whalley, W.; Barraclough, P. How do roots penetrate strong soil? Plant Soil 2003, 255, 93–104. [Google Scholar] [CrossRef]
- Singh, V.; Van Oosterom, E.J.; Jordan, D.R.; Messina, C.D.; Cooper, M.; Hammer, G.L. Morphological and architectural development of root systems in sorghum and maize. Plant Soil 2010, 333, 287–299. [Google Scholar] [CrossRef]
- Van Der Bom, F.J.T.; Williams, A.; Bell, M.J. Root architecture for improved resource capture: Trade-offs in complex environments. J. Exp. Bot. 2020, 71, 5752–5763. [Google Scholar] [CrossRef]
- Prince, S.J.; Murphy, M.; Mutava, R.N.; Durnell, L.A.; Valliyodan, B.; Shannon, J.G.; Nguyen, H.T. Root xylem plasticity to improve water use and yield in water-stressed soybean. J. Exp. Bot. 2017, 68, 2027–2036. [Google Scholar] [CrossRef]
- Richards, R.; Passioura, J. A breeding program to reduce the diameter of the major xylem vessel in the seminal roots of wheat and its effect on grain yield in rain-fed environments. Aust. J. Agric. Res. 1989, 40, 943–950. [Google Scholar] [CrossRef]
- Richards, R.A.; Passioura, J.B. Seminal root morphology and water use of wheat II. Genetic variation 1. Crop. Sci. 1981, 21, 253–255. [Google Scholar] [CrossRef]
- Benjamin, J.; Nielsen, D. Water deficit effects on root distribution of soybean, field pea and chickpea. Field Crop. Res. 2006, 97, 248–253. [Google Scholar] [CrossRef]
- Kashiwagi, J.; Krishnamurthy, L.; Crouch, J.; Serraj, R. Variability of root length density and its contributions to seed yield in chickpea (Cicer arietinum L.) under terminal drought stress. Field Crop. Res. 2006, 95, 171–181. [Google Scholar] [CrossRef]
- Manschadi, A.; Sauerborn, J.; Stützel, H.; Göbel, W.; Saxena, M. Simulation of faba bean (Vicia faba L.) root system development under Mediterranean conditions. Eur. J. Agron. 1998, 9, 259–272. [Google Scholar] [CrossRef]
- Serraj, R.; Krishnamurthy, L.; Kashiwagi, J.; Kumar, J.; Chandra, S.; Crouch, J. Variation in root traits of chickpea (Cicer arietinum L.) grown under terminal drought. Field Crop. Res. 2004, 88, 115–127. [Google Scholar] [CrossRef]
- Zaman-Allah, M.; Jenkinson, D.M.; Vadez, V. A conservative pattern of water use, rather than deep or profuse rooting, is critical for the terminal drought tolerance of chickpea. J. Exp. Bot. 2011, 62, 4239–4252. [Google Scholar] [CrossRef]
- Gorim, L.Y.; Vandenberg, A. Root traits, nodulation and root distribution in soil for five wild lentil species and lens culinaris (Medik.) Grown under well-watered conditions. Front. Plant Sci. 2017, 8, 1632. [Google Scholar] [CrossRef]
- Zaman-Allah, M.; Jenkinson, D.M.; Vadez, V. Chickpea genotypes contrasting for seed yield under terminal drought stress in the field differ for traits related to the control of water use. Funct. Plant Biol. 2011, 38, 270–281. [Google Scholar] [CrossRef]
- Gahoonia, T.S.; Ali, O.; Sarker, A.; Nielsen, N.E.; Rahman, M.M. Genetic variation in root traits and nutrient acquisition of lentil genotypes. J. Plant Nutr. 2006, 29, 643–655. [Google Scholar] [CrossRef]
- Gorim, L.Y.; Rabani, E.M.; Barlow, B.A.; de Silva, D.; Vandenberg, A. Are artificial media valid for root analysis? A case study comparing root traits of five lentil genotypes in artificial media versus soil. J. Soil Sci. Plant Health 2018, 2, 1. [Google Scholar]
- Manschadi, A.M.; Christopher, J.T.; Hammer, G.L.; Devoil, P. Experimental and modelling studies of drought-adaptive root architectural traits in wheat (Triticum aestivum L.). Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2010, 144, 458–462. [Google Scholar] [CrossRef]
- Wang, L.; Guo, M.; Lingling, W.; Ruan, W.; Mo, X.; Wu, Z.; Sturrock, C.J.; Wenyuan, R.; Lu, C.; Peng, J.; et al. LARGE ROOT ANGLE1, encoding OsPIN2, is involved in root system architecture in rice. J. Exp. Bot. 2018, 69, 385–397. [Google Scholar] [CrossRef]
- Waite, J.M.; Collum, T.D.; Dardick, C. AtDRO1 is nuclear localized in root tips under native conditions and impacts auxin localization. Plant Mol. Biol. 2020, 103, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Zhan, A.; Lynch, J.P. Reduced frequency of lateral root branching improves N capture from low-N soils in maize. J. Exp. Bot. 2015, 66, 2055–2065. [Google Scholar] [CrossRef]
- Liao, H.; Rubio, G.; Yan, X.; Cao, A.; Brown, K.M.; Lynch, J.P. Effect of phosphorus availability on basal root shallowness in common bean. Plant Soil 2001, 232, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Miguel, M.A.; Widrig, A.; Vieira, R.F.; Brown, K.M.; Lynch, J.P. Basal root whorl number: A modulator of phosphorus acquisition in common bean (Phaseolus vulgaris). Ann. Bot. 2013, 112, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Burridge, J.; Jochua, C.N.; Bucksch, A.; Lynch, J.P. Legume shovelomics: High—Throughput phenotyping of common bean (Phaseolus vulgaris L.) and cowpea (Vigna unguiculata subsp, unguiculata) root architecture in the field. Field Crop. Res. 2016, 192, 21–32. [Google Scholar] [CrossRef]
- Van Duivenbooden, N.; Pala, M.; Studer, C.; Bielders, C.; Beukes, D. Cropping systems and crop complementarity in dryland agriculture to increase soil water use efficiency: A review. NJAS Wagening. J. Life Sci. 2000, 48, 213–236. [Google Scholar] [CrossRef]
- Varney, G.T.; Canny, M.J. Rates of water uptake into the mature root system of maize plants. New Phytol. 1993, 123, 775–786. [Google Scholar] [CrossRef]
- Ahmed, M.A.; Zarebanadkouki, M.; Meunier, F.; Javaux, M.; Kaestner, A.; Carminati, A. Root type matters: Measurement of water uptake by seminal, crown, and lateral roots in maize. J. Exp. Bot. 2018, 69, 1199–1206. [Google Scholar] [CrossRef]
- Wiecheteck, L.H.; Giarola, N.F.; De Lima, R.P.; Tormena, C.A.; Torres, L.C.; De Paula, A.L. Comparing the classical permanent wilting point concept of soil (−15,000 hPa) to biological wilting of wheat and barley plants under contrasting soil textures. Agric. Water Manag. 2020, 230, 105965. [Google Scholar] [CrossRef]
- Wang, X.; Gan, Y.; Hamel, C.; Lemke, R.; McDonald, C. Water use profiles across the rooting zones of various pulse crops. Field Crop. Res. 2012, 134, 130–137. [Google Scholar] [CrossRef]
- Liu, L.; Gan, Y.; Bueckert, R.; Van Rees, K. Rooting systems of oilseed and pulse crops I: Temporal growth patterns across the plant developmental periods. Field Crop. Res. 2011, 122, 256–263. [Google Scholar] [CrossRef]
- Rawson, H.M.; Macpherson, H.G. Irrigated Wheat; Food and Agriculture Organization of the United Nations. 2000. Available online: http://www.fao.org/3/x8234e/x8234e00.htm#Contents (accessed on 1 October 2019).
- Palta, J.A.; Watt, M. Vigorous crop root systems: Form and function for improving the capture of water and nutrients. In Crop Physiology. Applications for Genetic Improvement and Agronomy; Sadras, V., Calderini, D., Eds.; Elsevier: Amsterdam, The Netherlands, 2009; pp. 309–325. [Google Scholar]
- Li, P.; Ma, B.; Palta, J.A.; Ding, T.; Cheng, Z.; Lv, G.; Xiong, Y. Wheat breeding highlights drought tolerance while ignores the advantages of drought avoidance: A meta-analysis. Eur. J. Agron. 2021, 122, 126196. [Google Scholar] [CrossRef]
- Siddiqui, N.; Léon, J.; Naz, A.A.; Ballvora, A. Genetics and genomics of root system variation in adaptation to drought stress in cereal crops. J. Exp. Bot. 2021, 72, 1007–1019. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rao, S.; Armstrong, R.; Silva-Perez, V.; Tefera, A.T.; Rosewarne, G.M. Pulse Root Ideotype for Water Stress in Temperate Cropping System. Plants 2021, 10, 692. https://doi.org/10.3390/plants10040692
Rao S, Armstrong R, Silva-Perez V, Tefera AT, Rosewarne GM. Pulse Root Ideotype for Water Stress in Temperate Cropping System. Plants. 2021; 10(4):692. https://doi.org/10.3390/plants10040692
Chicago/Turabian StyleRao, Shiwangni, Roger Armstrong, Viridiana Silva-Perez, Abeya T. Tefera, and Garry M. Rosewarne. 2021. "Pulse Root Ideotype for Water Stress in Temperate Cropping System" Plants 10, no. 4: 692. https://doi.org/10.3390/plants10040692
APA StyleRao, S., Armstrong, R., Silva-Perez, V., Tefera, A. T., & Rosewarne, G. M. (2021). Pulse Root Ideotype for Water Stress in Temperate Cropping System. Plants, 10(4), 692. https://doi.org/10.3390/plants10040692