
Seed Priming with Sorghum Water Extract Improves the Performance of Camelina (Camelina sativa (L.) Crantz.) under Salt Stress


 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Results
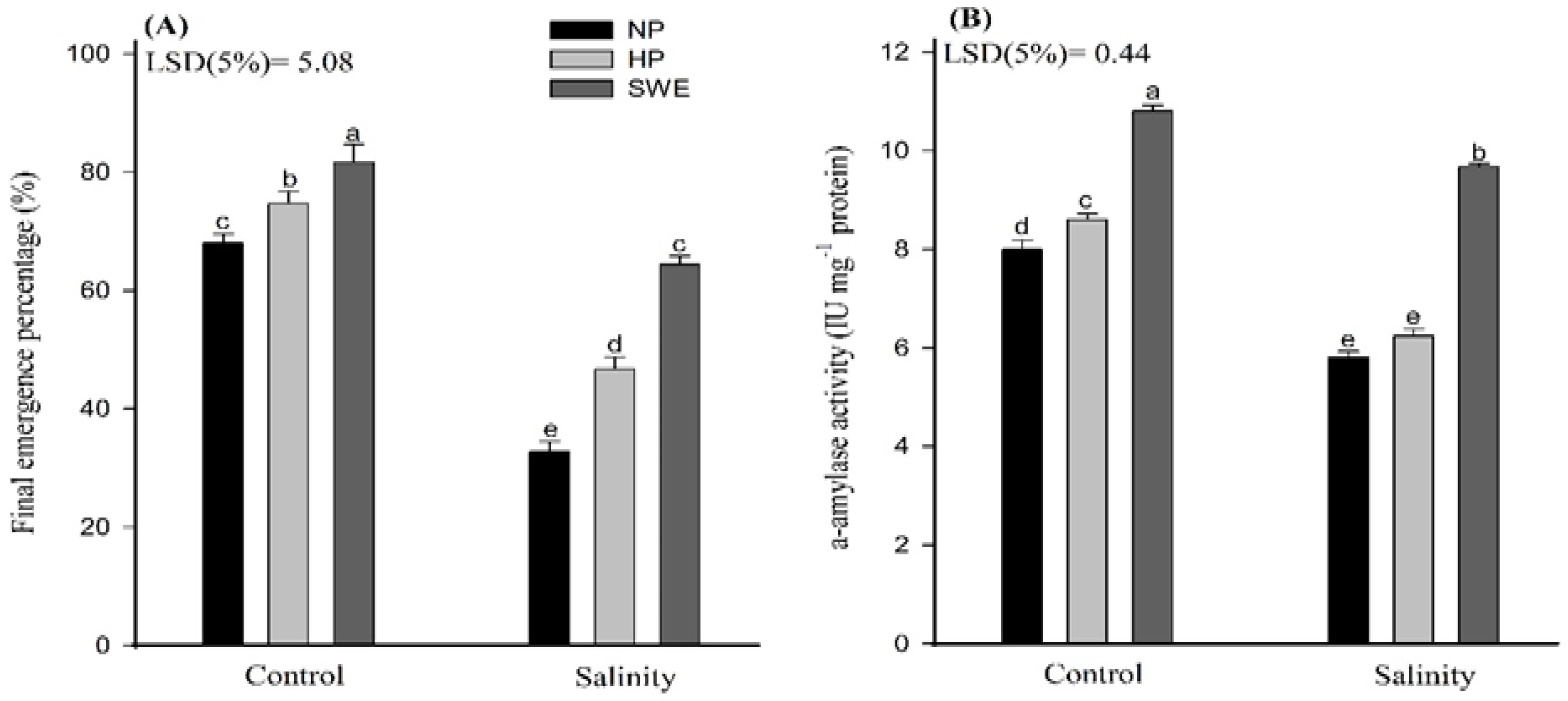
2.1. Emergence Traits
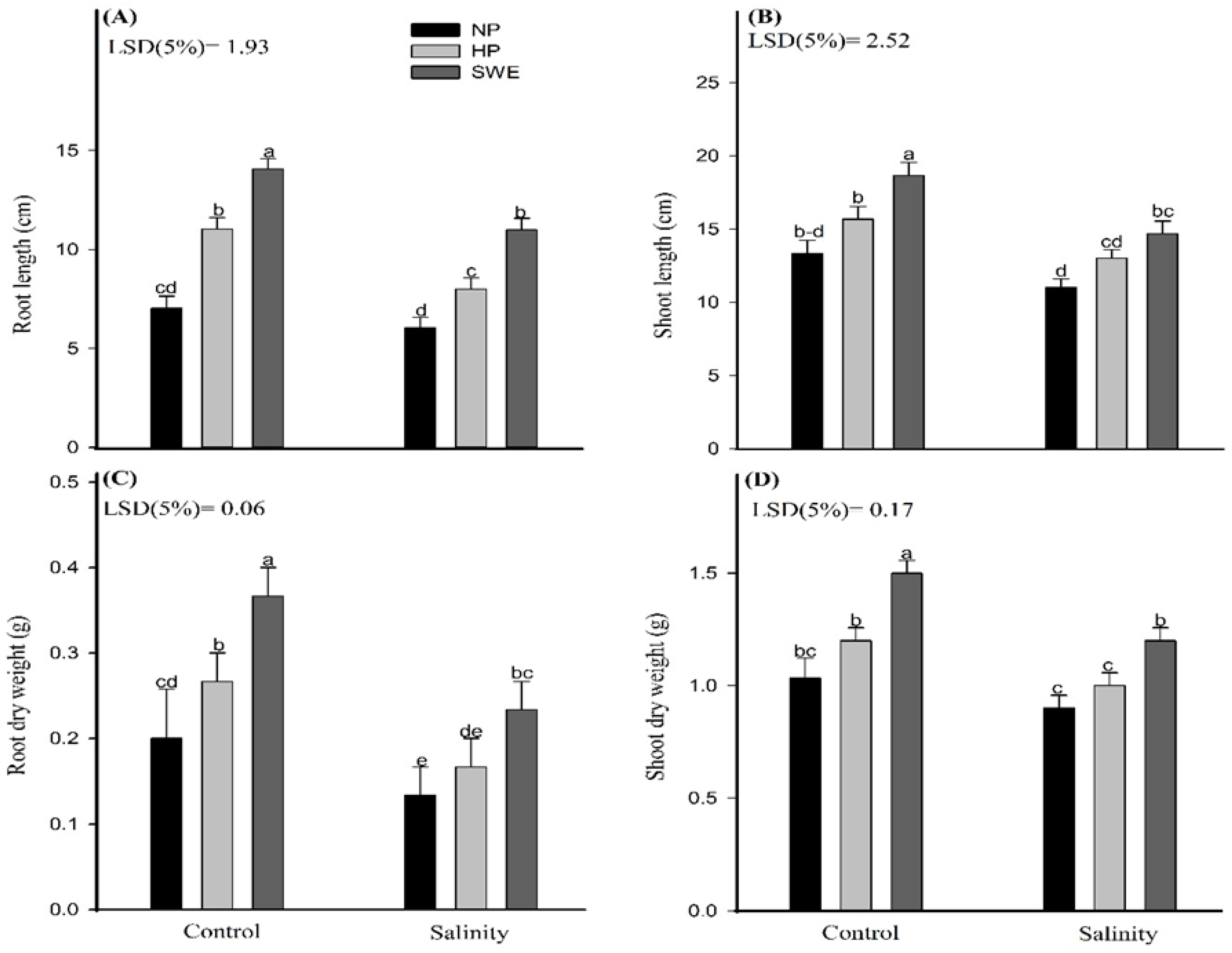
2.2. Seedling Growth
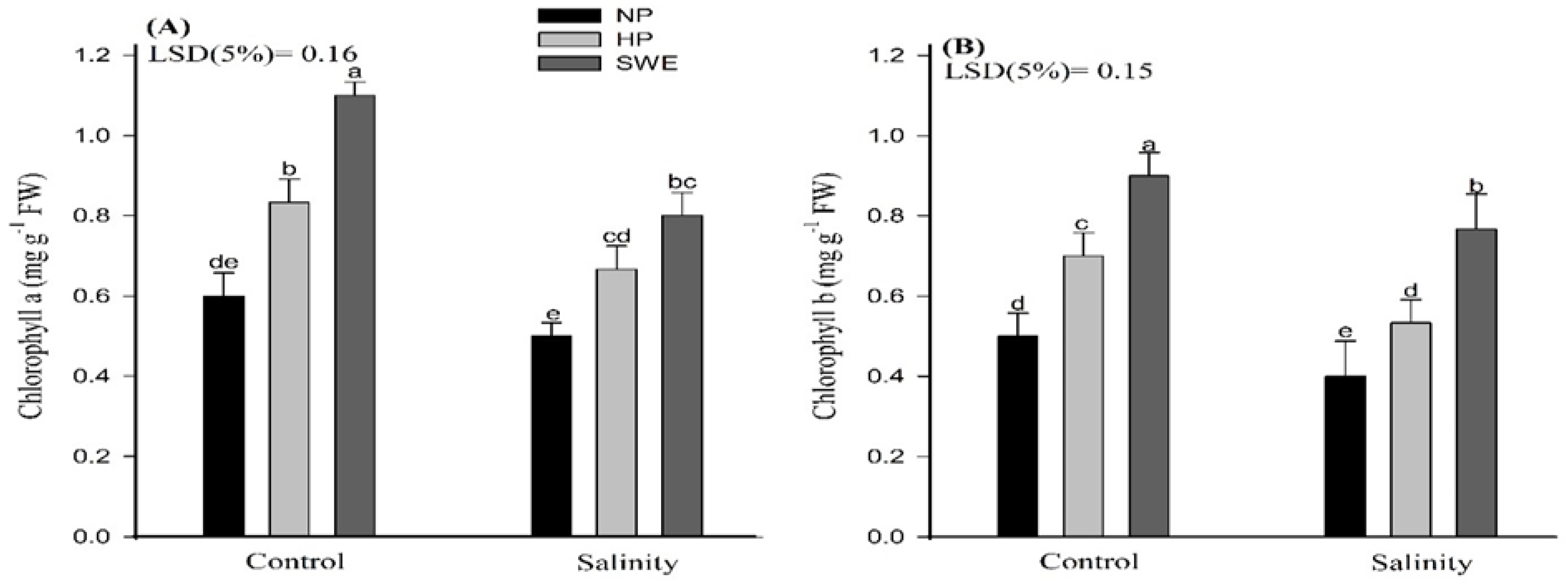
2.3. Chlorophyll Content
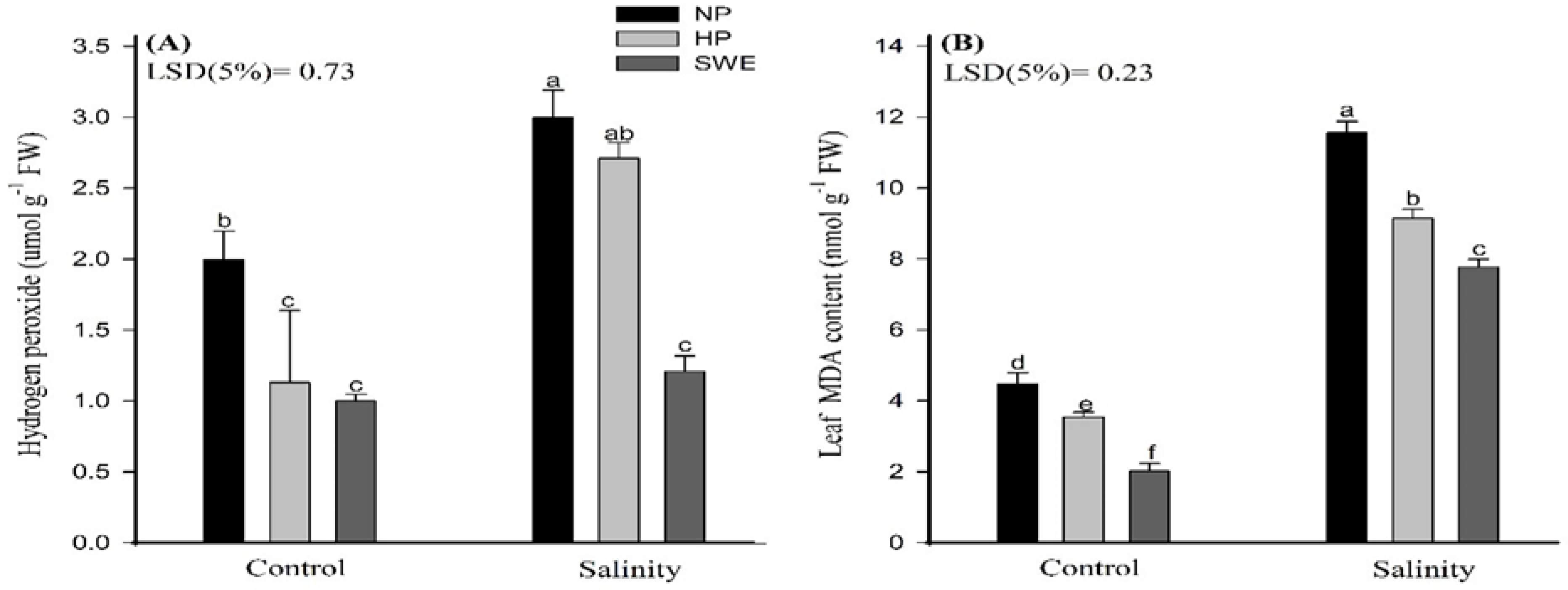
2.4. Leaf H2O2 and MDA Contents
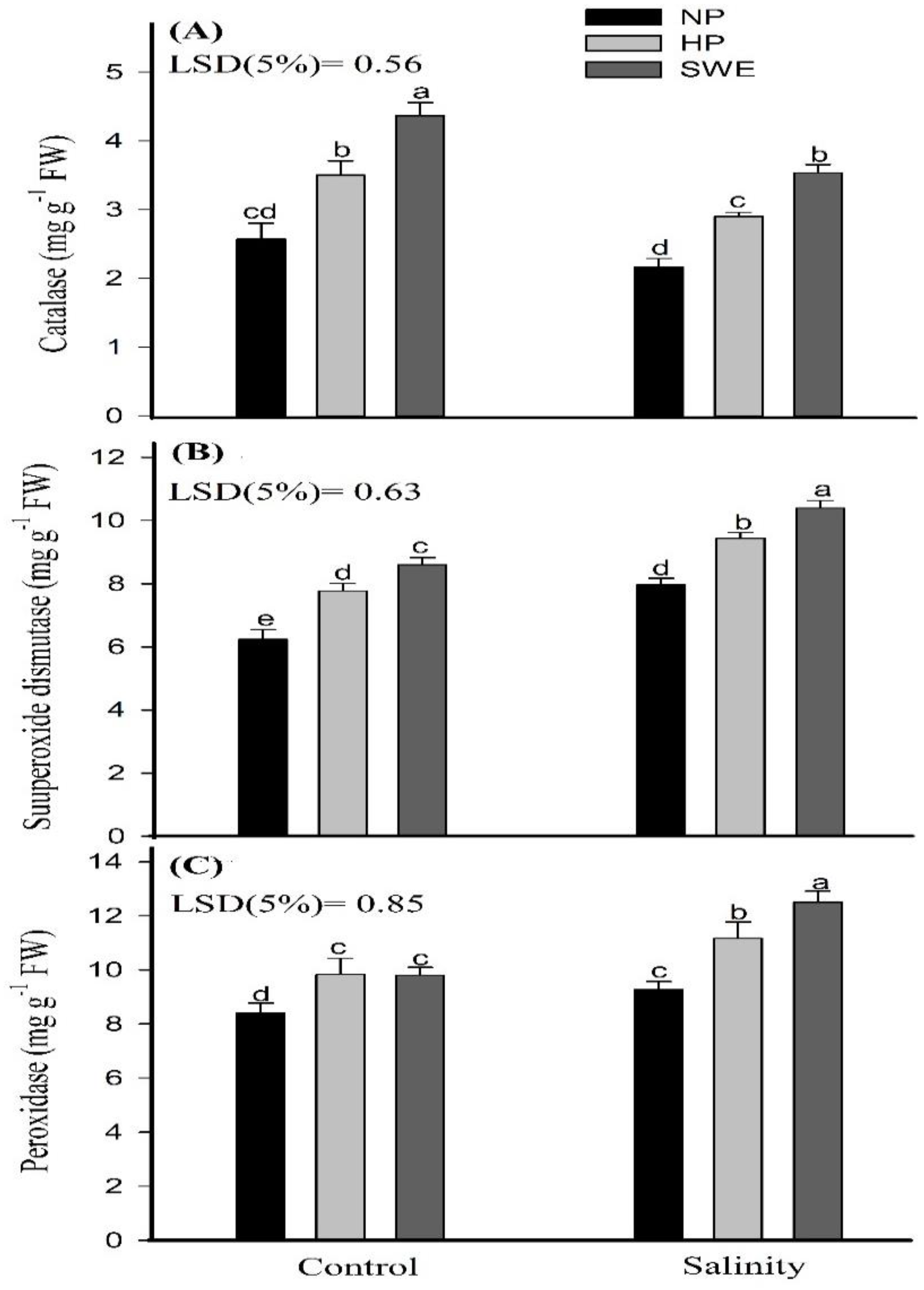
2.5. Antioxidant Enzymatic Activities
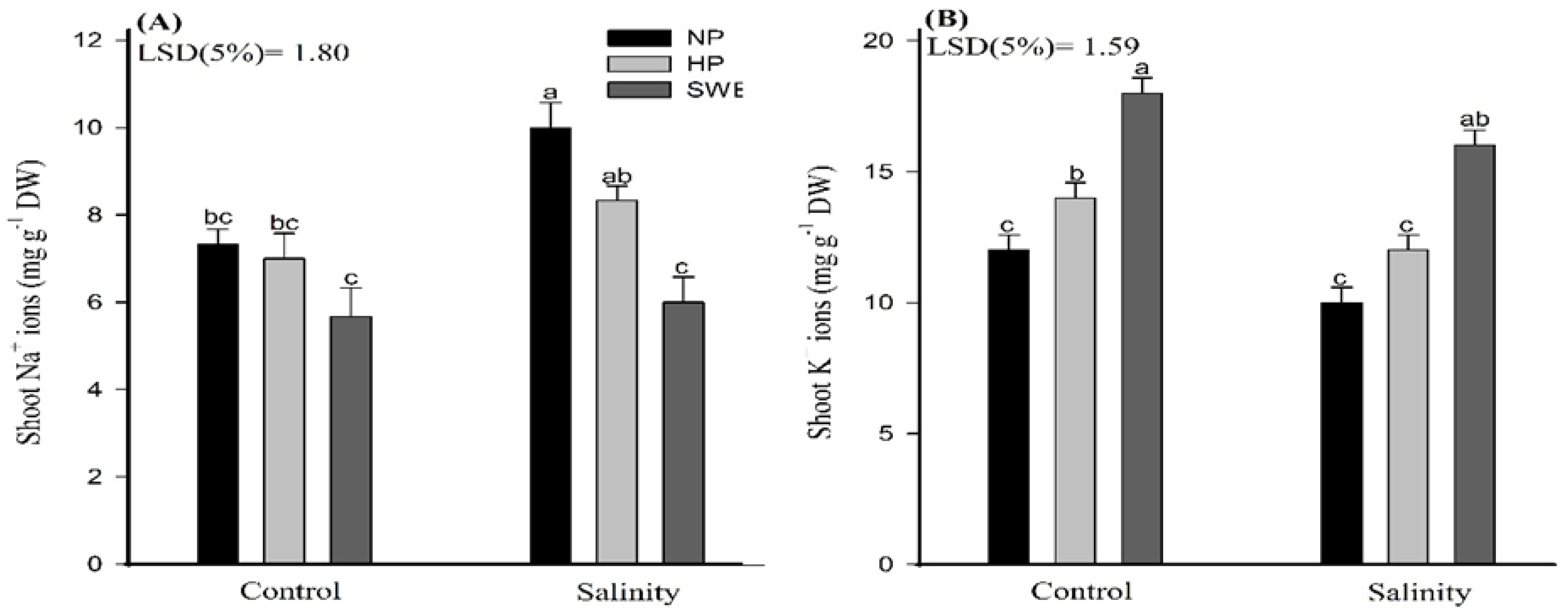
2.6. Mineral Ions Accumulation
3. Discussion
4. Materials and Methods
4.1. Experimental Detail
4.2. Experimental Design and Treatments
4.3. Observations
4.3.1. Final Emergence Percentage
4.3.2. Seedling Growth
4.3.3. α-Amylase Activity
4.3.4. Chlorophyll Contents Determination
4.3.5. Antioxidants Defense System
4.3.6. Determination of MDA and H2O2
4.3.7. Na+ and K+ Ions Determination
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yang, J.; Caldwell, C.; Corscadden, K.; He, Q.S.; Li, J. An evaluation of biodiesel production from Camelina sativa grown in Nova Scotia. Ind. Crop. Prod. 2016, 81, 162–168. [Google Scholar] [CrossRef]
- Adhikari, P.; Heo, J.; Nyachoti, C. Standardized total tract digestibility of phosphorus in camelina (Camelina sativa) meal fed to growing pigs without or phytase supplementation. Anim. Feed. Sci. Technol. 2016, 214, 104–109. [Google Scholar] [CrossRef]
- Ibrahim, F.M.; El Habbasha, S. Chemical composition, medicinal impacts and cultivation of camelina (Camelina sativa). Int. J. PharmTech Res. 2015, 8, 114–122. [Google Scholar]
- Gómez-Monedero, B.; Bimbela, F.; Arauzo, J.; Faria, J.; Ruiz, M.P. Pyrolysis of Red Eucalyptus, Camelina Straw, and Wheat Straw in an Ablative Reactor. Energy Fuels 2015, 29, 1766–1775. [Google Scholar] [CrossRef]
- Bonjean, A.; Le Goffic, F. La cameline. Camelina sativa (L.) Crantz: Une opportunité pour l’agriculture et l’industrie européennes. OCL Ol. Corps Gras Lipides 1999, 6, 28–34. [Google Scholar]
- Larsson, M. Cultivation and processing of Linum usitatissimum and Camelina sativa in southern Scandinavia during the Roman Iron Age. Veg. Hist. Archaeobotany 2013, 22, 509–520. [Google Scholar] [CrossRef]
- Blackshaw, R.; Johnson, E.; Gan, Y.; May, W.; McAndrew, D.; Barthet, V.; McDonald, T.J.; Wispinski, D. Alternative oilseed crops for biodiesel feedstock on the Canadian prairies. Can. J. Plant Sci. 2011, 91, 889–896. [Google Scholar] [CrossRef]
- Hunsaker, D.J.; French, A.N.; Clarke, T.R.; El-Shikha, D.M. Water use, crop coefficients, and irrigation management criteria for camelina production in arid regions. Irrig. Sci. 2010, 29, 27–43. [Google Scholar] [CrossRef]
- Berti, M.; Gesch, R.; Eynck, C.; Anderson, J.; Cermak, S. Camelina uses, genetics, genomics, production, and management. Ind. Crop. Prod. 2016, 94, 690–710. [Google Scholar] [CrossRef]
- Gao, L.; Caldwell, C.D.; Jiang, Y. Photosynthesis and Growth of Camelina and Canola in Response to Water Deficit and Applied Nitrogen. Crop. Sci. 2018, 58, 393–401. [Google Scholar] [CrossRef]
- Gesch, R.W. Influence of genotype and sowing date on camelina growth and yield in the north central U.S. Ind. Crop. Prod. 2014, 54, 209–215. [Google Scholar] [CrossRef]
- Eberle, C.A.; Thom, M.D.; Nemec, K.T.; Forcella, F.; Lundgren, J.G.; Gesch, R.W.; Riedell, W.E.; Papiernik, S.K.; Wagner, A.; Peterson, D.H.; et al. Using pennycress, camelina, and canola cash cover crops to provision pollinators. Ind. Crop. Prod. 2015, 75, 20–25. [Google Scholar] [CrossRef]
- Heydarian, Z.; Yu, M.; Gruber, M.; Coutu, C.; Robinson, S.J.; Hegedus, D.D. Changes in gene expression in Camelina sativa roots and vegetative tissues in response to salinity stress. Sci. Rep. 2018, 8, 9804. [Google Scholar] [CrossRef]
- Hussain, S.S.; Jamil, M.; Sadiq, M.; Mehdi, S.M. Alleviation of adverse effect of first irrigation to wheat crop in saline sodic soil. Soil Environ. 2012, 31, 1. [Google Scholar]
- Ghafoor, A.; Qadir, M.; Murtaza, G. Salt-Affected Soils: Principles of Management; Allied Book Centre: Punjab, Pakistan, 2004. [Google Scholar]
- Zhang, J. Salt-Affected Soil Resources in China. In Coastal Saline Soil Rehabilitation and Utilization Based on Forestry Approaches in China; Springer: Berlin, Germany, 2013; pp. 9–13. [Google Scholar]
- Steppuhn, H.; Falk, K.C.; Zhou, R. Emergence, height, grain yield and oil content of camelina and canola grown in saline media. Can. J. Soil Sci. 2010, 90, 151–164. [Google Scholar] [CrossRef]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Zahra, N.; Mahmood, S.; Raza, Z.A. Salinity stress on various physiological and biochemical attributes of two distinct maize (Zea mays L.) genotypes. J. Plant Nutr. 2018, 41, 1368–1380. [Google Scholar] [CrossRef]
- Jafar, M.Z.; Farooq, M.; Cheema, M.A.; Afzal, I.; Basra, S.M.; Wahid, M.A.; Aziz, T.; A Shahid, M. Improving the Performance of Wheat by Seed Priming Under Saline Conditions. J. Agron. Crop. Sci. 2011, 198, 38–45. [Google Scholar] [CrossRef]
- Hussain, S.; Khaliq, A.; Tanveer, M.; Matloob, A.; Hussain, H.A. Aspirin priming circumvents the salinity-induced effects on wheat emergence and seedling growth by regulating starch metabolism and antioxidant enzyme activities. Acta Physiol. Plant 2018, 40, 68. [Google Scholar] [CrossRef]
- Zheng, M.; Tao, Y.; Hussain, S.; Jiang, Q.; Peng, S.; Huang, J.; Cui, K.; Nie, L. Seed priming in dry direct-seeded rice: Consequences for emergence, seedling growth and associated metabolic events under drought stress. Plant Growth Regul. 2016, 78, 167–178. [Google Scholar] [CrossRef]
- Hussain, H.A.; Hussain, S.; Anjum, S.A.; Hussain, S. Seed Priming Toward Enhanced Chilling Tolerance in Field Crops: An Overview. In Priming and Pretreatment of Seeds and Seedlings; Springer: Berlin/Heidelberg, Germany, 2019; pp. 265–286. [Google Scholar]
- Farooq, M.; Rehman, A.; Al-Alawi, A.K.; Al-Busaidi, W.M.; Lee, D.-J. Integrated use of seed priming and biochar improves salt tolerance in cowpea. Sci. Hortic. 2020, 272, 109507. [Google Scholar] [CrossRef]
- Hussain, M.A.; Farooq, M.; Basra, S.M.A.; Lee, D.-J. Application of Moringa Allelopathy in Crop Sciences. In Allelopathy; Springer International Publishing: Cham, Switzerland, 2012; pp. 469–483. [Google Scholar]
- Farooq, M.; Bajwa, A.A.; Cheema, S.A.; Cheema, Z.A. Application of allelopathy in crop production. Int. J. Agric. Biol. 2013, 15, 1367–1378. [Google Scholar]
- Dawood, M.G. Stimulating Plant Tolerance Against Abiotic Stress Through Seed Priming. In Advances in Seed Priming; Springer International Publishing: Cham, Switzerland, 2018; pp. 147–183. [Google Scholar]
- Soliman, M.H.; Ahlam, H.H.; Hamdah, A.G.; Shroug, S. Allelopathic effect of Moringa oleifera leaves extract on seed germination and early seedling growth of faba bean (Vicia faba L.). Int. J. Agric. Technol. 2017, 13, 105–117. [Google Scholar]
- Farooq, O.; Hussain, Q.M.; Sarwar, N.; Nawaz, A.; Iqbal, M.M.; Shiaz, M. Seed Priming with Sorghum Water Extracts and Calcium Chloride Improves the Stand Establishment and Seedling Growth of Sunflower and Maize. Pak. J. Life Soc. Sci. 2018, 16, 2. [Google Scholar]
- Daur, I. Effects of hydro and hormonal priming on quinoa (Chenopodium quinoa Willd.) seed germination under salt and drought stress. Pak. J. Bot. 2018, 50, 1669–1673. [Google Scholar]
- Rashid, N.; Basra, S.M.; Shahbaz, M.; Iqbal, S.; Hafeez, M.B. Foliar applied moringa leaf extract induces terminal heat tolerance in quinoa. Int. J. Agric. Biol. 2018, 20, 157–164. [Google Scholar]
- Farooq, M.; Nadeem, F.; Arfat, M.Y.; Nabeel, M.; Musadaq, S.; Cheema, S.A.; Nawaz, A. Exogenous application of allelopathic water extracts helps improving tolerance against terminal heat and drought stresses in bread wheat (Triticum aestivum L. Em. Thell.). J. Agron. Crop. Sci. 2018, 204, 298–312. [Google Scholar] [CrossRef]
- Prabha, D.; Negi, S.; Kumari, P.; Negi, Y.K.; Chauhan, J. Effect of seed priming with some plant leaf extract on seedling growth characteristics and root rot disease in tomato. Int. J. Agric. Syst. 2016, 4, 46–51. [Google Scholar]
- Mavi, K.; Usunoğlu, F. Effects of pre-sowing treatments with allelopathic plant extracts on tree tomato (Solanum betaceum Cav.) seedling emergence and performance. Agron. Colomb. 2020, 38, 2. [Google Scholar]
- Muhammad, F.; Mazhar, H.; Abdul, W.; Rushna, M. Employing aqueous allelopathic extracts of sunflower in improving salinity tolerance of rice. J. Agric. Soc. Sci. 2011, 7, 75–80. [Google Scholar]
- Bogatek, R.; Gniazdowska, A. ROS and Phytohormones in Plant-Plant Allelopathic Interaction. Plant Signal. Behav. 2007, 2, 317–318. [Google Scholar] [CrossRef]
- Goldthwaite, J.J. Hormones in plant senescence. In Plant Hormones and Their Role in Plant Growth and Development; Springer: Berlin/Heidelberg, Germany, 1987; pp. 553–573. [Google Scholar]
- Available, N.A.N.; Duke, S.O. Ecophysiological aspects of allelopathy. Planta 2003, 217, 529–539. [Google Scholar] [CrossRef]
- A Macías, F.; Molinillo, J.M.G.; Varela, R.M.; Galindo, J.C.G. Allelopathy—A natural alternative for weed control. Pest Manag. Sci. 2007, 63, 327–348. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.Q.; Ye, S.F.; Zhang, M.F.; Hu, W.H. Effects of root exudates and aqueous root extracts of cucumber (Cucumis sativus) and allelochemicals, on photosynthesis and antioxidant enzymes in cucumber. Biochem. Syst. Ecol. 2003, 31, 129–139. [Google Scholar] [CrossRef]
- Salawu, S.O.; Bester, M.J.; Duodu, K.G. Phenolic Composition and Bioactive Properties of Cell Wall Preparations and Whole Grains of Selected Cereals and Legumes. J. Food Biochem. 2013, 38, 62–72. [Google Scholar] [CrossRef]
- Ben-Hammouda, M.; Kremer, R.J.; Minor, H.C.; Sarwar, M. A chemical basis for differential allelopathic potential of sorghum hybrids on wheat. J. Chem. Ecol. 1995, 21, 775–786. [Google Scholar] [CrossRef]
- Bajwa, A.A.; Farooq, M.; Nawaz, A. Seed priming with sorghum extracts and benzyl aminopurine improves the tolerance against salt stress in wheat (Triticum aestivum L.). Physiol. Mol. Biol. Plants 2018, 24, 239–249. [Google Scholar] [CrossRef]
- Al-Hussaini, M.K.; Alsaadawi, I.S. Mitigation of drought stress effect on growth and productivity of mung bean by foliar application of sorghum water extract. Iraqi J. Sci. 2013, 54, 560–568. [Google Scholar]
- Alsaadawi, I.; Al-Ekelle, M.; Al-Hamzawi, M. Differential allelopathic potential of grain sorghum genotypes to weeds. Allelopathy J. 2007, 19, 153–159. [Google Scholar]
- Afzal, I.; Akram, M.; Rehman, H.; Rashid, S.; Basra, S. Moringa leaf and sorghum water extracts and salicylic acid to alleviate impacts of heat stress in wheat. S. Afr. J. Bot. 2020, 129, 169–174. [Google Scholar] [CrossRef]
- Basra, S.; Iftikhar, M.; Afzal, I. Potential of moringa (Moringa oleifera) leaf extract as priming agent for hybrid maize seeds. Int. J. Agric. Biol. 2011, 13, 6. [Google Scholar]
- Bajwa, A.A.; Farooq, M. Seed priming with sorghum water extract and benzyl amino purine along with surfactant improves germination metabolism and early seedling growth of wheat. Arch. Agron. Soil Sci. 2016, 63, 319–329. [Google Scholar] [CrossRef]
- Saddiq, M.S.; Iqbal, S.; Afzal, I.; Ibrahim, A.M.H.; Bakhtavar, M.A.; Hafeez, M.B.; Jahanzaib; Maqbool, M.M. Mitigation of salinity stress in wheat (Triticum aestivum L.) seedlings through physiological seed enhancements. J. Plant Nutr. 2019, 42, 1192–1204. [Google Scholar] [CrossRef]
- Ahmad, W.; Noor, M.A.; Afzal, I.; Bakhtavar, M.A.; Nawaz, M.M.; Sun, X.; Zhou, B.; Ma, W.; Zhao, M. Improvement of Sorghum Crop through Exogenous Application of Natural Growth-Promoting Substances under a Changing Climate. Sustainability 2016, 8, 1330. [Google Scholar] [CrossRef]
- Li, Z.; Xu, J.; Gao, Y.; Wang, C.; Guo, G.; Luo, Y.; Huang, Y.; Hu, W.; Sheteiwy, M.S.; Guan, Y.; et al. The Synergistic Priming Effect of Exogenous Salicylic Acid and H2O2 on Chilling Tolerance Enhancement during Maize (Zea mays L.) Seed Germination. Front. Plant Sci. 2017, 8, 1153. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, E.A. Seed priming to alleviate salinity stress in germinating seeds. J. Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef]
- Barbosa, W.F.S.; Steiner, F.; de Oliveira, L.C.M.; Henrique, P.; das Chagas, M. Comparison of seed priming techniques with regards to germination and growth of watermelon seedlings in laboratory condition. Afr. J. Biotechnol. 2016, 15, 2596–2602. [Google Scholar]
- Oliveira, C.E.D.S.; Steiner, F.; Zuffo, A.M.; Zoz, T.; Alves, C.Z.; De Aguiar, V.C.B. Seed priming improves the germination and growth rate of melon seedlings under saline stress. Ciênc. Rural 2019, 49, 7. [Google Scholar] [CrossRef]
- Kubala, S.; Garnczarska, M.; Wojtyla, Ł.; Clippe, A.; Kosmala, A.; Żmieńko, A.; Lutts, S.; Quinet, M. Deciphering priming-induced improvement of rapeseed (Brassica napus L.) germination through an integrated transcriptomic and proteomic approach. Plant Sci. 2015, 231, 94–113. [Google Scholar] [CrossRef]
- Mokhtari, K.H.; Hosseini, C.S.Z.; Kazemeini, S.A. Allelopathic Effects of Sorghum on Milk Thistle (Silybum marianum L.) Seed Germination and Growth. Res. Crop Ecophysiol. 2014, 9, 115–123. [Google Scholar]
- Khaliq, A.; Matloob, A.; Mahmood, S.; Wahid, A. Seed Pre-Treatments Help Improve Maize Performance Under Sorghum Allelopathic Stress. J. Crop. Improv. 2013, 27, 586–605. [Google Scholar] [CrossRef]
- Niu, G.; Xu, W.; Rodriguez, D.S.; Sun, Y. Growth and Physiological Responses of Maize and Sorghum Genotypes to Salt Stress. ISRN Agron. 2012, 2012, 1–12. [Google Scholar] [CrossRef]
- Shakeri, E.; Emam, Y.; Tabatabaei, S.; Sepaskhah, A. Evaluation of grain sorghum (Sorghum bicolor L.) lines/cultivars under salinity stress using tolerance indices. Int. J. Plant Prod. 2017, 11, 101–116. [Google Scholar]
- De Lacerda, C.F.; Cambraia, J.; Oliva, M.A.; Ruiz, H.A.; Prisco, J.T. Solute accumulation and distribution during shoot and leaf development in two sorghum genotypes under salt stress. Environ. Exp. Bot. 2003, 49, 107–120. [Google Scholar] [CrossRef]
- Ashraf, M. Some important physiological selection criteria for salt tolerance in plants. Flora Morphol. Distrib. Funct. Ecol. Plants 2004, 199, 361–376. [Google Scholar] [CrossRef]
- Abdelhamid, M.T.; El-Masry, R.R.; Darwish, D.S.; Abdalla, M.M.F.; Oba, S.; Ragab, R.; El Sabagh, A.; El Kholy, M.H.; Omer, E. Mechanisms of Seed Priming Involved in Salt Stress Amelioration. In Priming and Pretreatment of Seeds and Seedlings; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; pp. 219–251. [Google Scholar]
- Tahira, T.; Riaz, A.; Muhammad, F.; Basra, S.M.A. Improving salt tolerance in barley by osmopriming and biopriming. Int. J. Agric. Biol. 2018, 20, 2455–2464. [Google Scholar]
- Shah, T.; Latif, S.; Khan, H.; Munsif, F.; Nie, L. Ascorbic Acid Priming Enhances Seed Germination and Seedling Growth of Winter Wheat under Low Temperature Due to Late Sowing in Pakistan. Agronomy 2019, 9, 757. [Google Scholar] [CrossRef]
- Kaur, I.; Sharma, R. Allelopathic effect of Ageratum conyzoides on chlorophyll content in the leaves of mungbean. Int. J. Recent Sci. Res. 2016, 7, 13296–13298. [Google Scholar]
- Mansour, M.M.F.; Ali, E.F.; Salama, K.H.A. Does Seed Priming Play a Role in Regulating Reactive Oxygen Species Under Saline Conditions? React. Oxyg. Nitrogen Sulfur Species Plants 2019, 437–488. [Google Scholar] [CrossRef]
- Qian, H.; Xu, X.; Chen, W.; Jiang, H.; Jin, Y.; Liu, W.; Fu, Z. Allelochemical stress causes oxidative damage and inhibition of photosynthesis in Chlorella vulgaris. Chemosphere 2009, 75, 368–375. [Google Scholar] [CrossRef]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The role of Na+ and K+ transporters in salt stress adaptation in glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef]
- Rathod, G.R.; Anand, A. Effect of seed magneto-priming on growth, yield and Na/K ratio in wheat (Triticum aestivum L.) under salt stress. Indian J. Plant Physiol. 2016, 21, 15–22. [Google Scholar] [CrossRef]
- Rahneshan, Z.; Nasibi, F.; Moghadam, A.A. Effects of salinity stress on some growth, physiological, biochemical parameters and nutrients in two pistachio (Pistacia vera L.) rootstocks. J. Plant Interact. 2018, 13, 73–82. [Google Scholar] [CrossRef]
- Ryan, J.; Estefan, G.; Rashid, A. Soil and Plant Analysis Laboratory Manual, 2nd ed.; International Center for Agricultural Research in the Dry Areas (ICARDA): Aleppo, Syria, 2001; p. 172. [Google Scholar]
- Cheema, Z.; Khaliq, A. Use of sorghum allelopathic properties to control weeds in irrigated wheat in a semi arid region of Punjab. Agric. Ecosyst. Environ. 2000, 79, 105–112. [Google Scholar] [CrossRef]
- Rhoades, J.; Chanduvi, F.; Lesch, S. Soil Salinity Assessment: Methods and Interpretation of Electrical Conductivity Measurements. No. 57. Food and Agriculture Organization United Nation. 1999. Available online: http://www.fao.org/3/x2002e/x2002e.pdf (accessed on 8 April 2021).
- Association of Official Seed Analysts. Seed Vigor Testing Handbook. Contribution No. 32 to the Handbook on Seed Testing. In Association of Official Seed Analysis, East Lansing; Association of Official Seed Analysts: Ithaca, NY, USA, 1983; Available online: http://www.aosaseed.com/ (accessed on 8 April 2021).
- Bernfeld, P. Amylases, alpha and beta. Method Enzymol. 1955, 149–158. Available online: http://www.sciepub.com/reference/44942 (accessed on 8 April 2021).
- Arnon, D.I. Copper Enzymes in Isolated Chloroplasts Polyphenoloxidase in beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Chance, B.; Maehly, A. Assay of catalases and peroxidases. Methods Enzym. 1955, 2, 764–775. [Google Scholar] [CrossRef]
- Cakmak, I.; Horst, W.J. Effect of aluminium on lipid peroxidation, superoxide dismutase, catalase, and peroxidase activities in root tips of soybean (Glycine max). Physiol. Plant 1991, 83, 463–468. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Yoshida, S.; Forno, D.A.; Cock, J.H. Laboratory Manual for Physiological Studies of Rice; International Rice Research Institute: Los Baños, PH, USA, 1971; p. 61. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FEP | α-Amylase Activity | RL | SL | RDW | SDW | Chl a | Chl b | |
| Treatment (T) | 99.09 *** | 103.01 *** | 17.39 *** | 3.58 * | 7.27 * | 7.29 * | 7.40 ** | 10.24 *** |
| Salt stress (S) | 417.12 *** | 156.06 *** | 35.59 *** | 10.43 ** | 11.36 ** | 17.41 ** | 24.07 *** | 25.71 *** |
| T × S | 15.75 ** | 287.53 *** | 25.67 *** | 18.15 *** | 29.09 *** | 20.66 *** | 20.87 *** | 14.52 *** |
| H2O2 | MDA | CAT | SOD | POD | Na+ | K+ | ||
| Treatment (T) | 13.53 *** | 3281.34 *** | 9.95 ** | 16.70 *** | 7.58 * | 3.60 * | 18.26 ** | |
| Salt stress (S) | 4.42 * | 3200.15 *** | 34.11 *** | 40.68 *** | 25.54 *** | 2.75 * | 65.22 *** | |
| T × S | 18.35 *** | 1579.85 *** | 21.41 *** | 92.13 *** | 51.95 *** | 14.38 *** | 33.91 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, P.; He, L.; Abbas, A.; Hussain, S.; Hussain, S.; Du, D.; Hafeez, M.B.; Balooch, S.; Zahra, N.; Ren, X.; et al. Seed Priming with Sorghum Water Extract Improves the Performance of Camelina (Camelina sativa (L.) Crantz.) under Salt Stress. Plants 2021, 10, 749. https://doi.org/10.3390/plants10040749
Huang P, He L, Abbas A, Hussain S, Hussain S, Du D, Hafeez MB, Balooch S, Zahra N, Ren X, et al. Seed Priming with Sorghum Water Extract Improves the Performance of Camelina (Camelina sativa (L.) Crantz.) under Salt Stress. Plants. 2021; 10(4):749. https://doi.org/10.3390/plants10040749
Chicago/Turabian StyleHuang, Ping, Lili He, Adeel Abbas, Sadam Hussain, Saddam Hussain, Daolin Du, Muhammad Bilal Hafeez, Sidra Balooch, Noreen Zahra, Xiaolong Ren, and et al. 2021. "Seed Priming with Sorghum Water Extract Improves the Performance of Camelina (Camelina sativa (L.) Crantz.) under Salt Stress" Plants 10, no. 4: 749. https://doi.org/10.3390/plants10040749
APA StyleHuang, P., He, L., Abbas, A., Hussain, S., Hussain, S., Du, D., Hafeez, M. B., Balooch, S., Zahra, N., Ren, X., Rafiq, M., & Saqib, M. (2021). Seed Priming with Sorghum Water Extract Improves the Performance of Camelina (Camelina sativa (L.) Crantz.) under Salt Stress. Plants, 10(4), 749. https://doi.org/10.3390/plants10040749