
Mycobiota Associated with the Vascular Wilt of Poplar


 ,
,
Abstract
:1. Introduction
2. Materials and Metods
2.1. Site and Sampling
2.2. DNA Extraction, Amplification and Illumina Sequencing
2.3. Bioinformatics Analysis
2.4. Statistical Analyses
3. Results
4. Discussion
4.1. Disease Characteristics
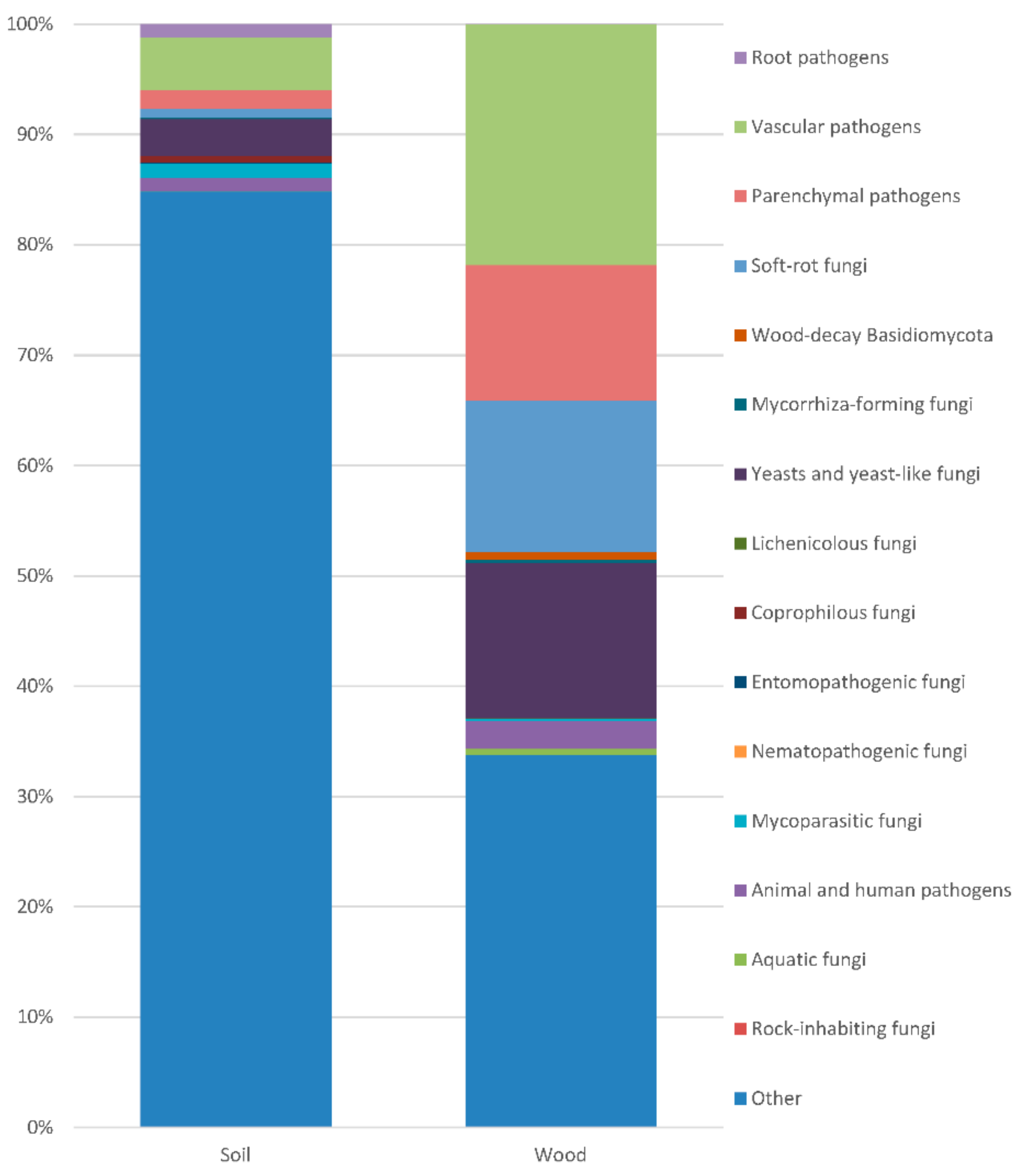
4.2. Pathogens in Diseased Poplar Trunk
4.3. Yeasts in Diseased Poplar Trunks
4.4. Mycorrhiza-Forming Fungi
4.5. The Endophytic State/Habit/Lifestyle of Fungi
4.6. Interactions among Fungi
4.7. Soil and Planting Material as the Source of the Inoculum
4.8. Colonization
4.9. Effects of Climate
4.10. Control and Mitigation
5. Conclusions
- 1.
- Populus hybrids may be subjected to various, thus far unidentified pathogenic agents.
- 2.
- New diseases may be asymptomatic, at least in the initial phase.
- 3.
- The indigenous microbiota can be involved in the development of the disease, but can also have an important role in limiting or preventing the development of pathogens.
- 4.
- The development of new diseases is related to climate change. It can lead to the near-total disappearance of some diseases, the sudden emergence of a new pathogens, or to the fungi already present becoming pathogenic.
- 5.
- Poplar wilt symptoms may be a consequence of various factors, the most important being climate and its effects on fungal development and the host–pathogen relationship.
- 6.
- Fungal diseases can spread from the soil or from introduced plant material, with the latter potentially introducing them into new areas.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Przybysz, K.; Przybysz, P. Poplar wood as a raw material for the paper industry in the twenty-first century. Ann. Warsaw Univ. Life Sci. 2013, 84, 56–59. [Google Scholar]
- Herve, C.; Ceulemans, R. Short-rotation coppiced vs. non-coppiced poplar: A comparative study at two different field sites. Biomass Bioenergy 1996, 11, 139–150. [Google Scholar] [CrossRef]
- Zajączkowski, K. Hodowla Lasu: Plantacje Drzew Szybko Rosnących; Powszechne Wydawnictwo Rolnicze i Leśne: Warszawa, Poland, 2013; pp. 1–168. [Google Scholar]
- Benetka, V.; Novotná, K.; Štochlová, P. Biomass production of Populus nigra L. clones grown in short rotation coppice systems in three different environments over four rotations. iFor. Biogeosci. For. 2014, 7, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Tigerstedt, P.; Haapanen, M. Growth and phenology of hybrid aspen clones (Populus tremula L. × Populus tremuloides Michx.). Silva Fenn. 2001, 35, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, P.-A.; Bálint, M.; Greshake, B.; Bandow, C.; Römbke, J.; Schmitt, I. Illumina metabarcoding of a soil fungal community. Soil Biol. Biochem. 2013, 65, 128–132. [Google Scholar] [CrossRef]
- Vilgalys, R.; Gonzalez, D. Organization of ribosomal DNA in the basidiomycete Thanatephorus praticola. Curr. Genet. 1990, 18, 277–280. [Google Scholar] [CrossRef]
- Gweon, H.S.; Oliver, A.; Taylor, J.; Booth, T.; Gibbs, M.; Read, D.S.; Griffiths, R.I.; Schonrogge, K. PIPITS: An automated pipeline for analyses of fungal internal transcribed spacer sequences from the Illumina sequencing platform. Methods Ecol. Evol. 2015, 6, 973–980. [Google Scholar] [CrossRef]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [Green Version]
- Bengtsson-Palme, J.; Ryberg, M.; Hartmann, M.; Branco, S.; Wang, Z.; Godhe, A.; De Wit, P.J.G.M.; Sánchez-García, M.; Ebersberger, I.; De Sousa, F.; et al. Improved software detection and extraction of ITS1 and ITS2 from ribosomal ITS sequences of fungi and other eukaryotes for analysis of environmental sequencing data. Methods Ecol. Evol. 2013, 4, 914–919. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian Classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; Mc Garrell, D.M.; Sun, Y.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magurran, A.E. Ecological Diversity and Its Measurement; Springer Science and Business Media, L.L.C.: Berlin/Heidelberg, Germany, 1988; pp. 1–180. [Google Scholar]
- Mugnai, L.; Graniti, A.; Surico, G. Esca (Black Measles) and Brown Wood-Streaking: Two old and elusive diseases of grapevines. Plant Dis. 1999, 83, 404–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlucci, A.; Lops, F.; Mostert, L.; Halleen, F.; Raimondo, M. Occurrence fungi causing black foot on young grapevines and nursery rootstock plants in Italy. Phytopathol. Mediterr. 2017, 56, 10–39. [Google Scholar]
- Larignon, P.; Fontaine, F.; Farine, S.; Clément, C. Esca et Black Dead Arm: Deux acteurs majeurs des mala-dies du bois chez la Vigne. Comptes Rendus Biol. 2009, 332, 765–783. [Google Scholar] [CrossRef] [PubMed]
- Van Niekerk, J.M.; Groenewald, J.Z.; Farr, D.F.; Fourie, P.H.; Halleen, F.; Crous, P.W. Reassessment of Phomopsis species on grapevine. Austral. Plant Pathol. 2005, 34, 27–39. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Leavitt, G.M.; Voegel, T.M.; Gubler, W.D. Identification and distribution of Botryosphaeria spp. associated with grapevine cankers in California. Plant Dis. 2006, 90, 1490–1503. [Google Scholar] [CrossRef] [Green Version]
- De Guido, M.A.; Pollastro, S.; Carlucci, A.; De Miccolis Angelini, R.M.; Faretra, F. Phomopsis viticola is easily transformed with Hph and Bmlr genes. J. Plant Pathol. 2013, 85, 43–52. [Google Scholar]
- Larignon, P.; Dubos, B. Fungi associated with esca disease in grapevine. Eur. J. Plant Pathol. 1997, 103, 147–157. [Google Scholar] [CrossRef]
- Arzanlou, M.; Narmani, A.; Moshari, S.; Khodaei, S.; Babai-Ahari, A. Truncatella angustata associated with grapevine trunk disease in northern Iran. Arch. Phytopathol. Plant Prot. 2013, 46, 1168–1181. [Google Scholar] [CrossRef]
- Bruno, G.; Sparapano, L. Effects of three esca-associated fungi on Vitis vinifera L.: V. Changes in the chemical and biological profile of xylem sap from diseased cv. Sangiovese vines. Physiol. Mol. Plant Pathol. 2007, 71, 210–229. [Google Scholar] [CrossRef]
- Gramaje, D.; Armengol, J.; Mohammadi, H.; Banihashemi, Z.; Mostert, L. Novel Phaeoacremonium species associated with Petri disease and esca of grapevine in Iran and Spain. Mycologia 2009, 101, 920–929. [Google Scholar] [CrossRef] [PubMed]
- Agusti-Brisach, C.; Armengol, J. Black-foot disease of grapevine: An update on taxonomy, epidemiology and management strategies. Phytopathol. Mediterr. 2013, 52, 245–261. [Google Scholar]
- Carlucci, A.; Cibelli, F.; Lops, F.; Phillips, A.; Ciccarone, C.; Raimondo, M. Pleurostomophora richardsiae asso-ciated with trunk diseases of grapevines in southern Italy. Phytopathol. Mediterr. 2015, 54, 109–123. [Google Scholar]
- Lombard, L.; Van Der Merwe, N.A.; Groenewald, J.Z.; Crous, P.W. Lineages in Nectriaceae: Re-evaluating the generic status of Ilyonectria and allied genera. Phytopathol. Mediterr. 2014, 53, 515–532. [Google Scholar]
- White, N.H.; Chilvers, G.A.; Evans, G. Antifungal activity of Cylindrocarpon radicicola Wr. Nat. Cell Biol. 1962, 195, 406–407. [Google Scholar] [CrossRef]
- Spies, C.; Moyo, P.; Halleen, F.; Mostert, L. Phaeoacremonium species diversity on woody hosts in the Western Cape Province of South Africa. Persoonia 2018, 40, 26–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabral, A.; Groenewald, J.Z.; Rego, C.; Oliveira, H.; Crous, P.W. Cylindrocarpon root rot: Multi-gene analysis reveals novel species within the Ilyonectria radicicola species complex. Mycol. Prog. 2011, 11, 655–688. [Google Scholar] [CrossRef] [Green Version]
- Liao, H.-L.; Bonito, G.; Rojas, J.A.; Hameed, K.; Wu, S.; Schadt, C.W.; Labbé, J.; Tuskan, G.A.; Martin, F.; Grigoriev, I.V.; et al. Fungal endophytes of Populus trichocarpa alter host phenotype, gene expression, and rhizobiome composition. Mol. Plant-Microbe Interact. 2019, 32, 853–864. [Google Scholar] [CrossRef]
- Prodi, A.; Sandalo, S.; Tonti, S.; Pisi, A. Phialophora-like fungi associated with kiwi fruit elephantiasis. J. Plan. Pathol. 2008, 90, 487–494. [Google Scholar]
- Diaz, B.; Murace, M.; Peri, P.; Keil, G.; Luna, L.; Otaño, M.Y. Natural and preservative-treated durability of Populus nigra cv Italica timber grown in Santa Cruz Province, Argentina. Int. Biodeterior. Biodegrad. 2003, 52, 43–47. [Google Scholar] [CrossRef]
- Xing, J.-Q.; Ikuo, M.; Wakako, O. Natural resistance of two plantation woods Populus × canadensis cv. and Cunninghamia lanceolata to decay fungi and termites. For. Stud. China 2005, 7, 36–39. [Google Scholar] [CrossRef]
- Del Frari, G.; Gobbi, A.; Aggerbeck, M.R.; Oliveira, H.; Hansen, L.H.; Ferreira, R.B. Characterization of the wood mycobiome of Vitis vinifera in a vineyard affected by esca. Spatial distribution of fungal communities and their putative relation with leaf symptoms. Front. Plant Sci. 2019, 10, 910. [Google Scholar] [CrossRef] [Green Version]
- Réblová, M.; Seifert, K.A. A new species of Chaetosphaeria with Menispora ciliata and phialophora-like anamorphs. Fungal Divers. 2008, 29, 99–105. [Google Scholar]
- Corda, A.K.J. Leones fungorum hucusque cognitorum. Praha Tomus 1837, 1, 1–32. [Google Scholar]
- Pirozynski, K.A. Xenotypa Petrak and Graphostroma gen. nov., segregates from Diatrypaceae. Can. J. Bot. 1974, 52, 2129–2135. [Google Scholar] [CrossRef]
- Zhao, G.; Liu, X.; Wu, W. Helicosporous hyphomycetes from China. Fungal Divers. 2007, 26, 313–524. [Google Scholar]
- Choi, Y.W. A Novel Helicosporium isolate and its antimicrobial and cytotoxic pigment. J. Microbiol. Biotechnol. 2012, 22, 1214–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hengstmengel, J. Notes on Hymenoscyphus—II. On three non-fructicolous species of the ‘fructi-genus-group’ with croziers. Persoonia 1996, 16, 191–207. [Google Scholar]
- Kowalski, T. Chalara fraxinea sp. nov. associated with dieback of ash (Fraxinus excelsior) in Poland. For. Pathol. 2006, 36, 264–270. [Google Scholar] [CrossRef]
- Kowalski, T.; Holdenrieder, O. The teleomorph of Chalara fraxinea, the causal agent of ash dieback. For. Pathol. 2009, 39, 304–308. [Google Scholar] [CrossRef]
- Réblová, M.; Gams, W.; Štěpánek, V. The new hyphomycete genera Brachyalara and Infundichalara, the similar Exochalara and species of ‘Phialophora sect. Catenulatae’ (Leotiomycetes). Fungal Divers. 2011, 46, 67–86. [Google Scholar] [CrossRef]
- Behnke-Borowczyk, J.; Kwaśna, H.; Kulawinek, B. Fungi associated with Cyclaneusma needle cast in Scots pine in the west of Poland. For. Pathol. 2019, 49, e12487. [Google Scholar] [CrossRef]
- Hutchison, L.J.; Untereiner, W.A.; Hiratsuka, Y. Knufia cryptophialidica gen. et sp. nov., a Dematiaceous Hyphomycete isolated from black galls of trembling aspen (Populus tremuloides). Mycologia 1995, 87, 902. [Google Scholar] [CrossRef]
- Fernando, A.; Currah, R. A comparative study of the effects of the root endophytes Leptodontidium orchi-dicola and Phialocephala fortinii (Fungi Imperfecti) on the growth of some subalpine. Can. J. Bot. 1996, 74, 1071–1078. [Google Scholar] [CrossRef]
- Wit, M.; Sierota, Z.; Osako, T.; Mirzwa-Mróz, E.; Wakuliński, W. Fusarium spp. na nadziemnych organach zamierających dębów—Nowe zagrożenie? (Fusarium spp. on the above–ground organs of dying—A new threat?). Sylwan 2015, 159, 403–410. [Google Scholar]
- Chen, J.L.; Tzean, S.S. Megacapitula villosa gen. et sp. nov. from Taiwan. Mycol. Res. 1993, 97, 347–350. [Google Scholar] [CrossRef]
- Behnke-Borowczyk, J.; Kwaśna, H.; Kokot, K.; Hałuszczak, M.; Łakomy, P. Abundance and diversity of fungi in oak wood. Dendrobiology 2018, 80, 143–160. [Google Scholar] [CrossRef]
- Rooney, S.N.; Eskalen, A.; Gubler, W.D. Recovery of Phaeomoniella chlamydospora and Phaeoacremonium in-flatipes from soil and grapevine tissues. Phytopathol. Mediterr. 2001, 40, S351–S356. [Google Scholar]
- Ariyawansa, H.A.; Phukhamsakda, C.; Thambugala, K.M.; Bulgakov, T.S.; Wanasinghe, D.N.; Perera, R.H.; Mapook, A.; Camporesi, E.; Kang, J.-C.; Jones, E.B.G.; et al. Revision and phylogeny of Leptosphaeriaceae. Fungal Divers. 2015, 74, 19–51. [Google Scholar] [CrossRef]
- El-Demerdash, A.; El-Demerdash, A. Chemical diversity and biological activities of Phaeosphaeria fungi genus: A systematic review. J. Fungi 2018, 4, 130. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Ariyawansa, H.A.; Bhat, D.J.; Boonmee, S.; Maharachchikumbura, S.S.N.; McKenzie, E.H.C.; Phookamsak, R.; Phukhamsakda, C.; et al. Fungal diversity notes 1–110, Taxonomic and phylogenetic contributions to fungal species. Fungal Divers. 2015, 72, 1–197. [Google Scholar] [CrossRef]
- Gams, W.; Stielow, B.; Gräfenhan, T.; Schroers, H.-J. The ascomycete genus Niesslia and associated Monocillium-like anamorphs. Mycol. Prog. 2019, 18, 5–76. [Google Scholar] [CrossRef]
- Jankowiak, R.; Bilański, P.; Ostafińska, A.; Linnakoski, R. Ophiostomatales associated with wounds on hardwood trees in Poland. Plant Pathol. 2019, 68, 1407–1424. [Google Scholar] [CrossRef]
- Kendrick, W.B. The Leptographium complex Verticicladiella Hughes. Can. J. Bot. 1961, 39, 1079–1085. [Google Scholar] [CrossRef]
- Kwaśna, H.; Mazur, A.; Łabędzki, A.; Kuźmiński, R.; Łakomy, P. Zbiorowiska grzybów w rozkładającym się drewnie dębu i sosny (Communities of fungi in decomposed wood of oak and pine). For. Res. Pap. 2016, 77, 261–275. [Google Scholar]
- Sogonov, M.V.; Castlebury, L.A.; Rossman, A.Y.; Mejía, L.C.; White, J.F., Jr. Leaf-inhabiting genera of the Gnomoniaceae, Diaporthales. Stud. Mycol. 2008, 62, 1–79. [Google Scholar] [CrossRef]
- Tanney, J.; Seifert, K. Mollisiaceae: An overlooked lineage of diverse endophytes. Stud. Mycol. 2020, 95, 293–380. [Google Scholar] [CrossRef]
- Behnke-Borowczyk, J.; Kwaśna, H.; Kartawik, N.; Sijka, B.; Bełka, M.; Łakomy, P. Effect of management on fungal communities in dead wood of Scots pine. For. Ecol. Manag. 2021, 479, 118528. [Google Scholar] [CrossRef]
- Oren, Y.; Sadowsky, A.; Gefen, D.; Solel, Z.; Kimchy, M. Scytalidium wilt of citrus. Eur. J. Plant Pathol. 2001, 107, 467–470. [Google Scholar] [CrossRef]
- Machado, A.R.; Pinho, D.B.; De Oliveira, S.A.S.; Pereira, O.L. New occurrences of Botryosphaeriaceae causing black root rot of cassava in Brazil. Trop. Plant Pathol. 2014, 39, 464–470. [Google Scholar] [CrossRef] [Green Version]
- De Meyer, E.M.; De Beer, Z.W.; Summerbell, R.C.; Moharram, A.; De Hoog, G.S.; Vismer, H.F.; Wingfield, M.J. Taxonomy and phylogeny of new wood- and soil-inhabiting Sporothrix species in the Ophiostoma stenoceras-Sporothrix schenckii complex. Mycologia 2008, 100, 647–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrzosek, M.; Sierota, Z.; Sikora, K.; Małecka, M.; Pawłowska, J. Bogactwo grzybów zasiedlających drewno pniaków świerkowych po roku od sztucznego zakażenia Phlebiopsis gigantea. (The diversity of fungi present in the wood of spruce stumps one year after artificial infection by Phlebiopsis gigantea. Stud. Mater. CEPL 2014, 16, 202–211. [Google Scholar]
- Koukol, O.; Kubátová, A. New European records of basidiomycete Burgoa anomala from coniferous litter and sediment in underground tunnel. Czech Mycol. 2015, 67, 241–247. [Google Scholar] [CrossRef]
- Goh, T.K.; Hyde, K.D. A synopsis of Trichocladium species, based on the literature. Fungal Divers. 1999, 2, 101–118. [Google Scholar]
- Nagahama, T. Yeast biodiversity in freshwater, marine and deep-sea environments. In Biodiversity and ecophysiology of yeasts; The Yeast Handbook; Rosa, C.A., Peter, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 241–263. [Google Scholar]
- Alias, S.A.; Jones, E.B.G. Marine Fungi from Mangroves of Malaysia; Institute Ocean and Earth Sciences, University of Malaya: Kuala Lumpur, Malaysia, 2009; pp. 1–108. [Google Scholar]
- Pang, K.L.; Jheng, J.S.; Jones, E.B.G. Marine Mangrove Fungi of Taiwan; National Taiwan Ocean University Press: Keelung, Taiwan, 2011; pp. 1–131. [Google Scholar]
- El-Elimat, T.; Raja, H.A.; Figueroa, M.; Falkinham, J.O.; Oberlies, N.H. Isochromenones, isobenzofuranone, and tetrahydronaphthalenes produced by Paraphoma radicina, a fungus isolated from a freshwater habitat. Phytochemistry 2014, 104, 114–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafellner, J. Distributional and other data for some species of Didymocyrtis (Dothideomycetes, Pleo-sporales, Phaeosphaeriaceae), including their Phoma-type anamorphs. Fritschiana 2015, 80, 43–88. [Google Scholar]
- Tibell, S.; Tibell, L.; Pang, K.-L.; Jones, E.G. A conspectus of the filamentous marine fungi of Sweden. Bot. Mar. 2019, 63, 141–153. [Google Scholar] [CrossRef]
- Kwaśna, H. Zdrowotność topoli na plantacjach w strefach ochronnych hut miedzi Legnica i Głogów. (Health of poplars in plantations in the sanitary protection zones of Legnica and Glogów copper mills). Sylwan 2017, 161, 639–647. [Google Scholar]
- Cellerino, G.P. Review of Fungal Diseases in Poplar; Food and Agriculture Organization of the United Nations: Rome, Italy, 1999. [Google Scholar]
- Kepley, J.B.; Jacobi, W.R. Pathogenicity of Cytospora fungi on six hardwood species. J. Arboric. 2000, 26, 326–332. [Google Scholar]
- Rosskopf, E.N.; Charudattan, R.; DeValerio, J.T.; Stall, W.M. Field evaluation of Phomopsis amaranthicola, a biological control agent of Amaranthus spp. Plant Dis. 2000, 84, 1225–1230. [Google Scholar] [CrossRef] [Green Version]
- Van Kan, J.A.L. Licensed to kill: The lifestyle of a necrotrophic plant pathogen. Trends Plant Sci 2006, 11, 247–253. [Google Scholar] [CrossRef]
- Bussaban, B.; Lumyong, L.; Lumyong, P.; McKenzie, E.H.C.; Hyde, K.D. Endophytic fungi from Amomum siamense. Can. J. Microbiol. 2001, 47, 943–948. [Google Scholar] [CrossRef]
- Kumaresan, V.; Suryanarayanan, T.S. Endophyte assemblages in young, mature and senescent leaves of Rhizophora apiculata: Evidence for the role of endophytes in mangrove litter degradation. Fungal Divers. 2002, 9, 81–91. [Google Scholar]
- Osono, T.; Takeda, H. Comparison of litter decomposing ability among diverse fungi in cool temperate deciduous forest in Japan. Mycologia 2002, 94, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Suryanarayanan, T.S.; Murali, T.S.; Venkatesan, G. Occurrence and distribution of fungal endophytes in tropical forests across a rainfall gradient. Can. J. Bot. 2002, 80, 818–826. [Google Scholar] [CrossRef]
- Yanna Ho, W.H.; Hyde, K.D. Fungal succession on fronds of Phoenix hanceana in Hong Kong. Fungal Divers. 2002, 10, 185–211. [Google Scholar]
- Tomita, F. Endophytes in Southeast Asia and Japan: Their taxonomic diversity and potential applications. Fungal Divers. 2003, 14, 187–204. [Google Scholar]
- Murali, T.S.; Suryanarayanan, T.S.; Geeta, R. Endophytic Phomopsis species: Host range and implications for diversity estimates. Can. J. Microbiol. 2006, 52, 673–680. [Google Scholar] [CrossRef]
- Hyde, K.D.; Bussaban, B.; Paulus, B.; Crous, P.W.; Lee, S.; McKenzie, E.H.C.; Photita, W.; Lumyong, S. Biodiversity of saprobic fungi. Biodivers. Conserv. 2007, 16, 17–35. [Google Scholar]
- Rossman, A.Y.; Farr, D.F.; Castlebury, L.A. A review of the phylogeny and biology of the Diaporthales. Mycoscience 2007, 48, 135–144. [Google Scholar] [CrossRef]
- Promputtha, I.; Hyde, K.D.; McKenzie, E.H.C.; Peberdy, J.F.; Lumyong, S. Can leaf degrading enzymes provide evidence that endophytic fungi becoming saprobes? Fungal Divers. 2010, 41, 89–99. [Google Scholar] [CrossRef]
- Promputtha, I.; Lumyong, S.; Vijaykrishna, D.; McKenzie, E.H.C.; Hyde, K.D.; Jeewon, R. A phylogenetic evaluation of whether endophytes become saprotrophs at host senescence. Microb. Ecol. 2007, 53, 579. [Google Scholar] [CrossRef] [PubMed]
- Kodsueb, R.; McKenzie, E.H.C.; Lumyong, S.; Hyde, K.D. Diversity of saprobic fungi on Magnoliaceae. Fungal Divers. 2008, 30, 37–53. [Google Scholar]
- Kodsueb, R.; McKenzie, E.H.C.; Lumyong, S.; Hyde, K.D. Fungal succession on woody litter of Magnolia liliiflora (Magnoliaceae). Fungal Divers. 2008, 30, 55–72. [Google Scholar]
- Botella, L.; Diez, J.J. Phylogenic diversity of fungal endophytes in Spanish stands of Pinus halepensis. Fungal Divers. 2010, 47, 9–18. [Google Scholar] [CrossRef]
- González, V.; Tello, M.L. The endophytic mycota associated with Vitis vinifera in central Spain. Fungal Divers. 2011, 47, 29–42. [Google Scholar] [CrossRef]
- Dai, C.C.; Chen, Y.; Tian, L.; Sh, Y. Correlation between invasion by endophytic fungus Phomopsis sp. and enzyme production. Afr. J. Agric. Res. 2010, 5, 1324–1340. [Google Scholar]
- Ceriani-Nakamurakare, E.; Slodowicz, M.; Gonzalez-Audino, P.; Dolinko, A.; Carmarán, C. Mycobiota associated with the ambrosia beetle Megaplatypus mutatus: Threat to poplar plantations. Forests 2016, 89, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Goidànich, G. Schema di una classificazione delle Stilbaceaeche erano riunite fin’ora nel genere Graphium Corda. Ann. Bot. 1935, 21, 40–50. [Google Scholar]
- Upadhyay, H.P. A monograph of Ceratocyslis and Ceratocystiopsi; University of Georgia Press: Athens, GA, USA, 1981; pp. 1–176. [Google Scholar]
- Seifert, K.A.; Okada, G. Graphium anamorphs of Ophiostoma species and similar anamorphs of other as-comycetes. In Ceratocyslis and Ophiostoma: Taxonomy, Ecology, and Pathology; Wingfield, M.J., Seifert, K.A., Webber, J.F., Eds.; American Phytopathological Society Press: St. Paul, MN, USA, 1993; pp. 27–41. [Google Scholar]
- De Gruyter, J.; Woudenberg, J.H.; Aveskamp, M.M.; Verkley, G.J.; Groenewald, J.Z.; Crous, P.W. Systematic reappraisal of species in Phoma section Paraphoma, Pyrenochaeta and Pleurophoma. Mycologia 2010, 102, 1066–1081. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Zhou, R.; Yuan, Y.; Xu, H.; Fu, J.; Li, H. Identification and characterization of Paraphoma chrysanthemicola causing leaf spot disease on Atractylodes japonica in China. J. Phytopathol. 2016, 164, 372–377. [Google Scholar] [CrossRef]
- Dhillon, G.; Sandhu, J.S.; Singh, P. Variation among poplar (Populus deltoides Bartr.) clones for growth, wood traits and tolerance to leaf spot diseases. Curr. Agric. Res. J. 2020, 8, 128–136. [Google Scholar] [CrossRef]
- Yokoya, K.; Postel, S.; Fang, R.; Sarasan, V. Endophytic fungal diversity of Fragaria vesca, a crop wild relative of strawberry, along environmental gradients within a small geographical area. PeerJ 2017, 5, e2860. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Schumacher, R.K.; Wingfield, M.J.; Lombard, L.; Giraldo, A.; Christensen, M.; Gardiennet, A.; Nakashima, C.; Pereira, O.; Smith, A.J.; et al. Fungal systematics and evolution: FUSE 1. Sydowia 2015, 67, 81–118. [Google Scholar]
- Domsch, K.H.; Gams, W.; Anderson, T.-H. Compendium of Soil Fungi. TAXON 1982, 31, 600. [Google Scholar] [CrossRef]
- Hale, M.D.; Eaton, R.A. Oscillatory growth of fungal hyphae in wood cell walls. Trans. Br. Mycol. Soc. 1985, 84, 277–288. [Google Scholar] [CrossRef]
- Nilsson, T. Microscopic studies on the degradation of cellophane and various cellulosic fibres by wood-attacking microfungi. Stud. For. Suec. 1947, 117, 1–27. [Google Scholar]
- Hallaksela, A.M.; Niemistö, P. Stem discoloration of planted silver birch. Scand. J. For. Res. 2008, 13, 169–176. [Google Scholar] [CrossRef]
- Coetsee, C.; Wingfield, M.J.; Crous, P.W.; Wingfield, B.D. Xenochalara, a new genus of dematiaceous hypho-mycetes for Chalara-like fungi with apical wall building conidial development. S. Afr. J. Bot. 2000, 66, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, A.; Scorzetti, G.; Fell, J.W. Diversity in the yeast Cryptoccocus albidus and related species as re-vealed by ribosomal DNA sequence analysis. Can. J. Microbiol. 2000, 46, 7–27. [Google Scholar] [CrossRef]
- Cai, L.; Jeewon, R.; Hyde, K.; Hyde, R. Phylogenetic evaluation and taxonomic revision of Schizothecium based on ribosomal DNA and protein coding genes. Fungal Divers. 2005, 19, 1–21. [Google Scholar]
- Arenz, B.E.; Held, B.W.; Jurgens, J.A.; Farrell, R.L.; Blanchette, R.A. Fungal diversity in soils and historic wood from the Ross Sea Region of Antarctica. Soil Biol. Biochem. 2006, 38, 3057–3064. [Google Scholar] [CrossRef]
- McGovern, R.J.; Horita, H.; Stiles, C.M.; Seijo, T.E. Host range of Itersonilia perplexans and management of Itersonilia petal blight of China Aster. Plant Health Prog. 2006, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Aveskamp, M.M.; de Gruyter, J.; Woudenberg, J.H.C.; Verkley, G.J.M.; Crous, P.W. Highlights of the Didymellaceae: A polyphasic approach to characterise Phoma and related pleosporalean genera. Stud Mycol 2010, 65, 1–60. [Google Scholar] [CrossRef] [PubMed]
- Branda, E.; Turchetti, B.; Diolaiuti, G.; Pecci, M.; Smiraglia, C.; Buzzini, P. Yeast and yeast-like diversity in the southernmost glacier of Europe (Calderone Glacier, Apennines, Italy). FEMS Microbiol. Ecol. 2010, 72, 354–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crous, P.W.; Groenewald, J.Z.; Diederich, P. Xenopolyscytalum pinae. Fungal Planet 55. Persoonia 2010, 5, 130–131. [Google Scholar]
- Kowalski, T.; Kehr, R.D. Two new species of Phialocephala occurring on Picea and Alnus. Can. J. Bot. 1995, 73, 26–32. [Google Scholar] [CrossRef]
- Markovskaja, S. Aero-aquatic fungi colonizing decaying leaves in woodland swampy pools of Aukštadvaris Regional Park (Lithuania). Bot. Lithuania 2012, 18, 123–132. [Google Scholar]
- Savchenko, K.G.; Heluta, V.P. Smut fungi of Ukraine, a checklist. Sydowia 2012, 64, 281–300. [Google Scholar]
- Hyde, K.D.; Jones, E.B.G.; Liu, J.-K.; Ariyawansa, H.; Boehm, E.W.; Boonmee, S.; Braun, U.; Chomnunti, P.; Crous, P.W.; Dai, D.-Q.; et al. Families of Dothideomycetes. Fungal Divers. 2013, 63, 1–313. [Google Scholar] [CrossRef]
- Quaedvlieg, W.; Verkley, G.; Shin, H.-D.; Barreto, R.; Alfenas, A.; Swart, W.; Groenewald, J.; Crous, P. Sizing up Septoria. Stud. Mycol. 2013, 75, 307–390. [Google Scholar] [CrossRef] [Green Version]
- Marin-Felix, Y.; Groenewald, J.; Cai, L.; Chen, Q.; Marincowitz, S.; Barnes, I.; Bensch, K.; Braun, U.; Camporesi, E.; Damm, U.; et al. Genera of phytopathogenic fungi: GOPHY 1. Stud. Mycol. 2017, 86, 99–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valenzuela-Lopez, N.; Cano-Lira, J.; Guarro, J.; Sutton, D.; Wiederhold, N.; Crous, P.; Stchigel, A. Coelomycetous Dothideomycetes with emphasis on the families Cucurbitariaceae and Didymellaceae. Stud. Mycol. 2017, 90, 1–69. [Google Scholar] [CrossRef]
- Markovskaja, S.; Kačergius, A.; Davydenko, K.; Fraser, S. First record of Neocatenulostroma germanicum on pines in Lithuania and Ukraine and its co-occurrence with Dothistroma spp. and other pathogens. For. Pathol. 2016, 46, 522–533. [Google Scholar] [CrossRef]
- LoBuglio, K.F. Cenococcum. In Ectomycorrhizal Fungi Key Genera in Profile; Cairney, J.W.G., Chambers, S.M., Eds.; Springer: Berlin/Heidelberg, Germany, 1999. [Google Scholar] [CrossRef]
- Cole, G.T.; Kendrick, B. (Eds.) Biology of Conidial Fungi; Academic Press: New York, NY, USA, 1981; pp. 1–680. ISBN 978-0-12-179501-6. [Google Scholar]
- Kwaśna, H.; Behnke-Borowczyk, J.; Gornowicz, R.; Łakomy, P. Effects of preparation of clear-cut forest sites on the soil mycobiota with consequences for Scots pine growth and health. For. Pathol. 2019, 49, e12494. [Google Scholar] [CrossRef]
- Boberg, J.B.; Ihrmark, K.; Lindahl, B.D. Decomposing capacity of fungi commonly detected in Pinus sylvestris needle litter. Fungal. Ecol. 2011, 4, 110–114. [Google Scholar] [CrossRef]
- Gramaje, D.; Úrbez-Torres, J.R.; Sosnowski, M.R. Managing grapevine trunk diseases with respect to etiology and epidemiology: Current strategies and future prospects. Plant Dis. 2018, 102, 12–39. [Google Scholar] [CrossRef] [Green Version]
- Su, L.; Zhu, H.; Niu, Y.; Guo, Y.; Du, X.; Guo, J.; Zhang, L.; Qin, C. Phylogeny and taxonomic revision of Kernia and Acaulium. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Prabhugaonkar, A.; Jalmi, P. Isolation of Acrodontium crateriforme as a pitcher trap inquiline. Curr. Res. Environ. Appl. Mycol. 2017, 7, 203–207. [Google Scholar] [CrossRef]
- Ingold, C. Aquatic hyphomycetes of decaying alder leaves. Trans. Br. Mycol. Soc. 1942, 25, 339–417. [Google Scholar] [CrossRef]
- Wang, X.; Houbraken, J.; Groenewald, J.; Meijer, M.; Andersen, B.; Nielsen, K.; Crous, P.; Samson, R. Diversity and taxonomy of Chaetomium and chaetomium-like fungi from indoor environments. Stud. Mycol. 2016, 84, 145–224. [Google Scholar] [CrossRef] [Green Version]
- Kendrick, W.B.; Burges, A. Biological aspects of the decay of Pinus sylvestris leaf litter. Nova Hedwigia 1962, 4, 313–359. [Google Scholar]
- Martinović, T.; Koukol, O.; Hirose, D. Distinct phylogeographic structure recognized within Desmazierella acicola. Mycologia 2016, 108, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Sun, G.; Zhai, X.; Batzer, J.; Mayfield, D.; Crous, P.; Groenewald, J.; Gleason, M. Dissoconiaceae associated with sooty blotch and flyspeck on fruits in China and the United States. Pers. Mol. Phylogeny Evol. Fungi 2012, 28, 113–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boonmee, S.; Ko, T.W.K.; Chukeatirote, E.; Hyde, K.D.; Chen, H.; Cai, L.; McKenzie, E.H.; Jones, E.B.G.; Kodsueb, R.; Hassan, B.A. Two new Kirschsteiniothelia species with Dendryphiopsis anamorphs cluster in Kirschsteiniotheliaceae fam. nov. Mycologia 2012, 104, 698–714. [Google Scholar] [CrossRef]
- Abass, M.; Hussein, N. Morphological, molecular and pathological study on Nigrospora oryzae and Nigrospora sphaerica, the leaf spot fungi of date palm. Basra J. Date Palm Res. 2014, 13, 1313. [Google Scholar]
- Brentu, F.C.; Cornelius, E.W.; Lawson, L.E.V.; Oduro, K.A.; Vicent, A. First Report of Pseudocercospora angolensis causing fruit and leaf spot of Citrus in Ghana. Plant Dis. 2013, 97, 1661. [Google Scholar] [CrossRef]
- Nagahama, T.; Hamamoto, M.; Nakase, T.; Horikoshi, K. Rhodotorula lamellibrachii sp. nov., a new yeast species from a tubeworm collected at the deep-sea floor in Sagami bay and its phylogenetic analysis. Antonie Leeuwenhoek 2001, 80, 317–323. [Google Scholar] [CrossRef]
- Hamamoto, M. Erythrobasidium Hamamoto, Sugiyama & Komagata. In The Yeasts, a Taxonomic Study, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: London, UK, 2011; Volume 3, pp. 1433–1435. [Google Scholar]
- Kumar, D.; Sigler, L.; Gibas, C.F.C.; Mohan, S.; Schuh, A.; Medeiros, B.C.; Peckham, K.; Humar, A. Graphium basitruncatum fungemia in a patient with acute leukemia. J. Clin. Microbiol. 2007, 45, 1644–1647. [Google Scholar] [CrossRef] [Green Version]
- Lackner, M.; de Hoog, G.S. Parascedosporium and its relatives: Phylogeny and ecological trends. IMA Fungus 2011, 2, 39–48. [Google Scholar] [CrossRef]
- Milenković, I.; Keča, N.; Karadžić, D.; Radulović, Z.; Nowakowska, J.A.; Oszako, T.; Sikora, K.; Corcobado, T.; Jung, T. Isolation and pathogenicity of Phytophthora species from poplar plantations in Serbia. Forests 2018, 9, 330. [Google Scholar] [CrossRef] [Green Version]
- Cadete, R.M.; Lopes, M.R.; Rosa, C.A. Yeasts associated with decomposing plant material and rotting wood. In Yeasts in Natural Ecosystems: Diversity; Buzzini, P., Lachance, M.-A., Yurkov, A., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 265–292. [Google Scholar] [CrossRef]
- González, A.; Martínez, A.T.; Almendros, G.; Grinbergs, J. A study of yeasts during the delignification and fungal transformation of wood into cattle feed in Chilean rain forest. Antonie Leeuwenhoek 1989, 55, 221–236. [Google Scholar] [CrossRef] [PubMed]
- Hagler, A.N.; Mendonca-Hagler, L.C.; Phaff, H.J. Candida populi, a new species of yeast occurring in exudates of Populus and Betula species. Int. J. Syst. Bacteriol. 1989, 39, 97–99. [Google Scholar] [CrossRef] [Green Version]
- Selosse, M.A.; Martos, F.; Perry, B.; Maj, P.; Roy, M.; Pailler, T. Saprotrophic fungal symbionts in tropical achlorophyllous orchids. Plant Signal. Behav. 2010, 5, 349–353. [Google Scholar] [CrossRef] [Green Version]
- Hofstetter, V.; Buyck, B.; Croll, D.; Viret, O.; Couloux, A.; Gindro, K. What if esca disease of grapevine were not a fungal disease? Fungal Divers. 2012, 54, 51–67. [Google Scholar] [CrossRef] [Green Version]
- Shakya, M.; Gottel, N.; Castro, H.; Yang, Z.K.; Gunter, L.; Labbe, J.; Muchero, W.; Bonito, G.; Vilgalys, R.; Tuskan, G.; et al. A multifactor analysis of fungal and bacterial community structure in the root microbiome of mature Populus deltoides trees. PLoS ONE 2013, 8, e76382. [Google Scholar] [CrossRef]
- Cregger, M.A.; Veach, A.M.; Yang, Z.K.; Crouch, M.J.; Vilgalys, R.; Tuskan, G.A.; Schadt, C.W. The Populus holobiont: Dissecting the effects of plant niches and genotype on the microbiome. Microbiome 2018, 6, 31. [Google Scholar] [CrossRef]
- Ridout, M.; Newcombe, G. Sydowia polyspora is both a foliar endophyte and a pre-emergent seed pathogen in Pinus ponderosa. Plant Dis. 2018, 102, 640–644. [Google Scholar] [CrossRef] [Green Version]
- Hamayun, M.; Khan, S.A.; Khan, A.L.; Rehman, G.; Sohn, E.-Y.; Shah, A.A.; Kim, S.-K.; Joo, G.-J.; Lee, I.-J. Phoma herbarum as a new gibberellin-producing and plant growth-promoting fungus. J. Microbiol. Biotechnol. 2009, 19, 1244–1249. [Google Scholar]
- Da Silva, R.R.; Da Rosa, N.G.; De Oliveira, L.C.G.; Juliano, M.A.; Juliano, L.; Rosa, J.C.; Cabral, H. Biochemical properties and catalytic specificity of a novel neutral serine peptidase secreted by fungus Pyrenochaetopsis sp. Appl. Biochem. Biotechnol. 2018, 187, 1158–1172. [Google Scholar] [CrossRef]
- Ullah, C.; Unsicker, S.B.; Reichelt, M.; Gershenzon, J.; Hammerbacher, A. Accumulation of catechin and proanthocyanidins in black poplar stems after infection by Plectosphaerella populi: Hormonal regulation, biosynthesis and antifungal activity. Front. Plant Sci. 2019, 10, 1441. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, H.F.; Mohan, S.K. Compendium of Onion and Garlic Diseases and Pests, 2nd ed.; The American Phytopathological Society: St. Paul, MN, USA, 2016; pp. 8–86. [Google Scholar] [CrossRef]
- Larignon, P. Réflexions sur l’esca: Ce que l’on sait déjà montre qu’il en reste beaucoup à apprendre: Vigne. Phytoma-La Défense des Végétaux 2004, 576, 28–31. [Google Scholar]
- Fourie, P.H.; Halleen, F.; van der Vyver, J.; Schreuder, W. Effects of Trichoderma treatments on the occurrence of decline pathogens in the roots and rootstocks of nursery grapevines. Phytopathol. Mediterr. 2001, 40, 473–478. [Google Scholar]
- Gerbore, J. Lutte Biologique Contre un Champignon Pathogène Impliqué dans L’esca de la Vigne, par Utilisation de L’oomycète Pythium oligandrum. Ph.D. Thesis, Punjab Agricultural University, Punjab, India, 2013; pp. 1–270. [Google Scholar]
- Yacoub, A.; Gerbore, J.; Magnin, N.; Chambon, P.; Dufour, M.C.; Corio-Costet, M.F.; Guyoneaud, R.; Rey, P. Ability of Pythium oligandrum strains to protect Vitis vinifera against Phaeomoniella chlamydsopora, a pathogen involved in Esca, by inducing plant resistance. Biol. Control 2016, 92, 7–16. [Google Scholar]
- Yacoub, A.; Gerbore, J.; Magnin, N.; Vallance, J.; Grizard, D.; Guyoneaud, R.; Rey, P. Induction of grapevine defence systems using the oomycete Pythium oligandrum against a pathogenic fungus involved in Esca. Phytopathol. Mediterr. 2014, 53, 574–575. [Google Scholar]
- Bertsch, C.; Ramírez-Suero, M.; Magninrobert, M.; Larignon, P.; Chong, J.; Aboumansour, E.; Spagnolo, A.; Clément, C.; Fontaine, F. Grapevine trunk diseases: Complex and still poorly understood. Plant Pathol. 2012, 62, 243–265. [Google Scholar] [CrossRef] [Green Version]
- Jones, E.E.; Hammond, S.; Blond, C.; Brown, D.S.; Ridgway, H.J. Interaction between arbuscular mycorrhizal fungi and rootstock cultivar on the susceptibility to infection by Ilyonectria species. Phytopathol. Mediterr. 2014, 53, 582–583. [Google Scholar]
- Petit, E.; Gubler, W.D. Influence of Glomus intraradices on black foot disease caused by Cylindrocarpon macrodidymum on Vitis rupestris under controlled conditions. Plant Dis. 2006, 90, 1481–1484. [Google Scholar] [CrossRef] [Green Version]
- Bonito, G.; Reynolds, H.; Robeson, M.S.; Nelson, J.; Hodkinson, B.P.; Tuskan, G.; Schadt, C.W.; Vilgalys, R. Plant host and soil origin influence fungal and bacterial assemblages in the roots of woody plants. Mol. Ecol. 2014, 23, 3356–3370. [Google Scholar] [CrossRef]
- Zhu, Y.J.; Yu, X.Y.; Wang, B.T.; Jin, L.; Jin, F.J. Description of Fusarium solani isolated from the soil of a poplar plantation in China. Int. J. Agric. Biol. 2020, 24, 663–670. [Google Scholar]
- Crous, P.; Braun, U.; Schubert, K.; Groenewald, J. Delimiting Cladosporium from morphologically similar genera. Stud. Mycol. 2007, 58, 33–56. [Google Scholar] [CrossRef]
- Baral, H.-O.; De Sloover, J.R.; Huhtinen, S.; Laukka, T.; Stenroos, S. An emendation of the genus Hyaloscypha to include Fuscoscypha (Hyaloscyphaceae, Helotiales, Ascomycotina). Karstenia 2009, 49, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Van Niekerk, J.M.; Bester, W.; Halleen, F.; Crous, P.W.; Fourie, P.H. The distribution and symptomatology of grapevine trunk disease pathogens are influenced by climate. Phytopathol. Mediterr. 2011, 50, 98–111. [Google Scholar]
- Úrbez-Torres, J.R.; Battany, M.; Bettiga, L.J.; Gispert, C.; McGourty, G.; Roncoroni, J.; Smith, R.J.; Verdegaal, P.; Gubler, W.D. Botryosphaeriaceae species spore-trapping studies in California vineyards. Plant Dis. 2010, 94, 717–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Úrbez-Torres, J.; Bruez, E.; Hurtado, J.; Gubler, W.D. Effect of temperature on conidial germination of Botryosphaeriaceae species infecting grapevines. Plant Dis. 2010, 94, 1476–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Niekerk, J.M.; Calitz, F.J.; Halleen, F.; Fourie, P.H. Temporal spore dispersal patterns of grapevine trunk pathogens in South Africa. Eur. J. Plant Pathol. 2010, 127, 375–390. [Google Scholar] [CrossRef]
- Szwed, M. Variability of precipitation in Poland under climate change. Theor. Appl. Clim. 2018, 135, 1003–1015. [Google Scholar] [CrossRef] [Green Version]
- Gramaje, D.; Armengol, J. Fungal trunk pathogens in the grapevine propagation process: Potential inoculum sources, detection, identification, and management strategies. Plant Dis. 2011, 95, 1040–1055. [Google Scholar] [CrossRef] [Green Version]
- Úrbez-Torres, J.R.; Haag, P.; Bowen, P.; Lowery, T.; O’Gorman, D. Development of a DNA macroarray for the detection and identification of fungal pathogens causing decline of young grapevines. Phytopathology 2015, 105, 1373–1388. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
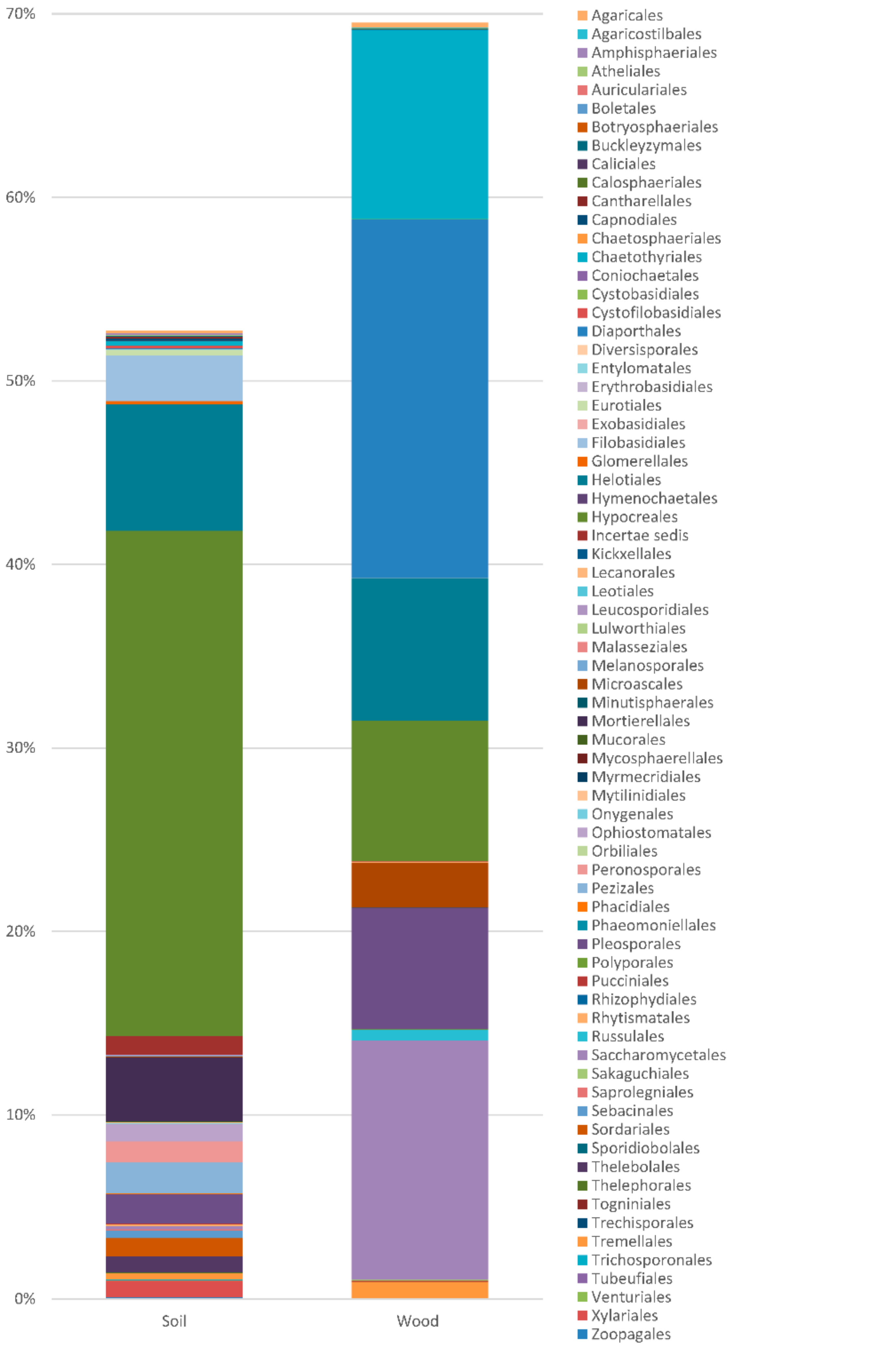
| No. | Taxon | Order | Soil | Wood | Trophic Group |
|---|---|---|---|---|---|
| Chromista | |||||
| Oomycota | |||||
| 1. | Aphanomyces spp. | Saprolegniales | 0.042 | Pathogens | |
| 2. | Elongisporangium anandrum (Drechsler) Uzuhasi, Tojo & Kakish | Peronosporales | 0.004 | Pathogen | |
| 3. | Globisporangium apiculatum (B. Paul) Uzuhashi, Tojo & Kakish. + G. heterothallicum W.A. Campb. & F.F. Hendrix + G. intermedium (de Bary) Uzuhashi, Tojo & Kakish. + G. macrosporum (Vaartaja & Plaäts-Nit.) Uzuhashi, Tojo & Kakish. + G. mamillatum (Meurs) Uzuhashi, Tojo & Kakish. + G. pleroticum (Takesi Itô) Uzuhashi, Tojo & Kakish. + G. sylvaticum (W.A. Campb. & F.F. Hendrix) Uzuhashi, Tojo & Kakish. + G. ultimum (Trow) Uzuhashi, Tojo & Kakish | Peronosporales | 1.010 | 0.001 | Pathogens |
| 4. | Hyaloperonospora cochleariae (Gäum.) Göker, Riethm., Voglmayr, Weiss & Oberw | Peronosporales | 0.017 | Pathogen | |
| 5. | Isoachlya intermedia (Coker & J.V. Harv.) Coker | Saprolegniales | 0.007 | Saprotroph | |
| 6. | Myzocytiopsis sp. | Peronosporales | 0.005 | Nematopathogenic | |
| 7. | Phytophthora brassicae De Cock & Man in ‘t Veld + P. citricola Sawada + P. clandestina P.A. Taylor, Pascoe & F.C. Greenh | Peronosporales | 0.040 | Pathogens | |
| 8. | Pythium conidiophorum Jokl. + P. oligandrum Drechsler + P. pachycaule Ali-Shtayeh + P. selbyi M.L. Ellis, Broders & Dorrance + P. vanterpoolii V. Kouyeas & H. Kouyeas + P. volutum Vanterp. & Truscott + Pythium spp. | Peronosporales | 0.053 | 0.001 | Pathogens |
| 9. | Thraustotheca clavata (de Bary) Humphrey | Saprolegniales | 0.021 | Saprotroph | |
| Frequency Oomycota | 1.199 | 0.002 | |||
| Number of taxa Oomycota | 26 | 2 | |||
| Fungi | |||||
| Blastocladiomycota | |||||
| Frequency Blastocladiomycota | 0.005 | ||||
| Number of taxa Blastocladiomycota | 1 | ||||
| Chytridiomycota | |||||
| 1. | Chytridiomycota | 0.004 | |||
| 2. | Rhizophydium sp. | Rhizophydiales | 0.004 | Pathogen | |
| Frequency Chytridiomycota | 0.008 | ||||
| Number of taxa Chytridiomycota | 2 | ||||
| Glomeromycota | |||||
| 1. | Entrophospora sp. | Diversisporales | 0.001 | ||
| Frequency Glomeromycota | 0.001 | Mycorrhizal | |||
| Number of taxa Glomeromycota | 2 | ||||
| Zygomycota | |||||
| 1. | Mortierella alpina Peyronel + M. amoeboidea W. Gams + M. antarctica Linnem. + M. elongata Linnem. + M. epicladia W. Gams & Emden + M. exigua Linnem. + M. fatshederae Linnem. + M. gamsii Milko + M. horticola Linnem. + M. humilis Linnem. + M. hyalina (Harz) W. Gams + Mortierella spp. | Mortierellales | 3.483 | 0.006 | Saprotrophs |
| 2. | Mortierellales | Mortierellales | 0.006 | ||
| 3. | Mucor racemosus Bull. | Mucorales | 0.012 | Saprotrophs | |
| 4. | Ramicandelaber sp. | Kickxellales | 0.004 | ||
| 5. | Rhizopus arrhizus A. Fisch. + R. oryzae Went & Prins. Geerl. | Mucorales | 0.019 | ||
| 6. | Syncephalis sp. | Zoopagales | 0.107 | Mycoparasite | |
| Frequency Zygomycota | 3.631 | 0.006 | |||
| Number of taxa Zygomycota | 18 | 3 | |||
| Ascomycota | |||||
| 1. | Acaulium retardatum (Udagawa & T. Muroi) Lei Su | Microascales | 0.004 | Saprotroph | |
| 2. | Acericola italica Wanas., Camporesi, E.B.G. Jones & K.D. Hyde | Pleosporales | 0.001 | ||
| 3. | Acremonium persicinum (Nicot) W. Gams + A. rutilum W. Gams | Hypocreales | 0.001 | 0.002 | Saprotrophs |
| 4. | Acrodontium crateriforme (J.F.H. Beyma) de Hoog | Incertae sedis | 0.013 | ||
| 5. | Alatospora acuminata Ingold + Alatospora sp. | Helotiales | 0.113 | 0.026 | |
| 6. | Alternaria alternata (Fr.) Keissl. + A. botrytis (Preuss) Woudenb. & Crous + A. infectoria E.G. Simmons + A. tenuissima (Kunze) Wiltshire + Alternaria sp. | Pleosporales | 0.065 | 0.039 | Pathogens |
| 7. | Amesia nigricolor (L.M. Ames) X. Wei Wang & Samson | Sordariales | 0.001 | Saprotroph | |
| 8. | Angustimassarina acerina Jayasiri, Thambug., R.K. Schumach. & K.D. Hyde + A. populi Thambug. & K.D. Hyde | Pleosporales | 0.354 | Mycoparasite | |
| 9. | Arthoniomycetes | 0.001 | 0.001 | ||
| 10. | Ascobolus sp. | Pezizales | 0.005 | Saprotroph, coprophilous | |
| 11. | Ascochyta skagwayensis (R. Sprague) Punith. | Pleosporales | 0.001 | Saprotroph, pathogen | |
| 12. | Ascomycete | 0.027 | |||
| 13. | Ascomycota | 1.123 | 0.215 | ||
| 14. | Aspergillus conicus Blochwitz + A. niger Tiegh. + A. penicillioides Speg. + A. versicolor (Vuill.) Tirab. | Eurotiales | 0.008 | 0.003 | Saprotrophs |
| 15. | Atrocalyx lignicola (Ying Zhang, J. Fourn. & K.D. Hyde) A. Hashim. & Kaz. Tanaka | Pleosporales | 0.009 | Saprotroph | |
| 16. | Aureobasidium melanogenum (Herm.-Nijh.) Zalar, Gostinčar & Gunde-Cim. + A. pullulans (de Bary & Löwenthal) G. Arnaud + Aureobasidium sp. | Dothideales | 0.003 | 0.013 | Saprotrophs, often aquatic |
| 17. | Bacidina sp. | Lecanorales | 0.018 | Lichenicolous | |
| 18. | Beauveria bassiana (Bals.-Criv.) Vuill. + Beauveria sp. | Hypocreales | 0.049 | 0.002 | Entomopathogenic |
| 19. | Blastobotrys malaysiensis Kurtzman + Blastobotrys sp. | Saccharomycetales | 0.009 | 0.013 | Saprotrophs |
| 20. | Boeremia exigua (Desm.) Aveskamp, Gruyter & Verkley + B. noackiana (Allesch.) Gruyter & Verkley | Pleosporales | 0.006 | 0.017 | Pathogens |
| 21. | Cadophora luteo-olivacea (J.F.H. Beyma) T.C. Harr. & McNew + C. spadicis Travadon, D.P. Lawr., Roon.-Lath., Gubler, W.F. Wilcox, Rolsh. & K. Baumgartner + Cadophora sp. | Helotiales | 0.114 | 1.435 | Pathogens |
| 22. | Candida sake (Saito & M. Ota) Uden & H.R. Buckley ex S.A. Mey. & Ahearn + C. subhashii M. Groenew., Sigler & S.E. Richardson + C. vartiovaarae (Capr.) Uden & H.R. Buckley + Candida sp. | Saccharomycetales | 0.093 | 0.012 | Saprotrophs |
| 23. | Capnobotryella renispora Sugiy | Capnodiales | 0.005 | Saprotroph | |
| 24. | Capnodiales | Capnodiales | 0.017 | ||
| 25. | Cenococcum geophilum Fr. | Mytilinidiales | 0.039 | Ectomycorrhizal | |
| 26. | Cephalothecaceae | Sordariales | 0.003 | Saprotrophs, mycoparasites | |
| 27. | Ceratostomataceae | Melanosporales | 0.004 | Saprotrophs, mycoparasite | |
| 28. | Cercophora sp. | Sordariales | 0.014 | Coprophilous | |
| 29. | Cercosporabeticola Sacc. | Capnodiales | 0.012 | Pathogen | |
| 30. | Chaetomiaceae | Sordariales | 0.085 | Saprotrophs | |
| 31. | Chaetomium globosum Kunze + Ch. piluliferum J. Daniels + Chaetomium sp. | Sordariales | 0.062 | 0.002 | Saprotrophs, endophytes |
| 32. | Chaetosphaeria vermicularioides (Sacc. & Roum.) W. Gams & Hol.-Jech. | Chaetosphaeriales | 0.005 | Saprotroph | |
| 33. | Chaetothyriales | Chaetothyriales | 0.104 | Parasites of humans and cold-blooded animals | |
| 34. | Chalara microspora (Corda) S. Hughes + Chalara sp. | Helotiales | 0.007 | 0.001 | Saprotroph |
| 35. | Chloridium paucisporum C.J.K. Wang & H.E. Wilcox | Helotiales | 0.001 | Ectendomycorrhizal | |
| 36. | Chrysosporium pseudomerdarium Oorschot | Onygenales | 0.004 | Endophyte | |
| 37. | Cistella albidolutea (Feltgen) Baral | Helotiales | 0.003 | Saprotroph | |
| 38. | Cladophialophora minutissima M.L. Davey & Currah + Cladophialophora sp. | Chaetothyriales | 0.002 | Saprotrophs, human pathogens | |
| 39. | Cladorrhinum flexuosum Madrid, Cano, Gené & Guarro | Sordariales | 0.008 | Saprotroph | |
| 40. | Cladosporium allicinum (Fr.) Bensch, U. Braun & Crous + C. cladosporioides (Fresen.) G.A. de Vries + C. colocasiae Sawada | Capnodiales | 0.096 | 0.015 | Saprotrophs, facultative plant pathogens, mycoparasites |
| 41. | Clonostachys divergens Schroers + C. parva (Schroers) Rossman, L. Lombard & Crous + C. rosea (Link) Schroers, Samuels + Clonostachys sp. | Hypocreales | 0.187 | 0.033 | Endophytes, mycoparasites |
| 42. | Coleophoma cylindrospora (Desm.) Höhn | Helotiales | 0.010 | Saprotroph | |
| 43. | Collophorina sp. | Leotiales | 0.001 | Saprotroph | |
| 44. | Coniochaetasp. | Coniochaetales | 0.015 | 0.002 | Pathogens, saprotrophs, endophytes, coprophilous, mycoparasite, human pathogens |
| 45. | Cordyceps bassiana Z.Z. Li, C.R. Li, B. Huang & M.Z. Fan + C. brongniartii Shimazu | Hypocreales | 0.047 | Enthomopathogenic, mycoparasite | |
| 46. | Cosmospora berkeleyana (P. Karst.) Gräfenhan, Seifert & Schroers | Hypocreales | 0.027 | Saprotroph, pathogen, mycoparasite | |
| 47. | Crocicreas sp. | Helotiales | 0.005 | Saprotrophs | |
| 48. | Cucurbitariaceae | Pleosporales | 0.076 | Saprotrophs, pathogens | |
| 49. | Cudoniella indica J. Webster, Eicker & Spooner | Helotiales | 0.002 | Saprotroph | |
| 50. | Cyathicula cyathoidea (Bull.) Thüm | Helotiales | 0.006 | Saprotrophs | |
| 51. | Cyphellophora sessilis (de Hoog) Réblová & Unter | Chaetothyriales | 0.001 | Pathogen | |
| 52. | Cytospora davidiana Y.L. Wang & X.Y. Zhang + C. leucostoma (Pers.) Sacc. + C. paratranslucens Norphanph., Bulgakov, T.C. Wen & K.D. Hyde + Cytospora sp. | Diaporthales | 0.012 | 13.720 | Pathogens |
| 53. | Dactylaria dimorphospora Veenb.-Rijks | Helotiales | 0.016 | Saprotroph | |
| 54. | Dactylonectria torresensis (A. Cabral, Rego & Crous) L. Lombard & Crous | Hypocreales | 0.008 | Pathogen | |
| 55. | Debaryomyces hansenii (Zopf) Lodder & Kreger-van Rij | Saccharomycetales | 0.023 | Pathogen | |
| 56. | Dendryphion europaeum Crous & R.K. Schumach. + D. nanum (Nees) S. Hughes | Pleosporales | 0.268 | 0.006 | Saprotroph |
| 57. | Dermateaceae | Helotiales | 0.002 | ||
| 58. | Desmazierella acicola Lib. | Pezizales | 0.001 | Saprotroph | |
| 59. | Diaporthe cynaroidis Marinc., M.J. Wingf. & Crous + D. foeniculina (Sacc.) Udayanga & Castl. + D. helicis Niessl + D. novem J.M. Santos, Vrandečić & A.J.L. Phillips + D. rudis (Fr.) Nitschke + Diaporthe sp. | Diaporthales | 0.017 | 3.327 | Pathogens, endophytes |
| 60. | Didymella macrostoma (Mont.) Qian Chen & L. C + D. pedeiae (Aveskamp, Gruyter & Verkley) Qian Chen & L. Cai + D. pinodes (Berk. & A. Bloxam) Petr. + D. pomorum (Thüm.) Qian Chen & L. Cai | Pleosporales | 0.039 | 0.036 | Pathogens |
| 61. | Didymosphaeria futilis (Berk. & Broome) Rehm | Pleosporales | 0.005 | Saprotroph | |
| 62. | Dissoconium eucalypti Crous & Carnegie | Capnodiales | 0.001 | Commensalist, mycoparasite | |
| 63. | Dothideomycetes | 0.018 | 0.014 | ||
| 64. | Emericellopsis glabra (J.F.H. Beyma) Backus & Orpurt + E. minima Stolk | Hypocreales | 0.179 | Endophytes | |
| 65. | Endophoma elongata Tsuneda & M.L. Dave | Incertae sedis | 0.005 | ||
| 66. | Epicoccum nigrum Link | Pleosporales | 0.002 | 0.001 | Endophyte, saprotroph, pathogen |
| 67. | Eurotiales | Eurotiales | 0.001 | ||
| 68. | Eurotiomycetes | 0.002 | 0.020 | ||
| 69. | Exophiala capensis Crous + E. equina (Pollacci) de Hoog, V.A. Vicente, Najafz., Harrak, Badali & Seyedm. + E. opportunistica de Hoog, V.A. Vicente, Najafz., Harrak, Badali & Seyedm. + Exophiala sp. | Chaetothyriales | 0.129 | 0.031 | Saprotrophs, human pathogens |
| 70. | Fusarium avenaceum (Fr.) Sacc. + F. equiseti (Corda) Sacc. + F. fujikuroi Nirenberg + F. oxysporum Schltdl. + F. petersiae L. Lombard + F. redolens Wollenw. + F. solani (Mart.) Sacc. + F. torulosum (Berk. & M.A. Curtis) Gruyter & J.H.M. Schneid. + Fusarium sp. + Neocosmospora solani (Mart.) L. Lombard & Crous | Hypocreales | 0.890 | 0.104 | Pathogens |
| 71. | Fusicolla aquaeductuum (Radlk. & Rabenh.) Gräfenhan, Seifert & Schroers + F. merismoides (Corda) Gräfenhan, Seifert & Schroers | Hypocreales | 0.096 | Pathogens | |
| 72. | Gibellulopsis nigrescens (Pethybr.) Zare, W. Gams & Summerb | Glomerellales | 0.009 | Saprotroph | |
| 73. | Gliomastix murorum var. felina (Marchal) S. Hughes | Hypocreales | 0.023 | Saprotroph | |
| 74. | Graphium basitruncatum (Matsush.) Seifert & G.Okada + G. penicillioides Corda | Microascales | 0.007 | 2.451 | Saprotrophs, plant and human pathogens |
| 75. | Gaphostroma platystomum (Schwein.) Piroz. | Xylariales | 0.004 | Saprotroph | |
| 76. | Halenospora varia (Anastasiou) E.B.G. Jones + Halenospora sp. | Helotiales | 0.443 | Saprotrophs, aquatic | |
| 77. | Halokirschsteiniothelia maritima (Linder) Boonmee & K.D. Hyde | Mytilinidiales | 0.023 | Saprotroph | |
| 78. | Halosphaeria quadri-remis (Höhnk) Kohlm | Microascales | 0.007 | Saprotroph | |
| 79. | Halosphaeriaceae | Microascales | 0.008 | ||
| 80. | Harzia acremonioides (Harz) Costantin + H. sphaerospora (Matsush.) D.W. Li & N.P. Schultes | Melanosporales | 0.028 | Saprotrophs | |
| 81. | Helicodendron luteoalbum Glen Bott + H. westerdijkiae Beverw | Helotiales | 0.009 | Saprotrophs | |
| 82. | Helicosporium sp. | Tubeufiales | 0.006 | Saprotrophs | |
| 83. | Helotiaceae | Helotiales | 0.005 | ||
| 84. | Helotiales | Helotiales | 3.087 | 4.565 | |
| 85. | Hemibeltrania sp. | Amphisphaeriales | 0.007 | Pathogen | |
| 86. | Herpotrichia pinetorum (Fuckel) G. Winter + Herpotrichia sp. | Pleosporales | 0.183 | 0.002 | Pathogens |
| 87. | Herpotrichiellaceae | Chaetothyriales | 0.004 | ||
| 88. | Hyalodendriella betulae Crous | Helotiales | 0.012 | 0.001 | Saprotroph, pathogen |
| 89. | Hyalopeziza sp. | Helotiales | 0.014 | Saprotroph | |
| 90. | Hyaloscypha bicolor (Hambl. & Sigler) Vohník, Fehrer & Réblová | Helotiales | 0.012 | Endophyte, saprotroph | |
| 91. | Hyaloscyphaceae | Helotiales | 0.003 | 0.040 | |
| 92. | Hymenoscyphus caudatus (P. Karst.) Dennis + H. imberbis (Bull.) Dennis | Helotiales | 0.007 | 0.017 | Pathogens, saprotrophs |
| 93. | Hypocreales | Hypocreales | 2.979 | ||
| 94. | Hypoxylon fragiforme (Pers.) J. Kickx f. | Xylariales | 0.469 | 0.002 | Saprotroph, pathogen |
| 95. | Ilyonectria crassa (Wollenw.) A. Cabral & Crous + I. cyclaminicola A. Cabral & Crous + I. destructans (Zinssm.) Rossman, L. Lombard & Crous + I. europaea A. Cabral, Rego & Crous + I. mors-panacis (A.A. Hildebr.) A. Cabral & Crous + I. robusta (A.A. Hildebr.) A. Cabral & Crous + Ilyonectria sp. + Cylindrocarpon sp. | Hypocreales | 2.031 | 6.710 | Saprotrophs, pathogens |
| 96. | Infundichalara microchona (W. Gams) Réblová & W. Gams + I. minuta Koukol | Helotiales | 0.014 | 0.001 | Saprotrophs, patogens, mycoparasitic |
| 97. | Jattaea taediosa (Sacc.) Réblová & Jaklitsch | Calosphaeriales | 0.005 | Endophyte | |
| 98. | Juxtiphoma eupyrena Sacc. | Pleosporales | 0.001 | Pathogen | |
| 99. | Knufia cryptophialidica L.J. Hutchison & Unter. + K. peltigerae (Fuckel) Réblová & Unter | Incertae sedis | 0.006 | 0.015 | Pathogens, lichenicolous |
| 100. | Lambertella tubulosa Abdullah & J. Webster | Helotiales | 1.445 | Saprotroph | |
| 101. | Lasiosphaeriaceae | Sordariales | 0.095 | 0.005 | |
| 102. | Lecania cyrtella (Ach.) Th. Fr. + L. naegelii (Hepp) Diederich & van den Boom | Lecanorales | 0.001 | 0.034 | Lichenicolous |
| 103. | Lecanorales | Lecanorales | 0.001 | ||
| 104. | Lemonniera terrestris Tubaki | Helotiales | 0.014 | Saprotroph, aquatic | |
| 105. | Leohumicola minima (de Hoog & Grinb.) Seifert & Hambl | Helotiales | 0.002 | Saprotroph | |
| 106. | Leotiomycetes | 0.003 | 0.876 | ||
| 107. | Lepraria caesiella R.C. Harris | Lecanorales | 0.002 | Lichenicolous | |
| 108. | Leptodontidium sp. | Helotiales | 0.011 | 0.254 | Endophyte, mycorrhizal |
| 109. | Leptosphaeria sp. | Pleosporales | 0.023 | Endophytes, saprotrophs, pathogens | |
| 110. | Leptosphaerulina australis McAlpine | Pleosporales | 0.014 | Endophyte | |
| 111. | Lophiostoma corticola (Fuckel) E.C.Y. Liew, Aptroot & K.D. Hyde + Lophiostoma sp. | Pleosporales | 0.788 | Pathogens | |
| 112. | Lophodermium pinastri (Schrad.) Chevall. + L. seditiosum Minter, Staley & Millar + Lophodermium sp. | Rhytismatales | 0.107 | 0.003 | Pathogens |
| 113. | Lophotrichus sp. | Microascales | 0.017 | Patogen, coprophilus, human pathogen | |
| 114. | Macroconia sphaeriae (Fuckel) Gräfenhan & Schroers | Hypocreales | 0.013 | Saprotroph, mycoparasitic | |
| 115. | Magnohelicospora fuscospora (Linder) R.F. Castañeda, Hern.-Restr. & Gené | Incertae sedis | 0.269 | Saprotroph | |
| 116. | Massarina sp. | Pleosporales | 0.002 | Saprotroph | |
| 117. | Megacapitula villosa J.L. Chen & Tzean | Incertae sedis | 0.001 | Saprotroph | |
| 118. | Melanospora kurssanoviana (Beliakova) Czerepan | Melanosporales | 0.009 | Saprotroph, mycoparasitic | |
| 119. | Metarhizium marquandii (Massee) Kepler, S.A. Rehner & Humber | Hypocreales | 0.495 | Endophyte | |
| 120. | Meyerozyma guilliermondii (Wick.) Kurtzman & M. Suzuki | Saccharomycetales | 0.003 | 0.022 | Coprophilous, human pathogen |
| 121. | Micarea adnata Coppins | Lecanorales | 0.006 | Lichenicolous | |
| 122. | Microascaceae | Microascales | 0.002 | ||
| 123. | Microdochium sp. | Amphisphaeriales | 0.063 | 0.001 | Pathogen |
| 124. | Microthecium fimicola (E.C. Hansen) Y. Marín, Stchigel, Guarro & Cano + M. quadrangulare (Dania García, Stchigel & Guarro) Y. Marín, Stchigel, Guarro & Cano | Melanosporales | 0.012 | 0.002 | Saprotrophs |
| 125. | Minutisphaera parafimbriatispora Raja, Oberlies, Shearer & A.N. Mill | Minutisphaerales | 0.017 | Saprotroph, aquatic | |
| 126. | Mollisia sp. | Helotiales | 0.021 | Saprotroph | |
| 127. | Monographella nivalis (Schaffnit) E. Müll | Amphisphaeriales | 0.004 | Pathogen | |
| 128. | Montagnulaceae | Pleosporales | 0.005 | Saprotrophs, endophytes, pathogens | |
| 129. | Mycofalcella calcarata Marvanová, Om-Kalth. & J. Webster | Helotiales | 0.002 | Saprotroph, aquatic | |
| 130. | Mycosphaerella tassiana (De Not.) Johanson | Capnodiales | 0.008 | Pathogen, saprotroph | |
| 131. | Myrmecridium schulzeri (Sacc.) Arzanlou, W. Gams & Crous | Myrmecridiales | 0.010 | Saprotroph | |
| 132. | Naevala perexigua (Roberge ex Desm.) K. Holm & L. Holm | Helotiales | 0.001 | Saprotroph | |
| 133. | Nakazawaea anatomiae (Zwillenb.) Kurtzman & Robnett + N. populi (Hagler, Mend.-Hagler & Phaff) Kurtzman & Robnett | Saccharomycetales | 0.016 | 12.941 | Saprotrophs |
| 134. | Nectria sp. | Hypocreales | 0.032 | Pathogens, saprotrophs | |
| 135. | Nectriaceae | Hypocreales | 0.432 | ||
| 136. | Neoascochytaexitialis (Morini) Qian Chen & L. Cai | Pleosporales | 0.012 | Pathogen | |
| 137. | Neobulgaria premnophila Roll-Hansen & H. Roll-Hansen + N. pura (Pers.) Petr. + Neobulgaria sp. | Helotiales | 0.684 | Saprotrophs | |
| 138. | Neocatenulostroma germanicum (Crous & U. Braun) Quaedvl. & Crous | Capnodiales | 0.001 | Pathogen | |
| 139. | Neocucurbitaria cava (Schulzer) Gruyter, Aveskamp & Verkley | Pleosporales | 0.002 | Saprotroph | |
| 140. | Neofabraea perennans Kienholz | Helotiales | 0.009 | Pathogen | |
| 141. | Neoleptosphaeria rubefaciens (Togliani) Gruyter, Aveskamp & Verkley | Pleosporales | 0.003 | Pathogen | |
| 142. | Neonectria candida (Ehrenb.) Rossman, L. Lombard & Crous + Neonectria sp. | Hypocreales | 0.560 | 0.763 | Pathogen |
| 143. | Neopyrenochaeta acicola ((Moug. & Lév.) Valenz.-Lopez, Crous, Stchigel, Guarro & Cano + N. inflorescentiae (Crous, Marinc. & M.J. Wingf.) Valenz.-Lopez, Crous, Stchigel, Guarro & Cano | Pleosporales | 0.014 | 0.058 | Pathogens, saprotrophs |
| 144. | Neosetophoma clematidis Wijayaw., Camporesi & K.D. Hyde | Pleosporales | 0.046 | Saprotroph | |
| 145. | Neurospora terricola Goch. & Backus | Sordariales | 0.004 | Saprotroph | |
| 146. | Niesslia mucida (W. Gams) W. Gams & Stielow | Hypocreales | 0.004 | Saprotroph | |
| 147. | Nigrograna mycophila Jaklitsch, Friebes & Voglmayr | Pleosporales | 0.007 | Saprotroph, mycoparasitic | |
| 148. | Nigrospora oryzae (Berk. & Broome) Petch | Incertae sedis | 0.535 | Saprotroph, pathogen | |
| 149. | Ochrocladosporium elatum (Harz) Crous & U. Braun | Pleosporales | 0.022 | 0.084 | Endophyte |
| 150. | Oedocephalum nayoroense Ts. Watan | Pezizales | 0.049 | Saprotroph | |
| 151. | Onygenales | Onygenales | 0.005 | ||
| 152. | Ophiostomataceae | Ophiostomatales | 0.790 | Pathogens | |
| 153. | Orbilia auricolor (A. Bloxam) Sacc. | Orbiliales | 0.026 | Saprotroph | |
| 154. | Orbiliaceae | Orbiliales | 0.006 | ||
| 155. | Pachyramichloridium pini (de Hoog & Rahman) C. Nakash., Videira & Crous | Capnodiales | 0.017 | Pathogen | |
| 156. | Papulaspora pisicola J.F.H. Beyma | Incertae sedis | 0.019 | Saprotroph | |
| 157. | Paraphoma chrysanthemicola (Hollós) Gruyter, Aveskamp & Verkley + P. radicina (McAlpine) Morgan-Jones & J.F. White + Paraphoma sp. | Pleosporales | 4.852 | Saprotrophs, pathogens | |
| 158. | Penicillium citreonigrum Dierckx + P. citreosulfuratum Biourge + P. georgiense S.W. Peterson & B.W. Horn + P. glandicola (Oudem.) Seifert & Samson + P. halotolerans Frisvad, Houbraken & Samson + P. lapidosum Raper & Fennell + P. nothofagi Houbraken, Frisvad & Samson + P. raphiae Houbraken, Frisvad & Samson + P. roseomaculatum Biourge + P. sacculum E. Dale + P. unicum Tzean, J.L. Chen & Shiu + P. virgatum Nirenberg & Kwaśna + Penicillium sp. + Talaromyces luteus C.R. Benj. | Eurotiales | 0.295 | 0.001 | Saprotrophs |
| 159. | Periconia sp. | Pleosporales | 0.012 | Endophyte | |
| 160. | Petriella sordida (Zukal) G.L. Barron & J.C. Gilman | Microascales | 0.001 | Coprophilous | |
| 161. | Phacidium lacerum Fr. + Phacidium sp. | Phacidiales | 0.027 | Saprotroph | |
| 162. | Phaeoacremonium cinereum Gramaje, Mohammadi, Banihash., Armengol & L. Mostert + P. hungaricum Essakhi, Mugnai, Surico & Crous | Togniniales | 0.044 | Pathogens | |
| 163. | Phaeoisaria loranthacearum Crous & R.K. Schumach. + P. sparsa B. Sutton | Xylariales | 0.347 | Saprotrophs, coprophilous | |
| 164. | Phaeomoniella sp. | Phaeomoniellales | 0.001 | ||
| 165. | Phaeosphaeria sp. | Pleosporales | 0.007 | Pathogens | |
| 166. | Phaeosphaeriaceae | Pleosporales | 0.013 | ||
| 167. | Phaeosphaeriopsis sp. | Pleosporales | 0.032 | Pathogens, saprotrophs | |
| 168. | Phialocephala sp. | Helotiales | 0.004 | Saprotrophs | |
| 169. | Phialophora sp. | Chaetothyriales | 10.291 | Saprotrophs, pathogens | |
| 170. | Phoma boeremae Gruyter + Phoma sp. | Pleosporales | 0.010 | 0.007 | Saprotrophs, pathogens |
| 171. | Phomopsis phaseoli (Desm.) Sacc. + P. velata (Sacc.) Traverso + Phomopsis sp. | Diaporthales | 1.186 | Pathogens, saprothrophs endophytes | |
| 172. | Physcia tenella (Scop.) DC. | Caliciales | 0.001 | Lichenicolous | |
| 173. | Pilophorus strumaticus Nyl. ex Cromb | Lecanorales | 0.001 | Lichenicolous | |
| 174. | Plagiostoma jonesii Senan. & K.D. Hyde | Diaporthales | 0.031 | Saprotroph, endophyte | |
| 175. | Plectosphaerella cucumerina (Lindf.) W. Gams + P. niemeijerarum L. Lombard | Glomerellales | 0.140 | 0.014 | Pathogens |
| 176. | Pleosporaceae | Pleosporales | 0.003 | ||
| 177. | Pleosporales | Pleosporales | 0.161 | 0.504 | |
| 178. | Pleotrichocladium opacum (Corda) Hern.-Restr., R.F. Castañeda & Gené | Pleosporales | 0.307 | 0.013 | Saprotroph, aquatic |
| 179. | Pleurophoma ossicola Crous, Krawczynski & H.-G. Wagner + Pleurophoma sp. | Xylariales | 0.016 | 0.005 | Saprotroph |
| 180. | Podospora appendiculata (Auersw. ex Niessl) Niessl + P. bulbillosa (W. Gams & Mouch.) X. Wei Wang & Houbraken. + P. leporina (Cain) Cain + Podospora sp. | Sordariales | 0.074 | Saprotroph, coprophilous | |
| 181. | Preussia flanaganii Boylan + P. typharum (Sacc.) Cain | Pleosporales | 0.058 | Saprotrophs, endophytes, coprophilous | |
| 182. | Pseudeurotium hygrophilum (Sogonov, W. Gams, Summerb. & Schroers) Minnis & D.L. Lindner + P. ovale Stolk + P. zonatum J.F.H. Beyma | Thelebolales | 0.804 | Saprotrophs, human pathogens | |
| 183. | Pseudocercospora angolensis (T. Carvalho & O. Mendes) Crous & U. Braun | Mycosphaerellales | 0.004 | Pathogen | |
| 184. | Pseudogymnoascus pannorum (Link) Minnis & D.L. Lindner + P. roseus Raillo | Thelebolales | 0.068 | Saprotrophs | |
| 185. | Pyrenochaeta sp. | Incertae sedis | 0.105 | 0.005 | Pathogen, saprotroph |
| 186. | Pyrenochaetopsis leptospora (Sacc. & Briard) Gruyter, Aveskamp & Verkley + P. microspora (Gruyter & Boerema) Gruyter, Aveskamp & Verkley | Pleosporales | 0.007 | 0.001 | Pathogens, saprotrophs, endophytes |
| 187. | Pyronemataceae | Pezizales | 0.081 | ||
| 188. | Saccharomyces cerevisiae (Desm.) Meyen | Saccharomycetales | 0.001 | Saprotroph | |
| 189. | Schizothecium glutinans (Cain) N. Lundq | Sordariales | 0.015 | Saprotroph, coprophilous | |
| 190. | Scolecobasidium constrictum E.V. Abbott + S. umbrinum (Ach.) Arnold | Incertae sedis | 0.016 | 0.002 | Saprotrophs, endophytes |
| 191. | Scutellinia scutellata (L.) Lambotte | Pezizales | 0.005 | Saprotroph | |
| 192. | Scytalidium lignicola Pesante + S. multiseptatum Hol.-Jech | Helotiales | 0.055 | 0.001 | Pathogens, saprotrophs, mycoparasitic |
| 193. | Sordariales | 0.008 | |||
| 194. | Sordariomycetes | 0.211 | 0.003 | ||
| 195. | Sphaeropsis sapinea (Fr.) Dyko & B. Sutton | Botryosphaeriales | 0.003 | Pathogen | |
| 196. | Sporormiaceae | Pleosporales | 0.003 | ||
| 197. | Sporothrix dentifunda Aghayeva & M.J. Wingf. + S. stenoceras (Robak) Z.W. de Beer, T.A. Duong & M.J. Wingf. + S. narcissi (Limber) Z.W. de Beer, T.A. Duong & M.J. Wingf | Ophiostomatales | 0.161 | 0.001 | Pathogens, saprotrophs |
| 198. | Stemphylium herbarum E.G. Simmons + S. majusculum E.G. Simmons + S. vesicarium (Wallr.) E.G. Simmons | Pleosporales | 0.027 | Pathogens | |
| 199. | Subramaniula flavipila X. Wei Wang & Samson | Sordariales | 0.014 | Saprotroph | |
| 200. | Sydowia polyspora (Bref. & Tavel) E. Müll | Dothideales | 0.004 | 1.028 | Pathogen, endophyte, saprotroph |
| 201. | Tetracladium furcatum Descals + T. setigerum (Grove) Ingold + Tetracladium sp. | Helotiales | 1.171 | 0.862 | Saprotrophs |
| 202. | Thelonectria blackeriella + T. olida (Wollenw.) Wollenw. + T. nodosa Salgado & P. Chaverri | Hypocreales | 0.012 | 0.006 | Pathogens |
| 203. | Tricharina sp. | Pezizales | 1.55 | Saprotrophs | |
| 204. | Trichocladium asperum Harz + T. griseum (Traaen) X. Wei Wang & Houbraken | Sordariales | 0.593 | Saprotrophs | |
| 205. | Trichoderma aerugineum Jaklitsch + T. hamatum (Bonord.) Bainier + T. koningiopsis Samuels, Carm. Suárez & H.C. Evans + T. martiale Samuels + T. neokoningii Samuels & Soberanis + T. piluliferum J. Webster & Rifai + T. Polysporum (Link) Rifai + T. pubescens Bissett + T. stilbohypoxyli Samuels & Schroers + T. viride Pers. + Trichoderma sp. | Hypocreales | 19.464 | 0.001 | Saprotrophs |
| 206. | Tricladium splendens Ingold | Helotiales | 0.040 | 0.057 | Saprotroph, acquatic |
| 207. | Truncatella an gustata (Pers.) S. Hughes + T. restionacearum S.J. Lee & Crous | Amphisphaeriales | 0.003 | 0.001 | Pathogens |
| 208. | Valsa malicola Z. Urb. + V. sordida Sacc. + V. leucostoma (Pers.) Fr. | Diaporthales | 0.012 | 0.214 | Pathogens |
| 209. | Valsaceae | Diaporthales | 0.003 | ||
| 210. | Venturia hystrioides (Dugan, R.G. Roberts & Hanlin) Crous & U. Braun | Venturiales | 0.018 | Pathogen | |
| 211. | Venturiaceae sp. | Venturiales | 0.001 | ||
| 212. | Verticillium dahliae Kleb. + V. longisporum (C. Stark) Karapapa, Bainbr. & Heale | Glomerellales | 0.029 | Pathogens, saprotrophs | |
| 213. | Volutella ciliata (Alb. & Schwein.) Fr. + Volutella sp. | Hypocreales | 0.009 | 0.009 | Saprotrophs, pathogen |
| 214. | Xanthoparmelia subchalybaeizans (Hale) G. Amo, A. Crespo, Elix & Lumbsch | Lecanorales | 0.005 | Lichenicolous | |
| 215. | Xenochalara sp. | Helotiales | 0.033 | Saprotroph | |
| 216. | Xenopolyscytalum pinea Crous + Xenopolyscytalum sp. | Helotiales | 0.001 | 0.001 | Saprotrophs |
| 217. | Xenoramularia arxii Videira, Crous & U. Braun | Capnodiales | 0.001 | Pathogen | |
| 218. | Xylariales | Xylariales | 0.061 | ||
| 219. | Yamadazyma mexicana (M. Miranda, Holzschu, Phaff & Starmer) Billon-Grand | Saccharomycetales | 0.039 | Saprotroph | |
| 220. | Yarrowia lipolytica (Wick., Kurtzman & Herman) Van der Walt & Arx | Saccharomycetales | 0.001 | Saprotroph | |
| 221. | Zalerion sp. | Lulworthiales | 0.001 | Saprotroph, aquatic | |
| 222. | Zopfiella marina Furuya & Udagawa + Z. pilifera Udagawa & Furuya | Sordariales | 0.027 | Saprotrophs, aquatic | |
| Frequency of Ascomycota | 45.299 | 68.697 | |||
| Number of taxa Ascomycota | 263 | 178 | |||
| Basidiomycota | |||||
| 1. | Aecidium sp. | Pucciniales | 0.034 | Pathogen | |
| 2. | Agaricales | 0.054 | |||
| 3. | Agaricomycetes | 0.008 | 0.074 | ||
| 4. | Agaricostilbomycetes | 0.001 | |||
| 5. | Apiotrichum dulcitum (Berkhout) Yurkov & Boekhout + A. gracile (Weigmann & A. Wolff) Yurkov & Boekhout | Trichosporonales | 0.047 | Saprotrophs | |
| 6. | Armillaria mellea (Vahl) P. Kumm | Agaricales | 0.025 | Pathogen | |
| 7. | Athelia acrospora Jülich | Atheliales | 0.001 | Saprotroph | |
| 8. | Atheliaceae | Atheliales | 0.023 | ||
| 9. | Aurantiporus fissilis (Berk. & M.A. Curtis) H. Jahn ex Ryvarden | Polyporales | 0.002 | Saprotroph, pathogen | |
| 10. | Auriculariales | 0.004 | |||
| 11. | Basidiomycota | 0.031 | 0.038 | ||
| 12. | Bensingtonia sp. | Agaricostilbales | 0.001 | Saprotroph | |
| 13. | Bjerkandera adusta (Willd.) P. Karst | Polyporales | 0.002 | Saprotroph, pathogen | |
| 14. | Buckleyzyma aurantiaca (Saito) Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout | Buckleyzymales | 0.048 | 0.007 | Saprotroph |
| 15. | Bullera crocea Buhagiar | Tremellales | 0.008 | 0.001 | Saprotroph |
| 16. | Bulleromyces albus Boekhout & Á. Fonseca | Tremellales | 0.001 | 0.001 | Saprotroph |
| 17. | Burgoa anomala (Hotson) Goid | Cantharellales | 0.009 | Saprotroph | |
| 18. | Camarophyllus sp. | Agaricales | 0.001 | Mycorrhizal | |
| 19. | Cantharellales | 0.002 | |||
| 20. | Chondrostereum purpureum (Pers.) Pouzar | Agaricales | 0.018 | Pathogen, saprotroph | |
| 21. | Coprinellus disseminatus (Pers.) J.E. Lange | Agaricales | 0.230 | Saprotroph | |
| 22. | Cryptococcus tephrensis Vishniac + Cryptococcus sp. | Tremellales | 0.220 | 0.406 | Saprotrophs, endophytes |
| 23. | Curvibasidium pallidicorallinum Golubev, Fell & N.W. Golubev | Incertae sedis | 0.001 | Mycocinogenic | |
| 24. | Cystobasidiomycetes | 0.003 | |||
| 25. | Cystobasidium pinicola (F.Y. Bai, L.D. Guo & J.H. Zhao) Yurkov, Kachalkin, H.M. Daniel, M. Groenew., Libkind, V. de Garcia, Zalar, Gouliam., Boekhout & Begerow + C. psychroaquaticum A.M. Yurkov, Kachalkin, H.M. Daniel, M. Groenew., Libkind, V. de Garcia, Zalar, Gouliamova, Boekhout & Begerow | Cystobasidiales | 0.002 | 0.016 | Saprotrophs, mycoparasitic |
| 26. | Cystofilobasidiales | Cystofilobasidiales | 0.004 | 0.001 | |
| 27. | Cystofilobasidium infirmominiatum (Fell, I.L. Hunter & Tallman) Hamam., Sugiy. & Komag. + C. macerans J.P. Samp. | Cystofilobasidiales | 0.012 | 0.001 | Saprotrophs, acquatic |
| 28. | Daedaleopsis confragosa (Bolton) J. Schröt | Polyporales | 0.001 | Saprotroph | |
| 29. | Efibulobasidium sp. | Sebacinales | 0.020 | Mycorrhizal | |
| 30. | Entyloma gaillardianum Vánky + E. polysporum (Peck) Farl. | Entylomatales | 0.044 | Pathogens | |
| 31. | Erythrobasidiales | Erythrobasidiales | 0.001 | 0.001 | |
| 32. | Erythrobasidium hasegawae (Y. Yamada & Komag.) Hamam., Sugiy. & Komag | Erythrobasidiales | 0.008 | Saprotroph | |
| 33. | Exidiopsis sp. | Auriculariales | 0.001 | Saprotroph | |
| 34. | Exobasidium arescens Nannf. + Exobasidium sp. | Exobasidiales | 0.001 | 0.001 | Pathogen |
| 35. | Fellomyces sp. | Tremellales | 0.001 | Saprotroph | |
| 36. | Fellozyma inositophila (Nakase & M. Suzuki) Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout | Incertae sedis | 0.007 | Saprotroph | |
| 37. | Fibulobasidium inconspicuum Bandoni | Tremellales | 0.004 | 0.379 | Saprotroph |
| 38. | Filobasidium wieringae (Á. Fonseca, Scorzetti & Fell) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout | Filobasidiales | 0.008 | Saprotroph | |
| 39. | Fomitopsis pinicola (Sw.) P. Karst | Polyporales | 0.005 | Pathogen, saprotroph | |
| 40. | Geotrichopsis mycoparasitica Tzean & Estey | Incertae sedis | 0.033 | Mycoparasitic | |
| 41. | Gymnopus androsaceus (L.) Della Magg. & Trassin | Agaricales | 0.001 | Saprotroph, mycoparasitic | |
| 42. | Hannaella zeae (O. Molnár & Prillinger) F.Y. Bai & Q.M. Wang | Tremellales | 0.047 | Saprotroph, endophyte | |
| 43. | Hebeloma mesophaeum (Pers.) Quél | Agaricales | 0.007 | Mycorrhizal | |
| 44. | Hydnaceae | Cantharellales | 0.004 | ||
| 45. | Hygrophoraceae | Agaricales | 0.008 | ||
| 46. | Hymenogaster arenarius Tul. & C. Tul. | Agaricales | 0.005 | Ectomycorrhizal | |
| 47. | Hyphodontia pallidula (Bres.) J. Erikss | Hymenochaetales | 0.003 | Saprotroph | |
| 48. | Hypochnicium lundellii (Bourdot) J. Erikss | Polyporales | 0.012 | Saprotroph | |
| 49. | Inocybe curvipes P. Karst | Agaricales | 0.043 | Ectomycorrhizal | |
| 50. | Itersonilia perplexans Derx | Cystofilobasidiales | 0.001 | Pathogen | |
| 51. | Kockovaella machilophila Cañ.-Gib., M. Takash., Sugita & Nakase | Tremellales | 0.001 | ||
| 52. | Kondoa yuccicola (Nakase & M. Suzuki) Q.M. Wang, M. Groenew., F.Y. Bai & Boekhout | Agaricostilbales | 0.012 | Saprotroph | |
| 53. | Kwoniella newhampshirensis K. Sylvester, Q.M. Wang & Hittinger + K. pini (Golubev & I. Pfeiff.) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout | Tremellales | 0.016 | 0.003 | Entomopathogenic |
| 54. | Laccaria sp. | Agaricales | 0.001 | Ectomycorrhizal | |
| 55. | Lachnella alboviolascens (Alb. & Schwein.) Fr. | Agaricales | 0.007 | Saprotroph | |
| 56. | Leptosporomyces galzinii (Bourdot) Jülich | Atheliales | 0.054 | Saprotroph | |
| 57. | Leucosporidiales | Leucosporidiales | 0.007 | ||
| 58. | Malassezia globosa Midgley, E. Guého & J. Guillot + M. restricta E. Guého, J. Guillot & Midgley + | Malasseziales | 0.016 | 0.001 | Human pathogens |
| 59. | Marasmius cohaerens (Pers.) Cooke & Quél | Agaricales | 0.008 | Saprotroph | |
| 60. | Microbotryomycetes | 0.042 | |||
| 61. | Minimedusa polyspora (Hotson) Weresub & P.M. LeClair | Cantharellales | 0.069 | Saprotroph, mycoparasitic | |
| 62. | Mrakia frigida (Fell, Statzell, I.L. Hunter & Phaff) Y. Yamada & Komag. + Mrakia sp. | Cystofilobasidiales | 0.012 | 0.001 | Saprotroph |
| 63. | Mycena aurantiomarginata (Fr.) Quél. + M. galericulata (Scop.) Gray | Agaricales | 0.003 | 0.001 | Saprotroph |
| 64. | Naganishia cerealis (Passoth, A.-C. Andersson, Olstorpe, Theelen, Boekhout & Schnürer) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout + N. diffluens (Zach) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout | Tremellales | 0.021 | 0.001 | Saprotroph |
| 65. | Oberwinklerozyma silvestris Golubev & Scorzetti ex Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout | Incertae sedis | 0.012 | ||
| 66. | Oliveonia sp. | Auriculariales | 0.008 | Saprotroph | |
| 67. | Peniophora sp. | Russulales | 0.593 | Pathogen, saprotroph | |
| 68. | Phaeotremella frondosa (Fr.) Spirin & V. Malysheva + P. roseotincta (Lloyd) V. Malysheva | Tremellales | 0.001 | 0.123 | Saprotrophs, mycoparasites |
| 69. | Phloeomana speirea (Fr.) Redhead | Agaricales | 0.024 | Saprotroph, aquatic | |
| 70. | Piskurozyma sp. | Filobasidiales | 0.024 | Saprotroph | |
| 71. | Psathyrella squamosa (P. Karst.) A.H. Sm. | Agaricales | 0.004 | Saprotroph | |
| 72. | Rhodotorula glutinis (Fresen.) F.C. Harrison + Rhodotorula sp. | Sporidiobolales | 0.003 | 0.001 | Saprotrophs |
| 73. | Saitozyma podzolica (Babeva & Reshetova) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout | Tremellales | 0.001 | Saprotroph | |
| 74. | Sakaguchia lamellibrachiae (Nagah., Hamam., Nakase & Horikoshi) Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout | Sakaguchiales | 0.027 | Saprotroph | |
| 75. | Sebacinales | Sebacinales | 0.392 | 0.001 | |
| 76. | Serendipita vermifera Oberw | Sebacinales | 0.017 | Endophyte, mycorrhizal | |
| 77. | Serpula himantioides (Fr.) P. Karst | Boletales | 0.001 | Saprotroph, pathogen | |
| 78. | Sirotrema translucens (H.D. Gordon) Bandoni | Tremellales | 0.001 | Saprotroph | |
| 79. | Sistotremastrum sp. | Trechisporales | 0.001 | Saprotroph | |
| 80. | Slooffia pilatii (F.H. Jacob, Faure-Reayn. & Berton) Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout | Incertae sedis | 0.001 | Saprotroph | |
| 81. | Solicoccozyma fuscescens (Golubev) Yurkov + S. phenolica (Á. Fonseca, Scorzetti & Fell) A.M. Yurkov + S. terrea (Di Menna) A.M. Yurkov + S. terricola (T.A. Pedersen) Yurkov | Filobasidiales | 2.451 | 0.004 | Saprotrophs |
| 82. | Sporobolomyces roseus Kluyver & C.B. Niel + Sporobolomyces sp. | 0.008 | 0.001 | ||
| 83. | Stilbum sp. | Agaricostilbales | 0.018 | Saprotroph | |
| 84. | Symmetrospora coprosmae (Hamam. & Nakase) Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout + S. gracilis (Derx) Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout | Incertae sedis | 0.005 | 0.001 | Saprotrophs |
| 85. | Tausonia pullulans (Lindner) Xin Zhan Liu, F.Y. Bai, J.Z. Groenew. & Boekhout | Cystofilobasidiales | 0.094 | 0.012 | Saprotrophs |
| 86. | Thelephoraceae | Thelephorales | 0.058 | Pathogens | |
| 87. | Tomentella sp. | Thelephorales | 0.001 | Ectomycorrhizal | |
| 88. | Tremella encephala Pers. | Tremellales | 0.003 | Saprotroph | |
| 89. | Tremellales | 0.014 | 0.001 | Saprotrophs | |
| 90. | Tremellomycetes | 0.003 | |||
| 91. | Tricholomataceae | Agaricales | 0.004 | ||
| 92. | Trichosporon otae Sugita, Takshima & Kikuchi | Trichosporonales | 0.003 | Human pathogen | |
| 93. | Tulasnellaceae | Cantharellales | 0.005 | ||
| 94. | Typhula incarnata Lasch | Agaricales | 0.004 | Pathogen | |
| 95. | Pappia fissilis (Berk. & M.A. Curtis) Zmitr | Polyporales | 0.004 | Saprotroph | |
| 96. | Vishniacozyma carnescens (Verona & Luchetti) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout + V. globispora (B.N. Johri & Bandoni) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout + V. victoriae (M.J. Montes, Belloch, Galiana, M.D. García, C. Andrés, S. Ferrer, Torr.-Rodr. & J. Guinea) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout | Tremellales | 0.007 | 0.005 | Pathogens, saprotrophs |
| Frequency Basidiomycota | 4.119 | 2.076 | |||
| Number of Basidiomycota taxa | 81 | 59 | |||
| Frequency | |||||
| Oomycota | 1.199 | 0.002 | |||
| Culturable fungi | 53.062 | 70.780 | |||
| Non-culturable fungi | 25.645 | 17.435 | |||
| Other Kingdoms | 15.822 | 11.728 | |||
| No sequence in NCBI database | 4.272 | 0.055 | |||
| Number | |||||
| Total OTUs | 69,467 a | 70,218 a | |||
| Culturable fungal OTUs | 44,506 a | 53,592 a | |||
| Taxa | 474 a | 309 a | |||
| Fungal taxa | 364 a | 242 a | |||
| Margalef’s diversity index–DMg | 65.54 | 21.72 | |||
| Shannon’s diveristy index–H | 2.55 | 0.77 | |||
| Simpson’s diversity index–D | 0.21 | 0.74 | |||
| Shannon’s evenness index–E | 0.39 | 0.17 | |||
| Berger-Parker’s dominance index–d | 0.20 | 0.46 | |||
 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwaśna, H.; Szewczyk, W.; Baranowska, M.; Gallas, E.; Wiśniewska, M.; Behnke-Borowczyk, J. Mycobiota Associated with the Vascular Wilt of Poplar. Plants 2021, 10, 892. https://doi.org/10.3390/plants10050892
Kwaśna H, Szewczyk W, Baranowska M, Gallas E, Wiśniewska M, Behnke-Borowczyk J. Mycobiota Associated with the Vascular Wilt of Poplar. Plants. 2021; 10(5):892. https://doi.org/10.3390/plants10050892
Chicago/Turabian StyleKwaśna, Hanna, Wojciech Szewczyk, Marlena Baranowska, Ewa Gallas, Milena Wiśniewska, and Jolanta Behnke-Borowczyk. 2021. "Mycobiota Associated with the Vascular Wilt of Poplar" Plants 10, no. 5: 892. https://doi.org/10.3390/plants10050892
APA StyleKwaśna, H., Szewczyk, W., Baranowska, M., Gallas, E., Wiśniewska, M., & Behnke-Borowczyk, J. (2021). Mycobiota Associated with the Vascular Wilt of Poplar. Plants, 10(5), 892. https://doi.org/10.3390/plants10050892