
Identification of Susceptibility Genes in Castanea sativa and Their Transcription Dynamics following Pathogen Infection


 , ,
, , 

Abstract
:1. Introduction
2. Results
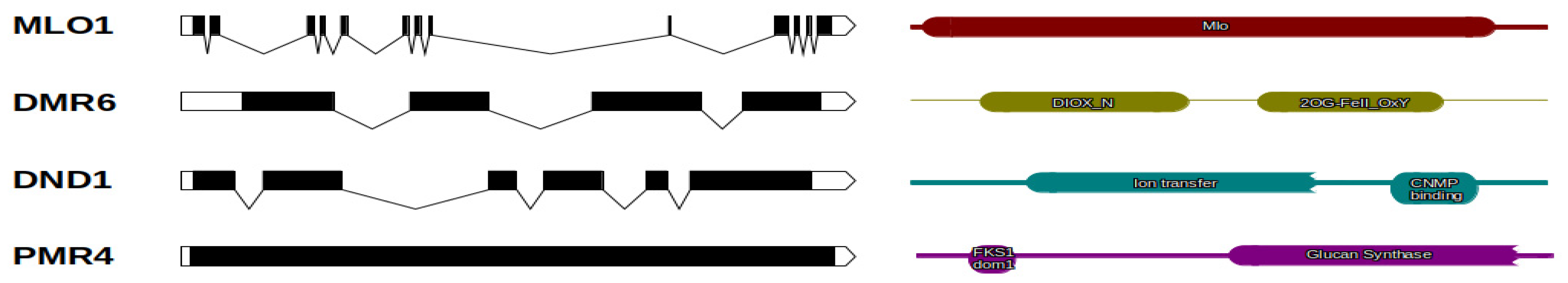
2.1. Genes Identification and Structure
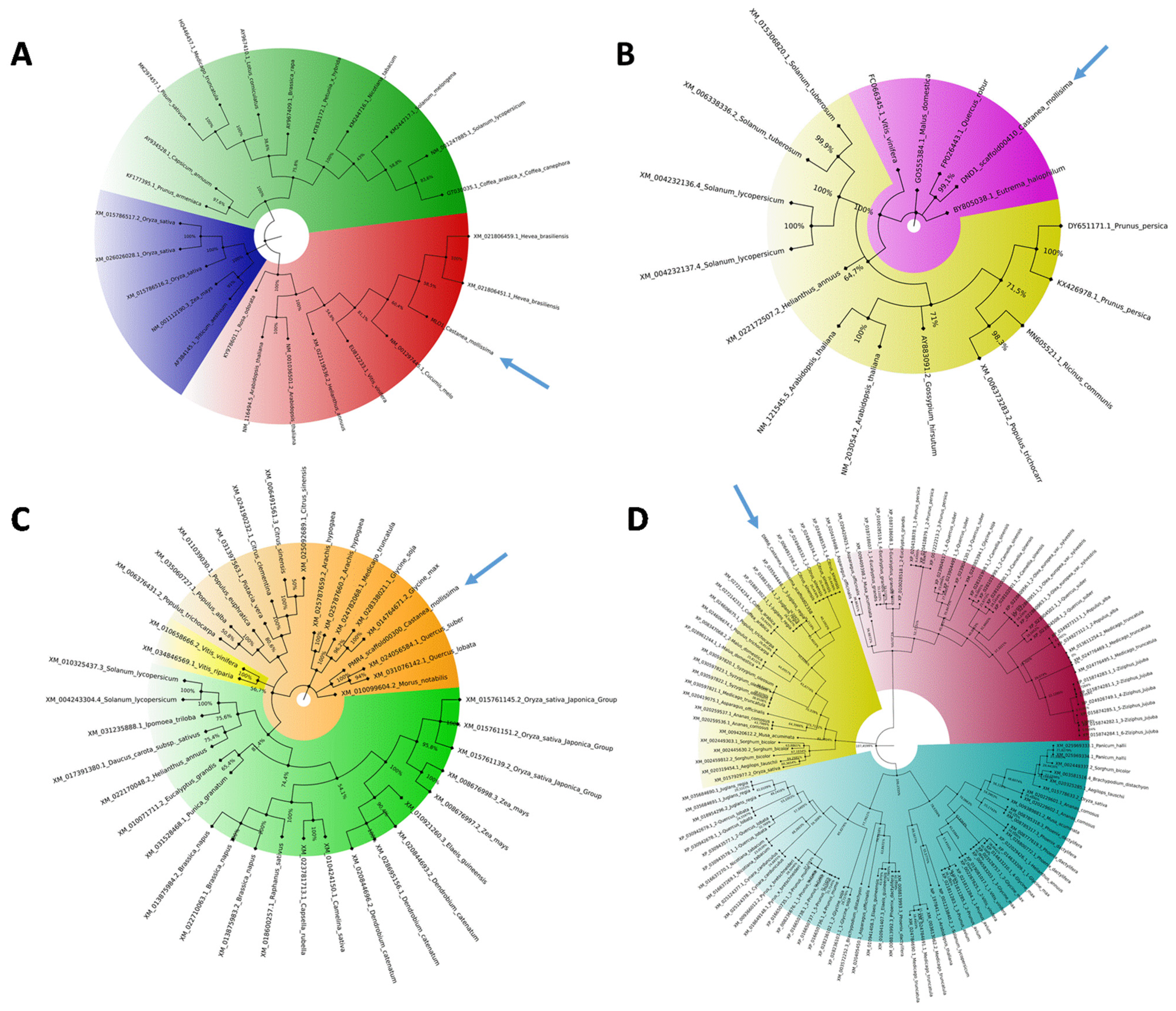
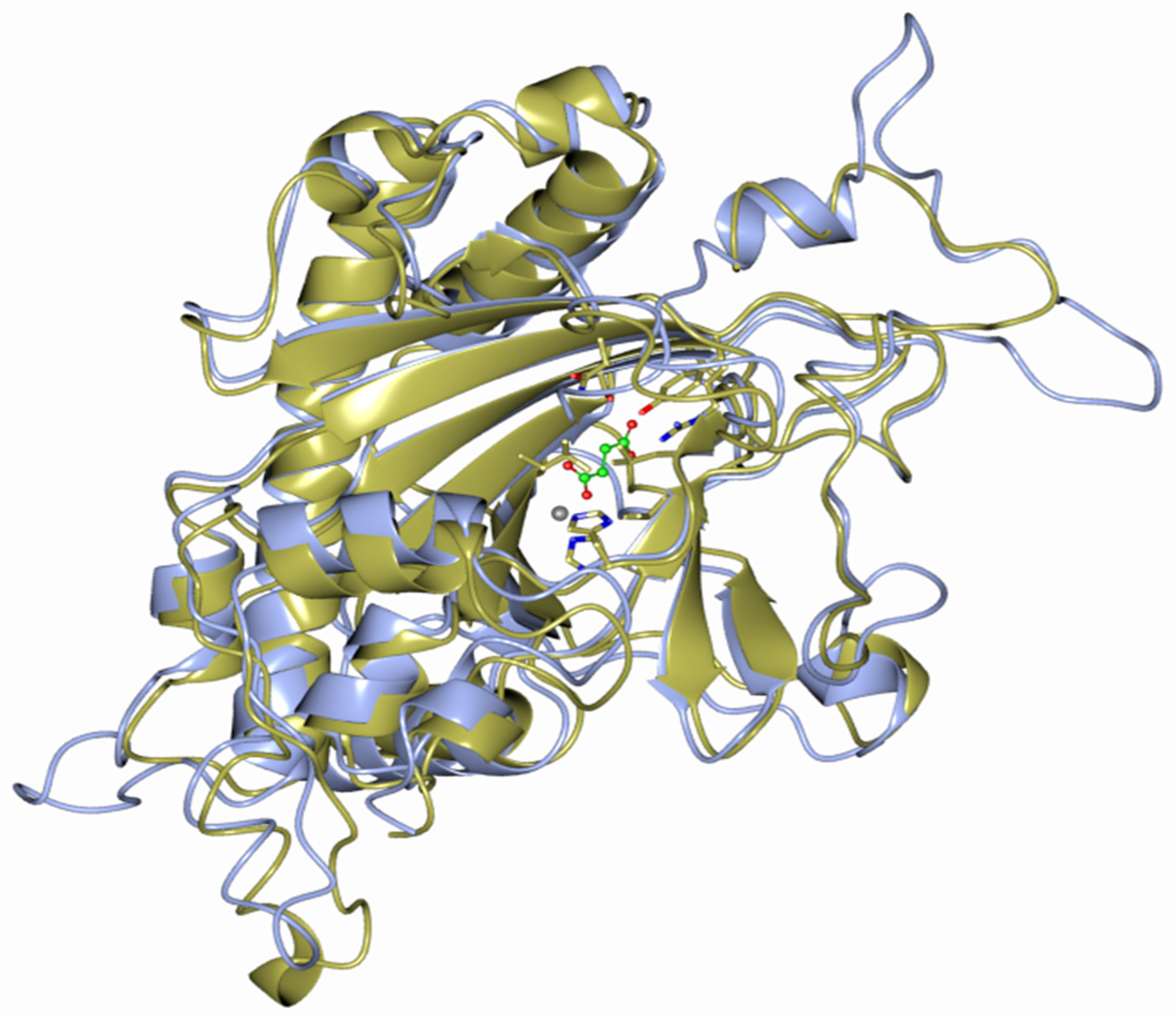
2.2. Phylogenetic and Modeling Analysis
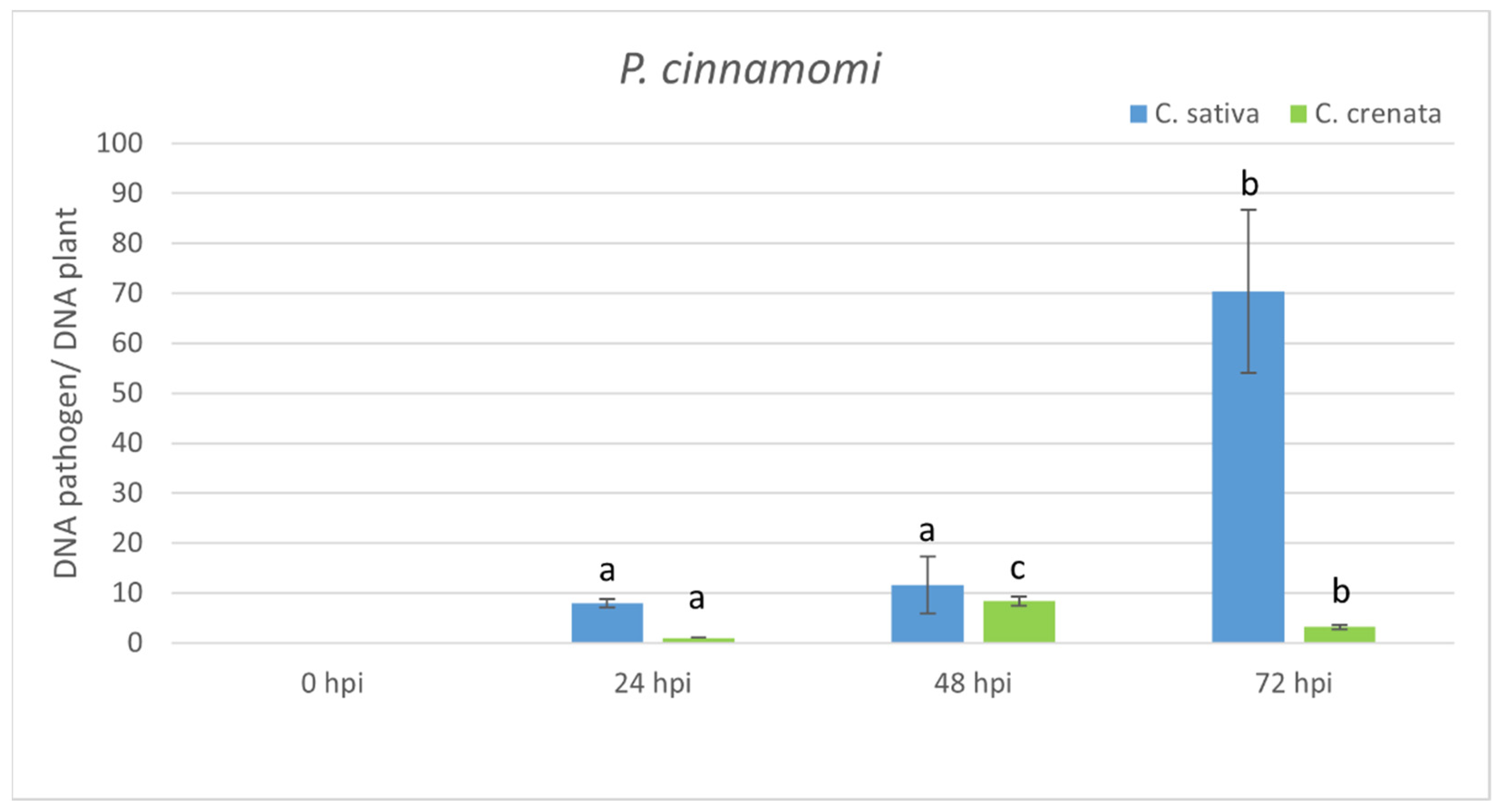
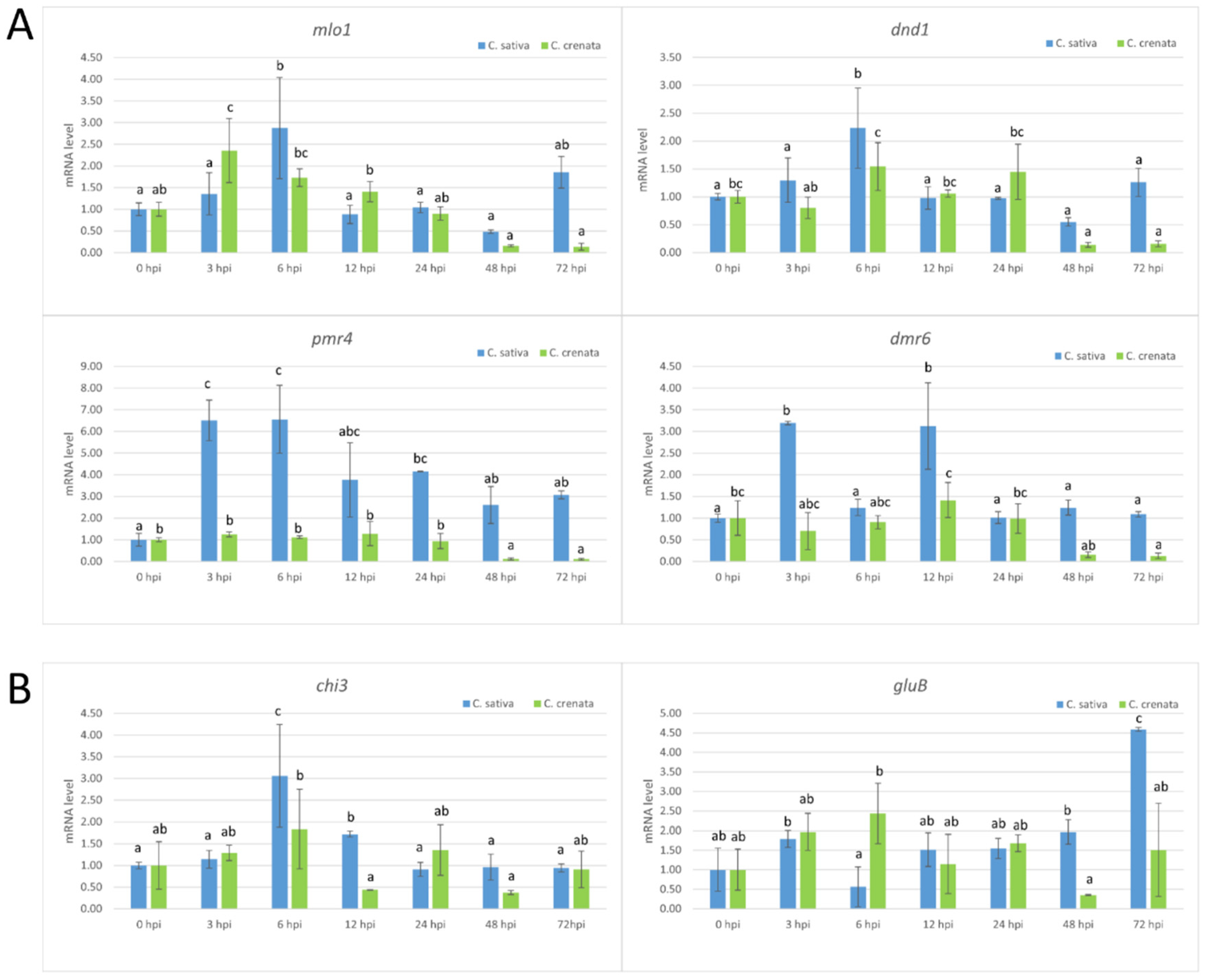
2.3. Transcriptional Profiling in Response to P. cinnamomi Infection
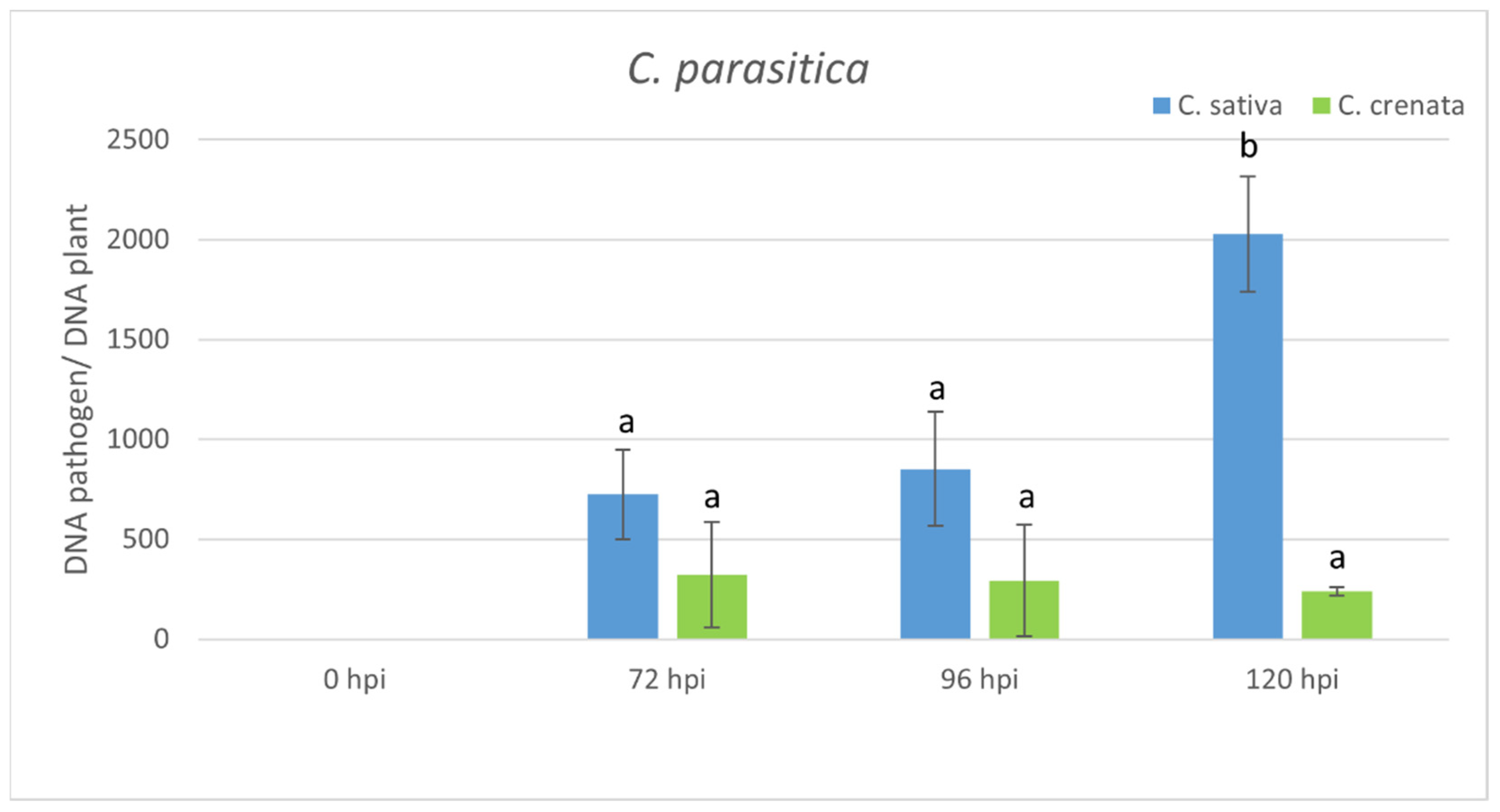
2.4. Transcriptional Profiling in Response to C. parasitica Infection
3. Discussion
4. Materials and Methods
4.1. Identification of Chestnut S-genes Orthologues
4.2. Phylogenetic Analysis
4.3. Protein Modeling
4.4. Pathogens Inoculation and Samples Collection
4.5. RNA Extraction and Real-Time qPCR Quantification
4.6. Pathogen Quantification
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Torello Marinoni, D.; Nishio, S.; Valentini, N.; Shirasawa, K.; Acquadro, A.; Portis, E.; Alma, A.; Akkak, A.; Pavese, V.; Cavalet-Giorsa, E.; et al. Development of High-Density Genetic Linkage Maps and Identification of Loci for Chestnut Gall Wasp Resistance in Castanea spp. Plants 2020, 9, 1048. [Google Scholar] [CrossRef] [PubMed]
- Conedera, M.; Krebs, P.; Tinner, W.; Pradella, M.; Torriani, D. The cultivation of Castanea sativa (Mill.) in Europe, from its origin to its diffusion on a continental scale. Veg. Hist. Archaeobot. 2004, 13, 161–179. [Google Scholar] [CrossRef] [Green Version]
- Müllerová, J.; Hédl, R.; Szabó, P. Coppice abandonment and its implications for species diversity in forest vegetation. For. Ecol. Manag. 2015, 343, 88–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zentmyer, G.A. Phytophthora cinnamomi and The Diseases it Causes; The American Phytopathological Society: St. Paul, MN, USA, 1980; Volume 10, p. 96. [Google Scholar]
- Cristinzio, G.; Grassi, G. Valutazione di resistenza a Phytophthora cambivora e Phytophthora cinnamomi in cultivar di Castanea sativa. Monti Boschi 1993, 1, 54–58. [Google Scholar]
- Serrazina, S.; Machado, H.; Costa, R.; Duque, P.; Malhó, R. Expression of Castanea crenata Allene Oxide Synthase in Arabidopsis Improves the Defense to Phytophthora cinnamomi. bioRxiv 2020. [Google Scholar] [CrossRef]
- Maurel, M.; Robin, C.; Simonneau, T.; Loustau, D.; Dreyer, E.; Desprez-Loustau, M.-L. Stomatal conductance and root-to-shoot signalling in chestnut saplings exposed to Phytophthora cinnamomi or partial soil drying. Funct. Plant Biol. 2004, 31, 41–51. [Google Scholar] [CrossRef]
- Vannini, A.; Vettraino, A.M. Ink disease in chestnuts: Impact on the European chestnut. For. Snow Landsc. Res. 2002, 76, 345–350. [Google Scholar]
- Robin, C.; Smith, I.; Hansen, E.M. Phythophthora cinnamomi. For. Phytophthoras 2012, 2, 1. [Google Scholar] [CrossRef]
- Gonthier, P.; Robin, C. Diseases. In The Chestnut Handbook: Crop and Forest Management; Beccaro, G., Alma, A., Bounous, G., Gomes-Laranjo, J., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2019; pp. 297–316, Chapter 11; ISBN 9781138334021. [Google Scholar]
- Hayden, K.J.; Garbelotto, M.; Dodd, R.; Wright, J.W. Scaling up from greenhouse resistance to fitness in the field for a host of an emerging forest disease. Evol. App. 2013, 6, 970–982. [Google Scholar] [CrossRef]
- Bounous, G. Il Castagno: Risorsa Multifunzionale in Italia e nel Mondo; Edagricole: Bologna, Italy, 2014. [Google Scholar]
- Rigling, D.; Prospero, S. Cryphonectria parasitica, the causal agent of chestnut blight: Invasion history, population biology and disease control. Mol. Plant Pathol. 2018, 1, 7–20. [Google Scholar] [CrossRef] [Green Version]
- Lione, G.; Giordano, L.; Turina, M.; Gonthier, P. Hail-Induced Infections of the Chestnut Blight Pathogen Cryphonectria parasitica Depend on Wound Size and May Lead to Severe Diebacks. Phytopathology 2020, 110, 1280–1293. [Google Scholar] [CrossRef]
- Trapiello, E.; González-Varela, G.; González, A.J. Chestnut Blight Control by Agrochemicals in Castanea sativa under Managed Conditions. J. Plant Dis. Prot. 2015, 122, 120–124. [Google Scholar] [CrossRef]
- Westbrook, J.W.; Zhang, Q.; Mandel, M.K.; Jenkins, E.V.; Barth, L.E.; Jenkins, J.W.; Grimwood, J.; Schmutz, J.; Holliday, J.A. Genomic selection analyses reveal tradeoff between blight tolerance and genome inheritance from American chestnut (Castenea dentata) in [C. dentata x C. mollissima] x C. dentata backcross populations. BioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Kubisiak, T.L.; Hebard, F.V.; Nelson, C.D.; Zhang, J.; Bernatzky, R.; Huang, H.; Anagnostakis, S.L.; Doudrick, R.L. Molecular Mapping of Resistance to Blight in an Interspecific Cross in the genus Castanea. Phytopathology 1997, 87, 751–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubisiak, T.L.; Nelson, C.D.; Staton, M.E.; Zhebentyayeva, T.; Smith, C.; Olukolu, B.A.; Fang, G.-C.; Hebard, F.V.; Anagnostakis, S.; Wheeler, N.; et al. A transcriptome-based genetic map of Chinese chestnut (Castanea mollissima) and identification of regions of segmental homology with peach (Prunus persica). Tree Genet. Genomes 2012, 9, 557–571. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Oakes, A.D.; Newhouse, A.E.; Baier, K.M.; Maynard, C.A.; Powell, W.A. A threshold level of oxalate oxidase transgene expression reduces Cryphonectria parasitica-induced necrosis in a transgenic American chestnut (Castanea dentata) leaf bioassay. Transgenic Res. 2013, 22, 973–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcolin, E.; Manetti, M.C.; Pelleri, F.; Conedera, M.; Pezzatti, G.B.; Lingua, E.; Pividori, M. Seed regeneration of sweet chestnut (Castanea sativa Miller) under different coppicing approaches. For. Ecol. Manag. 2020, 472, 118273. [Google Scholar] [CrossRef]
- Aebi, A.; Schönrogge, K.; Melika, G.; Alma, A.; Bosio, G.; Quacchia, A.; Picciau, L.; Abe, Y.; Moriya, S.; Yara, K.; et al. Parasitoid recruitment to the globally invasive chestnut gall wasp Dryocosmus kuriphilus. In Ecology and Evolution of Galling Arthropods and Their Associates; Ozaki, K., Yukwa, J., Ohgushi, T., Price, P.W., Eds.; Springer: Tokyo, Japan, 2006; pp. 103–121. [Google Scholar]
- Van Schie, C.C.; Takken, F.L. Susceptibility genes 101: How to be a good host. Annu. Rev. Phytopathol. 2014, 52, 551–581. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.S.; Mukhtar, M.S.; Mansoor, S. Genome editing: Targeting susceptibility genes for plant disease resistance. Trends Biotechnol. 2018, 36, 898–906. [Google Scholar] [CrossRef] [PubMed]
- Pavan, S.; Jacobsen, E.; Visser, R.G.; Bai, Y. Loss of susceptibility as a novel breeding strategy for durable and broad-spectrum resistance. Mol. Breed. 2010, 25, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Devoto, A.; Hartmann, H.A.; Piffanelli, P.; Elliott, C.; Simmons, C.; Taramino, G.; Goh, C.S.; Cohen, F.E.; Emerson, B.C.; Schulze-Lefert, P.; et al. Molecular phylogeny and evolution of the plant-specific seven-transmembrane MLO family. J. Mol. Evol. 2003, 56, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Büschges, R.; Hollricher, K.; Panstruga, R.; Simons, G.; Wolter, M.; Frijters, A.; van Daelen, R.; van der Lee, T.; Diergaarde, P.; Groenendijk, J.; et al. The barley Mlo gene: A novel control element of plant pathogen resistance. Cell 1997, 88, 695–705. [Google Scholar] [CrossRef] [Green Version]
- Appiano, M.; Pavan, S.; Catalano, D.; Zheng, Z.; Bracuto, V.; Lotti, C.; Visser, R.G.; Ricciardi, L.; Bai, Y. Identification of candidate MLO powdery mildew susceptibility genes in cultivated Solanaceae and functional characterization of tobacco NtMLO1. Transgenic Res. 2015, 24, 847–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santillán Martínez, M.I.; Bracuto, V.; Koseoglou, E.; Appiano, M.; Jacobsen, E.; Visser, R.G.F.; Wolters, A.A.; Bai, Y. CRISPR/Cas9-targeted mutagenesis of the tomato susceptibility gene PMR4 for resistance against powdery mildew. BMC Plant Biol. 2020, 20, 284. [Google Scholar] [CrossRef]
- Sun, K.; Wolters, A.M.A.; Loonen, A.E.; Huibers, R.P.; van der Vlugt, R.; Goverse, A.; Jacobsen, E.; Visser, R.G.; Bai, Y. Down-regulation of Arabidopsis DND1 orthologs in potato and tomato leads to broad-spectrum resistance to late blight and powdery mildew. Transgenic Res. 2016, 25, 123–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Damme, M.; Huibers, R.P.; Elberse, J.; Van Den Ackerveken, G. Arabidopsis DMR6 encodes a putative 2OG-Fe(II) oxygenase that is defense-associated but required for susceptibility to downy mildew. Plant J. 2008, 54, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Zeilmaker, T.; Ludwig, N.R.; Elberse, J.; Seidl, M.F.; Berke, L.; Van Doorn, A.; Schuurink, R.C.; Snel, B.; Van den Ackerveken, G. Downy mildew resistant 6 and DMR6-like oxygenase 1 are partially redundant but distinct suppressors of immunity in Arabidopsis. Plant J. 2015, 81, 210–222. [Google Scholar] [CrossRef]
- Liyanage, K.K.; Khan, S.; Herath, V.; Brooks, S.; Mortimer, P.E.; Nadir, S.; Hyde, K.D.; Xu, J. Genome Wide Identification of the MLO Gene Family Associated with Powdery Mildew Resistance in Rubber Trees (Hevea brasiliensis). Trop. Plant Biol. 2020, 13, 331–342. [Google Scholar] [CrossRef]
- Filiz, E.; Vatansever, R. Genome-wide identification of mildew resistance locus O (MLO) genes in tree model poplar (Populus trichocarpa): Powdery mildew management in woody plants. Eur. J. Plant Pathol. 2018, 152, 95–109. [Google Scholar] [CrossRef]
- Pessina, S.; Lenzi, L.; Perazzolli, M.; Campa, M.; Dalla Costa, L.; Urso, S.; Valè, G.; Salamini, F.; Velasco, R.; Malnoy, M. Knockdown of MLO genes reduces susceptibility to powdery mildew in grapevine. Hortic. Res. 2016, 3, 16016. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Pérez-Sierra, A.; Durán, A.; Jung, M.H.; Balci, Y.; Scanu, B. Canker and decline diseases caused by soil- and airborne Phytophthora species in forests and woodlands. Pers. Mol. Phylogeny Evol. Fungi 2018, 40, 182–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira-Lorenzo, S.; Ballester, A.; Corredoira, E.; Bounous, G.; Botta, R.; Beccaro, G.L.; Kubisiak, T.L.; Conedera, M.; Krebs, P.; Yamamoto, T.; et al. Chestnut. In Fruit Breeding; Badenes, M.L., Byrne, D.H., Eds.; Springer Science + Business Media: New York, NY, USA, 2012; pp. 729–769. [Google Scholar]
- Barakat, A.; Staton, M.; Cheng, C.H.; Park, J.; Yassin, N.B.M.; Ficklin, S.; Yeh, C.C.; Hebard, F.; Baier, K.; Powell, W.; et al. Chestnut resistance to the blight disease: Insights from transcriptome analysis. BMC Plant Biol. 2012, 12, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrazina, S.; Santos, C.; Machado, H.; Pesquita, C.; Vicentini, R.; Pais, M.S.; Sebastiana, M.; Costa, R. Castanea root transcriptome in response to Phytophthora cinnamomi challenge. Tree Genet. Genomes 2015, 11, 19. [Google Scholar] [CrossRef]
- Santos, C.; Machado, H.; Correia, I.; Gomes, F.; Gomes-Laranjo, J.; Costa, R. Phenotyping Castanea hybrids for Phytophthora cinnamomi resistance. Plant Pathol. 2015, 64, 901–910. [Google Scholar] [CrossRef]
- Santos, C.; Duarte, S.; Tedesco, S.; Fevereiro, P.; Costa, R.L. Expression profiling of Castanea genes during resistant and susceptible interactions with the oomycete pathogen Phytophthora cinnamomi reveal possible mechanisms of immunity. Front. Plant Sci. 2017, 8, 515. [Google Scholar] [CrossRef] [Green Version]
- Santos, C.; Nelson, C.D.; Zhebentyayeva, T.; Machado, H.; Gomes-Laranjo, J.; Costa, R.L. First interspecific genetic linkage map for Castanea sativa x Castanea crenata revealed QTLs for resistance to Phytophthora cinnamomi. PLoS ONE 2017, 12, e0184381. [Google Scholar] [CrossRef] [Green Version]
- Barakat, A.; DiLoreto, D.S.; Zhang, Y.; Smith, C.; Baier, K.; Powell, W.A.; Wheeler, N.; Sederoff, R.; Carlson, J.E. Comparison of the transcriptomes of American chestnut (Castanea dentata) and Chinese chestnut (Castanea mollissima) in response to the chestnut blight infection. BMC Plant Biol. 2009, 9, 51. [Google Scholar] [CrossRef] [Green Version]
- van Esse, H.P.; Reuber, T.L.; van der Does, D. Genetic modification to improve disease resistance in crops. New Phytol. 2020, 225, 70–86. [Google Scholar] [CrossRef] [Green Version]
- Wan, D.Y.; Guo, Y.; Cheng, Y.; Hu, Y.; Xiao, S.; Wang, Y.; Wen, Y.Q. CRISPR/Cas9-mediated mutagenesis of VvMLO3 results in enhanced resistance to powdery mildew in grapevine (Vitis vinifera). Hortic. Res. 2020, 7, 116. [Google Scholar] [CrossRef]
- Clough, S.J.; Fengler, K.A.; Yu, I.C.; Lippok, B.; Smith, R.K.; Bent, A.F. The Arabidopsis dnd1 “defense, no death” gene encodes a mutated cyclic nucleotide-gated ion channel. Proc. Natl. Acad. Sci. USA 2000, 97, 9323–9328. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, M.T.; Stein, M.; Hou, B.H.; Vogel, J.P.; Edwards, H.; Somerville, S.C. Loss of a callose synthase results in salicylic acid-dependent disease resistance. Science 2003, 301, 969–972. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, L.; Zhao, J.; Li, Y.; Wang, J.; Guo, R.; Gan, S.; Liu, C.J.; Zhang, K. S5H/DMR6 encodes a salicylic acid 5-hydroxylase that fine-tunes salicylic acid homeostasis. Plant Physiol. 2017, 175, 1082–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huibers, R.P.; Loonen, A.E.; Gao, D.; Van den Ackerveken, G.; Visser, R.G.; Bai, Y. Powdery mildew resistance in tomato by impairment of SlPMR4 and SlDMR1. PLoS ONE 2013, 8, e67467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saiz-Fernández, I.; Milenković, I.; Berka, M.; Černý, M.; Tomšovský, M.; Brzobohatý, B.; Kerchev, P. Integrated Proteomic and Metabolomic Profiling of Phytophthora cinnamomi Attack on Sweet Chestnut (Castanea sativa) Reveals Distinct Molecular Reprogramming Proximal to the Infection Site and Away from It. Int. J. Mol. Sci. 2020, 21, 8525. [Google Scholar] [CrossRef]
- Lovat, C.-A.; Donnelly, D.J. Mechanisms and metabolomics of the host-pathogen interactions between Chestnut (Castanea species) and Chestnut blight (Cryphonectria parasitica). For. Pathol. 2019, 49, e12562. [Google Scholar] [CrossRef]
- Fernandes, P.; Machado, H.; Silva, M.D.C.; Costa, R.L. A Histopathological Study Reveals New Insights Into Responses of Chestnut (Castanea spp.) to Root Infection by Phytophthora cinnamomi. Phytopathology 2021. [Google Scholar] [CrossRef] [PubMed]
- Veluthakkal, R.; Dasgupta, M.G. Pathogenesis-related genes and proteins in forest tree species. Trees 2010, 24, 993–1006. [Google Scholar] [CrossRef]
- Puthoff, D.P.; Holzer, F.M.; Perring, T.M.; Walling, L.L. Tomato Pathogenesis-related Protein Genes are Expressed in Response to Trialeurodes vaporariorum and Bemisia tabaci Biotype B Feeding. J. Chem. Ecol. 2010, 36, 1271–1285. [Google Scholar] [CrossRef] [Green Version]
- Molinari, S.; Fanelli, E.; Leonetti, P. Expression of tomato salicylic acid (SA)-responsive pathogenesis-related genes in Mi-1-mediated and SA-induced resistance to root-knot nematodes. Mol. Plant Pathol. 2014, 15, 255–264. [Google Scholar] [CrossRef]
- Broglie, K.; Chet, I.; Holliday, M.; Cressman, R.; Biddle, P.; Knowlton, S.; Mauvais, C.J.; Broglie, R. Transgenic plants with enhanced resistance to the fungal pathogen, Rhizoctonia solani. Science 1991, 254, 1194–1197. [Google Scholar] [CrossRef] [PubMed]
- Mauch, F.; Staehelin, L.A. Functional Implications of the Subcellular Localization of Ethylene-Induced Chitinase and [beta]-1,3-Glucanase in Bean Leaves. Plant Cell 1989, 1, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Schlumbaum, A.; Mauch, F.; Vögeli, U.; Boller, T. Plant chitinases are potent inhibitors of fungal growth. Nature 1986, 324, 365–367. [Google Scholar] [CrossRef]
- Shain, L.; Miller, J.B.; Spalding, R.J. Responses of American and Chinese Chestnut to Cryphonectria parasitica and Ethylene. In Proceedings of the International Chestnut Conference; Double, M.L., MaCDonald, D.H., Eds.; West Virginia University Press: Morgantown, WV, USA, 1994; pp. 97–101. [Google Scholar]
- Vannini, A.; Caruso, C.; Leonardi, L.; Rugini, E.; Chiarot, E.; Caporale, C.; Buonocore, V. Antifungal properties of chitinases from Castanea sativa against hypovirulent and virulent strains of the chestnut blight fungus Cryphonectria parasitica. Physiol. Mol. Plant Pathol. 1999, 55, 29–35. [Google Scholar] [CrossRef]
- Corredoira, E.; San José, M.C.; Vieitez, A.M.; Allona, I.; Aragoncillo, C.; Ballester, A. Agrobacterium-mediated transformation of European chestnut somatic embryos with a Castanea sativa (Mill.) endochitinase gene. New For. 2016, 47, 669–684. [Google Scholar] [CrossRef]
- Zhu, H.; Li, C.; Gao, C. Applications of CRISPR–Cas in agriculture and plant biotechnology. Nat. Rev. Mol. Cell Biol. 2020, 21, 661–677. [Google Scholar] [CrossRef]
- Schenke, D.; Cai, D. Applications of CRISPR/Cas to Improve Crop Disease Resistance: Beyond Inactivation of Susceptibility Factors. iScience 2020, 23, 101478. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, S.; Kesiraju, K.; Saakre, M.; Rathinam, M.; Raman, V.; Pattanayak, D.; Sreevathsa, R. Genome editing for resistance to insect pests: An emerging tool for crop improvement. ACS Omega 2020, 5, 20674–20683. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cheng, X.; Shan, Q.; Zhang, Y.; Liu, J.; Gao, C.; Qiu, J.L. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat. Biotechnol. 2014, 32, 947–951. [Google Scholar] [CrossRef]
- Malnoy, M.; Viola, R.; Jung, M.H.; Koo, O.-J.; Kim, S.; Kim, J.-S.; Velasco, R.; Kanchiswamy, C.N. DNA-Free Genetically Edited Grapevine and Apple Protoplast Using CRISPR/Cas9 Ribonucleoproteins. Front. Plant Sci. 2016, 7, 1904. [Google Scholar] [CrossRef]
- Nekrasov, V.; Wang, C.; Win, J.; Lanz, C.; Weigel, D.; Kamoun, S. Rapid generation of a transgene-free powdery mildew resistant tomato by genome deletion. Sci. Rep. 2017, 7, 482. [Google Scholar] [CrossRef] [Green Version]
- de Toledo Thomazella, D.P.; Brail, Q.; Dahlbeck, D.; Staskawicz, B.J. CRISPR-Cas9 mediated mutagenesis of a DMR6 ortholog in tomato confers broad-spectrum disease resistance. bioRxiv 2016, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Lione, G.; Danti, R.; Fernandez-Conradi, P.; Ferreira-Cardoso, J.V.; Lefort, F.; Marques, G.; Meyer, J.B.; Prospero, S.; Radòcz, L.; Robin, C.; et al. The emerging pathogen of chestnut Gnomoniopsis castaneae: The challenge posed by a versatile fungus. Eur. J. Plant Pathol. 2019, 153, 671–685. [Google Scholar] [CrossRef]
- Webb, B.; Sali, A. Comparative Protein Structure Modeling Using MODELLER. Curr. Protoc. Bioinform. 2016, 54, 5.6.1–5.6.37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marinoni, D.; Akkak, A.; Bounous, G.; Edwards, K.J.; Botta, R. Development and characterization of microsatellite markers in Castanea sativa (Mill.). Mol. Breed. 2003, 11, 127–136. [Google Scholar] [CrossRef]
- Zampieri, E.; Giordano, L.; Lione, G.; Vizzini, A.; Sillo, F.; Balestrini, R.; Gonthier, P. A nonnative and a native fungal plant pathogen similarly stimulate ectomycorrhizal development but are perceived differently by a fungal symbiont. New Phytol. 2017, 213, 1836–1849. [Google Scholar] [CrossRef]
- Robin, C.; Morel, O.; Vettraino, A.M.; Perlerou, C.; Diamondis, S.; Vannini, A. Genetic variation in susceptibility to Phytophthora cambivora in European chestnut (Castanea sativa). For. Ecol. Manag. 2006, 226, 199–207. [Google Scholar] [CrossRef]
- Dinis, L.-T.; Peixoto, F.; Zhang, C.; Martins, L.; Costa, R.; Gomes-Laranjo, J. Physiological and biochemical changes in resistant and sensitive chestnut (Castanea) plantlets after inoculation with Phytophthora cinnamomi. Physiol. Mol. Plant Pathol. 2011, 75, 146–156. [Google Scholar] [CrossRef]
- Wu, C.T.; Leubner-Metzger, G.; Meins, F. Class I beta-1,3-glucanase and chitinase are expressed in the micropylar endosperm of tomato seeds prior to radicle emergence. Plant Physiol. 2001, 126, 1299–1313. [Google Scholar] [CrossRef] [Green Version]
- Popov, A.P.; Tsvetkov, I.L.; Belov, A.A.; Konichev, A.S.; Ivanushkina, N.E.; Kochkina, G.A.; Ozerskaya, S.M. Molecular genetic identification of the phytopathogenic fungus Cryphonectria parasitica. Microbiology 2010, 79, 223–228. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Scaffold | ORF Length (bp) | N° Exons | Size (aa) | Domains | PFAM DOMAINS |
|---|---|---|---|---|---|---|
| MLO1 | scaffold00101 | 1425 | 13 | 474 | Mlo | PF03094 |
| DMR6 | scaffold02358 | 1128 | 4 | 375 | 2OG-FeII_Oxy; DIOX_N | PF03171;PF14226 |
| DND1 | scaffold00410 | 1407 | 6 | 468 | cNMP_binding; Ion_trans | PF00027;PF00520 |
| PMR4 | scaffold00300 | 5346 | 1 | 1781 | FKS1_dom1; Glucan_synthase | PF14288;PF02364 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavese, V.; Moglia, A.; Gonthier, P.; Torello Marinoni, D.; Cavalet-Giorsa, E.; Botta, R. Identification of Susceptibility Genes in Castanea sativa and Their Transcription Dynamics following Pathogen Infection. Plants 2021, 10, 913. https://doi.org/10.3390/plants10050913
Pavese V, Moglia A, Gonthier P, Torello Marinoni D, Cavalet-Giorsa E, Botta R. Identification of Susceptibility Genes in Castanea sativa and Their Transcription Dynamics following Pathogen Infection. Plants. 2021; 10(5):913. https://doi.org/10.3390/plants10050913
Chicago/Turabian StylePavese, Vera, Andrea Moglia, Paolo Gonthier, Daniela Torello Marinoni, Emile Cavalet-Giorsa, and Roberto Botta. 2021. "Identification of Susceptibility Genes in Castanea sativa and Their Transcription Dynamics following Pathogen Infection" Plants 10, no. 5: 913. https://doi.org/10.3390/plants10050913
APA StylePavese, V., Moglia, A., Gonthier, P., Torello Marinoni, D., Cavalet-Giorsa, E., & Botta, R. (2021). Identification of Susceptibility Genes in Castanea sativa and Their Transcription Dynamics following Pathogen Infection. Plants, 10(5), 913. https://doi.org/10.3390/plants10050913