
Deciphering the Molecular Mechanism Responsible for Efficiently Inhibiting Metastasis of Human Non-Small Cell Lung and Colorectal Cancer Cells Targeting the Matrix Metalloproteinases by Selaginella repanda

 ,
,  ,
, 
 ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. High Resolution-Liquid Chromatography-Mass Spectroscopy (HR-LC-MS) Identification of S. repanda Phytoconstituents
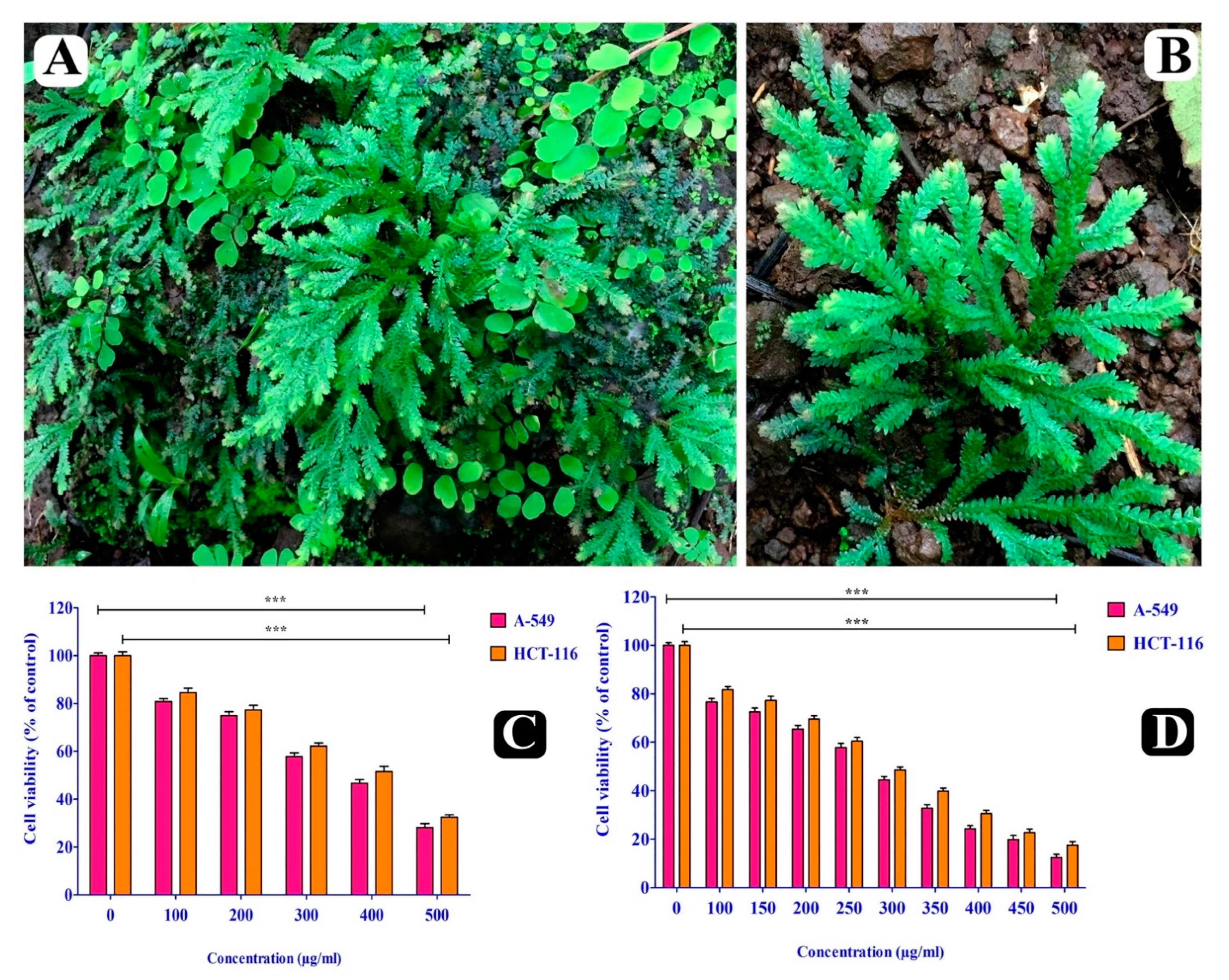
2.2. Cytotoxic Effect of S. repanda Crude Extract
2.3. Anti-Migratory Effect of S. repanda Crude Extract
2.4. Anti-Invasion Effect of S. repanda Crude Extract
2.5. Anti-Adhesion Effect of S. repanda Crude Extract
2.6. Changes in Transcriptional Level of Metastasis Related Genes
3. Discussion
4. Materials and Methods
4.1. Plant Material Collection and Extraction
4.2. HR-LC–MS Analysis
4.3. Cell culture and Treatment
4.4. Cell Viability Analysis Using MTT Assay
4.5. Wound Closure Assay
4.6. Invasion Assay
4.7. Cell Adhesion Assay
4.8. Expression Levels Determination of Metastasis Related Genes
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021. [Google Scholar] [CrossRef]
- Coghlin, C.; I Murray, G. Current and emerging concepts in tumour metastasis. J. Pathol. 2010, 222, 1–15. [Google Scholar] [CrossRef]
- Adnan, M.; Khan, S.; Al-Shammari, E.; Patel, M.; Saeed, M.; Hadi, S.; Information, P.E.K.F.C. In pursuit of cancer metastasis therapy by bacteria and its biofilms: History or future. Med. Hypotheses 2017, 100, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Sak, K. Chemotherapy and Dietary Phytochemical Agents. Chemother. Res. Pr. 2012, 2012, 282570. [Google Scholar] [CrossRef] [Green Version]
- Baskar, R.; Dai, J.; Wenlong, N.; Yeo, R.; Yeoh, K.-W. Biological response of cancer cells to radiation treatment. Front. Mol. Biosci. 2014, 1, 24. [Google Scholar] [CrossRef] [Green Version]
- Chikara, S.; Nagaprashantha, L.D.; Singhal, J.; Horne, D.; Awasthi, S.; Singhal, S.S. Oxidative stress and dietary phyto-chemicals: Role in cancer chemoprevention and treatment. Cancer Lett. 2018, 413, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Sharma, B.; Kanwar, S.S.; Kumar, A. Lead Phytochemicals for Anticancer Drug Development. Front. Plant Sci. 2016, 7, 1667. [Google Scholar] [CrossRef] [Green Version]
- Adnan, M.; Siddiqui, A.J.; Jamal, A.; Hamadou, W.S.; Awadelkareem, A.M.; Sachidanandan, M.; Patel, M. Evidence-Based Medicinal Potential and Possible Role of Selaginella in the Prevention of Modern Chronic Diseases: Ethnopharmacological and Ethnobotanical Perspective. Rec. Nat. Prod. 2021, 15, 330–355. [Google Scholar] [CrossRef]
- Adnan, M.; Siddiqui, A.J.; Hamadou, W.S.; Patel, M.; Ashraf, S.A.; Jamal, A.; Awadelkareem, A.M.; Sachidanandan, M.; Snoussi, M.; De Feo, V. Phytochemistry, Bioactivities, Pharmacokinetics and Toxicity Prediction of Selaginella repanda with Its Anticancer Potential against Human Lung, Breast and Colorectal Carcinoma Cell Lines. Molecules 2021, 26, 768. [Google Scholar] [CrossRef] [PubMed]
- Han, B.-H.; Chi, H.-J.; Han, Y.-N.; Ryu, K.-S. Screening on the anti-inflammatory activity of crude drugs. Korean J. Pharmacogn. 1972, 3, 205–209. [Google Scholar]
- Itokawa, H.; Mihashi, S.; Watanabe, K.; Natsumoto, H.; Hamanaka, T. Studies on the constituents of crude drugs having inhibitory activity against contraction of the ileum caused by histamine or barium chloride (1) screening test for the activ-ity of commercially available crude drugs and the related plant materials. Shoyakugaku Zasshi 1983, 37, 223–228. [Google Scholar]
- Macfoy, C.A.; Sama, A.M. Medicinal plants in pujehun district of sierra leone. J. Ethnopharmacol. 1983, 8, 215–223. [Google Scholar] [CrossRef]
- Han, D.S.L.; Lee, H.K. Ethnobotanical survey in Korea. In Proceedings of the Fifth Asian Symposium on Medicinal Plants and Spices, Seoul, Korea, 20–24 August 1984; Volume 5, p. 125. [Google Scholar]
- Winkelman, M. Frequently used medicinal plants in Baja California Norte. J. Ethnopharmacol. 1986, 18, 109–131. [Google Scholar] [CrossRef]
- Darias, V.; Bravo, L.; Rabanal, R.; Mateo, C.; Luis, R.; Pérez, A. New contribution to the ethnopharmacological study of the canary islands. J. Ethnopharmacol. 1989, 25, 77–92. [Google Scholar] [CrossRef]
- Ono, K.; Nakane, H.; MENG, Z.-M.; OSE, Y.; SAKAI, Y.; MIZUNO, M. Differential inhibitory effects of various herb ex-tracts on the activities of reverse transcriptase and various deoxyribonucleic acid (DNA) polymerases. Chem. Pharm. Bull. 1989, 37, 1810–1812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Z.; Sakai, Y.; Ose, Y.; Sato, T.; Nagase, H.; Kito, H.; Matsuda, H.; Sato, M.; Mizuno, M. Antimutagenic activity of medical plants in traditional Chinese medicines. Mutat. Res. Mutagen. Relat. Subj. 1988, 203, 378–379. [Google Scholar] [CrossRef]
- Lin, R.; Skaltsounis, A.-L.; Seguin, E.; Tillequin, F.; Koch, M. Phenolic Constituents of Selaginella doederleinii. Planta Medica 1994, 60, 168–170. [Google Scholar] [CrossRef]
- De Sá, P.G.S.; Nunes, X.P.; De Lima, J.T.; Filho, J.A.D.S.; Fontana, A.P.; Siqueira, J.D.S.; Quintans-Júnior, L.J.; Damasceno, P.K.F.; Branco, C.R.C.; Branco, A.; et al. Antinociceptive effect of ethanolic extract of Selaginella convoluta in mice. BMC Complement. Altern. Med. 2012, 12, 187. [Google Scholar] [CrossRef] [Green Version]
- Almeida, J.R.G.; de Sa, P.G.S.; Macedo, L.A.R.; Filho, J.A.; Oliviera, V.R.; Filho, J.M.B. Phytochemistry of the genus Sela-ginella (Selaginellaceae). J. Med. Plants Res. 2013, 7, 1858–1868. [Google Scholar]
- Harbone, J.B.; Williams, C.A. ChemInform Abstract: Advances in Flavonoid Research since 1992. Phytochemistry 2010, 32, 481–504. [Google Scholar] [CrossRef]
- Pellati, F.; Benvenuti, S.; Magro, L.; Melegari, M.; Soragni, F. Analysis of phenolic compounds and radical scavenging activity of Echinacea spp. J. Pharm. Biomed. Anal. 2004, 35, 289–301. [Google Scholar] [CrossRef]
- Lan, L.; Wang, Y.; Pan, Z.; Wang, B.; Yue, Z.; Jiang, Z.; Li, L.; Wang, C.; Tang, H. Rhamnetin induces apoptosis in human breast cancer cells via the miR-34a/Notch-1 signaling pathway. Oncol. Lett. 2018, 17, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.-C.; Lin, H.-H.; Hsu, C.-H.; Wang, C.-J.; Chiang, T.-A.; Chen, J.-H. Inhibitory effects of andrographolide on migra-tion and invasion in human non-small cell lung cancer A549 cells via down-regulation of PI3K/Akt signaling pathway. Eur. J. Pharmacol. 2010, 632, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.-B.; Hsieh, M.-J.; Lin, C.-W.; Chiou, H.-L.; Lin, P.-Y.; Chen, T.-Y.; Yang, S.-F. The antimetastatic effects of resvera-trol on hepatocellular carcinoma through the downregulation of a metastasis-associated protease by SP-1 modulation. PLoS ONE 2013, 8, e56661. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.-F.; Chen, M.-K.; Hsieh, Y.-S.; Yang, J.-S.; Zavras, A.-I.; Hsieh, Y.-H.; Su, S.-C.; Kao, T.-Y.; Chen, P.-N.; Chu, S.-C. Antimetastatic effects of Terminalia catappa L. on oral cancer via a down-regulation of metastasis-associated prote-ases. Food Chem. Toxicol. 2010, 48, 1052–1058. [Google Scholar] [CrossRef]
- Giannelli, G.; Bergamini, C.; Fransvea, E.; Marinosci, F.; Quaranta, V.; Antonaci, S. Human Hepatocellular Carcinoma (HCC) Cells Require Both α3β1 Integrin and Matrix Metalloproteinases Activity for Migration and Invasion. Lab. Investig. 2001, 81, 613–627. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.; Sachidanandan, M.; Adnan, M. Serine arginine protein kinase 1 (SRPK1): A moonlighting protein with theranostic ability in cancer prevention. Mol. Biol. Rep. 2018, 46, 1487–1497. [Google Scholar] [CrossRef]
- Liotta, L.A. An attractive force in metastasis. Nat. Cell Biol. 2001, 410, 24–25. [Google Scholar] [CrossRef]
- Reddy, M.N.; Adnan, M.; Alreshidi, M.M.; Saeed, M.; Patel, M. Evaluation of Anticancer, Antibacterial and Antioxidant Properties of a Medicinally Treasured Fern Tectaria coadunata with its Phytoconstituents Analysis by HR-LCMS. Anti-Cancer Agents Med. Chem. 2020, 20, 1845–1856. [Google Scholar] [CrossRef]
- Siddiqui, A.J.; Danciu, C.; Ashraf, S.A.; Moin, A.; Singh, R.; Alreshidi, M.; Patel, M.; Jahan, S.; Kumar, S.; Alkhinjar, M.I. Plants-derived biomolecules as potent antiviral phytomedicines: New insights on ethnobotanical evidences against coro-naviruses. Plants 2020, 9, 1244. [Google Scholar] [CrossRef]
- Patel, M.; Ashraf, M.S.; Siddiqui, A.J.; Ashraf, S.A.; Sachidanandan, M.; Snoussi, M.; Adnan, M.; Hadi, S. Profiling and Role of Bioactive Molecules from Puntius sophore (Freshwater/Brackish Fish) Skin Mucus with Its Potent Antibacterial, Antiadhesion, and Antibiofilm Activities. Biomolecules 2020, 10, 920. [Google Scholar] [CrossRef] [PubMed]
- Surti, M.; Patel, M.; Adnan, M.; Moin, A.; Ashraf, S.A.; Siddiqui, A.J.; Snoussi, M.; Deshpande, S.; Reddy, M.N. Ili-maquinone (marine sponge metabolite) as a novel inhibitor of SARS-CoV-2 key target proteins in comparison with sug-gested COVID-19 drugs: Designing, docking and molecular dynamics simulation study. RSC Adv. 2020, 10, 37707–37720. [Google Scholar] [CrossRef]
- Adnan, M.; Patel, M.; Deshpande, S.; Alreshidi, M.; Siddiqui, A.J.; Reddy, M.N.; Emira, N.; De Feo, V. Effect of Adiantum philippense Extract on Biofilm Formation, Adhesion with Its Antibacterial Activities Against Foodborne Pathogens, and Characterization of Bioactive Metabolites: An in vitro-in silico Approach. Front. Microbiol. 2020, 11, 823. [Google Scholar] [CrossRef]
- Mseddi, K.; Alimi, F.; Noumi, E.; Veettil, V.N.; Deshpande, S.; Adnan, M.; Hamdi, A.; Elkahoui, S.; Alghamdi, A.; Kadri, A.; et al. Thymus musilii Velen. as a promising source of potent bioactive compounds with its pharmacological properties: In vitro and in silico analysis. Arab. J. Chem. 2020, 13, 6782–6801. [Google Scholar] [CrossRef]
- Ahn, S.-H.; Mun, Y.-J.; Lee, S.-W.; Kwak, S.; Choi, M.-K.; Baik, S.-K.; Kim, Y.-M.; Woo, W.-H. Selaginella tamariscina Induces Apoptosis via a Caspase-3-Mediated Mechanism in Human Promyelocytic Leukemia Cells. J. Med. Food 2006, 9, 138–144. [Google Scholar] [CrossRef]
- Lee, I.-S.; Nishikawa, A.; Furukawa, F.; Kasahara, K.-I.; Kim, S.-U. Effects of Selaginella tamariscina on in vitro tumor cell growth, p53 expression, G1 arrest and in vivo gastric cell proliferation. Cancer Lett. 1999, 144, 93–99. [Google Scholar] [CrossRef]
- Lee, J.S.; Lee, M.S.; Oh, W.K.; Sul, J.Y. Fatty acid synthase inhibition by amentoflavone induces apoptosis and antiprolif-eration in human breast cancer cells. Biol. Pharm. Bull. 2009, 32, 1427–1432. [Google Scholar] [CrossRef] [Green Version]
- Woo, E.-R.; Pokharel, Y.R.; Yang, J.W.; Lee, S.Y.; Kang, K.W. Inhibition of nuclear factor-κB activation by 2′, 8″-biapigenin. Biol. Pharm. Bull. 2006, 29, 976–980. [Google Scholar] [CrossRef] [Green Version]
- Setyawan, A.D. Review: Natural products from Genus Selaginella (Selaginellaceae). Nusant. Biosci. 2016, 3, 3. [Google Scholar] [CrossRef]
- Yang, J.-S.; Lin, C.-W.; Hsieh, Y.-S.; Cheng, H.-L.; Lue, K.-H.; Yang, S.-F.; Lu, K.-H. Selaginella tamariscina (Beauv.) pos-sesses antimetastatic effects on human osteosarcoma cells by decreasing MMP-2 and MMP-9 secretions via p38 and Akt signaling pathways. Food Chem. Toxicol. 2013, 59, 801–807. [Google Scholar] [CrossRef]
- Yang, J.-S.; Lin, C.-W.; Hsin, C.-H.; Hsieh, M.-J.; Chang, Y.-C. Selaginella tamariscina Attenuates Metastasis via Akt Pathways in Oral Cancer Cells. PLoS ONE 2013, 8, e68035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.-J.; Duh, C.-Y.; Chen, J.-F. New cytotoxic biflavonoids from Selaginella delicatula. Planta Med. 2005, 71, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.-C.; Kuo, Y.-C.; Chou, C.-J. Cytotoxic Biflavonoids from Selaginella delicatula. J. Nat. Prod. 2000, 63, 627–630. [Google Scholar] [CrossRef]
- Sun, C.-M.; Syu, W.-J.; Huang, Y.-T.; Chen, C.-C.; Ou, J.-C. Selective cytotoxicity of ginkgetin from Selaginella moellendorffii. J. Nat. Prod. 1997, 60, 382–384. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Sun, C.-M.; Chuang, H.-H.; Chang, P.-T. Studies on the cytotoxic mechanisms of ginkgetin in a human ovarian adenocarcinoma cell line. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2000, 362, 82–90. [Google Scholar] [CrossRef]
- Sato, H.; Takino, T.; Okada, Y.; Cao, J.; Shinagawa, A.; Yamamoto, E.; Seiki, M. A matrix metalloproteinase expressed on the surface of invasive tumour cells. Nat. Cell Biol. 1994, 370, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.-B.; Hsieh, M.-J.; Hsieh, Y.-H.; Chien, M.-H.; Chiou, H.-L.; Yang, S.-F. Antimetastatic effects of norcantharidin on hepatocellular carcinoma by transcriptional inhibition of MMP-9 through modulation of NF-kB activity. PLoS ONE 2012, 7, e31055. [Google Scholar] [CrossRef]
- Ghiso, J.A.A.; Alonso, D.F.; Farías, E.F.; Gomez, D.E.; Joffè, E.B.D.K. Deregulation of the signaling pathways controlling urokinase production. Its relationship with the invasive phenotype. JBIC J. Biol. Inorg. Chem. 1999, 263, 295–304. [Google Scholar] [CrossRef]
- Hwang, E.-S.; Lee, H.J. Allyl isothiocyanate and its N-acetylcysteine conjugate suppress metastasis via inhibition of inva-sion, migration, and matrix metalloproteinase-2/-9 activities in SK-Hep1 human hepatoma cells. Exp. Biol. Med. 2006, 231, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Määttä, M.; Soini, Y.; Liakka, A.; Autio-Harmainen, H. Differential expression of matrix metalloproteinase (MMP)-2, MMP-9, and membrane type 1-MMP in hepatocellular and pancreatic adenocarcinoma: Implications for tumor progres-sion and clinical prognosis. Clin. Cancer Res. 2000, 6, 2726–2734. [Google Scholar]
- Ogata, R.; Torimura, T.; Kin, M.; Ueno, T.; Tateishi, Y.; Kuromatsu, R.; Shimauchi, Y.; Sakamoto, M.; Tamaki, S.; Sata, M.; et al. Increased expression of membrane type 1 matrix metalloproteinase and matrix metalloproteinase-2 with tumor dedifferentiation in hepatocellular carcinomas. Hum. Pathol. 1999, 30, 443–450. [Google Scholar] [CrossRef]
- Tam, E.M.; Moore, T.R.; Butler, G.S.; Overall, C.M. Characterization of the Distinct Collagen Binding, Helicase and Cleavage Mechanisms of Matrix Metalloproteinase 2 and 14 (Gelatinase A and MT1-MMP). J. Biol. Chem. 2004, 279, 43336–43344. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, T. The evolution of chemosystematics. Phytochemistry 2007, 68, 2887–2895. [Google Scholar] [CrossRef] [PubMed]
- Larsen, T.O.; Smedsgaard, J.; Nielsen, K.F.; Hansen, M.E.; Frisvad, J.C. Phenotypic taxonomy and metabolite profiling in microbial drug discovery. Nat. Prod. Rep. 2005, 22, 672–695. [Google Scholar] [CrossRef] [PubMed]
- Fellows, L.; Smith, P.M. The Chemotaxonomy of Plants. Kew Bull. 1978, 32, 806. [Google Scholar] [CrossRef]
- Hegnauer, R. Phytochemistry and plant taxonomy—An essay on the chemotaxonomy of higher plants. Phytochemistry 1986, 25, 1519–1535. [Google Scholar] [CrossRef]
- Chao, L.R.; Seguin, E.; Tillequin, F.; Koch, M. New Alkaloid Glycosides from Selaginella doederleinii. J. Nat. Prod. 1987, 50, 422–426. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Long, C.-L.; Yang, F.-M.; Wang, X.; Sun, Q.-Y.; Wang, H.-S.; Shi, Y.-N.; Tang, G.-H. Pyrrolidinoindoline Alkaloids from Selaginella moellendorfii. J. Nat. Prod. 2009, 72, 1151–1154. [Google Scholar] [CrossRef]
- Zheng, X.-K.; Bi, Y.-F.; Feng, W.-S.; Shi, S.-P.; Wang, J.-F.; Niu, J.-Z. Study on chemical constituents of Selaginella tamariscina (Beauv.) Spring. Acta Pharm. Sin. 2004, 39, 266–268. [Google Scholar]
- Zheng, X.; Liao, D.F.; Zhu, B.Y.; Tuo, Q.H.; Xu, Y.L. Study on chemical constituents of Selaginella pulvinata. Zhongcaoyao 2001, 32, 17–18. [Google Scholar]
- Bi, Y.-F.; Zheng, X.-K.; Feng, W.-S.; Shi, S.-P. Isolation and structural identification of chemical constituents from Selaginella tamariscina (Beauv.) Spring. Acta Pharm. Sin. 2004, 39, 41–45. [Google Scholar]
- Fischer, M.; Kandler, M. Identifizierung von selaginose und deren verbreitung in der gattung Selaginella. Phytochemistry 1975, 14, 2629–2633. [Google Scholar] [CrossRef]
- Moore, B.D.; Isidoro, E.; Seemann, J.R. Distribution of 2-carboxyarabinitol among plants. Phytochemistry 1993, 34, 703–707. [Google Scholar] [CrossRef]
- Ma, L.-Y.; Ma, S.-C.; Wei, F.; Lin, R.-C.; But, P.P.-H.; Lee, S.H.-S.; Lee, S.F. Uncinoside A and B, Two New Antiviral Chromone Glycosides from Selaginella uncinata. Chem. Pharm. Bull. 2003, 51, 1264–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.Y.; Wei, F.; Ma, S.C.; Lin, R.C. Two new chromone glycosides from Selaginella uncinata. Chin. Chem. Lett. 2002, 13, 748–751. [Google Scholar]
- Kunert, O.; Swamy, R.C.; Kaiser, M.; Presser, A.; Buzzi, S.; Rao, A.A.; Schühly, W. Antiplasmodial and leishmanicidal activity of biflavonoids from Indian Selaginella bryopteris. Phytochem. Lett. 2008, 1, 171–174. [Google Scholar] [CrossRef]
- Cao, Y.; Tan, N.H.; Chen, J.J.; Zeng, G.Z.; Ma, Y.B.; Wu, Y.P.; Yan, H.; Yang, J.; Lu, L.F.; Wang, Q. Bioactive flavones and biflavones from Selaginella moellendorfii Hieron. Fitoterapia 2010, 81, 253–258. [Google Scholar] [CrossRef]
- Ma, S.-C.; But, P.P.-H.; Ooi, V.E.-C.; He, Y.-H.; Lee, S.H.-S.; Lee, S.-F.; Lin, R.-C. Antiviral Amentoflavone from Selaginella sinensis. Biol. Pharm. Bull. 2001, 24, 311–312. [Google Scholar] [CrossRef] [Green Version]
- Silva, G.L.; Chai, H.; Gupta, M.P.; Farnsworth, N.R.; Cordell, G.A.; Pezzuto, J.M.; Beecher, C.W.; Kinghorn, A.D. Cytotoxic biflavonoids from Selaginella willdenowii. Phytochemistry 1995, 40, 129–134. [Google Scholar] [CrossRef]
- López-Sáez, J.A.; Alonso, M.J.P.-; Negueruela, A.V. Biflavonoids of Selaginella denticulata Growing in Spain. Z. Nat. C 1994, 49, 267–270. [Google Scholar] [CrossRef]
- Qasim, M.A.; Roy, S.K.; Kamil, M.; Ilyas, M. Phenolic constituents of Selaginellaceae. Indian J. Chem. 1985, 24, 220. [Google Scholar]
- Chakravarthy, B.K.; Rao, Y.V.; Gambhir, S.S.; Gode, K.D. Isolation of Amentoflavone from Selaginella rupestris and its Pharmacological Activity on Central Nervous System, Smooth Muscles and Isolated Frog Heart Preparations. Planta Med. 1981, 43, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Huneck, S.; Khaidav, T. Amentoflavone from Selaginella sanguinolenta. Die Pharm. 1985, 40, 431. [Google Scholar]
- Lee, I.R.; Song, J.Y.; Lee, Y.S. Cytotoxicity of folkloric medicines in murine and human cancer cells. Korean J. Pharmacogn. 1992, 23, 132–136. [Google Scholar]
- Feng, W.S.; Li, K.K.; Zheng, X.K. A new norlignan lignanoside from Selaginella moellendorfii Hieron. Acta Pharm. Sin. B 2011, 1, 36–39. [Google Scholar] [CrossRef] [Green Version]
- Zhu, T.M.; Chen, K.L.; Zhou, W.B. A new flavones glycoside from Selaginella moellendorfii Hieron. Chin. Chem. Lett. 2008, 19, 1456–1458. [Google Scholar] [CrossRef]
- Lin, L.C.; Chou, C.J. Three new biflavonoids from Selaginella delicatula. Chin. J. Pharm. 2000, 52, 211–218. [Google Scholar]
- Yang, J.W.; Pokharel, Y.R.; Kim, M.-R.; Woo, E.-R.; Choi, H.K.; Kang, K.W. Inhibition of inducible nitric oxide synthase by sumaflavone isolated from Selaginella tamariscina. J. Ethnopharmacol. 2006, 105, 107–113. [Google Scholar] [CrossRef]
- Zhang, L.-P.; Liang, Y.-M.; Wei, X.-C.; Cheng, D.-L. A New Unusual Natural Pigment from Selaginella sinensis and Its Noticeable Physicochemical Properties. J. Org. Chem. 2007, 38, 3824–3921. [Google Scholar] [CrossRef]
- Cheng, X.-L.; Ma, S.-C.; Yu, J.-D.; Yang, S.-Y.; Xiao, X.-Y.; Hu, J.-Y.; Lu, Y.; Shaw, P.-C.; But, P.P.-H.; Lin, R.-C. Selaginellin A and B, Two Novel Natural Pigments Isolated from Selaginella tamariscina. Chem. Pharm. Bull. 2008, 56, 982–984. [Google Scholar] [CrossRef] [Green Version]
- Tan, G.-S.; Xu, K.-P.; Li, F.-S.; Wang, C.-J.; Li, T.-Y.; Hu, C.-P.; Shen, J.; Zhou, Y.-J.; Li, Y.-J. Selaginellin C, a new natural pigment from Selaginella pulvinata Maxim (Hook et Grev.). J. Asian Nat. Prod. Res. 2009, 11, 1001–1004. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Chen, J.J.; Tan, N.H.; Oberer, L.; Wagner, T.; Wu, Y.P.; Zeng, G.Z.; Yan, H.; Wang, Q. Antimicrobial selaginellin derivatives from Selaginella pulvinata. Bioorganic Med. Chem. Lett. 2010, 20, 2456–2460. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Yang, J.; Wang, Q.; Chen, J.-J.; Tan, N.-H.; Wu, Y.-P. Structure determination of selaginellins G and H from Selaginella pulvinata by NMR spectroscopy. Magn. Reson. Chem. 2010, 48, 656–659. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-F.; Xu, K.-P.; Li, F.-S.; Shen, J.; Hu, C.-P.; Zou, H.; Yang, F.; Liu, G.-R.; Xiang, H.-L.; Zhou, Y.-J.; et al. A New Flavonoid from Selaginella tamariscina (Beauv.) Spring. Chem. Pharm. Bull. 2010, 58, 549–551. [Google Scholar] [CrossRef] [Green Version]
- Chiu, P.-L.; Patterson, G.W.; Salt, T.A. Sterol composition of pteridophytes. Phytochemistry 1988, 27, 819–822. [Google Scholar] [CrossRef]
- Zheng, X.; Du, J.; Xu, Y.; Zhu, B.; Liao, D. A new steroid from Selaginella pulvinata. Fitoterapia 2007, 78, 598–599. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Sun, J.Y.; Xie, N.G.; Shi, Y.G. Chemical constituents of daeycai (Selaginella doederleinii). Zhongcaoyao 1995, 26, 397–399. [Google Scholar]
- Che, D.H.; Yu, J.G. Analysis on the chemical constituents of jiangnanjuanbai (Selaginella moellendorfii Hieron). Chung Tsao Yao 1986, 17, 4. [Google Scholar]
- Baraya, Y.S.; Wong, K.K.; Yaacob, N.S. Strobilanthes crispus inhibits migration, invasion and metastasis in breast cancer. J. Ethnopharmacol. 2019, 233, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Justus, C.R.; Leffler, N.; Ruiz-Echevarria, M.; Yang, L.V. In vitro Cell Migration and Invasion Assays. J. Vis. Exp. 2014, e51046. [Google Scholar] [CrossRef] [Green Version]
- Burg-Roderfeld, M.; Roderfeld, M.; Wagner, S.; Henkel, C.; Grötzinger, J.; Roeb, E. MMP-9-hemopexin domain hampers adhesion and migration of colorectal cancer cells. Int. J. Oncol. 2007, 30, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Adnan, M.; Morton, G.; Hadi, S. Analysis of rpoS and bolA gene expression under various stress-induced environments in planktonic and biofilm phase using 2−ΔΔCT method. Mol. Cell. Biochem. 2011, 357, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, A.J.; Bhardwaj, J.; Goyal, M.; Prakash, K.; Adnan, M.; Alreshidi, M.M.; Patel, M.; Soni, A.; Redman, W. Immune responses in liver and spleen against Plasmodium yoelii pre-erythrocytic stages in Swiss mice model. J. Adv. Res. 2020, 24, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Byambaragchaa, M.; De La Cruz, J.; Yang, S.H.; Hwang, S.-G. Anti-metastatic Potential of Ethanol Extract of Saussurea involucrata against Hepatic Cancer in vitro. Asian Pac. J. Cancer Prev. 2013, 14, 5397–5402. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytocompounds | Formula | Class | m/z | RT (min) | Mass |
|---|---|---|---|---|---|
| Urocanic acid | C6H6N2O2 | monocarboxylic acid | 131.2 | 24.845 | 138.04267 |
| Hordenine | C10H15NO | alkaloid | 167.2 | 23.071 | 169.23958 |
| Oleamide | C18H35NO | fatty acid | 286.1 | 20.177 | 281.27135 |
| Hexadecanamide | C16H33NO | fatty acid amide | 250.4 | 19.849 | 255.25575 |
| Arachidonic acid | C20H32O2 | polyunsaturated fatty acid | 301.8 | 18.601 | 304.2397 |
| 4-Coumaric acid | C9H8O3 | phenol | 168.2 | 18.265 | 164.04718 |
| 2-Arachidonoyl glycerol | C23H38O4 | fatty acid derivative | 371.0 | 16.012 | 378.27635 |
| Valine | C5H11NO2 | amino acid | 118.4 | 13.901 | 117.07901 |
| Genistein | C15H10O5 | isoflavone | 275.3 | 13.542 | 270.05214 |
| Diosmetin | C16H12O6 | flavonoid | 294.3 | 13.534 | 300.06279 |
| 2-Amino-1,3,4-octadecanetriol | C18H39NO3 | phenol | 311.5 | 13.499 | 317.29231 |
| Glycitein | C16H12O5 | flavonoid | 288.9 | 13.477 | 284.06807 |
| Rhamnetin | C16H12O7 | flavonoid | 311.2 | 12.452 | 316.05766 |
| Formononetin | C16H12O4 | phenol | 265.3 | 12.157 | 268.07329 |
| Luteolin | C15H10O6 | flavonoid | 278.6 | 11.735 | 286.04723 |
| Apigenin | C15H10O5 | flavone | 272.4 | 11.177 | 270.05235 |
| Glycitin | C22H22O10 | isoflavone | 442.2 | 11.088 | 446.12065 |
| (-)-Caryophyllene oxide | C15H24O | epoxide | 228.4 | 10.861 | 220.18227 |
| Kuromanin | C21H20O11 | pigment | 458.1 | 8.976 | 448.09996 |
| Kaempferol | C15H10O6 | flavonoid | 294.6 | 8.662 | 286.04726 |
| Sedanolide | C12H18O2 | isobenzofuran | 199.5 | 8.578 | 194.13039 |
| α-Pinene-2-oxide | C10H16O | terpenoid | 148.9 | 8.494 | 152.11989 |
| Quercetin-3β-D-glucoside | C21H20O12 | flavonoid | 260.3 | 8.438 | 464.09465 |
| Quercetin | C15H10O7 | flavonoid | 308.6 | 8.414 | 302.04192 |
| Vitexin | C21H20O10 | flavonoid | 436.8 | 8.319 | 432.10497 |
| Rutin | C27H30O16 | flavonoid | 615.4 | 8.291 | 610.15239 |
| 4-Methoxycinnamic acid | C10H10O3 | phenol | 186.2 | 7.754 | 178.06275 |
| Norharman | C11H8N2 | alkaloid | 164.7 | 6.725 | 168.06847 |
| 4-Hydroxycoumarin | C9H6O3 | benzopyrone | 158.8 | 6.568 | 162.0314 |
| Methyl cinnamate | C10H10O2 | cinnamic acid ester | 164.5 | 6.458 | 162.06775 |
| Isoferulic acid | C10H10O4 | phenol | 198.6 | 6.367 | 194.05762 |
| Scopoletin | C10H8O4 | coumarin | 196.3 | 5.969 | 192.04198 |
| Citral | C10H16O | terpenoid | 145.8 | 5.575 | 152.11989 |
| Pulegone | C10H16O | terpenoid | 158.9 | 4.94 | 152.11989 |
| Caffeic acid | C9H8O4 | phenol | 186.2 | 4.692 | 180.04181 |
| 7-Hydroxycoumarine | C9H6O3 | phenol | 169.4 | 4.651 | 162.0314 |
| Chlorogenic acid | C16H18O9 | phenol | 360.5 | 4.646 | 354.09435 |
| Kynurenic acid | C10H7NO3 | quinoline carboxylic acid | 181.7 | 3.823 | 189.04239 |
| Coumarin | C9H6O2 | phenol | 148.6 | 3.784 | 146.0365 |
| 3-Methylcrotonylglycine | C7H11NO3 | amino acid | 152.6 | 3.325 | 157.0737 |
| 4-Hydroxyphenylacetic acid | C8H8O3 | benzenoid | 156.3 | 3.314 | 152.04714 |
| 8-Hydroxyquinoline | C9H7NO | alkaloid | 153.2 | 2.965 | 145.05255 |
| Maltol | C6H6O3 | sugar | 130.4 | 2.278 | 126.03161 |
| Guvacoline | C7H11NO2 | pyridine alkaloid | 145.2 | 1.539 | 141.07878 |
| L-Phenylalanine | C9H11NO2 | amino acid | 164.2 | 1.375 | 165.07883 |
| L-Norleucine | C6H13NO2 | amino acid | 136.5 | 1.134 | 131.09453 |
| L-Pyroglutamic acid | C5H7NO3 | amino acid | 133.1 | 1.04 | 129.0425 |
| D-Glucosamine | C6 H13 NO5 | amino sugar | 183.3 | 0.946 | 179.079 |
| Betaine | C5H11NO2 | amino acid | 122.2 | 0.935 | 117.07901 |
| L(-)-Carnitine | C7H15NO3 | amino acid derivative | 157.6 | 0.93 | 161.10489 |
| Acetylcholine | C7H15 NO2 | essential nutrient (vitamin) | 149.7 | 0.850 | 145.11 |
| α-Lactose | C12H22O11 | sugar | 349.9 | 0.839 | 342.11521 |
| Choline | C5H13N O | essential nutrient (vitamin) | 111.4 | 0.798 | 103.09988 |
| Sl. No | Primer | Sequence |
|---|---|---|
| 1 | MMP-2 | sense–5′-GGCCCTGTCACTCCTGAGAT-3′ antisense–5′-GGCATCCAGGTTATCGGGGA-3′ |
| 2 | MMP-9 | sense–5′- CGGAGCACGGAGACGGGTAT-3′ antisense–5′- TGAAGGGGAAGACGCACAGC-3′ |
| 3 | MT1-MMP | sense–5′-TGGGTAGCGATGAAGTCTTC-3′ antisense–5′-AGTAAAGCAGTCGCTTGGGT-3′ |
| 4 | TIMP-1 | sense–5′- GATCCAGCGCCCAGAGAGACACC-3′ antisense–5′-TTCCACTCCGGGCAGCATT-3′ |
| 5 | GAPDH | sense–5′- CGAGATCCCTCCAAAATCAA-3′ antisense–5′-AGGTCCACCACTGACACGTT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adnan, M.; Siddiqui, A.J.; Hamadou, W.S.; Snoussi, M.; Badraoui, R.; Ashraf, S.A.; Jamal, A.; Awadelkareem, A.M.; Sachidanandan, M.; Hadi, S.; et al. Deciphering the Molecular Mechanism Responsible for Efficiently Inhibiting Metastasis of Human Non-Small Cell Lung and Colorectal Cancer Cells Targeting the Matrix Metalloproteinases by Selaginella repanda. Plants 2021, 10, 979. https://doi.org/10.3390/plants10050979
Adnan M, Siddiqui AJ, Hamadou WS, Snoussi M, Badraoui R, Ashraf SA, Jamal A, Awadelkareem AM, Sachidanandan M, Hadi S, et al. Deciphering the Molecular Mechanism Responsible for Efficiently Inhibiting Metastasis of Human Non-Small Cell Lung and Colorectal Cancer Cells Targeting the Matrix Metalloproteinases by Selaginella repanda. Plants. 2021; 10(5):979. https://doi.org/10.3390/plants10050979
Chicago/Turabian StyleAdnan, Mohd, Arif Jamal Siddiqui, Walid Sabri Hamadou, Mejdi Snoussi, Riadh Badraoui, Syed Amir Ashraf, Arshad Jamal, Amir Mahgoub Awadelkareem, Manojkumar Sachidanandan, Sibte Hadi, and et al. 2021. "Deciphering the Molecular Mechanism Responsible for Efficiently Inhibiting Metastasis of Human Non-Small Cell Lung and Colorectal Cancer Cells Targeting the Matrix Metalloproteinases by Selaginella repanda" Plants 10, no. 5: 979. https://doi.org/10.3390/plants10050979