
The Relevance of a Physiological-Stage Approach Study of the Molecular and Environmental Factors Regulating Seed Germination in Wild Plants

 ,
,  ,
,
Abstract
:1. Introduction
2. Molecular Bases of Seed Maturation
2.1. Accumulation of Reserves
2.2. Tolerance to Desiccation
3. Molecular Bases of Dormancy
3.1. Regulation of Dormancy: The Role of Hormones and Dormancy-Specific Regulators
3.2. Dormancy Break by Different Treatments
4. Molecular Bases of Germination
5. Domestication, Seed Management and the Need for Rewilding Seed Science

{kind=link}
| Species | Gene Name | Characteristic | Type/Function | Type of Mutation | Domestication Phase | Reference |
|---|---|---|---|---|---|---|
| Brassica rapa | TT8 | Lighter-colored testa | Transcription factor | Insertion, LOF | Diversification and dispersion | [85] |
| Glycine max | GmG | Reduced dormancy, lighter-colored testa | CAAX amino-terminal protease protein | Splicing defect, premature stop | Onset of domestication | [76] |
| Oryza sativa | qSD7-1/qPC7 | Dormancy | Transcription factor | Deletion | Onset of domestication | [96,97] |
| Oryza sativa | Sdr4 | Reduced dormancy | Not characterized | Insertion | Onset of domestication | |
| Oryza sativa | OsG | Reduced dormancy | CAAX amino-terminal protease protein | Non-synonymous SNP | Onset of domestication | [76] |
| Oryza sativa | GIF1 | Reserve accumulation | Cell wall invertase | Cis-regulatory, restricted expression | Onset of domestication | [98] |
| Oryza sativa | Bh4 | Seed color | Amino acid transporter | Deletion/LOF, premature stop | Onset of domestication | [99] |
| Oryza sativa | GS3 | Seed size and weight increase | Transmembranal protein | Premature stop | Diversification and dispersion | [17] |
| Oryza sativa | OsGRF4 | Seed size increase | Transcription factor | Mutation in regulator target site | Variety improvement | [100] |
| Oryza sativa | GS5 | Seed size increase | Serine-carboxypeptidase | Indels, cis-regulatory | Diversification and improvement | [101] |
| Oryza sativa | GW2 | Seed size increase | E3-ubiquitin ligase | Deletion, premature stop | Unknown | [15] |
| Oryza sativa | GW6a | Seed size/weight increase, plant biomass | Histone H4-acetyltransferase, transcription regulator | Cis-regulatory | Variety improvement | [102] |
| Oryza sativa | Ae1 | Amylose properties | Alpha-amylase | SNPs | Onset of domestication | [103] |
| Panicum miliaceum | PmGBSSI | Starch properties | Granule-bound starch synthase | Deletion, LOF, frameshift, missense | Diversification and dispersion | [85] |
| Setaria italica | WAXY | Starch properties | Granule-bound starch synthase | Insertion LOF | Diversification and dispersion | [104] |
| Solanum lycopersicum | SolyG | Reduced dormancy | CAAX amino-terminal protease protein | Non-synonymous SNP | Onset of domestication | [76] |
| Triticum aestivum | WAP2 (Q) | Reduced seed shattering, plant height and other pleiotropic effects | Transcription factor | Missense, cis-regulatory, premature stop | Onset of domestication | [105] |
| Zea mays | Bt2 (SSU) | Amylopectin properties | ADP-glucose pyrophosphorylase | Signal peptide, expression site | Onset of domestication and diversification | [106] and references therein |
| Zea mays | Vgt1 | Flowering time QTL | AP2-like transcription factor | Cis-regulatory | Diversification and dispersion | [107] |
| Zea mays | HEX9 | Glycolysis pathway, trans eQTL effects | Hexokinase 9 | Cis-regulatory | Not determined | [76] |
| Zea mays | Adh2 | Resistance to hypoxia | Alcohol dehydrogenase | Small tandem repeats, indels | Onset of domestication | [108] |
| Zea mays | Su1 | Starch biosynthesis, sweetness | Isoamylase | Insertion LOF, missense, premature stop | Onset of domestication | [108] |
| Zea mays | DULL1 | Starch synthesis | soluble starch synthase | Insertion, reduced expression | Diversification | [94] |
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds; Springer: New York, NY, USA, 2013; ISBN 978-1-4614-4692-7. [Google Scholar]
- Fenner, M.; Thompson, K. The Ecology of Seeds, 1st ed.; Cambridge University Press: Cambridge, UK, 2005; ISBN 978-0-521-65311-4. [Google Scholar]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology; Springer: New York, NY, USA, 2008; ISBN 978-0-387-78340-6. [Google Scholar]
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed Dormancy and the Control of Germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef]
- Long, R.L.; Gorecki, M.J.; Renton, M.; Scott, J.K.; Colville, L.; Goggin, D.E.; Commander, L.E.; Westcott, D.A.; Cherry, H.; Finch-Savage, W.E. The Ecophysiology of Seed Persistence: A Mechanistic View of the Journey to Germination or Demise: The Ecophysiology of Seed Persistence. Biol. Rev. 2015, 90, 31–59. [Google Scholar] [CrossRef]
- McDonald, M.B.; Copeland, L.O. Seed Formation and Development. In Seed Production; Springer: Boston, MA, USA, 1997; pp. 19–28. ISBN 978-1-4613-6825-0. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Academic Press: San Diego, CA, USA, 1998; ISBN 978-0-12-080260-9. [Google Scholar]
- Miernyk, J.A. Seed Proteomics. In Plant Proteomics, Methods in Molecular Biology (Methods and Protocols); Jorrin-Novo, J.V., Komatsu, S., Weckwerth, W., Wienkoop, S., Eds.; Humana Press: Totowa, NJ, USA, 2014; Volume 1072, pp. 361–377. ISBN 978-1-62703-630-6. [Google Scholar]
- He, H.; Willems, L.A.J.; Batushansky, A.; Fait, A.; Hanson, J.; Nijveen, H.; Hilhorst, H.W.M.; Bentsink, L. Effects of Parental Temperature and Nitrate on Seed Performance Are Reflected by Partly Overlapping Genetic and Metabolic Pathways. Plant Cell Physiol. 2016, 57, 473–487. [Google Scholar] [CrossRef] [Green Version]
- Farnsworth, E. The Ecology and Physiology of Viviparous and Recalcitrant Seeds. Annu. Rev. Ecol. Syst. 2000, 31, 107–138. [Google Scholar] [CrossRef]
- Gillman, J.D.; Biever, J.J.; Ye, S.; Spollen, W.G.; Givan, S.A.; Lyu, Z.; Joshi, T.; Smith, J.R.; Fritschi, F.B. A Seed Germination Transcriptomic Study Contrasting Two Soybean Genotypes That Differ in Terms of Their Tolerance to the Deleterious Impacts of Elevated Temperatures during Seed Fill. BMC Res. Notes 2019, 12, 522. [Google Scholar] [CrossRef]
- Carbonero, P.; Iglesias-Fernández, R.; Vicente-Carbajosa, J. The AFL Subfamily of B3 Transcription Factors: Evolution and Function in Angiosperm Seeds. J. Exp. Bot. 2017, 68, 871–880. [Google Scholar] [CrossRef]
- Liu, H.; Liang, X.; Lu, Q.; Li, H.; Liu, H.; Li, S.; Varshney, R.; Hong, Y.; Chen, X. Global Transcriptome Analysis of Subterranean Pod and Seed in Peanut (Arachis hypogaea L.) Unravels the Complexity of Fruit Development under Dark Condition. Sci. Rep. 2020, 10, 13050. [Google Scholar] [CrossRef]
- Liu, S.-S.; Chen, J.; Li, S.-C.; Zeng, X.; Meng, Z.-X.; Guo, S.-X. Comparative Transcriptome Analysis of Genes Involved in GA-GID1-DELLA Regulatory Module in Symbiotic and Asymbiotic Seed Germination of Anoectochilus roxburghii (Wall.) Lindl. (Orchidaceae). Int. J. Mol. Sci. 2015, 16, 30190–30203. [Google Scholar] [CrossRef] [Green Version]
- Song, X.-J.; Huang, W.; Shi, M.; Zhu, M.-Z.; Lin, H.-X. A QTL for Rice Grain Width and Weight Encodes a Previously Unknown RING-Type E3 Ubiquitin Ligase. Nat. Genet. 2007, 39, 623–630. [Google Scholar] [CrossRef]
- Shomura, A.; Izawa, T.; Ebana, K.; Ebitani, T.; Kanegae, H.; Konishi, S.; Yano, M. Deletion in a Gene Associated with Grain Size Increased Yields during Rice Domestication. Nat. Genet. 2008, 40, 1023–1028. [Google Scholar] [CrossRef]
- Yan, S.; Zou, G.; Li, S.; Wang, H.; Liu, H.; Zhai, G.; Guo, P.; Song, H.; Yan, C.; Tao, Y. Seed Size Is Determined by the Combinations of the Genes Controlling Different Seed Characteristics in Rice. Theor. Appl. Genet. 2011, 123, 1173–1181. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.; Buijs, G.; Ligterink, W.; Hilhorst, H. Evolutionary Ecophysiology of Seed Desiccation Sensitivity. Funct. Plant Biol. 2018, 45, 1083–1095. [Google Scholar] [CrossRef]
- Wyse, S.V.; Dickie, J.B. Predicting the Global Incidence of Seed Desiccation Sensitivity. J. Ecol. 2017, 105, 1082–1093. [Google Scholar] [CrossRef] [Green Version]
- Berjak, P.; Pammenter, N.W. From Avicennia to Zizania: Seed Recalcitrance in Perspective. Ann. Bot. 2008, 101, 213–228. [Google Scholar] [CrossRef]
- Dickie, J.B.; Pritchard, H.W. Systematic and evolutionary aspects of desiccation tolerance in seeds. In Desiccation and Survival in Plants: Drying without Dying; Black, M., Pritchard, H.W., Eds.; CABI: Wallingford, UK, 2002; pp. 239–259. ISBN 978-0-85199-534-2. [Google Scholar]
- Delahaie, J.; Hundertmark, M.; Bove, J.; Leprince, O.; Rogniaux, H.; Buitink, J. LEA Polypeptide Profiling of Recalcitrant and Orthodox Legume Seeds Reveals ABI3-Regulated LEA Protein Abundance Linked to Desiccation Tolerance. J. Exp. Bot. 2013, 64, 4559–4573. [Google Scholar] [CrossRef]
- Artur, M.A.S.; Zhao, T.; Ligterink, W.; Schranz, E.; Hilhorst, H.W.M. Dissecting the Genomic Diversification of Late Embryogenesis Abundant (LEA) Protein Gene Families in Plants. Genome Biol. Evol. 2019, 11, 459–471. [Google Scholar] [CrossRef]
- Kijak, H.; Ratajczak, E. What Do We Know About the Genetic Basis of Seed Desiccation Tolerance and Longevity? Int. J. Mol. Sci. 2020, 21, 3612. [Google Scholar] [CrossRef]
- Miguel, A.; de Vega-Bartol, J.; Marum, L.; Chaves, I.; Santo, T.; Leitão, J.; Varela, M.C.; Miguel, C.M. Characterization of the Cork Oak Transcriptome Dynamics during Acorn Development. BMC Plant Biol. 2015, 15, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Rodríguez, M.C.; Jorrín-Novo, J.V.; Castillejo, M.A. Toward Characterizing Germination and Early Growth in the Non-Orthodox Forest Tree Species Quercus ilex through Complementary Gel and Gel-Free Proteomic Analysis of Embryo and Seedlings. J. Proteom. 2019, 197, 60–70. [Google Scholar] [CrossRef]
- Sghaier-Hammami, B.; Hammami, S.B.M.; Baazaoui, N.; Gómez-Díaz, C.; Jorrín-Novo, J.V. Dissecting the Seed Maturation and Germination Processes in the Non-Orthodox Quercus ilex Species Based on Protein Signatures as Revealed by 2-DE Coupled to MALDI-TOF/TOF Proteomics Strategy. IJMS 2020, 21, 4870. [Google Scholar] [CrossRef]
- Hilhorst, H.W.M. Definitions and Hypotheses of Seed Dormancy. In Seed Development, Dormancy and Germination; Bradford, K.J., Nonogaki, H., Eds.; Blackwell Publishing Ltd: Oxford, UK, 2007; pp. 50–71. ISBN 978-0-470-98884-8. [Google Scholar]
- Benech-Arnold, R.L.; Sánchez, R.A.; Forcella, F.; Kruk, B.C.; Ghersa, C.M. Environmental Control of Dormancy in Weed Seed Banks in Soil. Field Crops Res. 2000, 67, 105–122. [Google Scholar] [CrossRef]
- Yao, Z.; Tian, F.; Cao, X.; Xu, Y.; Chen, M.; Xiang, B.; Zhao, S. Global Transcriptomic Analysis Reveals the Mechanism of Phelipanche aegyptiaca Seed Germination. IJMS 2016, 17, 1139. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liu, S.; Yuan, G.; Zhao, P.; Yang, W.; Jia, J.; Cheng, L.; Qi, D.; Chen, S.; Liu, G. Comparative Transcriptome Analysis Provides Insights into the Distinct Germination in Sheepgrass (Leymus chinensis) during Seed Development. Plant Physiol. Biochem. 2019, 139, 446–458. [Google Scholar] [CrossRef]
- Chahtane, H.; Kim, W.; Lopez-Molina, L. Primary Seed Dormancy: A Temporally Multilayered Riddle Waiting to Be Unlocked. EXBOTJ 2016, erw377. [Google Scholar] [CrossRef]
- Penfield, S. Seed Dormancy and Germination. Curr. Biol. 2017, 27, R874–R878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baskin, J.M.; Baskin, C.C. Evolutionary Considerations of Claims for Physical Dormancy-Break by Microbial Action and Abrasion by Soil Particles. Seed Sci. Res. 2000, 10, 409–413. [Google Scholar] [CrossRef]
- Gama-Arachchige, N.S.; Baskin, J.M.; Geneve, R.L.; Baskin, C.C. Identification and Characterization of Ten New Water Gaps in Seeds and Fruits with Physical Dormancy and Classification of Water-Gap Complexes. Ann. Bot. 2013, 112, 69–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, H.; de Souza Vidigal, D.; Snoek, L.B.; Schnabel, S.; Nijveen, H.; Hilhorst, H.; Bentsink, L. Interaction between Parental Environment and Genotype Affects Plant and Seed Performance in Arabidopsis. J. Exp. Bot. 2014, 65, 6603–6615. [Google Scholar] [CrossRef] [Green Version]
- Soppe, W.J.J.; Bentsink, L. Seed Dormancy Back on Track; Its Definition and Regulation by DOG1. New Phytol. 2020, 228, 816–819. [Google Scholar] [CrossRef] [Green Version]
- Graeber, K.; Linkies, A.; Steinbrecher, T.; Mummenhoff, K.; Tarkowská, D.; Turečková, V.; Ignatz, M.; Sperber, K.; Voegele, A.; de Jong, H.; et al. DELAY OF GERMINATION 1 Mediates a Conserved Coat-Dormancy Mechanism for the Temperature- and Gibberellin-Dependent Control of Seed Germination. Proc. Natl. Acad. Sci. USA 2014, 111, E3571. [Google Scholar] [CrossRef] [Green Version]
- Née, G.; Xiang, Y.; Soppe, W.J. The Release of Dormancy, a Wake-up Call for Seeds to Germinate. Curr. Opin. Plant Biol. 2017, 35, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Carrera-Castaño, G.; Calleja-Cabrera, J.; Pernas, M.; Gómez, L.; Oñate-Sánchez, L. An Updated Overview on the Regulation of Seed Germination. Plants 2020, 9, 703. [Google Scholar] [CrossRef]
- Ashikawa, I.; Mori, M.; Nakamura, S.; Abe, F. A Transgenic Approach to Controlling Wheat Seed Dormancy Level by Using Triticeae DOG1-like Genes. Transgenic Res. 2014, 23, 621–629. [Google Scholar] [CrossRef]
- Huo, H.; Wei, S.; Bradford, K.J. DELAY OF GERMINATION1 (DOG1) Regulates Both Seed Dormancy and Flowering Time through MicroRNA Pathways. Proc. Natl. Acad. Sci. USA 2016, 113, E2199. [Google Scholar] [CrossRef] [Green Version]
- BLAST. National Library of Medicine (US) National Center for Biotechnology Information Bethesda (MD). 2004. Available online: https//ncbi.nlm.nih.gov/gene/ (accessed on 18 May 2021).
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Barrero, J.M.; Jacobsen, J.V.; Talbot, M.J.; White, R.G.; Swain, S.M.; Garvin, D.F.; Gubler, F. Grain Dormancy and Light Quality Effects on Germination in the Model Grass Brachypodium distachyon. New Phytol. 2012, 193, 376–386. [Google Scholar] [CrossRef]
- Cantoro, R.; Crocco, C.D.; Benech-Arnold, R.L.; Rodríguez, M.V. In Vitro Binding of Sorghum bicolor Transcription Factors ABI4 and ABI5 to a Conserved Region of a GA 2-OXIDASE Promoter: Possible Role of This Interaction in the Expression of Seed Dormancy. J. Exp. Bot. 2013, 64, 5721–5735. [Google Scholar] [CrossRef] [Green Version]
- Topham, A.T.; Taylor, R.E.; Yan, D.; Nambara, E.; Johnston, I.G.; Bassel, G.W. Temperature Variability Is Integrated by a Spatially Embedded Decision-Making Center to Break Dormancy in Arabidopsis Seeds. Proc. Natl. Acad. Sci. USA 2017, 114, 6629–6634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holdsworth, M.J.; Bentsink, L.; Soppe, W.J.J. Molecular Networks Regulating Arabidopsis Seed Maturation, After-ripening, Dormancy and Germination. New Phytol. 2008, 179, 33–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Li, W.; Fang, C.; Xu, F.; Liu, Y.; Wang, Z.; Yang, R.; Zhang, M.; Liu, S.; Lu, S.; et al. Parallel Selection on a Dormancy Gene during Domestication of Crops from Multiple Families. Nat. Genet. 2018, 50, 1435–1441. [Google Scholar] [CrossRef] [PubMed]
- Gubler, F.; Millar, A.A.; Jacobsen, J.V. Dormancy Release, ABA and Pre-Harvest Sprouting. Curr. Opin. Plant Biol. 2005, 8, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Tai, L.; Wang, H.-J.; Xu, X.-J.; Sun, W.-H.; Ju, L.; Liu, W.-T.; Li, W.-Q.; Sun, J.; Chen, K.-M. Pre-Harvest Sprouting in Cereals: Genetic and Biochemical Mechanisms. J. Exp. Bot. 2021, 72, 2857–2876. [Google Scholar] [CrossRef]
- Wu, Q.; Bai, X.; Wu, X.; Xiang, D.; Wan, Y.; Luo, Y.; Shi, X.; Li, Q.; Zhao, J.; Qin, P.; et al. Transcriptome Profiling Identifies Transcription Factors and Key Homologs Involved in Seed Dormancy and Germination Regulation of Chenopodium quinoa. Plant Physiol. Biochem. 2020, 151, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Xia, Q.; Saux, M.; Ponnaiah, M.; Gilard, F.; Perreau, F.; Huguet, S.; Balzergue, S.; Langlade, N.; Bailly, C.; Meimoun, P.; et al. One Way to Achieve Germination: Common Molecular Mechanism Induced by Ethylene and After-Ripening in Sunflower Seeds. IJMS 2018, 19, 2464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huarte, H.R.; Puglia, G.; Prjibelski, A.; Raccuia, S.A. Seed Transcriptome Annotation Reveals Enhanced Expression of Genes Related to ROS Homeostasis and Ethylene Metabolism at Alternating Temperatures in Wild Cardoon. Plants 2020, 9, 1225. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Sun, P.; Liao, D.; Sun, T.; Zhu, J.; Li, X. Transcriptomic Analysis of American Ginseng Seeds during the Dormancy Release Process by RNA-Seq. PLoS ONE 2015, 10, e0118558. [Google Scholar] [CrossRef]
- Cao, D.; Xu, H.; Zhao, Y.; Deng, X.; Liu, Y.; Soppe, W.J.J.; Lin, J. Transcriptome and Degradome Sequencing Reveals Dormancy Mechanisms of Cunninghamia lanceolata Seeds. Plant Physiol. 2016, 172, 2347. [Google Scholar] [CrossRef] [Green Version]
- Jia, Z.; Zhao, B.; Liu, S.; Lu, Z.; Chang, B.; Jiang, H.; Cui, H.; He, Q.; Li, W.; Jin, B.; et al. Embryo Transcriptome and MiRNA Analyses Reveal the Regulatory Network of Seed Dormancy in Ginkgo biloba. Tree Physiol. 2021, 41, 571–588. [Google Scholar] [CrossRef]
- Song, Q.; Cheng, S.; Chen, Z.; Nie, G.; Xu, F.; Zhang, J.; Zhou, M.; Zhang, W.; Liao, Y.; Ye, J. Comparative Transcriptome Analysis Revealing the Potential Mechanism of Seed Germination Stimulated by Exogenous Gibberellin in Fraxinus hupehensis. BMC Plant Biol. 2019, 19, 199. [Google Scholar] [CrossRef] [Green Version]
- Rana, B.; Sreenivasulu, Y. Protein Changes during Ethanol Induced Seed Germination in Aconitum heterophyllum. Plant Sci. 2013, 198, 27–38. [Google Scholar] [CrossRef]
- Qu, C.; Zuo, Z.; Cao, L.; Huang, J.; Sun, X.; Zhang, P.; Yang, C.; Li, L.; Xu, Z.; Liu, G. Comprehensive Dissection of Transcript and Metabolite Shifts during Seed Germination and Post-Germination Stages in Poplar. BMC Plant Biol. 2019, 19, 279. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Maqueo, X.; Soriano, D.; Velázquez-Rosas, N.; Alvarado-López, S.; Jiménez-Durán, K.; del Mar Garciadiego, M.; Gamboa-deBuen, A. The Seed Water Content as a Time-Independent Physiological Trait during Germination in Wild Tree Species Such as Ceiba aesculifolia. Sci. Rep. 2020, 10, 10429. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Tian, M.; Wang, C.; Zhang, Y. Transcriptome Sequencing and Differential Gene Expression Analysis Reveal the Mechanisms Involved in Seed Germination and Protocorm Development of Calanthe tsoongiana. Gene 2021, 772, 145355. [Google Scholar] [CrossRef]
- Dekkers, B.J.W.; Pearce, S.; van Bolderen-Veldkamp, R.P.; Marshall, A.; Widera, P.; Gilbert, J.; Drost, H.-G.; Bassel, G.W.; Müller, K.; King, J.R.; et al. Transcriptional Dynamics of Two Seed Compartments with Opposing Roles in Arabidopsis Seed Germination. Plant Physiol. 2013, 163, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Liu, S.S.; Kohler, A.; Yan, B.; Luo, H.M.; Chen, X.M.; Guo, S.X. ITRAQ and RNA-Seq Analyses Provide New Insights into Regulation Mechanism of Symbiotic Germination of Dendrobium officinale Seeds (Orchidaceae). J. Proteome Res. 2017, 16, 2174–2187. [Google Scholar] [CrossRef]
- Waterworth, W.M.; Bray, C.M.; West, C.E. The Importance of Safeguarding Genome Integrity in Germination and Seed Longevity. J. Exp. Bot. 2015, 66, 3549–3558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howell, K.A.; Narsai, R.; Carroll, A.; Ivanova, A.; Lohse, M.; Usadel, B.; Millar, A.H.; Whelan, J. Mapping Metabolic and Transcript Temporal Switches during Germination in Rice Highlights Specific Transcription Factors and the Role of RNA Instability in the Germination Process. Plant Physiol. 2009, 149, 961–980. [Google Scholar] [CrossRef] [Green Version]
- Tognacca, R.S.; Servi, L.; Hernando, C.E.; Saura-Sanchez, M.; Yanovsky, M.J.; Petrillo, E.; Botto, J.F. Alternative Splicing Regulation During Light-Induced Germination of Arabidopsis thaliana Seeds. Front. Plant Sci. 2019, 10, 1076. [Google Scholar] [CrossRef]
- Xu, H.; Lantzouni, O.; Bruggink, T.; Benjamins, R.; Lanfermeijer, F.; Denby, K.; Schwechheimer, C.; Bassel, G.W. A Molecular Signal Integration Network Underpinning Arabidopsis Seed Germination. Curr. Biol. 2020, 30, 3703–3712.e4. [Google Scholar] [CrossRef]
- Mérai, Z.; Graeber, K.; Wilhelmsson, P.; Ullrich, K.K.; Arshad, W.; Grosche, C.; Tarkowská, D.; Turečková, V.; Strnad, M.; Rensing, S.A.; et al. Aethionema arabicum: A Novel Model Plant to Study the Light Control of Seed Germination. J. Exp. Bot. 2019, 70, 3313–3328. [Google Scholar] [CrossRef] [PubMed]
- Nelson, E.B. The Seed Microbiome: Origins, Interactions, and Impacts. Plant Soil 2018, 422, 7–34. [Google Scholar] [CrossRef]
- Shade, A.; Jacques, M.-A.; Barret, M. Ecological Patterns of Seed Microbiome Diversity, Transmission, and Assembly. Curr. Opin. Microbiol. 2017, 37, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Gundel, P.E.; Rudgers, J.A.; Ghersa, C.M. Incorporating the Process of Vertical Transmission into Understanding of Host–Symbiont Dynamics. Oikos 2011, 120, 1121–1128. [Google Scholar] [CrossRef]
- Frank, A.; Saldierna Guzmán, J.; Shay, J. Transmission of Bacterial Endophytes. Microorganisms 2017, 5, 70. [Google Scholar] [CrossRef] [Green Version]
- Van der Heijden, M.G.; de Bruin, S.; Luckerhoff, L.; van Logtestijn, R.S.; Schlaeppi, K. A Widespread Plant-Fungal-Bacterial Symbiosis Promotes Plant Biodiversity, Plant Nutrition and Seedling Recruitment. ISME J. 2016, 10, 389–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, G.; Raaijmakers, J.M. Saving Seed Microbiomes. ISME J. 2018, 12, 1167–1170. [Google Scholar] [CrossRef]
- Wang, T.; Song, Z.; Wang, X.; Xu, L.; Sun, Q.; Li, L. Functional Insights into the Roles of Hormones in the Dendrobium officinale-tulasnella sp. Germinated Seed Symbiotic Association. Int. J. Mol. Sci. 2018, 19, 3484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA Prediction Server: Biological Network Integration for Gene Prioritization and Predicting Gene Function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Ronemus, M.; Vaughn, M.W.; Martienssen, R.A. MicroRNA-Targeted and Small Interfering RNA–Mediated MRNA Degradation Is Regulated by Argonaute, Dicer, and RNA-Dependent RNA Polymerase in Arabidopsis. Plant Cell 2006, 18, 1559–1574. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.; Ambaru, B.; Thakkar, P.; Marcotte, E.M.; Rhee, S.Y. Rational Association of Genes with Traits Using a Genome-Scale Gene Network for Arabidopsis thaliana. Nat. Biotechnol. 2010, 28, 149–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irving, L.J.; Cameron, D.D. Chaper 3 You are What You Eat: Interactions Between Root Parasitic Plants and Their Hosts. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2009; Volume 50, pp. 87–138. ISBN 0065-2296. [Google Scholar]
- Westwood, J.H.; Yoder, J.I.; Timko, M.P.; de Pamphilis, C.W. The Evolution of Parasitism in Plants. Trends Plant Sci. 2010, 15, 227–235. [Google Scholar] [CrossRef]
- Brun, G.; Braem, L.; Thoiron, S.; Gevaert, K.; Goormachtig, S.; Delavault, P. Seed Germination in Parasitic Plants: What Insights Can We Expect from Strigolactone Research? J. Exp. Bot. 2018, 69, 2265–2280. [Google Scholar] [CrossRef]
- Toh, S.; Kamiya, Y.; Kawakami, N.; Nambara, E.; McCourt, P.; Tsuchiya, Y. Thermoinhibition Uncovers a Role for Strigolactones in Arabidopsis Seed Germination. Plant Cell Physiol. 2012, 53, 107–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, R.S.; Purugganan, M.D. Evolution of Crop Species: Genetics of Domestication and Diversification. Nat. Rev. Genet. 2013, 14, 840–852. [Google Scholar] [CrossRef]
- Purugganan, M.D. Evolutionary Insights into the Nature of Plant Domestication. Curr. Biol. 2019, 29, R705–R714. [Google Scholar] [CrossRef] [Green Version]
- Purugganan, M.D.; Fuller, D.Q. The Nature of Selection during Plant Domestication. Nature 2009, 457, 843–848. [Google Scholar] [CrossRef]
- Perrino, E.V.; Wagensommer, R.P.; Medagli, P. Aegilops (Poaceae) in Italy: Taxonomy, Geographical Distribution, Ecology, Vulnerability and Conservation. Syst. Biodivers. 2014, 12, 331–349. [Google Scholar] [CrossRef]
- Allaby, R.G. Domestication Syndrome in Plants. In Encyclopedia of Global Archaeology; Smith, C., Ed.; Springer: New York, NY, USA, 2014; pp. 2182–2184. ISBN 978-1-4419-0426-3. [Google Scholar]
- Kilian, B.; Graner, A. NGS Technologies for Analyzing Germplasm Diversity in Genebanks. Brief. Funct. Genom. 2012, 11, 38–50. [Google Scholar] [CrossRef] [Green Version]
- Pedrini, S.; Dixon, K.W. International Principles and Standards for Native Seeds in Ecological Restoration. Restor. Ecol. 2020, 28. [Google Scholar] [CrossRef]
- Heywood, V.; Casas, A.; Ford-Lloyd, B.; Kell, S.; Maxted, N. Conservation and Sustainable Use of Crop Wild Relatives. Agric. Ecosyst. Environ. 2007, 121, 245–255. [Google Scholar] [CrossRef]
- Wu, D.; Liang, Z.; Yan, T.; Xu, Y.; Xuan, L.; Tang, J.; Zhou, G.; Lohwasser, U.; Hua, S.; Wang, H.; et al. Whole-Genome Resequencing of a Worldwide Collection of Rapeseed Accessions Reveals the Genetic Basis of Ecotype Divergence. Mol. Plant 2019, 12, 30–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson-Wagner, R.; Briskine, R.; Schaefer, R.; Hufford, M.B.; Ross-Ibarra, J.; Myers, C.L.; Tiffin, P.; Springer, N.M. Reshaping of the Maize Transcriptome by Domestication. Proc. Natl. Acad. Sci. USA 2012, 109, 11878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koenig, D.; Jiménez-Gómez, J.M.; Kimura, S.; Fulop, D.; Chitwood, D.H.; Headland, L.R.; Kumar, R.; Covington, M.F.; Devisetty, U.K.; Tat, A.V.; et al. Comparative Transcriptomics Reveals Patterns of Selection in Domesticated and Wild Tomato. Proc. Natl. Acad. Sci. USA 2013, 110, E2655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, X.-Y.; Foley, M.E.; Horvath, D.P.; Anderson, J.V.; Feng, J.; Zhang, L.; Mowry, C.R.; Ye, H.; Suttle, J.C.; Kadowaki, K.; et al. Association Between Seed Dormancy and Pericarp Color Is Controlled by a Pleiotropic Gene That Regulates Abscisic Acid and Flavonoid Synthesis in Weedy Red Rice. Genetics 2011, 189, 1515–1524. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, K.; Takeuchi, Y.; Ebana, K.; Miyao, A.; Hirochika, H.; Hara, N.; Ishiyama, K.; Kobayashi, M.; Ban, Y.; Hattori, T.; et al. Molecular Cloning of Sdr4, a Regulator Involved in Seed Dormancy and Domestication of Rice. Proc. Natl. Acad. Sci. USA 2010, 107, 5792–5797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, E.; Wang, J.; Zhu, X.; Hao, W.; Wang, L.; Li, Q.; Zhang, L.; He, W.; Lu, B.; Lin, H.; et al. Control of Rice Grain-Filling and Yield by a Gene with a Potential Signature of Domestication. Nat. Genet. 2008, 40, 1370–1374. [Google Scholar] [CrossRef] [PubMed]
- Poets, A.M.; Fang, Z.; Clegg, M.T.; Morrell, P.L. Barley Landraces Are Characterized by Geographically Heterogeneous Genomic Origins. Genome Biol. 2015, 16, 173. [Google Scholar] [CrossRef] [Green Version]
- Duan, P.; Ni, S.; Wang, J.; Zhang, B.; Xu, R.; Wang, Y.; Chen, H.; Zhu, X.; Li, Y. Regulation of OsGRF4 by OsmiR396 Controls Grain Size and Yield in Rice. Nat. Plants 2015, 2, 15203. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fan, C.; Xing, Y.; Jiang, Y.; Luo, L.; Sun, L.; Shao, D.; Xu, C.; Li, X.; Xiao, J.; et al. Natural Variation in GS5 Plays an Important Role in Regulating Grain Size and Yield in Rice. Nat. Genet. 2011, 43, 1266–1269. [Google Scholar] [CrossRef]
- Song, X.J.; Kuroha, T.; Ayano, M.; Furuta, T.; Nagai, K.; Komeda, N.; Segami, S.; Miura, K.; Ogawa, D.; Kamura, T.; et al. Rare Allele of a Previously Unidentified Histone H4 Acetyltransferase Enhances Grain Weight, Yield, and Plant Biomass in Rice. Proc. Natl. Acad. Sci. USA 2015, 112, 76–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, B.-F.; Si, L.; Wang, Z.; Jingjie Zhu, Y.Z.; Shangguan, Y.; Lu, D.; Fan, D.; Li, C.; Lin, H.; Qian, Q.; et al. Genetic Control of a Transition from Black to Straw-White Seed Hull in Rice Domestication. Plant Physiol. 2011, 155, 1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukunaga, K.; Kawase, M.; Kato, K. Structural Variation in the Waxy Gene and Differentiation in Foxtail Millet [Setaria italica (L.) P. Beauv.]: Implications for Multiple Origins of the Waxy Phenotype. Mol. Gen. Genom. 2002, 268, 214–222. [Google Scholar] [CrossRef]
- Simons, K.J.; Fellers, J.P.; Trick, H.N.; Zhang, Z.; Tai, Y.-S.; Gill, B.S.; Faris, J.D. Molecular Characterization of the Major Wheat Domestication Gene Q. Genetics 2006, 172, 547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbi, J.; Debieu, M.; Rousselet, A.; Montalent, P.; Le Guilloux, M.; Manicacci, D.; Tenaillon, M.I. Contrasted Patterns of Selection since Maize Domestication on Duplicated Genes Encoding a Starch Pathway Enzyme. Theor. Appl. Genet. 2011, 122, 705–722. [Google Scholar] [CrossRef]
- Salvi, S.; Sponza, G.; Morgante, M.; Tomes, D.; Niu, X.; Fengler, K.A.; Meeley, R.; Ananiev, E.V.; Svitashev, S.; Bruggemann, E.; et al. Conserved Noncoding Genomic Sequences Associated with a Flowering-Time Quantitative Trait Locus in Maize. Proc. Natl. Acad. Sci. USA 2007, 104, 11376. [Google Scholar] [CrossRef] [Green Version]
- Hufford, M.B.; Xu, X.; van Heerwaarden, J.; Pyhäjärvi, T.; Chia, J.-M.; Cartwright, R.A.; Elshire, R.J.; Glaubitz, J.C.; Guill, K.E.; Kaeppler, S.M.; et al. Comparative Population Genomics of Maize Domestication and Improvement. Nat. Genet. 2012, 44, 808–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
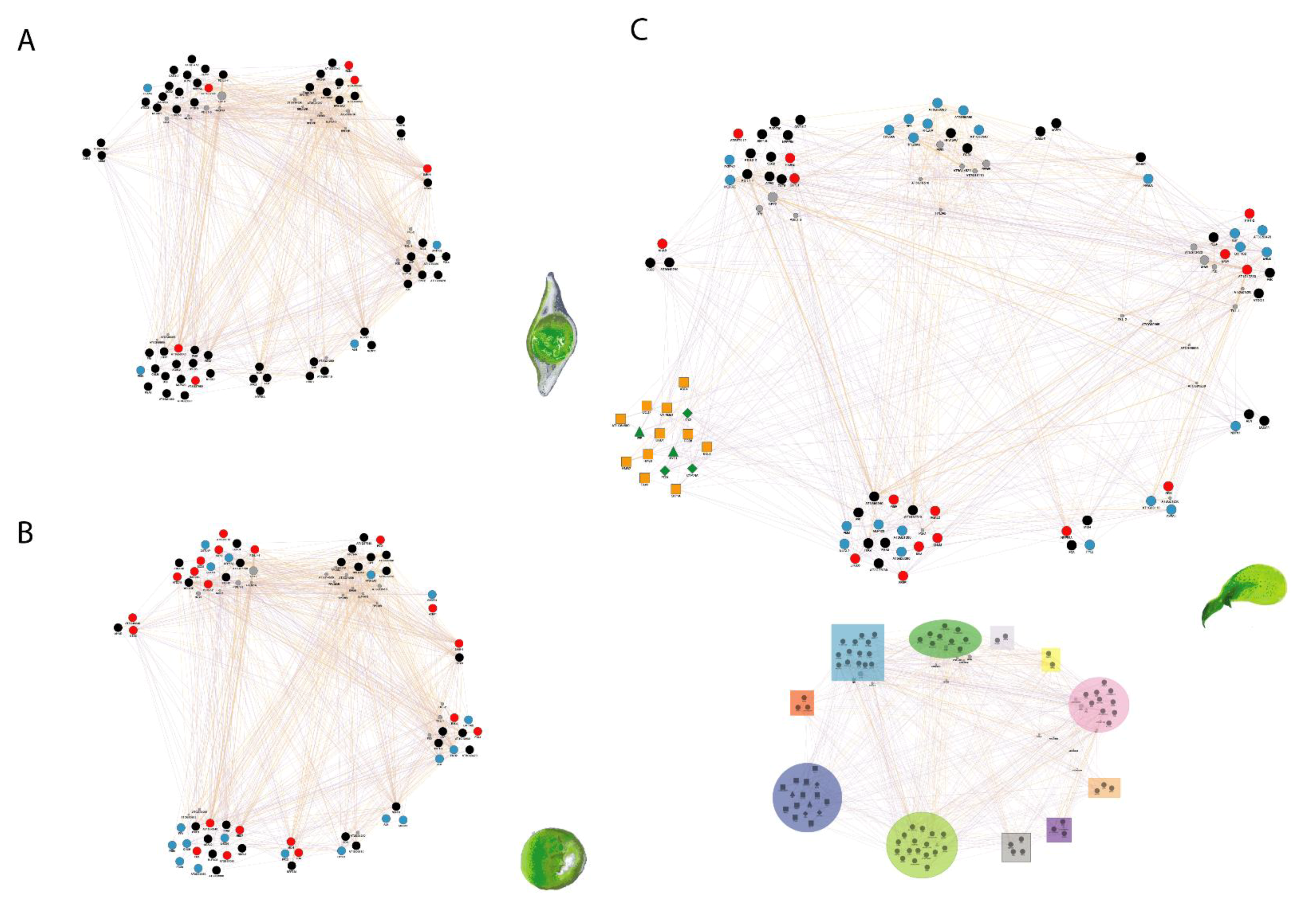
| Name | Family | Clade | Manage-ment | Original Climate | Life History | Storage Behavior | Dormancy |
|---|---|---|---|---|---|---|---|
| Aconitum heterophyllum | Ranunculaceae | D | W | Sub-alpine | Herbaceous, perennial | Possibly orthodox | Morpho-physiological |
| Aethionema arabicum | Brassicaceae | D | Ma | Semi-arid | Herbaceous, annual | Orthodox | Morphotype-dependent, physiological |
| Arabidopsis thaliana | Brassicaceae | D | Ma | Temperate | Herbaceous, annual | Orthodox | Physiological |
| Arachis hypogaea | Fabaceae | D | Do | Alpine | Herbaceous, annual | Orthodox | Physiological |
| Avicennia marina | Acanthaceae | D | W | Subtropical | Shrub, tree, perennial | Recalcitrant | Viviparous |
| Brassica rapa | Brassicaceae | D | Do | Semi-arid | Herbaceous, annual | Orthodox | Physiological |
| Castanospermum australe | Fabaceae | D | Ma | Tropical, subtropical | Tree, perennial | Recalcitrant | Not determined |
| Ceiba aesculifolia | Malvaceae | D | W | Tropical, subtropical | Tree, perennial | Orthodox | Non dormant |
| Chenopodium quinoa | Amaranthaceae | D | Do | Temperate | Herbaceous, annual | Orthodox | PHS |
| Cynara cardunculus var. sylvestris | Asteraceae | D | W | Mediterranean | Herbaceous, perennial | Possibly orthodox | Physiological |
| Fraxinus hupehensis | Oleaceae | D | W | Temperate, no dry season | Tree, perennial | Not determined | Not determined |
| Glycine max | Fabaceae | D | Do | Subtropical | Herbaceous, annual | Orthodox | Non-dormant |
| Lactuca sativa | Asteraceae | D | Do | Mediterranean, temperate | Herbaceous, annual | Orthodox | Physiological dormancy |
| Medicago truncatula | Fabaceae | D | Do | Mediterranean | Herbaceous, annual | Orthodox | Physical |
| Phelipanche aegyptiaca | Orobanchaceae | D | W | Temperate, desertic | Herbaceous | Not determined | Not determined |
| Populus nigra x Populus simonii | Salicaceae | D | Do | Temperate | Tree, perennial | Possibly Orthodox | Possibly non dormant |
| Quercus ilex | Fagaceae | D | Cu | Mediterranean | Tree, perennial | Recalcitrant | Non dormant |
| Quercus robur | Fagaceae | D | Cu | Temperate | Tree, perennial | Recalcitrant | Non dormant |
| Nicotiana tabacum | Solanaceae | D | Cu | Tropical | Herbaceous, annual | Orthodox | Physiological |
| Solanum lycopersicum | Solanaceae | D | Do | Tropical | Herbaceous, annual | Orthodox | Physiological |
| Anoectochilus roxburghii | Orchidaceae | M | W | Temperate, no dry season | Herbaceous, perennial | Orthodox | Not determined |
| Calanthe tsoongiana | Orchidaceae | M | W | Temperate, subtropical | Herbaceous, perennial | Not determined | Not determined |
| Dendrobium officinale | Orchidaceae | M | W | Tropical, subtropical | Herbaceous, perennial | Orthodox | Not determined |
| Leymus chinensis | Poaceae | M | Ma? | Temperate | Herbaceous, perennial | Orthodox | Possibly physiological |
| Oryza sativa | Poaceae | M | Do | Temperate and tropical | Herbaceous, annual | Orthodox | Cultivar-dependent. Physiological to PHS susceptible |
| Paris polyphylla | Melanthiaceae | M | Cu? | Temperate | Herbaceous, perennial | Not determined | Morpho-physiological |
| Triticum aestivum | Poaceae | M | Do | Semi-arid | Herbaceous, annual | Orthodox | Non-dormant, PHS susceptible |
| Zea mays | Poaceae | M | Do | Tropical | Herbaceous, annual | Orthodox | Non-dormant, PHS susceptible |
| Zea mexicana ssp. parviglumis | Poaceae | M | W | Tropical | Herbaceous, annual | Orthodox | Physiological |
| Cunninghamia lanceolata | Cupressaceae | G | Cu? | Temperate | Tree, perennial | Probably orthodox | Physiological |
| Ginkgo biloba | Ginkgoaceae | G | Cu | Mediterranean | Tree, perennial | Recalcitrant | Morpho-physiological |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Maqueo, X.; Figueroa-Corona, L.; Martínez-Villegas, J.A.; Soriano, D.; Gamboa-deBuen, A. The Relevance of a Physiological-Stage Approach Study of the Molecular and Environmental Factors Regulating Seed Germination in Wild Plants. Plants 2021, 10, 1084. https://doi.org/10.3390/plants10061084
Gómez-Maqueo X, Figueroa-Corona L, Martínez-Villegas JA, Soriano D, Gamboa-deBuen A. The Relevance of a Physiological-Stage Approach Study of the Molecular and Environmental Factors Regulating Seed Germination in Wild Plants. Plants. 2021; 10(6):1084. https://doi.org/10.3390/plants10061084
Chicago/Turabian StyleGómez-Maqueo, Ximena, Laura Figueroa-Corona, Jorge Arturo Martínez-Villegas, Diana Soriano, and Alicia Gamboa-deBuen. 2021. "The Relevance of a Physiological-Stage Approach Study of the Molecular and Environmental Factors Regulating Seed Germination in Wild Plants" Plants 10, no. 6: 1084. https://doi.org/10.3390/plants10061084
APA StyleGómez-Maqueo, X., Figueroa-Corona, L., Martínez-Villegas, J. A., Soriano, D., & Gamboa-deBuen, A. (2021). The Relevance of a Physiological-Stage Approach Study of the Molecular and Environmental Factors Regulating Seed Germination in Wild Plants. Plants, 10(6), 1084. https://doi.org/10.3390/plants10061084