
Integrative Taxonomic, Ecological and Genotyping Study of Charophyte Populations from the Egyptian Western-Desert Oases and Sinai Peninsula

 ,
, 
 ,
,  ,
, 
Abstract
1. Introduction
2. Results and Discussion
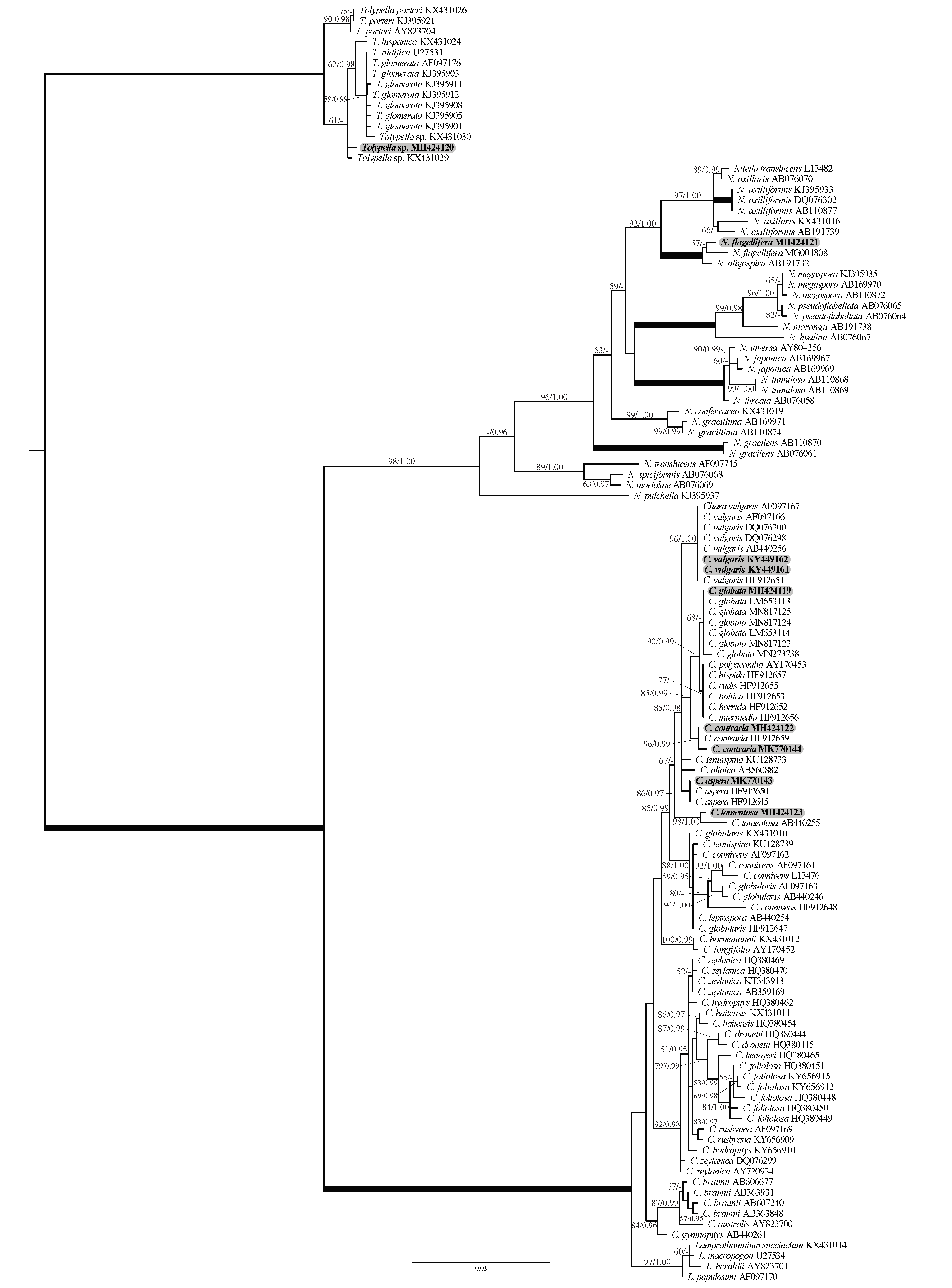
2.1. Phylogenetic Affinities of the Charophyte Specimens Investigated
2.2. Morphotaxonomy, Autecology, and Biogeography of the Charophyte Specimens Studied
2.2.1. Chara aspera Willdenow (Figure 5A–K)
- Description: Plants green, dioecious, up to 40 cm tall, without incrustations (Figure 5A). Axes moderately slender, 350–450 µm in diameter. Cortex triplostichous, isostichous to tylacanthous (Figure 5B,E). Spine-cells solitary, papilliform (Figure 5B,E). Stipulodes diplostephanous (in 2 tiers), 2 sets per branchlet, acuminate, uppers somewhat longer than lowers (Figure 5B). Internodes corticated, 1–3 times longer than the branchlets (Figure 5A,B). Branchlets 6–9 in a whorl, straight and spreading, 1.2–1.8 cm long (Figure 5A); each branchlet consisting of 5–6 corticated segments (Figure 5C,D); end segment 1–2-celled, naked (Figure 5F,G). Bract-cells usually 5, well developed, unilateral, shorter to longer than oogonium (Figure 5H). Bracteoles 2, somewhat longer than the bract-cells and exceeding the mature oogonium (Figure 5I,J). Gametangia on separate plants and the female thalli only observed. Oogonia solitary at the 2–3 lowest branchlet nodes, 690–750 µm long (without coronula) × 450–500 μm wide, with 12–13 convolutions. Coronula 50–85 µm long × 50–100 (–120) μm wide (Figure 5K). Oospores and bulbils not observed.
- Distribution in Egypt: This charophyte species has already been recorded in Egypt [32].
- General distribution and ecology: Cosmopolitan species in Europe [4], Atlantic Islands [45], North America [46], Africa and Middle East [15,32,47], and Asia [48]. So far, it has not yet been recorded in South America, the Pacific Islands, and Australia [47]. In North Africa, Muller et al. [5] pointed out that this species is frequently common in coastal ponds and marshes. During the present study, the Chara aspera population was found in a mineral spring-fed agricultural ditch in the Siwa Oasis. Hydrochemical conditions in this Saharan biotope were as follows: high water temperature (°C): 27.7; neutral pH: 7.32; high electrical conductivity (μS·cm−1): 6350; Na+ and Ca2+ were the major cations: 1140 and 276 mg·L−1, respectively; Cl− was the major anion: 2104 mg·L−1; low N, NO3− and NH4+: 460 and 68 µg·L−1, respectively; high P, TP and SRP: 260 and 55 µg·L−1, respectively (Table 1). In a similar study, Zouaïdia et al. [15] reported C. aspera in nitrate-rich brackish wetlands (NO3−: 400–840 μg·L−1), with moderate orthophosphate levels (30–48 μg·L−1). Additionally, Caisová and Gąbka [49] and Urbaniak and Gąbka [50] highlighted that C. aspera has a wide range of ecological preferences in fresh and brackish calcareous waters, mainly in drainage canals and lakes.
- Remarks: There is general agreement between the characteristics of our C. aspera population and the information provided by Wood and Imahori [47]; however, the following aberrant taxonomic features were noted: (1) the spine cells are solitary and papilliform (vs. well-developed spine cells that may be solitary and in fascicles of 2–3 cells, often with bulbous bases, and up to 2.5 times as long as the axis diameter), (2) stipulodes are distinctly much shorter (vs. stipulodes 0.5–2 times as long as the axis diameter and often as long as the basal branchlet segment), (3) branchlets 6–9 in a whorl and each one consisting of 5–6 corticated segments (vs. 8–9 branchlets with 6–8 segments of which 5–7 are 2-corticate), (4) oogonia convolutions with 12–13 turns (vs. 13–15 turns in the protologue), and (5) coronula much smaller (i.e., 50–85 µm long × 50–100 (–120) μm wide vs. 75–100 µm long × 120–200 μm wide). In agreement with our taxonomic observations, the recent integrative study by Langangen et al. [51] on the charophytes inhabiting the warm Troll Springs in Svalbard (Spitsbergen) recorded a morphologically aberrant C. aspera population in these groundwater-dependent biomes, but the plants were still genetically identical to other specimens of C. aspera from several European countries. Taxonomically, they showed that these plants were ecorticated and sterile, stipulodes were absent, branchlets with 5–10 segments, cortex of the branchlets rudimentary or missing, and eventually branchlets tipped with 2–3 ecorticated cells. Our observations and findings of Langangen et al. [51] confirm the phenotypic variations in the spine-cells, stipulodes, and cortication in C. aspera. Accordingly, Blindow et al. [28] pointed out that the variability in lengths of spines, stipulodes, and bract cells, as well as branchlet cortication, are traits of limited value for species delineation.
2.2.2. Chara contraria A.Braun ex Kützing (Figure 6A–K and Figure 7A–K)
- Description: Plants olive green to green, monoecious, 15–50 cm tall, without incrustations (Figure 6A) or moderately incrusted (Figure 7A). Axes moderately slender, 350–685 µm in diameter. Cortex diplo- to triplostichous, isostichous to tylacanthous (Figure 6F–H and Figure 7E). Spine-cells variable, solitary, papilliform (Figure 6G and Figure 7E) or often shortly cylindrical (Figure 6F), up to 200 µm long. Stipulodes diplostephanous (in 2 tiers), 2 sets per branchlet, short, obtuse or blunt (Figure 6E and Figure 7E). Internodes corticated, 2–3 times as long as the branchlets, 3–4 cm long (Figure 6A and Figure 7A). Branchlets 7–10 in a whorl (Figure 6C and Figure 7B), incurved, (0.8–) 1–1.5 cm long; each branchlet consists of 5–7 segments of which the basal 3–5 segments corticated, diplostichous; end segment 2–3-celled, naked (Figure 6C,D and Figure 7C); terminal cell conical to long acuminate (Figure 6I and Figure 7D). Bract-cells 5, unilateral; anteriors longer than oogonium; posteriors smaller or rudimentary. Bracteoles 2, longer than the anterior bract-cells and 2–3 (–4) times longer than the mature oogonium (Figure 7C). Gametangia conjoined at each 1–4th branchlet nodes, solitary or rarely geminate, without mucus (Figure 6D and Figure 7C). Oogonia solitary (Figure 6J and Figure 7F) or geminate (Figure 7G), 590–825 µm long (without coronula) × 390–530 μm wide, with 13–14 convolutions. Coronula 90–100 (–110) µm long × 80–90 (–100) μm wide, cells oblong, blunt. Oospores dark brown to black (Figure 6K and Figure 7H), (490–) 670–710 µm long × 350–450 μm wide; striae of 10–14 prominent ridges (Figure 6K and Figure 7I), with rounded-shaped granulate ornamentation covering fossae and ridges (Figure 7J,K); fossae ca. 40–42 μm across. Antheridia small, 300–355 μm in diameter (Figure 6J and Figure 7F).
- Distribution in Egypt: Previously recorded in Upper Egypt [36].
- General distribution and ecology: Nearly cosmopolitan in all continents [4,47]. In North Africa, Muller et al. [5] showed that Chara contraria is rather rare in the Mediterranean region and can be found in various biotopes. With respect to its conservation status, Langangen [52] classified it as “Near Threatened” species. During the present study, it was found in a nutrient-rich artificial muddy pool in Wadi El-Arbaeen, Saint Catherine Protectorate, South Sinai and in an agricultural ditch in the El-Dakhla Oasis (the Western Desert of Egypt). Values of environmental variables were as follows: water temperature (°C): 20.6–21.9; circumneutral pH: 6.93–7.46; conductivity (μS·cm−1): 760–2960; Na+ and Ca2+ were the major cations: 62–361 and 62–147 mg·L−1, respectively; Cl− was the major anion: 146–768 mg·L−1; low N, NO3−: 2320–5400 µg·L−1; NH4+: 57–924 µg·L−1; high P, TP and SRP: 150–1680 and 40–276 µg·L−1, respectively (Table 1). Thus, C. contraria can be found in circumneutral to slightly alkaline, fresh to slightly-brackish calcium-rich, meso–eutrophic waters. These observations are in agreement with the findings of Caisová and Gąbka [49] and Muller et al. [5].
- Remarks: Our specimens are consistent with the diagnosis of the protologue illustrated in Wood and Imahori [47,53]. Besides the clear-cut differences in some morphotaxonomic features with the most morphologically close species C. vulgaris (in particular tylacanthous cortication in C. contraria vs. aulacanthous in C. vulgaris), the two species are also well separated genetically (Figure 1).
2.2.3. Chara globata W.Migula (Figure 8A–K, Figure 9A–K and Figure S3A–L)
- Description: Plants green to olive green (Figure 8A,B), monoecious, 20–85 (–95) cm tall, unencrusted to heavily incrusted, forming a massive growth inside the main springhead and the outlet channel of thermal mineral desert spring (Figure S3A,B). Axes predominately stout, 610–1580 (–2000) µm in diameter (Figure 8C and Figure S3C). The internodes usually longer than the branchlets, 1.5–4 (–5) times longer than the branchlets, up to 8 cm long (Figure 8A–C and Figure S3C), the upper parts of thalli look like spherical loose heads (Figure 8C,D). Cortex irregularly diplo- to triplostichous, slightly isostichous to distinctly tylacanthous (Figure 8F–I and Figure S3F). Spine-cells mostly solitary (Figure 8F,H and Figure S3E) or rarely in a bunch of four (only one very long and the other surrounding three distinctly very short) (Figure 8G), subulate, with thickened cell walls at their ends. Stipulodes diplostephanous (in 2 tiers), 2 sets per branchlet, well developed, long aculeiform with acute ends (Figure 8E and Figure S3D). The branchlets usually straight, but still slightly arcuate, 9–10 in a whorl, 1.5–2 (–2.5) cm long (Figure 8A–D and Figure S3C); each branchlet consists of 6–7 segments of which the basal 3–4 segments corticated, diplostichous; the distal segments 3–4 ecorticate (Figure 8D,J and Figure S3G); terminal cell distinctly acuminate and shorter than adjacent bract-cells (Figure 8K). Bract-cells 5–6, verticillate, strongly developed, acuminate, (1.5–) 3–4 mm in length (Figure 8D,J and Figure S3G). Bracteoles 2, usually shorter than the bract-cells. Gametangia conjoined, solitary (Figure 9A), occurring at the 3 lowest nodes between corticated segments (Figure 8C,J and Figure S3G). Oogonia solitary, 825–950 μm long (without coronula) × 535–590 μm wide, with 11–14 convolutions (Figure 9B). Coronula 100–120 µm long × 100–150 μm wide, cells more or less apiculate at apex (Figure 9B). Ripe oospores black (Figure 9C–E and Figure S3H), with 11–15 striae having prominent ridges and ending at the base with a basket–like protrusion (Figure 9F,G), 860–910 µm long (incl. protrusions) × 415–535 μm wide, oospore wall smooth (Figure 9H,J and Figure S3K) to pustular (Figure 9I, Figure S3L) to slightly papillate covering fossae and ridges (Figure 9K); fossae 50–87 μm across (Figure 9H,I). Antheridia solitary, octoscutate, 460–520 μm in diameter (Figure 9A).
- Distribution in Egypt: Only recorded in the Sinai Peninsula—the Asian part of the Egyptian territory—based on the recent study carried out by Romanov et al. [43] on the herbarium specimens collected from the end of the 1960s to the beginning of the 1970s from Sinai and stored at Tel Aviv University Herbarium.
- General distribution and ecology: Rare, but still flagship, temperate species with disjunctive biogeographical distribution, particularly in the arid and semiarid regions. Fresh–brackish, moderately alkaliphilic (pH: 7.1–8.0) species preferring waters rich in sodium sulphates, and calcium/magnesium bicarbonates [43]. So far, it has only been recorded in Asia (China, Iran, Israel, Kazakhstan, Kyrgyzstan, Russia, Turkmenistan and Uzbekistan) [43,47,54,55,56,57], Europe (Romania and the European part of Russia) [57,58], the Sahara–Arabian Desert in Sinai Peninsula [43], and in North Africa (only in Tunisia) [57]. During the present study, C. globata was found in the thermal mineral desert spring ‘Ain Wazedi’ in the Siwa Oasis. This Saharan biotope was characterized by the following hydrochemical characteristics:—high water temperature (°C): 26.5; circumneutral pH: 6.85; high conductivity (μS·cm−1): 6280; low DO (mg·L−1): 3.4; Na+ and Ca2+ were the major cations: 1430 and 94 mg·L−1, respectively; Cl− and SO42− were the major anions: 1795 and 989 mg·L−1, respectively; low N, NO3−, NO2− and NH4+: 200, 120, and 270 µg·L−1, respectively; high P, TP and SRP: 135 and 100 µg·L−1, respectively (Table 1).
- Remarks: The diagnostic taxonomic features of the Siwa C. globata population fitted better the specimens recently described by Romanov et al. [43] than the protologue redescribed by Wood and Imahori [47,53]. However, our specimens still differ from the description in Romanov et al. [43] by the following taxonomic features: (1) stem cortex irregularly diplo- to triplostichous, slightly isostichous to distinctly tylacanthous (vs. consistently a tylacanthous diplostichous stem cortex), (2) spine-cells mostly solitary, long acuminate and rarely in a bunch of four (only one very long and the other surrounding three distinctly very short) (vs. only solitary and variable in length from short conical–papillose to conical to long subulate), (3) gametangia usually present at the 3 lowest nodes of the corticated segments (vs. gametangia occurring at the 2–4 lowest nodes between corticated segments and rarely between ecorticate segments), and (4) ripe oospores are obviously dominant (vs. oospores low or absent in the majority of the specimens). We think that all these phenotypic variations are environmentally-induced and with a low taxonomic value. Taxonomically, Romanov et al. [43] also proposed that C. globata should be transferred and assigned to the subsection Chara in the section Chara, instead of the section Grovesia having a triplostichous stem cortex, in terms of the taxonomic observations obtained (i.e., consistently and generally tylacanthous diplostichous stem cortex, solitary spine–cells, and stipulodes in two tiers), corresponding well to the section Chara [47]. On the contrary, the Siwa C. globata specimens investigated in the present study are mainly characterized by the presence of isostichous to tylacanthous diplo- to triplostichous stem cortex (Figure 8F–I). Ling et al. [55] also documented irregular triplostichous tylacanthous cortex in Chinese specimens of C. globata. Additionally, the subsection Chara placement proposed by Romanov et al. [43] was not supported by crossing experiments conducted by Proctor [59,60], who pointed to the affinity of C. globata towards the subsection Hartmania. However, the combined morphotaxonomic and phylogenetic data obtained in this study (Figure 1), as well as work of Romanov et al. [43], showed that C. globata has more or less a closer affinity to species of the subsection Hartmania but that it is still different genetically and taxonomically (in particular in the presence of the verticillate bract-cells and arcuate branchlets mainly in the apical parts of thalli). In our opinion, the accurate taxonomic placement of C. globata is still problematic and more integrative studies are needed.
2.2.4. Chara tomentosa Linnaeus (Figure 10A–M)
- Description: Plants dioecious, small, robust, moderately to heavily encrusted, parts of the plants red in color, up to 8 cm tall (Figure 10A). Axes stout, 410–480 µm in diameter (Figure 10B). Stem cortex diplostichous, tylacanthous (Figure 10E). Internodes 1.5–2.0 (–2.5) times longer than the branchlets, upper internodes much shorter, up to 3 cm long (Figure 10A,B). Spine-cells variable, mostly solitary, sometimes ovoid with thickened walls (Figure 10E,F). Stipulodes diplostephanous (in 2 tiers), 2 sets per branchlet, short, ovoid (Figure 10D). Branchlets 6 in a whorl, slightly incurved, 1–1.2 cm long; each branchlet consisting of 6–7 segments of which the basal 4–5 segments 2-corticated (Figure 10C); end segment 2–3-celled, ecorticated; penultimate cells swollen, constricted at the nodes, broader than other segments (Figure 10G); terminal cell acute to mucronate (Figure 10H,I). Bract-cells 5, verticillate, acuminate; anteriors longer than oogonium; posteriors much smaller or similar to anteriors (Figure 10J). Bracteoles 2, somewhat longer than anterior bract-cells, acuminate (Figure 10K). Gametangia on separate plants, female thalli only observed. Oogonia solitary, heavily incrusted, at each 1st–3rd lowest branchlet nodes (Figure 10C), 450–490 µm long (without coronula) × 345–360 μm wide, mostly with 14 convolutions. Coronula 90–115 µm long × 95–125 μm wide. Oospores brown to dark brown (Figure 10L), 450–485 µm long × 295–317 μm wide; striae of 13–14 prominent and slightly flanged ridges apparently prolonged at the base into claws, with smooth to irregularly patterned small projections covering fossae and ridges; fossae ca. 30–33 μm across (Figure 10M).
- Distribution in Egypt: This charophyte species has been previously recorded in the Siwa Oasis [36].
- General distribution and ecology: Europe [4], North Africa [5,47], and Asia [62]. Old records in North and South Americas resulted from erroneous identifications [47]. Caisová and Gąbka [49] highlighted that it is occasionally present in marshes and large fish ponds. Urbaniak and Gabka [50] reported its occurrence in meso- to eutrophic calcium-rich lakes. In North Africa, it has been categorized as a fresh–brackish, rare species with a limited distribution in the Maghreb countries [5]. As regards its conservation status, Stewart and Church [63] considered it to be a “vulnerable” species in Britain and Ireland. During the present study, C. tomentosa was found in a brackish, calcium-rich, shallow marsh in the El-Dakhla Oasis. The main physical and chemical features of this Saharan biotope were: high temperature (°C): 27.2; slightly alkaline pH: 7.53; high conductivity (μS·cm−1): 7700; DO (mg·L−1): 7.1; Na+ and Ca2+ as major cations: 1072 and 654 mg·L−1, respectively; Cl− and SO42− as major anions: 2543 and 582 mg·L−1, respectively; NO3− and NH4+: 1765 and 114 µg·L−1, respectively; TP and SRP: 715 and 295 µg·L−1, respectively (Table 1).
- Remarks: The diagnostic taxonomic features of the C. tomentosa population in our study better fitted the description illustrated by Corillion and Guerlesquin [36] than the specimens redescribed by Wood and Imahori [47,53] in the following: (1) branchlet segments 6–7 of which the basal 4–5 segments corticated and end segment 2–3-celled, ecorticated vs. 3–5(–6) segments of which (1–)2–3(–5) corticated and 1–3 naked, and (2) oogonia present at each 1–3th lowest branchlet nodes vs. 2nd–3rd lowest branchlet nodes [47,53]. The smaller dimensions of oogonia and oospores might be related to the desert environmental conditions.
2.2.5. Chara vulgaris Linnaeus (Figure 11A–O)
- Description: Plants monoecious, green in color, without incrustations, up to 45 cm tall (Figure 11A,B). Axes slender or moderately stout, 400–650 µm in diameter. Stem cortex diplostichous, generally aulacanthous (Figure 11D,O) to more or less isostichous (Figure 11C,M,N). Internodes 1–2 (–2.5) times as long as the branchlets, upper internodes much shorter, up to 3 cm long (Figure 11A,B). Spine-cells variable, solitary, papilliform (Figure 11M,N) or short, obtuse (Figure 11D). Stipulodes diplostephanous (in 2 tiers), 2 sets per branchlet, short, oblong-ovate (Figure 11C). Branchlets (6–)7–9(–10) in a whorl, incurved in the upper parts of the plants (Figure 11B) and more spreading in the lower parts of the plants (Figure 11A), 1–2 cm long; segments 4–5 of which the basal 3–4 segments 2-corticated, poorly developed; end segment 1–2-celled (Figure 11E,K), ecorticated; end cell conical (Figure 11L). Bract-cells unilateral, anteriors developed, 1–2 pairs longer than oogonium (about 5–13 times the length of oogonia), exceeding adjacent segment; posteriors usually rudimentary or absent (Figure 11E,K). Bracteoles similar to anterior elongated bract-cells. Gametangia conjoined at the 1–3th lowest branchlet nodes (adjacent to corticated segments) (Figure 11E,K). Oogonia solitary, 472–590 µm long (without coronula) × 300–415 μm wide, with 13–14 convolutions. Coronula 70–95 µm long × 55–74 μm wide (Figure 11F). Oospores dark brown to black, 495–565 µm long × 295–320 μm wide (Figure 11G,H); striae of 11–12 prominent and slightly flanged ridges (Figure 11I), with granulate ornamentation (with pores) on both fossae and ridges (Figure 11J); fossa ca. 45 μm across. Antheridia solitary, octoscutate, 260–390 in diameter (Figure 11F).
- General distribution and ecology: Cosmopolitan species in all freshwater biotopes, and occasionally in brackish waters [4,49]. It typically occurs in meso–eutrophic habitats [15]. In North Africa, it is very common in the Maghreb countries [5]. During the present study, it was found in the outlet channel of the thermal mineral desert spring ‘Ain Al-Maamal’ in the Siwa Oasis, and an agricultural ditch in the El-Farafra Oasis. Ranges of the physical and chemical variables determined: water temperature (°C): 23.8–31.6; slightly-acidic to almost neutral pH: 6.16–6.71; high conductivity (μS·cm−1): 960–4470; Na+ and Ca2+ as major cations: 25–545 and 30–135 mg·L−1, respectively; Cl− was the major anion: 131–1235 mg·L−1; SO42−: 36–300 mg·L−1; HCO3−: 221–314 mg·L−1; NO3−: 200–400 µg·L−1; NH4+: 150–271 µg·L−1; TP and SRP: 20–100 and 17–48 µg·L−1, respectively (Table 1). In agreement with our ecological assessment, Zouaïdia et al. [15] pointed out that C. vulgaris is a species tolerating hyper-eutrophic water conditions (up to 780 μg·L−1 for PO43−, 898 μg·L−1 for NO3−, and 140 μg·L−1 for NO2−).
- Remarks: Strong resemblance of our population with the one described by Wood and Imahori [47]. Although our specimens are characterized by the presence of elongated bract-cells, approximately 5–13 times the length of oogonia, they still genetically and taxonomically belong to the polymorphic species Chara vulgaris; not to the variety longibracteata. The same observation has already been reported by Saber et al. [44] during their integrative study on an aberrant C. vulgaris population from the Springs of Moses on the Sinai Peninsula, where they noticed that the bract-cells and bracteoles were clearly longer (ca. 4–12 times) than the oogonia. Similarly, Muller et al. [5] considered this phenotypic variation as a North-African “morphotype/phenoecodeme” due to high insolation.
2.2.6. Nitella flagellifera J.Groves & G.O.Allen (Figure 12A–M)
- Description: Plants monoecious, diffuse, delicate, pale green in color, up to 18 cm tall. Axes moderately slender, 400–435 µm in diameter. Internodes 1–2 times as long as the branchlets (Figure 12A,B). Branchlets fertile and sterile similar, 6–7 in a whorl, 3 cm long, 2–3 furcate, primary ray about half of the branchlet length, secondaries 4–5 of which one is the central ray and relatively more stout than the laterals, tertiaries 3–4 of which one may be central, quaternaries 2–4 (Figure 12C,D). Dactyls 2–4, 2-celled, elongated (Figure 12E,F); penultimate cell narrowed abruptly at distal end; end cell persistent, acute and conical, 100–120 µm long × 43–50 μm wide (Figure 12G,H). Heads not formed. Gametangia conjoined at the 1st–3rd lowest branchlet nodes, without mucous (Figure 12C,D). Oogonia solitary, (190–)200–225 µm long (incl. coronula) × 102–130 μm wide, with 7–8 convolutions (Figure 12I,J); coronula 50–68 µm long × 65–70 μm wide, upper cells more or less longer than lowers (Figure 12K,L). Oospores not observed. Antheridia solitary, 195–220 μm in diameter (Figure 12M).
- Distribution in Egypt: This is the first record of this charophyte both in North Africa and in Egypt.
- General distribution and ecology: According to current knowledge, this charophyte appears to have a limited biogeographical distribution. It has been recorded in Brazil, in South America [25,65,66,67], and Japan, India, and Bangladesh in Asia [47,68]. During the present study, N. flagellifera was found in a nutrient-rich agricultural ditch in the El-Dakhla Oasis, the Western Desert of Egypt. Main physical and chemical characteristics of this biotope were: high water temperature (°C): 31.5; alkaline pH: 8.13; high conductivity (μS·cm−1): 6670; low DO (mg·L−1): 3.8; Na+, K+ and Ca2+ as major cations: 1368, 136 and 110 mg·L−1, respectively; Cl− and SO42− as major anions: 2378 and 356 mg·L−1, respectively; NO3− and NH4+: 1570 and 86 µg·L−1, respectively; TP and SRP: 2750 and 421 µg·L−1, respectively (Table 1). Based on the TP and SRP concentrations, N. flagellifera can be considered an eutraphentic species that can tolerate high levels of pollution.
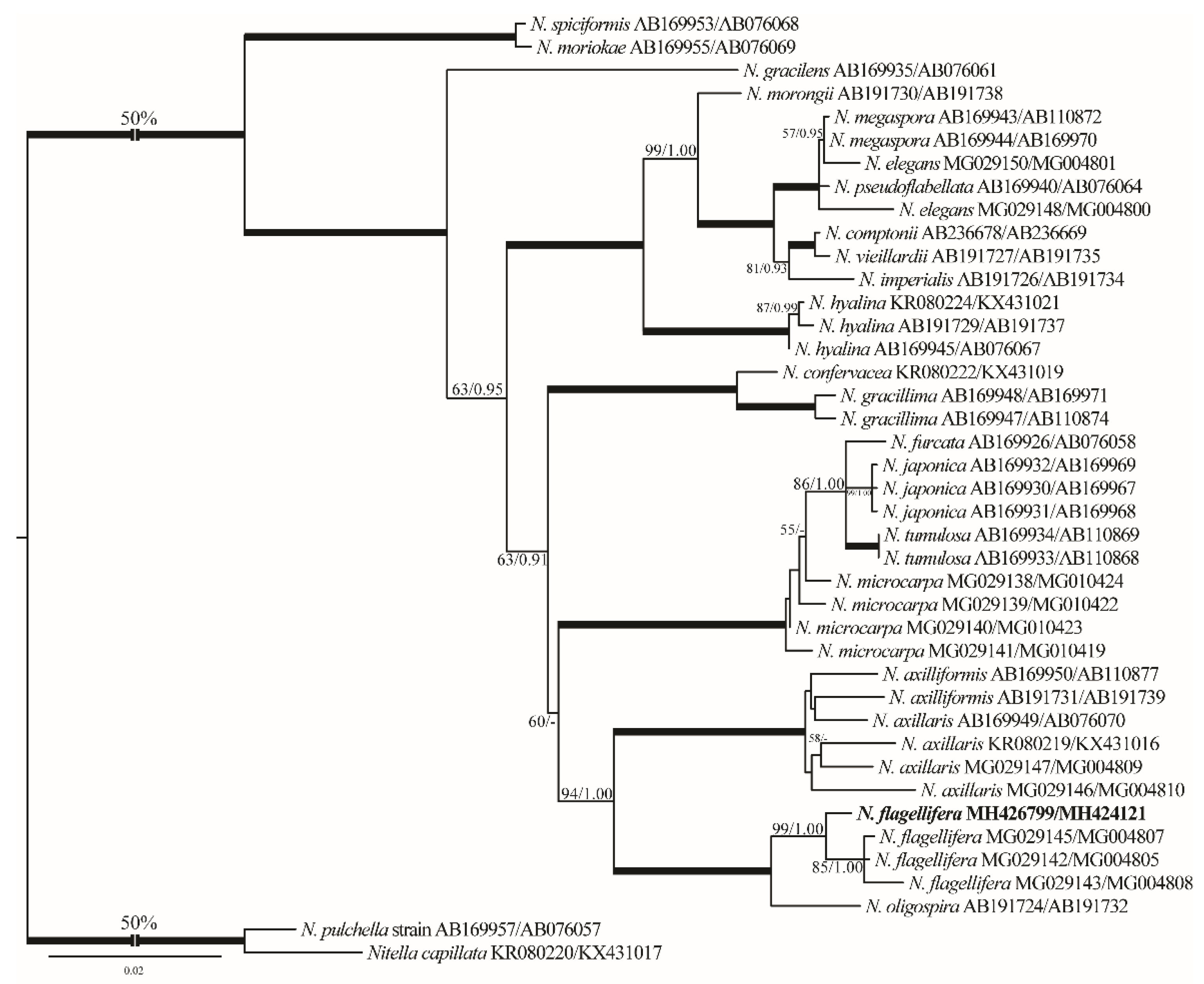
- Remarks: This species is considered a new record for Egypt and also for North Africa based on the published literature ([5,32,36,47] and references therein). Our N. flagellifera rbcL and ITS1 gene sequences are also the first ones for North Africa. From the taxonomic and phylogenetic points of view, our N. flagellifera specimens coincide with the specimens redescribed by Wood and Imahori [47] and also with the findings of Borges and Necchi [25]. Noteworthy, gametangia in our study were noticed at the first node of the branchlet (Figure 12C), and this taxonomic observation has also been documented for the Brazilian N. flagellifera population investigated by Borges and Necchi [25], and other previous studies (e.g., [66,67]). Contrarily, Wood and Imahori [47] noted the lack of gametangia at this position. Blindow et al. [69] pointed out the presence of high phenotypic plasticity and some taxonomic discrepancies in the key characters of the Subfamily Nitelleae, which hamper species identification. N. flagellifera also resembles morphologically and phylogenetically the closest species N. oligospira [25,47]. However, N. flagellifera is still different taxonomically by having a secondary central ray (Figure 12C,D), a unique taxonomic feature that can be easily used to distinguish it from N. oligospira. These two species have more or less similar distribution patterns, are phylogenetically closely related, and also occupy a distinctive position in the genus tree (Figure 3).
2.2.7. Tolypella sp. (A.Braun) A.Braun (Figure 13A–J)
- Description: Plants monoecious, pale green to green, unencrusted, fragile, up to 22 cm tall, with few coarse heads. Axes moderately slender, 500–850 µm in diameter. Internodes 1–2 times as long as the branchlets, becoming shorter towards the apex, up to 5 cm long. Sterile and fertile branchlets different (Figure 13A–D). The first node of the main axis produces 6–7 sterile branchlets and 2–4 secondary axes. The sterile branchlets are undivided and in a series of 3–5 elongated cells (Figure 13B,C). The fertile whorls produced by the secondary axes, short and grouped into fertile heads. Heads few to numerous, 3–14 per shoot (Figure 13A–D). The fertile branchlets apparently consist in a central row of cells (the “rachis”) that is a succession of nodes and internodes. These nodes carry the gametangia as well as 3 rays of 2–3 cells. All terminal cells are elongated, obtuse (Figure 13E). Gametangia conjoined at the fertile branchlet nodes, usually 1 central adaxial antheridium with 1–2(–3) lateral oogonia (Figure 13F). Oogonia 275–335 μm (incl. coronula) long × 250–280 μm wide, with 8–9 convolutions; coronula 30–35 μm high × 40–55 μm wide. Oospores brown to golden brown to slightly dark brown, (275–)320–354 μm long × 215–241 μm wide (Figure 13G); striae of 7–8 prominent, flanged ridges (Figure 13H,I); fossae and ridges with smooth ornamentation (Figure 13J); fossae 37–43 μm across. Antheridia solitary, small, sessile, 105–140 μm in diameter (Figure 13F).
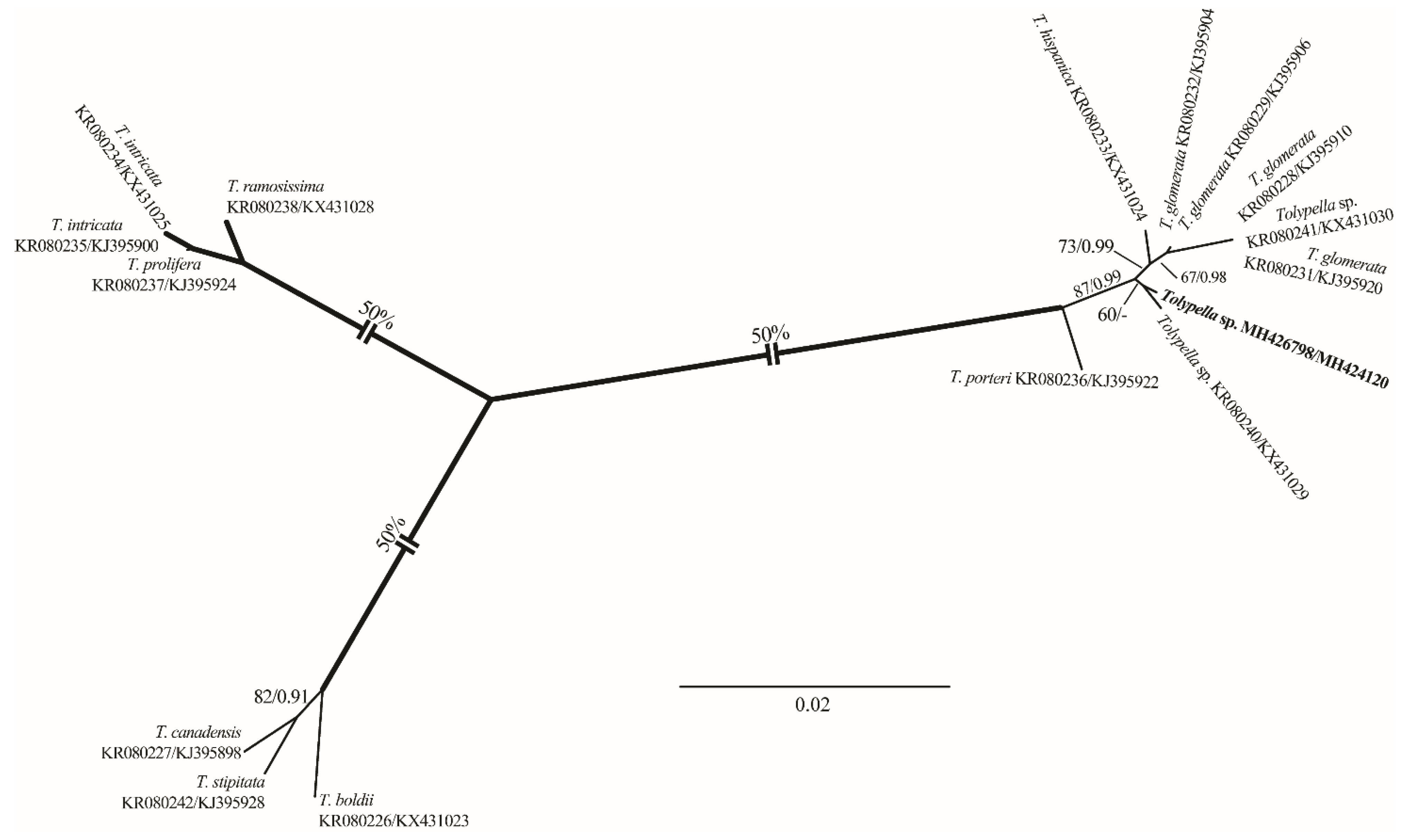
- Distribution in Egypt: This is the first record worldwide of this genetically distinctive charophyte. We therefore designated it with the working name ‘Tolypella sp. PBA–1704 from a desert, freshwater wetland’ mainly based on its concatenated rbcL+ITS1 phylogenetic placement.
- General distribution and ecology: Our Tolypella sp. specimens only showed a weak affinity to Tolypella sp. from Australia based on the concatenated data set of 16 rbcL and ITS1 sequences. During the present study, Tolypella sp. PBA–1704 was found in a shallow meso–eutrophic wetland in the El-Dakhla Oasis, the Western Desert of Egypt. This biotope had the following hydrochemical characteristics: relatively-low temperature (°C): 19.2; slightly-alkaline pH: 7.64; medium-low conductivity (μS·cm−1): 360; low DO (mg·L−1): 3.3; Ca2+ as the major cation: 38 mg·L−1; SO42− and HCO3− as major anions: 88 and 82 mg·L−1, respectively; NO3− and NH4+: 421 and 19 µg·L−1, respectively; TP and SRP: 217 and 62 µg·L−1, respectively (Table 1).
- Remarks: In spite of the high morphotaxonomic similarities between our Tolypella specimens and the cosmopolitan species T. glomerata [47], it is apparently still distinct phylogenetically from that taxon (Figure 1 and Figure 2), and we therefore designated it with the working name ‘Tolypella sp. PBA–1704 from a desert, freshwater wetland’ mainly based on its concatenated rbcL+ITS1 phylogenetic placement. Further in-depth taxonomic and molecular studies on this interesting Tolypella taxon are necessary to propose it as a (morphologically) cryptic species new to science or to recognize it as belonging to a wide genetic variability of T. glomerata.
3. Materials and Methods
3.1. Charophyte Sampling, Processing, and Morphological Identification
3.2. Hydrochemical Characterization of the Sampling Sites
3.3. DNA Extraction, Amplification, and Sequencing
3.4. Phylogenetics Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lewis, L.A.; McCourt, R.M. Green algae and the origin of land plants. Am. J. Bot. 2004, 91, 1535–1556. [Google Scholar] [CrossRef] [PubMed]
- Timme, R.E.; Bachvaroff, T.R.; Delwiche, C.F. Broad phylogenomic sampling and the sister lineage of land plants. PLoS ONE 2012, 7, e29696. [Google Scholar] [CrossRef] [PubMed]
- Palacio-López, K.P.; Tinaz, B.; Holzinger, A.; Domozych, D.S. Arabinogalactan proteins and the extracellular matrix of charophytes: A sticky business. Front. Plant Sci. 2019, 10, 447. [Google Scholar] [CrossRef] [PubMed]
- Krause, W. Charales (Charophyceae). In Süßwasserflora von Mitteleuropa; Ettl, H., Gärtner, G., Heynig, H., Mollenhauer, D., Eds.; Fischer Verlag: Stuttgart, Germany, 1997; Volume 18, ISBN 978-343-725-056-9. [Google Scholar]
- Muller, S.D.; Rhazi, L.; Soulie-Märsche, I.; Benslama, M.; Bottollier-Curtet, M.; Daoud-Bouattour, A.; Belair, G.D.; Ghrabi-Gammar, Z.; Grillas, P.; Paradis, L.; et al. Diversity and distribution of Characeae in the Maghreb (Algeria, Morocco, Tunisia). Cryptogam. Algol. 2017, 38, 201–251. [Google Scholar] [CrossRef]
- Schubert, H.; Blindow, I.; Schories, D.; Mages, M.; von Tümpling, W.; Woelfl, S. Biogeography of Chilean Charophytes—Determined by climate or by water chemistry? Bot. Lett. 2018, 165, 129–145. [Google Scholar] [CrossRef]
- Zalat, A.A.; Al-Wosabi, M.A.; El-Sheekh, M.M.; Al-Subbary, A.K.A. New record of charophytes (Characeae, Charophyta) from Socotra Island, Indian Ocean, Yemen. Thalassas 2020, 36, 437–445. [Google Scholar] [CrossRef]
- Schubert, H.; Blindow, I. Charophytes of the Baltic Sea; The Baltic Marine Biologists, Ed.; Gantner Verlag: Ruggell, Germany, 2003; ISBN 978-390-616-606-3. [Google Scholar]
- Torn, K.; Kovtun-Kante, A.; Herkül, K.; Martin, G.; Mäemets, H. Distribution and predictive occurrence model of charophytes in Estonian waters. Aquat. Bot. 2015, 120, 142–149. [Google Scholar] [CrossRef]
- Grillas, P.; Gauthier, P.; Yavercovski, N.; Perennou, C. Les Mares Temporaires Méditerranéennes. Volume 1: Enjeux de Conservation, Fonctionnement et Gestion; Station Biologique de la Tour du Valat: Arles, France, 2004; ISBN 2-9103-6849-1. [Google Scholar]
- Mjelde, M.; Swe, T.; Langangen, A.; Ballot, A. A contribution to the knowledge of charophytes in Myanmar; morphological and genetic identification and ecology notes. Bot. Lett. 2020, 168, 102–109. [Google Scholar] [CrossRef]
- Blindow, I. Decline of Charophytes during eutrophication: Comparison with angiosperms. Freshw. Biol. 1992, 28, 9–14. [Google Scholar] [CrossRef]
- Azzella, M.M. Italian volcanic lakes: A diversity hotspot and refuge for European charophytes. J. Limnol. 2014, 73. [Google Scholar] [CrossRef]
- Lambert-Servien, E.; Clemenceau, G.; Gabory, O.; Douillard, E.; Haury, J. Stoneworts (Characeae) and associated macrophyte species as indicators of water quality and human activities in the Pays-de-la-Loire region, France. Hydrobiologia 2006, 570, 107–115. [Google Scholar] [CrossRef]
- Zouaïdia, H.; De Bélair, G.; Benslama, M.; Soulié-Märsch, I.; Muller, S.D. Intérêt des Characeae comme bio-indicateurs de pollution des eaux: Le cas des zones humides de Numidie (Nord-Est Algérien). Rev. Ecol. Terre Vie 2015, 70, 121–133. [Google Scholar]
- Becker, R.; Doege, A.; Schubert, H.; van de Weyer, K. Bioindikation mit Characeen. In Armleuchteralgen: Die Characeen Deutschlands; Springer: Berlin/Heidelberg, Germany, 2016; pp. 97–137. ISBN 978-3-662-47797-7. [Google Scholar]
- Soulié-Märsche, I. Diversity of quaternary aquatic environments in NE Africa as shown by fossil Charophytes. In Geoscientific Research in Northeast Africa; Thorweihe, U., Schandelmeier, H., Eds.; CRC Press: London, UK, 1993; pp. 575–579. ISBN 978-0-203-75339-2. [Google Scholar]
- Zalat, A.A. Charophyte gyrogonites from holocene lacustrine sediments of the Fayoum Depression, Egypt. Neues Jahrb. Geol. Paläontol. Mon. 1996, 502–516. [Google Scholar] [CrossRef]
- Soulié-Märsche, I.; Benammi, M.; Gemayel, P. Biogeography of living and fossil Nitellopsis (Charophyta) in relationship to new finds from Morocco. J. Biogeog. 2002, 29, 1703–1711. [Google Scholar] [CrossRef]
- Soulié-Märsche, I.; Benkaddour, A.; El Khiati, N.; Gemayel, P.; Ramdani, M. Charophytes, indicateurs de paléobathymétrie du lac Tigalmamine (Moyen Atlas, Maroc). Geobios 2008, 41, 435–444. [Google Scholar] [CrossRef]
- Sakayama, H.; Kasai, F.; Nozaki, H.; Watanabe, M.M.; Kawachi, M.; Shigyo, M.; Nishihiro, J.; Washitani, I.; Krienitz, L.; Ito, M. Taxonomic reexamination of Chara globularis (Charales, Charophyceae) from Japan based on oospore morphology and rbcL gene sequences, and the description of C. leptospora sp. nov. J. Phycol. 2009, 45, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.C.; Rodrigues, A.; Moe, T.F.; Ballot, A. DNA barcoding the genus Chara: Molecular evidence recovers fewer taxa than the classical morphological approach. J. Phycol. 2015, 51, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Pérez, W.; Casanova, M.T.; Hall, J.D.; McCourt, R.M.; Karol, K.G. Phylogenetic congruence of ribosomal operon and plastid gene sequences for the Characeae with an emphasis on Tolypella (Characeae, Charophyceae). Phycologia 2017, 56, 230–237. [Google Scholar] [CrossRef]
- Borges, F.R.; Necchi, O. Taxonomy and phylogeny of Chara (Charophyceae, Characeae) from Brazil with emphasis on the midwest and southeast regions. Phytotaxa 2017, 302, 101–121. [Google Scholar] [CrossRef]
- Borges, F.R.; Necchi, O. Taxonomy and phylogeny of Nitella (Charophyceae, Characeae) from Brazil with emphasis on the midwest and southeast regions. Phytotaxa 2018, 356, 181–198. [Google Scholar] [CrossRef]
- Urbaniak, J.; Combik, M. Genetic and morphological data fail to differentiate Chara intermedia from C. baltica, or C. polyacantha and C. rudis from C. hispida. Eur. J. Phycol. 2013, 48, 253–259. [Google Scholar] [CrossRef][Green Version]
- Schneider, S.C.; Nowak, P.; Ammon, U.V.; Ballot, A. Species differentiation in the genus Chara (Charophyceae): Considerable phenotypic plasticity occurs within homogenous genetic groups. Eur. J. Phycol. 2016, 51, 282–293. [Google Scholar] [CrossRef]
- Blindow, I.; Marquardt, R.; Schories, D.; Schubert, H. Charophyceae of Chile—Taxonomy and distribution. Part 1. Subfamily Chareae. Nova Hedwig. 2018, 107, 1–47. [Google Scholar] [CrossRef]
- Trbojević, I.; Marković, A.; Blaženčić, J.; Simić, G.S.; Nowak, P.; Ballot, A.; Schneider, S. Genetic and morphological variation in Chara contraria and a taxon morphologically resembling Chara connivens. Bot. Lett. 2020, 167, 187–200. [Google Scholar] [CrossRef]
- Braun, A. Die Characeen Afrika’s zusammengestellt von Alexander Braun. In Monatsberichte der Königlichen Akademie der Wissenschaften; Akademische Buchdruckerei: Berlin, Germany, 1868. [Google Scholar]
- Corillion, R. Contribution à l’étude des Characées de Tunisie et bilan actuel de la flore charologique tunisienne. Bull. Soc. Phycol. Fr. 1977, 22, 47–59. [Google Scholar]
- Corillion, R. Les Characées du Nord de l’Afrique: Éléments floristiques et distribution. Bull. Soc. Études Sci. Anjou Nouv. Sér. 1978, 10, 27–34. [Google Scholar]
- Elkhiati, N. Biotypologie et Biogéographie des Charophycées du Maroc. Ph.D. Thesis, Faculty of Science Semlalia, Marrakech, Morocco, 1995; 155p. [Google Scholar]
- Cantonati, M.; Poikane, S.; Pringle, C.M.; Stevens, L.E.; Turak, E.; Heino, J.; Richardson, J.S.; Bolpagni, R.; Borrini, A.; Cid, N.; et al. Characteristics, main impacts, and stewardship of natural and artificial freshwater environments: Consequences for biodiversity conservation. Water 2020, 12, 260. [Google Scholar] [CrossRef]
- Saber, A.A.; El-Sheekh, M.; Nikulin, A.Y.; Cantonati, M.; Saber, H. Taxonomic and ecological observations on some algal and cyanobacterial morphospecies new for or rarely recorded in either Egypt or Africa. Egypt. J. Bot. 2021, 61, 283–301. [Google Scholar] [CrossRef]
- Corillion, R.; Guerlesquin, M. Notes phytogéographiques sur les Charophycées d’Egypte. Rev. Algol. 1971, 10, 177–191. [Google Scholar]
- Shaaban, A.S. The algal flora of Egyptian oases. II—On the algae of Siwa Oasis. Proc. Egypt. Bot. Soc. 1985, 4, 1–10. [Google Scholar]
- El-Awamri, A.A.; Shaaban, A.S.; Hamed, A.F. Algae in Saint Catherine region (South Sinai, Egypt). Egypt. J. Bot. 1996, 36, 145–168. [Google Scholar]
- Hamed, A.F. Geographical distribution of aquatic and soil algae of Wadi Gharandel, southwest Sinai, Egypt. Az. J. Microbiol. 2001, 53, 54–77. [Google Scholar]
- Shanab, S.M.M. Algal flora of Ain Helwan I. algae of the worm spring. Egypt. J. Phycol. 2006, 7, 209–231. [Google Scholar] [CrossRef]
- Shaaban, A.S.; Mansour, H.A.; Saber, A.A. Unveiling algal biodiversity of El-Farafra Oasis (Western Desert, Egypt) and potential relevance of its use in water bio-assessment: Special interest on springs and drilled wells. Egypt. J. Phycol. 2015, 16, 47–75. [Google Scholar] [CrossRef]
- Abd El Fatah, H.M.; Ibrahim, M.; Ibrahim, S.D. Morphological and molecular diversity among selected Chara L. species (Characeae) from different habitats in Egypt. Egypt. J. Exp. Biol. 2015, 11, 129–139. [Google Scholar]
- Romanov, R.E.; Gontcharov, A.A.; Barinova, S.S. Chara globata Mig. (Streptophyta: Charales): Rare species revised. Fottea 2015, 15, 39–50. [Google Scholar] [CrossRef]
- Saber, A.A.; Ballot, A.; Schneider, S.C.; Cantonati, M. Morphological and molecular features of a Chara vulgaris population from desert springs on the Sinai Peninsula (Springs of Moses, Egypt). Bot. Lett. 2018, 165, 77–89. [Google Scholar] [CrossRef]
- Hrafnsdottir, T.; Ingimarsson, F.; Ingvason, H.R.; Stefánsson, S.M.; Þorvaldsdóttir, E.G.; Malmquist, H.J.; Langangen, A. New finds of charophytes in Iceland with an update on the distribution of the charophyte flora. Nord. J. Bot. 2019, 37, e02111. [Google Scholar] [CrossRef]
- Langangen, A. Some charophytes (Chlorophyta, Charales) from Alaska, USA. Acta Entomol. Musei Natl. Pragae Ser. B Hist. Nat. 2001, 57, 51–56. [Google Scholar]
- Wood, R.D.; Imahori, K. A Revision of the Characeae. First Part: Monograph of the Characeae; Verlag Von J. Cramer: Weinheim, Germany, 1965; pp. 1–904. [Google Scholar]
- Barinova, S.; Niyatbekov, T. Alpha-biodiversity of nondiatom algae in the Pamir aquatic habitats, Tajikistan. Biodivers. Int. J. 2018, 2, 210–237. [Google Scholar] [CrossRef]
- Caisová, L.; Gąbka, M. Charophytes (Characeae, Charophyta) in the Czech Republic: Taxonomy, autecology and distribution. Fottea 2009, 9, 1–43. [Google Scholar] [CrossRef]
- Urbaniak, J.; Gąbka, M. Polish Charophytes. An Illustrated Guide to Identification; Uniwersytet Przyrodniczy we Wroclawiu: Wrocław, Poland, 2014; ISBN 978-83-7717-166-0. [Google Scholar]
- Langangen, A.; Ballot, A.; Nowak, P.; Schneider, S.C. Charophytes in warm springs on Svalbard (Spitsbergen): DNA barcoding identifies Chara aspera and Chara canescens with unusual morphological traits. Bot. Lett. 2020, 167, 179–186. [Google Scholar] [CrossRef]
- Langangen, A. Lokalitetsliste 2021 Status pr. 1. Februar 2021. Norges Kransalger Hefte 13; Privately Published: Oslo, Norway, 2021. [Google Scholar]
- Wood, R.D.; Imahori, K. A Revision of the Characeae. Second Part: Iconography of the Characeae; Verlag Von J. Cramer: Weinheim, Germany, 1964. [Google Scholar]
- Migula, W. Characeae Rossicae ex herbario Horti Petropolitani, determinatae et descriptae a prof. W. Migula (Karlsruhe). Acta Horti Petropolitani 1904, 23, 533–539. [Google Scholar]
- Ling, Y.J.; Xie, S.L.; Langangen, A. Charales of China. Nova Hedwig. 2000, 71, 69–94. [Google Scholar]
- Noedoost, F.; Riahi, H.; Sheidai, M.; Ahmadi, A. Distribution of charophytes from Iran with three new records of Characeae (Charales, Chlorophyta). Cryptogam. Algol. 2015, 36, 389–405. [Google Scholar] [CrossRef]
- Romanov, R.E. New interesting records of charophytes (Charales, Charophyceae) from Eurasia and Africa. Webbia 2019, 74, 159–166. [Google Scholar] [CrossRef]
- Romanov, R.E.; Chemeris, E.V.; Zhakova, L.V.; Ivanova, A.V.; Palagushkina, O.V. The charophytes (Charales, Charophyceae) from the Middle Volga Region (Russia): Synopsis of localities and species protection. Nat. Conserv. Res. 2018, 3, 1–20. [Google Scholar] [CrossRef]
- Proctor, V.W. Chara globularis Thuillier (= C. fragilis Desvaux): Breeding patterns within a cosmopolitan complex. Limnol. Oceanogr. 1971, 16, 422–436. [Google Scholar] [CrossRef]
- Proctor, V.W. The nature of charophyte species. Phycologia 1975, 14, 97–113. [Google Scholar] [CrossRef]
- Cantonati, M.; Fensham, R.J.; Stevens, L.E.; Gerecke, R.; Glazier, D.S.; Goldscheider, N.; Knight, R.L.; Richardson, J.S.; Springer, A.E.; Tockner, K. Urgent plea for global protection of springs. Conserv. Biol. 2020, 35, 378–382. [Google Scholar] [CrossRef]
- Ahmadi, A.; Riahi, H.; Sheidai, M.; Raam, J.V. Some Charophytes (Characeae, Charophyta) from Central and Western of Iran Including Chara kohrangiana species nova. Cryptogam. Algol. 2012, 33, 359–390. [Google Scholar] [CrossRef]
- Stewart, N.F.; Church, J.M. Red Data Books of Britain and Ireland: Stoneworts; Joint Nature Conservation Committee: Peterborough, UK, 1993; ISBN 978-1-873701-24-9. [Google Scholar]
- El-Naggar, M.E.E. Comparative biochemical studies on the vegetative and reproductive stages of Chara vulgaris. J. King Saud Univ. Sci. 1995, 7, 191–204. [Google Scholar]
- Bueno, N.C.; Bicudo, C.E.M. Characeae (Charophyceae) do pantanal de Mato Grosso do Sul, Brasil: Nitella. Hoehnea 1997, 24, 29–55. [Google Scholar]
- Bueno, N.C.; Meurer, T.; Biolo, S.; Bicudo, C.E.M. Novos registros de Nitella (Chlorophyta, Characeae) para regiões dos Estados de Mato Grosso e Mato Grosso do Sul, Brasil. Hoehnea 2011, 38, 385–396. [Google Scholar] [CrossRef]
- Bueno, N.C.; Prado, J.F.; Meurer, T.; Bicudo, C.E.d.M. Nitella (Streptophyta, Characeae) from southern Brazil. Iheringia Sér. Bot. 2016, 71, 132–154. [Google Scholar]
- Ahmed, Z.U.; Khondker, M.; Begum, Z.N.T.; Hassan, M.A.; Kabir, S.M.H.; Ahmad, M.; Ahmed, A.T.A.; Rhaman, A.K.A. Algae, Charophyta—Rhodophyta: Achnanthaceae–Vaucheriaceae. In Encyclopedia of Flora and Fauna of Bangladesh, Volume-4 Algae: Charophyta-Rhodophyta (Achnanthaceae–Vaucheriaceae); Zia Uddin, A., Ed.; Asiatic Society of Bangladesh: Dhaka, Bangladesh, 2009; ISBN 984-300-000-286-0. [Google Scholar]
- Blindow, I.; Schories, D.; Schubert, H. Charophytes of Chile—Taxonomy and distribution. Part 2. Subfamily Nitelleae. Nova Hedwig. 2018, 107, 49–90. [Google Scholar] [CrossRef]
- Powell, O.; Fensham, R. The history and fate of the Nubian Sandstone Aquifer springs in the oasis depressions of the Western Desert, Egypt. Hydrogeol. J. 2016, 24, 395–406. [Google Scholar] [CrossRef]
- Abdel-Shafy, H.I.; Kamel, A.H. Groundwater in Egypt issue: Resources, location, amount, contamination, protection, renewal, future overview. Egypt. J. Chem. 2016, 59, 321–362. [Google Scholar] [CrossRef]
- Urbaniak, J. A SEM and light microscopy study of the oospore wall ornamentation in Polish charophytes (Charales, Charophyceae)—Genus Chara. Nova Hedwig. 2011, 93, 1–28. [Google Scholar] [CrossRef]
- Chapman, H.D.; Pratt, P.F. Methods of Analysis for Soils, Plants, and Waters; Division of Agricultural Sciences, University of California: Riverside, CA, USA, 1978. [Google Scholar]
- Clesceri, L.S.; Greenberg, A.E.; Eaton, A.D. Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Public Health Association: Washington, DC, USA, 2000. [Google Scholar]
- Echt, C.S.; Erdahl, L.A.; McCoy, T.J. Genetic segregation of random amplified polymorphic DNA in diploid cultivated alfalfa. Genome 1992, 35, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Kiselev, K.V.; Dubrovina, A.S.; Tyunin, A.P. The methylation status of plant genomic DNA influences PCR efficiency. J. Plant Physiol. 2015, 175, 59–67. [Google Scholar] [CrossRef]
- Zurawski, G.; Clegg, M.T. Evolution of higher-plant chloroplast DNA-encoded genes: Implications for structure-function and phylogenetic studies. Annu. Rev. Plant Physiol. 1987, 38, 391–418. [Google Scholar] [CrossRef]
- Karol, K.G. Phylogenetic Studies of the Charales: The Closest Living Relatives of Land Plants. Ph.D. Thesis, University of Maryland, Baltimore, MD, USA, 2004; 164p. [Google Scholar]
- Pryer, K.M.; Smith, A.R.; Hunt, J.S.; Dubuisson, J.Y. rbcL data reveal two monophyletic groups of filmy ferns (Filicopsida: Hymenophyllaceae). Am. J. Bot. 2001, 88, 1118–1130. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.D.; Fucikova, K.; Lo, C.; Lewis, L.A.; Karol, K.G. An assessment of proposed DNA barcodes in freshwater green algae. Cryptogam. Algol. 2010, 31, 529–555. [Google Scholar] [CrossRef]
- Bonfield, J.K.; Smith, K.F.; Staden, R. A New DNA sequence assembly program. Nucleic Acids Res. 1995, 23, 4992–4999. [Google Scholar] [CrossRef] [PubMed]
- Galtier, N.; Gouy, M.; Gautier, C. SEAVIEW and PHYLO_WIN: Two graphic tools for sequence alignment and molecular phylogeny. Comput. Appl. Biosci. 1996, 12, 543–548. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods), Version 4.0b10; Sinauer Associates, Inc.: Sunderland, MA, USA, 2002. [Google Scholar]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed]
- Akaike, H. A New look at the statistical model identification. IEEE Trans. Automat. Contr. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [PubMed]
- Lambert, S.J.; Davy, A.J. Water quality as a threat to aquatic plants: Discriminating between the effects of nitrate, phosphate, boron and heavy metals on Charophytes. New Phytol. 2011, 189, 1051–1059. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Unit | Chara aspera | Chara contraria | Chara globata | Chara tomentosa | Chara vulgaris | Nitella flagellifera | Tolypella sp. | ||
|---|---|---|---|---|---|---|---|---|---|---|
| (PBA–1801) | (PBA–1603) | (PBA–1701) | (PBA–1604) | (PBA– 1702) | (PBA–1601) | (PBA–1602) | (PBA–1703) | (PBA–1704) | ||
| Temperature | °C | 27.7 | 21.9 | 20.6 | 26.5 | 27.2 | 31.6 | 23.8 | 31.5 | 19.2 |
| pH | 7.32 | 6.93 | 7.46 | 6.85 | 7.53 | 6.71 | 6.16 | 8.13 | 7.64 | |
| Conductivity | µS·cm−1 | 6350 | 760 | 2960 | 6280 | 7700 | 4470 | 960 | 6670 | 360 |
| T.D.S. | mg·L−1 | 3260 | 400 | 2010 | 3110 | 6160 | 2240 | 460 | 5335 | 244 |
| DO | mg·L−1 | 2.6 | 4.2 | 6.3 | 3.4 | 7.1 | 2.2 | 1.9 | 3.8 | 3.3 |
| Ca2+ | mg·L−1 | 276.0 | 61.8 | 146.9 | 94 | 653.7 | 134.9 | 29.9 | 110 | 38.2 |
| K+ | mg·L−1 | 59.3 | 19.3 | 89.7 | 28.6 | 146.6 | 32.1 | 24.8 | 135.6 | 18.4 |
| Mg2+ | mg·L−1 | 104 | 16.1 | 52 | 46.4 | 72.9 | 79.2 | 16.6 | 104.9 | 5.7 |
| Na+ | mg·L−1 | 1139.6 | 62.1 | 360.9 | 1430.3 | 1072.3 | 545 | 25.5 | 1368.2 | 17.2 |
| Cl− | mg·L−1 | 2104.2 | 146.4 | 768.5 | 1794.8 | 2543.2 | 1235.5 | 131 | 2378.1 | 15 |
| SO42- | mg·L−1 | 399.8 | 74.6 | 356.5 | 989 | 582.2 | 300 | 36 | 355.7 | 88.1 |
| HCO3− | mg·L−1 | 350.7 | 117.1 | 31.2 | 86.3 | 316.1 | 220.6 | 314.4 | 161.2 | 81.8 |
| CO32− | mg·L−1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| NO2− | µg·L−1 | 15 | 462 | 0 | 120 | 0 | 11 | 8 | 46 | 0 |
| NO3− | µg·L−1 | 460 | 5400 | 2320 | 200 | 1765 | 200 | 400 | 1570 | 421 |
| NH4+ | µg·L−1 | 68 | 924 | 57 | 270 | 114 | 271 | 150 | 86 | 19 |
| TP | µg·L−1 | 260 | 150 | 1680 | 135 | 715 | 20 | 100 | 2750 | 217 |
| SRP | µg·L−1 | 55 | 40 | 276 | 100 | 295 | 17 | 48 | 421 | 62 |
| SiO2 | mg·L−1 | 9.3 | 15.2 | 4.8 | 0.33 | 7.9 | 7.7 | 7.1 | 6.2 | 1.5 |
| Fe | µg·L−1 | 475 | 264 | 1500 | 430 | 5500 | 19 | 59 | 2100 | 175 |
| Mn | µg·L−1 | 105 | 120 | 315 | 120 | 191 | 1.30 | 4 | 450 | 17 |
| Cu | µg·L−1 | 17 | 15 | 81 | 30 | 130 | 0.3 | 0.36 | 134 | 52 |
| Zn | µg·L−1 | 135 | 11 | 255 | 300 | 360 | 1.7 | 8 | 263 | 81 |
| Species | Codes | Site Description | Collection Date | Latitude (N) | Longitude (E) | Elevation (m) | GenBank Accession Numbers | |
|---|---|---|---|---|---|---|---|---|
| rbcL | ITS1 | |||||||
| Chara aspera | PBA–1801 | mineral spring-fed agricultural ditch in the Siwa Oasis | 6 May 2018 | 29° 13′ 9″ | 25° 31′ 59″ | −9 | MK770143 | MK705919 |
| Chara contraria | PBA–1603 | nutrient-rich artificial muddy pool in Wadi El-Arbaeen, Saint Catherine Protectorate, South Sinai | 6 October 2016 | 28º 32′ 14.8″ | 33º 57′ 41.8″ | 1732 | MK770144 | MK705918 |
| PBA–1701 | agricultural ditch in El-Dakhla Oasis | 6 March 2017 | 25° 29′ 32.687″ | 29° 6′ 52.889″ | 126 | MH424122 | MH426800 | |
| Chara globata | PBA–1604 | thermal mineral desert spring ‘Ain Wazedi’ in the Siwa Oasis | 14 October 2016 | 29° 14′ 24.3″ | 25° 29′ 45.4″ | −20 | MH424119 | – |
| Chara tomentosa | PBA–1702 | shallow marsh in El-Dakhla Oasis | 7 March 2017 | 25° 32′ 13.48559″ | 29° 3′ 13.7921″ | 112 | MH424123 | – |
| Chara vulgaris | PBA–1601 | outlet channel of the thermal mineral desert spring ‘Ain Al-Maamal’ in the Siwa Oasis | 14 October 2016 | 29° 12′ 21.4″ | 25° 31′ 52.3″ | −9.9 | KY449161 | – |
| PBA–1602 | agricultural ditch in El-Farafra Oasis | 16 October 2016 | 27° 03′ 26.1″ | 27° 57′ 47.4″ | 104 | KY449162 | – | |
| Nitella flagellifera | PBA–1703 | agricultural ditch in El-Dakhla Oasis | 6 March 2017 | 25° 33′ 35.6623″ | 28° 59′ 4.83688″ | 107 | MH424121 | MH426799 |
| Tolypella sp. | PBA–1704 | shallow wetland in El-Dakhla Oasis | 6 March 2017 | 25° 33′ 56.68333″ | 28° 56′ 42.04014″ | 92 | MH424120 | MH426798 |
| Dataset/Parameter | Chara | Chara+Nitella+Tolypella | Nitella | Tolypella |
|---|---|---|---|---|
| Marker | ITS1 | rbcL | rbcL+ITS1 | rbcL+ITS1 |
| Number of sequences | 19 | 121 | 41 | 16 |
| Model | HKY+G | GTR+I+G | GTR+I+G | TrN+G |
| Number of runs/Markov chains for BI | 4/4 | 4/4 | 4/4 | 4/4 |
| Number of generations for BI | 600,000 | 10,000,000 | 1,000,000 | 500,000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saber, A.A.; Gontcharov, A.A.; Nikulin, A.Y.; Nikulin, V.Y.; Rayan, W.A.; Cantonati, M. Integrative Taxonomic, Ecological and Genotyping Study of Charophyte Populations from the Egyptian Western-Desert Oases and Sinai Peninsula. Plants 2021, 10, 1157. https://doi.org/10.3390/plants10061157
Saber AA, Gontcharov AA, Nikulin AY, Nikulin VY, Rayan WA, Cantonati M. Integrative Taxonomic, Ecological and Genotyping Study of Charophyte Populations from the Egyptian Western-Desert Oases and Sinai Peninsula. Plants. 2021; 10(6):1157. https://doi.org/10.3390/plants10061157
Chicago/Turabian StyleSaber, Abdullah A., Andrey A. Gontcharov, Arthur Yu. Nikulin, Vyacheslav Yu. Nikulin, Walaa A. Rayan, and Marco Cantonati. 2021. "Integrative Taxonomic, Ecological and Genotyping Study of Charophyte Populations from the Egyptian Western-Desert Oases and Sinai Peninsula" Plants 10, no. 6: 1157. https://doi.org/10.3390/plants10061157
APA StyleSaber, A. A., Gontcharov, A. A., Nikulin, A. Y., Nikulin, V. Y., Rayan, W. A., & Cantonati, M. (2021). Integrative Taxonomic, Ecological and Genotyping Study of Charophyte Populations from the Egyptian Western-Desert Oases and Sinai Peninsula. Plants, 10(6), 1157. https://doi.org/10.3390/plants10061157