
The Antiviral and Virucidal Activities of Voacangine and Structural Analogs Extracted from Tabernaemontana cymosa Depend on the Dengue Virus Strain

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
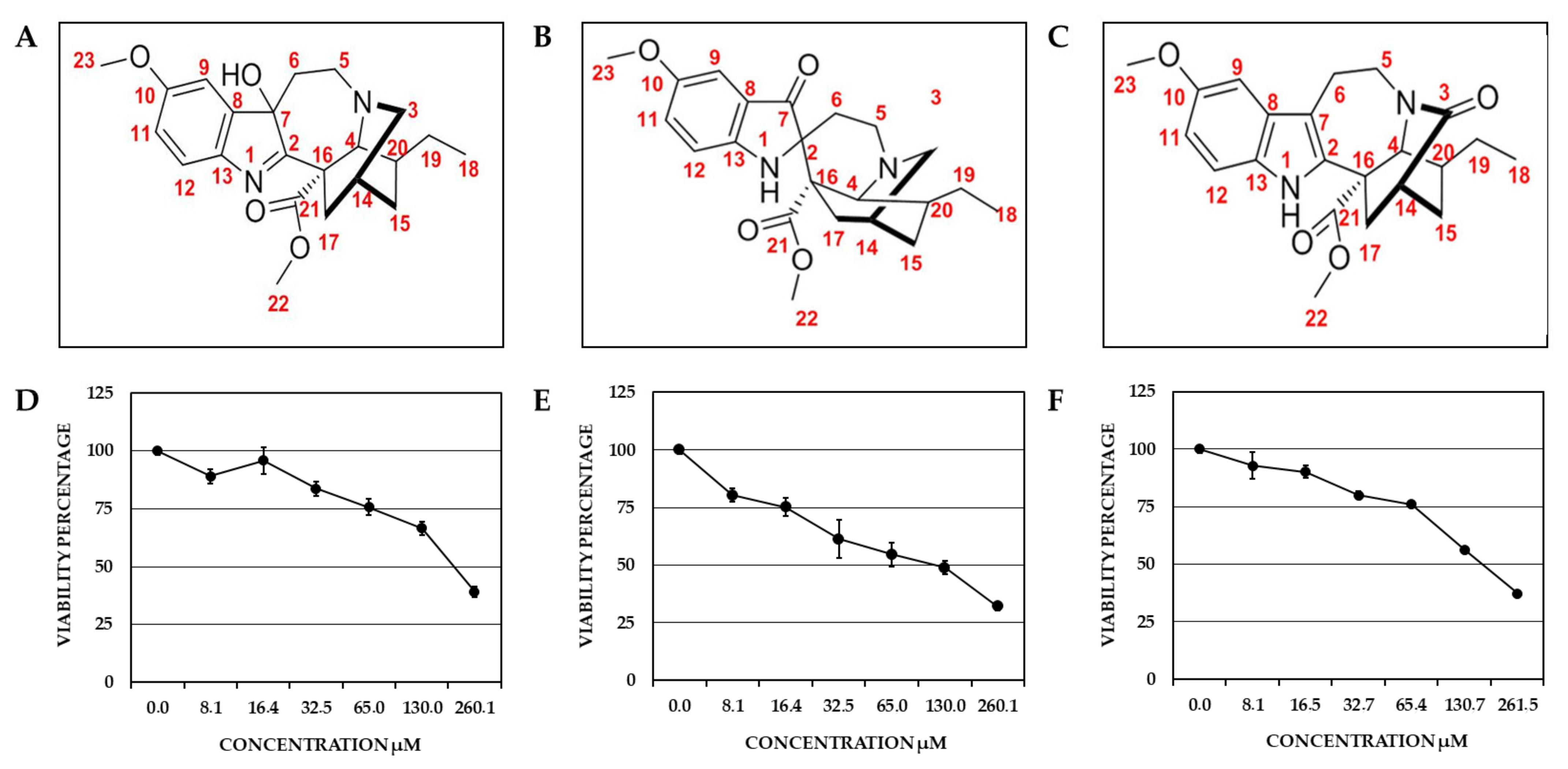
2.1. Identification of the Compounds Derived from T. cymosa and Their Toxic Effect on Vero Cells
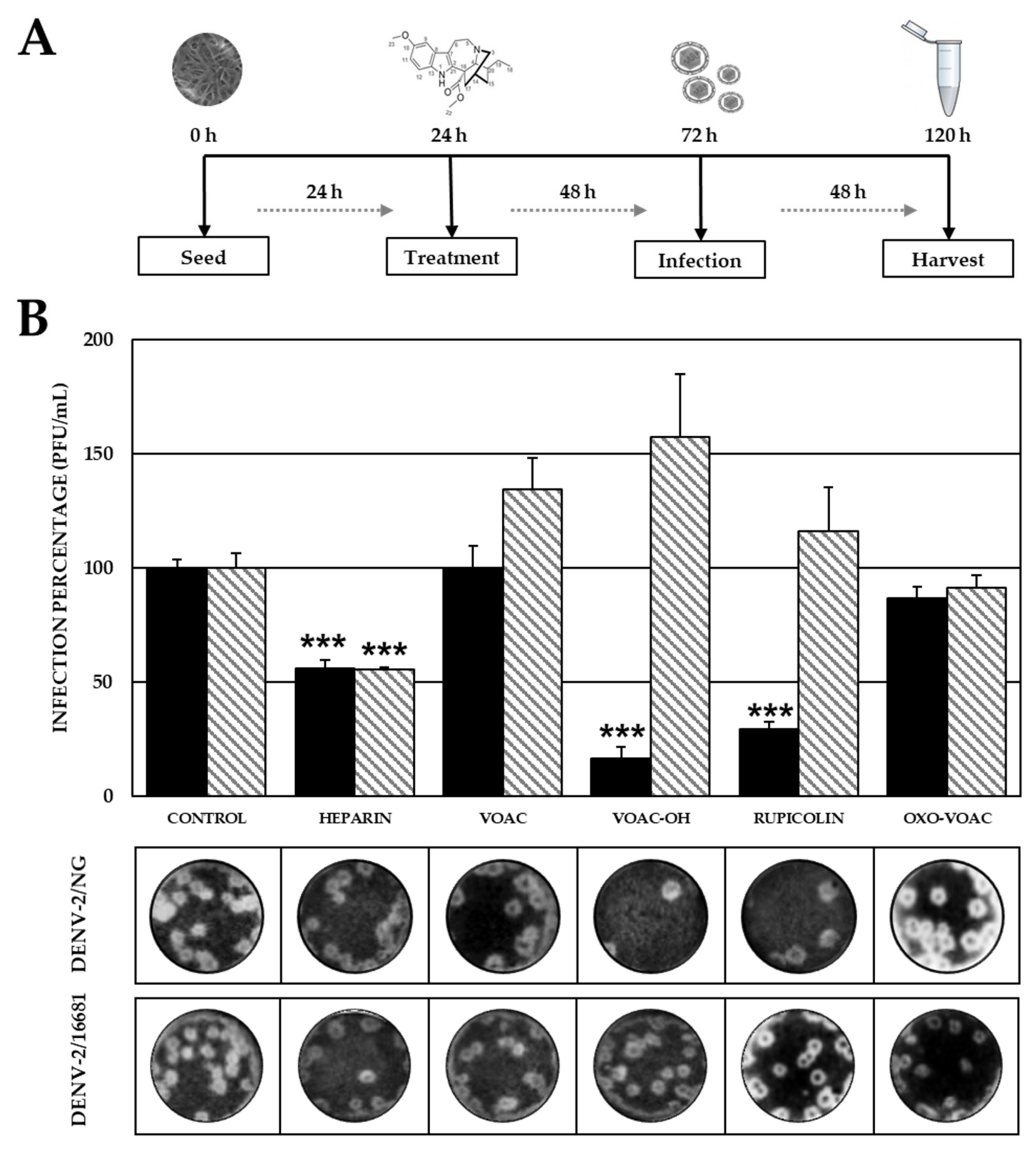
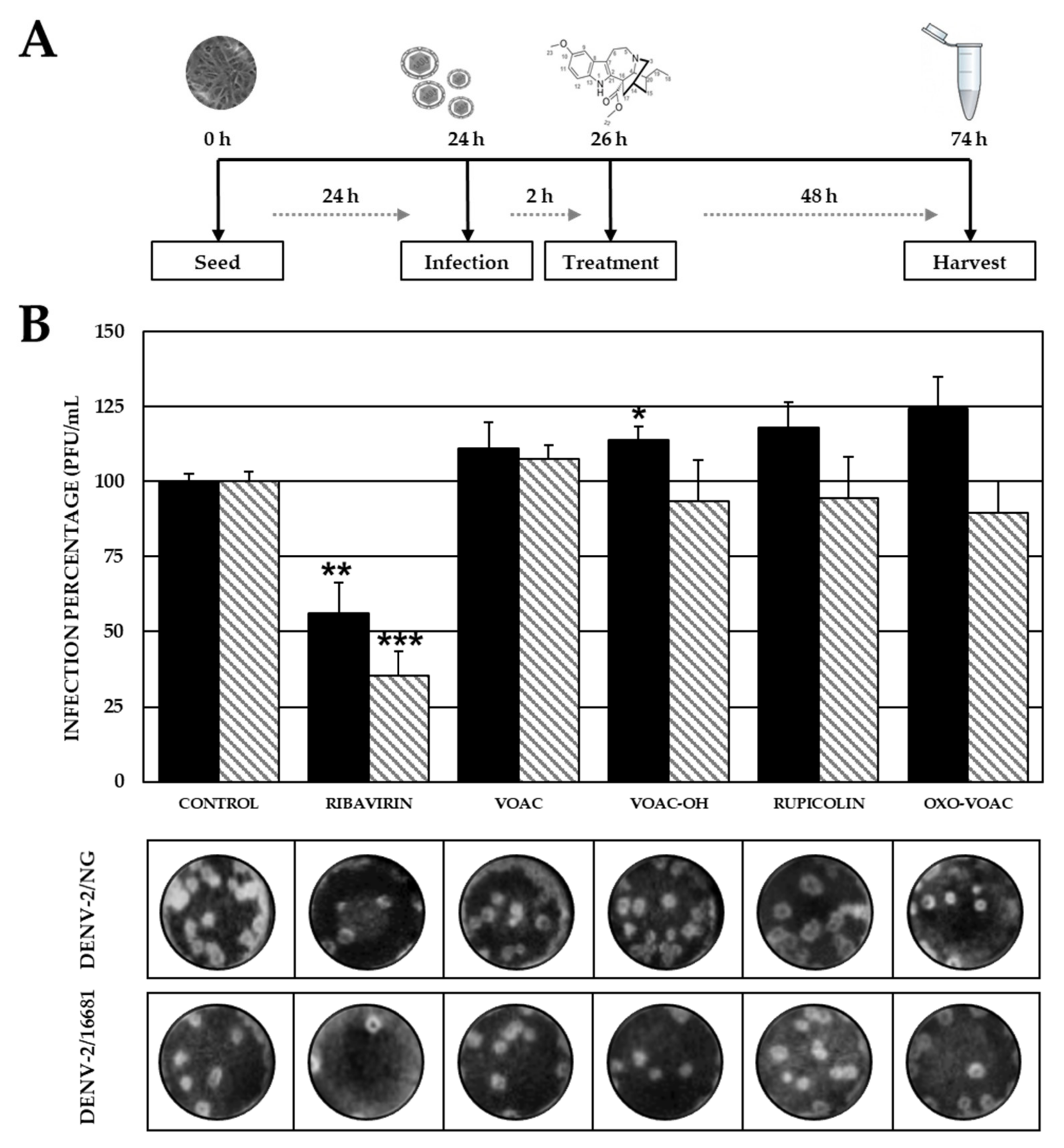
2.2. Pre-Treatment with VOAC-OH and Rupicoline Inhibits the Infection by DENV-2/NG Strain
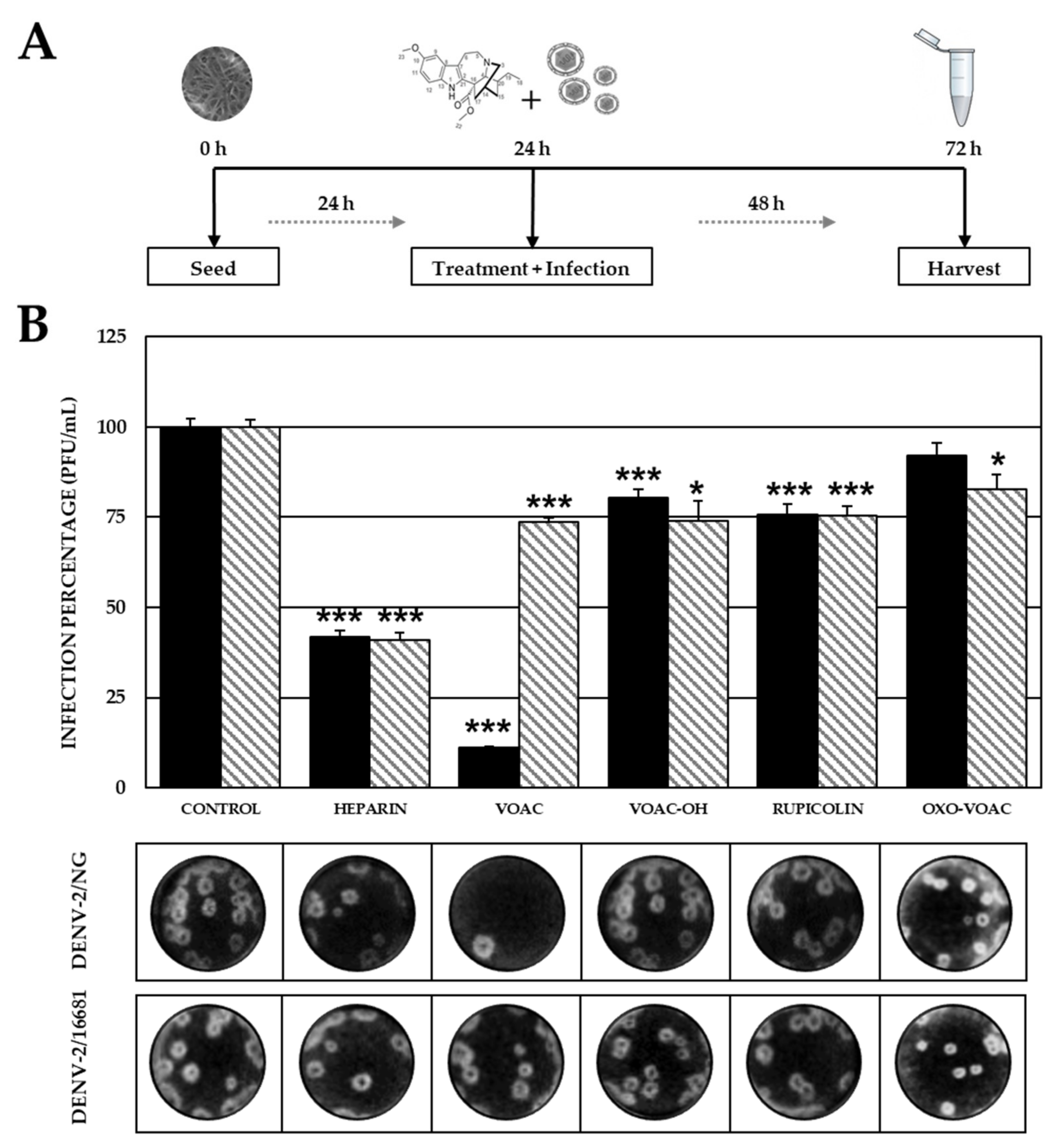
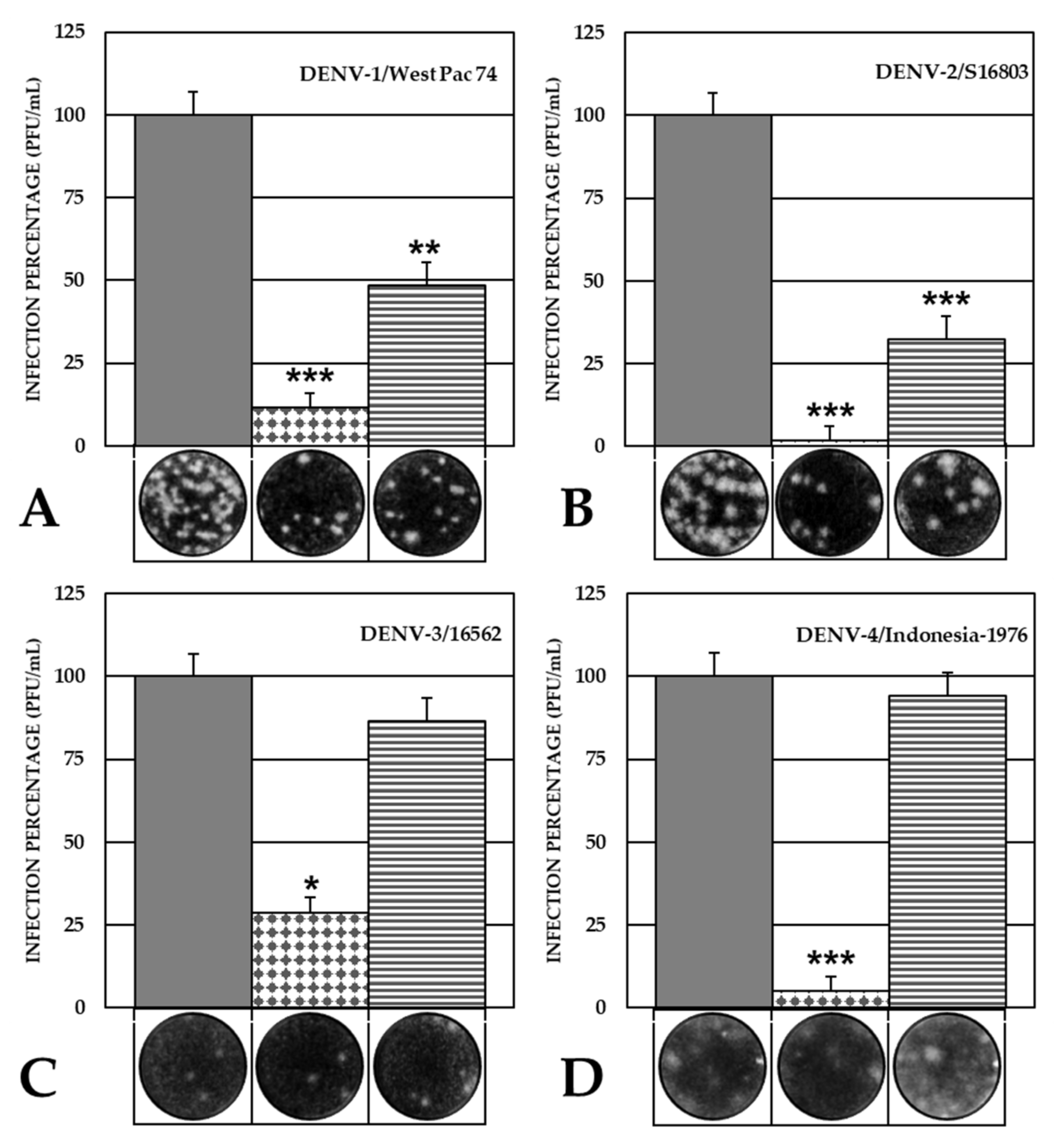
2.3. The VOAC Virucidal Effect Depends on the Serotype of the Virus
2.4. None of the Compounds Inhibit Post-Entry Stages of the Viral Reproductive Cycle
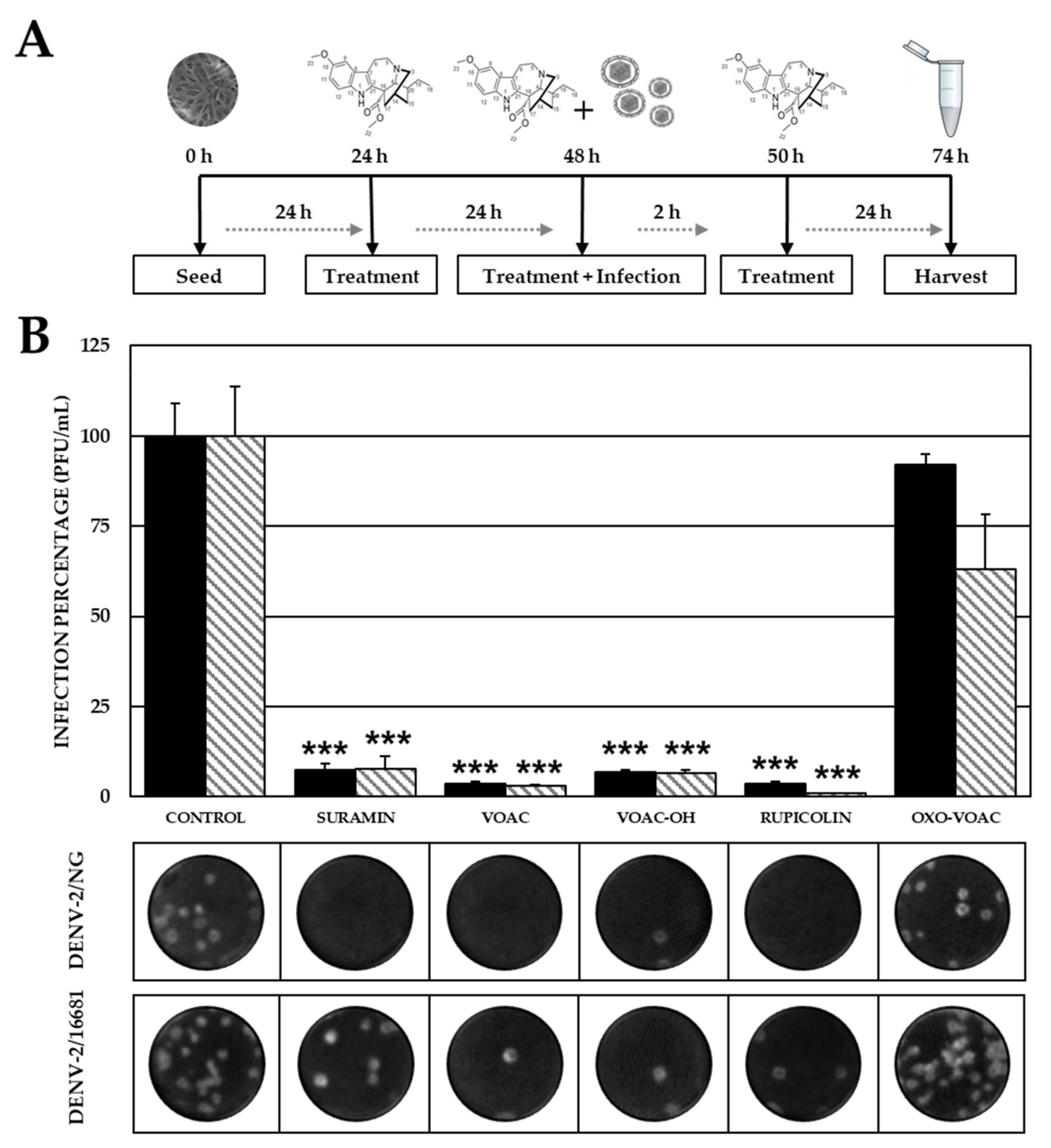
2.5. The Combined Antiviral Approach Using VOAC, VOAC-OH and Rupicoline Favors the Antiviral Effect Compared with Individual Approaches
2.6. Non-Synonymous Amino Acid Differences in the Envelope Proteins of DENV-2/NG and DENV-16681
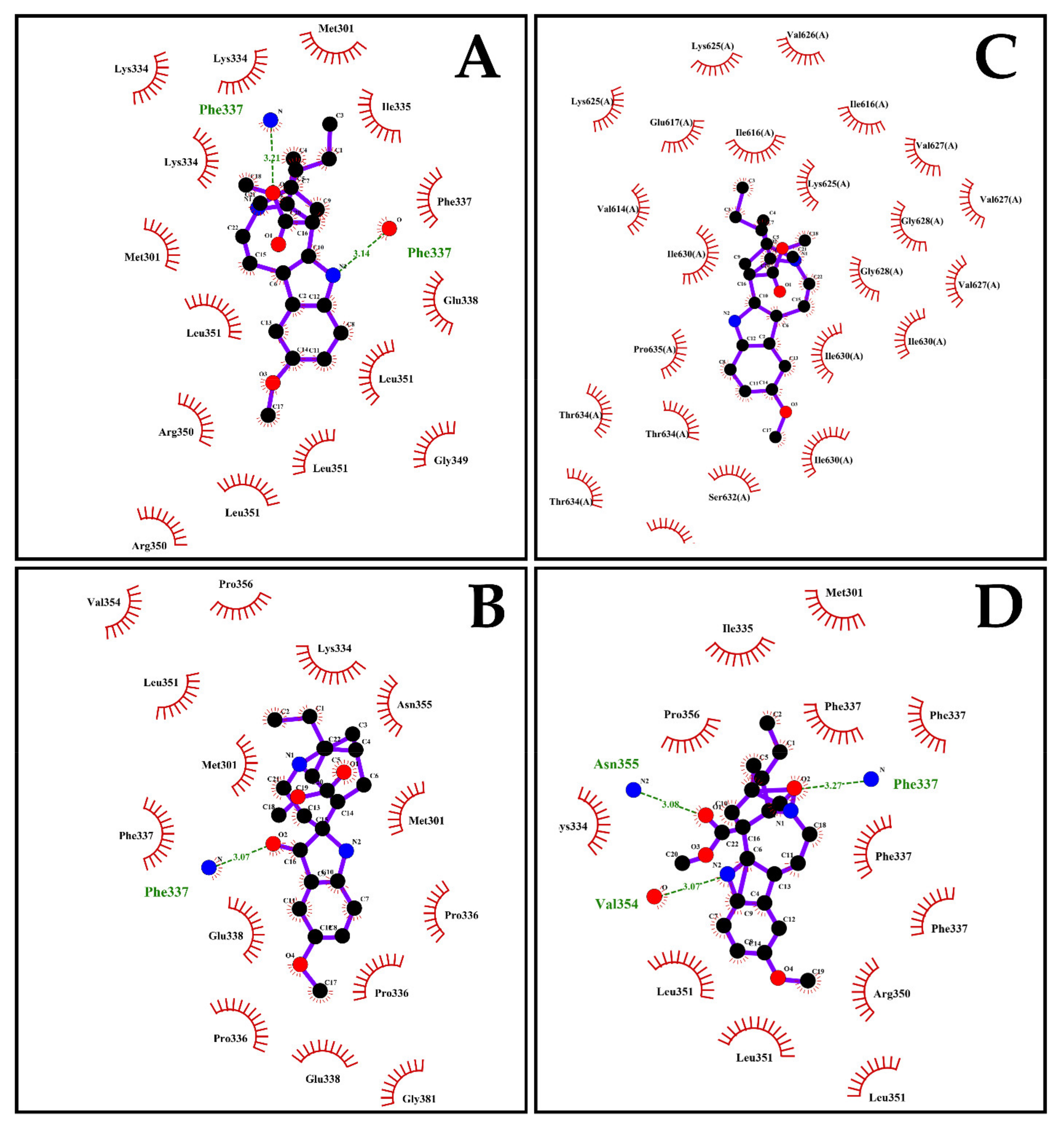
2.7. The Energy of Binding of Indole Alkaloids and the Structural E Protein of the DENV Serotypes Is Favorable via Molecular Docking
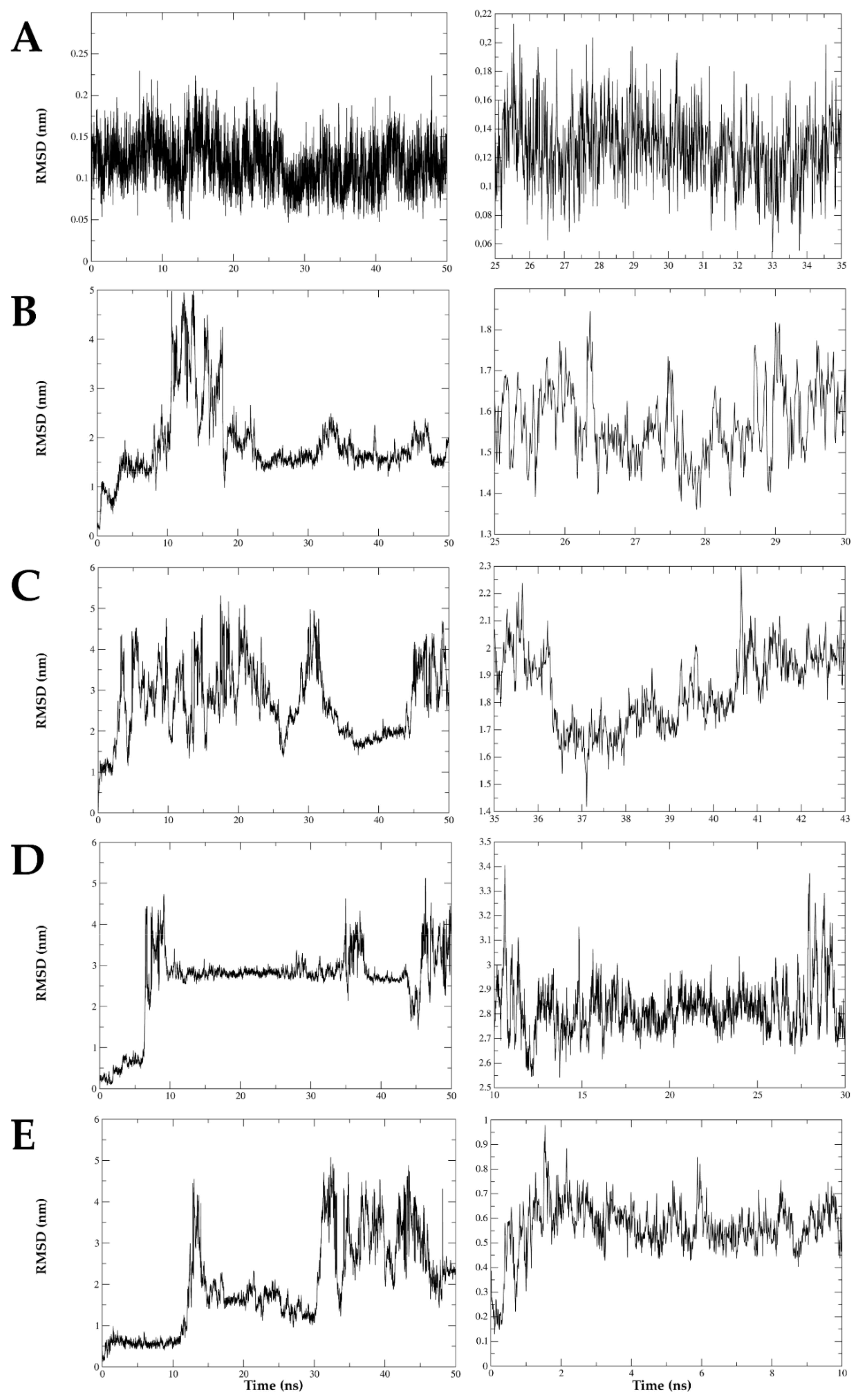
2.8. The Interaction between DENV E Protein Domain III and VOAC Is Stable over Time via Molecular Dynamics
3. Discussion
4. Materials and Methods
4.1. Isolation and Identification of Compounds
4.2. Virus and Cell Maintenance
4.3. Determination of the Cytotoxicity of Isolated Compounds
4.4. Antiviral In Vitro Assays
4.4.1. Antiviral Strategies
4.4.2. Quantification of Infectious Viral Particles by Plaque Assay
4.4.3. Statistical Analysis
4.5. In Silico Studies
4.5.1. Gene Sequence Analysis
4.5.2. Selection of the Target Protein
4.5.3. Selection of Ligand
4.5.4. Molecular Docking
4.5.5. Molecular Dynamics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tapia-Conyer, R.; Betancourt-Cravioto, M.; Méndez-Galván, J. Dengue: An escalating public health problem in Latin America. Paediatr. Int. Child Health 2012, 32, 14–17. [Google Scholar] [CrossRef] [Green Version]
- Gubler, D.J. Aedes aegypti and Aedes aegypti-Borne Disease Control in the 1990s: Top Down or Bottom Up. Am. J. Trop. Med. Hyg. 1989, 40, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Istúriz, R.E.; Gubler, D.J.; Del Castillo, J.B. Dengue and dengue hemorrhagic fever in Latin America and the Caribbean. Infect. Dis. Clin. 2000, 14, 121–140. [Google Scholar] [CrossRef]
- Apte-Sengupta, S.; Sirohi, D.; Kuhn, R.J. Coupling of replication and assembly in flaviviruses. Curr. Opin. Virol. 2014, 9, 134–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, T.J.; Hahn, C.S.; Galler, R.; Rice, C.M. Flavivirus genome organization, expression, and replication. Annu. Rev. Microbiol. 1990, 44, 649–688. [Google Scholar] [CrossRef] [PubMed]
- Rico-Hesse, R.; Nisalak, A.; Harrison, L.M.; Vaughn, D.W.; Green, S.; Ennis, F.A.; Kalayanarooj, S.; Rothman, A.L. Molecular evolution of dengue type 2 virus in Thailand. Am. J. Trop. Med. Hyg. 1998, 58, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Leitmeyer, K.C.; Vaughn, D.W.; Watts, D.M.; Salas, R.; Villalobos, I.; Chacon, D.; Ramos, C.; Rico-Hesse, R. Dengue Virus Structural Differences That Correlate with Pathogenesis. J. Virol. 1999, 73, 4738–4747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Betancur, V.; Marín-Villa, M.; Martínez-Gutierrez, M. Infection of epithelial cells with dengue virus promotes the expression of proteins favoring the replication of certain viral strains. J. Med. Virol. 2014, 86, 1448–1458. [Google Scholar] [CrossRef]
- Martínez-Betancur, V.; Martinez-Gutierrez, M. Proteomic profile of human monocytic cells infected with dengue virus. Asian Pac. J. Trop. Biomed. 2016, 6, 914–923. [Google Scholar] [CrossRef]
- Cruz-Oliveira, C.; Freire, J.M.; Conceição, T.M.; Higa, L.M.; Castanho, M.A.; Da Poian, A.T. Receptors and routes of dengue virus entry into the host cells. FEMS Microbiol. Rev. 2015, 39, 155–170. [Google Scholar] [CrossRef] [Green Version]
- Mosso, C.; Galván-Mendoza, I.J.; Ludert, J.E.; del Angel, R.M. Endocytic pathway followed by dengue virus to infect the mosquito cell line C6/36 HT. Virology 2008, 378, 193–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elshuber, S.; Allison, S.L.; Heinz, F.X.; Mandl, C.W. Cleavage of protein prM is necessary for infection of BHK-21 cells by tick-borne encephalitis virusFN1. J. Gen. Virol. 2003, 84, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Magden, J.; Kääriäinen, L.; Ahola, T. Inhibitors of virus replication: Recent developments and prospects. Appl. Microbiol. Biotechnol. 2005, 66, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Denaro, M.; Smeriglio, A.; Barreca, D.; De Francesco, C.; Occhiuto, C.; Milano, G.; Trombetta, D. Antiviral activity of plants and their isolated bioactive compounds: An update. Phytother. Res. 2020, 34, 742–768. [Google Scholar] [CrossRef] [PubMed]
- Cortez-Gallardo, V.; Macedo-Ceja, J.P.; Hernández-Arroyo, M.; Arteaga-Aureoles, G.; Espinosa-Galván, D.; Rodríguez-Landa, J.F. Farmacognosia: Breve historia de sus orígenes y su relación con las ciencias médicas. Rev. Bioméd. 2004, 15, 123–136. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Castro, C.; Diaz-Castillo, F.; Martinez-Gutierrez, M. Ethanol extracts of Cassia grandis and Tabernaemontana cymosa inhibit the in vitro replication of dengue virus serotype 2. Asian Pac. J. Trop. Dis. 2015, 5, 98–106. [Google Scholar] [CrossRef]
- Gómez-Calderón, C.; Mesa-Castro, C.; Robledo, S.; Gómez, S.; Bolivar-Avila, S.; Diaz-Castillo, F.; Martínez-Gutierrez, M. Antiviral effect of compounds derived from the seeds of Mammea americana and Tabernaemontana cymosa on Dengue and Chikungunya virus infections. BMC Complement. Altern. Med. 2017, 17, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Trujillo-Correa, A.I.; Quintero-Gil, D.C.; Diaz-Castillo, F.; Quiñones, W.; Robledo, S.M.; Martinez-Gutierrez, M. In vitro and in silico anti-dengue activity of compounds obtained from Psidium guajava through bioprospecting. BMC Complement. Altern. Med. 2019, 19, 1–16. [Google Scholar] [CrossRef]
- Endress, M.E.; Liede-Schumann, S.; Meve, U. An updated classification for Apocynaceae. Phytotaxa 2014, 159, 175–194. [Google Scholar] [CrossRef]
- Dey, A.; Mukherjee, A.; Chaudhury, M. Alkaloids from apocynaceae: Origin, pharmacotherapeutic properties, and structure-activity studies. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2017; Volume 52, pp. 373–488. [Google Scholar]
- Anbukkarasi, M.; Thomas, P.A.; Sheu, J.-R.; Geraldine, P. In vitro antioxidant and anticataractogenic potential of silver nanoparticles biosynthesized using an ethanolic extract of Tabernaemontana divaricata leaves. Biomed. Pharmacother. 2017, 91, 467–475. [Google Scholar] [CrossRef]
- Thambi, P.T.; Kuzhivelil, B.; Sabu, M.; Jolly, C. Antioxidant and antiinflammatory activities of the flowers of Tabernaemontana coronaria (L) R.BR. Indian J. Pharm. Sci. 2006, 68, 352–355. [Google Scholar]
- De Almeida, L.; Cintra, A.C.; Veronese, E.L.; Nomizo, A.; Franco, J.J.; Arantes, E.C.; Giglio, J.R.; Sampaio, S.V. Anticrotalic and antitumoral activities of gel filtration fractions of aqueous extract from Tabernaemontana catharinensis (Apocynaceae). Comp. Biochem. Physiol. Part C Toxicol. Pharm. 2004, 137, 19–27. [Google Scholar] [CrossRef]
- Van Beek, T.; Kuijlaars, F.; Thomassen, P.; Verpoorte, R.; Svendsen, A.B. Antimicrobially active alkaloids from Tabernaemontana pachysiphon. Phytochemistry 1984, 23, 1771–1778. [Google Scholar] [CrossRef]
- Díaz Castillo, F.; Morelos Cardona, S.M.; Carrascal Medina, M.; Pájaro González, Y.; Gómez Estrada, H. Actividad larvicida de extractos etanólicos de Tabernaemontana cymosa y Trichilia hirta sobre larvas de estadio III y IV de Aedes aegypti (Diptera: Culicidae). Rev. Cuba. Plantas Med. 2012, 17, 256–267. [Google Scholar]
- Pereira, P.S.; França, S.D.C.; De Oliveira, P.V.A.; Breves, C.M.D.S.; Pereira, S.I.V.; Sampaio, S.V.; Nomizo, A.; Dias, D.A. Chemical constituents from Tabernaemontana catharinensis root bark: A brief NMR review of indole alkaloids and in vitro cytotoxicity. Quím. Nova 2008, 31, 20–24. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Cordell, G.A. Heyneanine Hydroxyindolenine, A New Indole Alkaloid from Ervatamia coronaria var. plena. J. Nat. Prod. 1988, 51, 528–531. [Google Scholar] [CrossRef]
- Husain, K.; Said, I.M.; Din, L.B.; Takayama, H.; Kitajima, M.; Aimi, N. Alkaloids from The Roots of Tabernaemontana Macrocarpa Jack. Nat. Prod. Sci. 1997, 3, 42–48. [Google Scholar]
- Achenbach, H.; Benirschke, M.; Torrenegra, R. Alkaloids and other compounds from seeds of Tabernaemontana cymosa. Phytochemistry 1997, 45, 325–335. [Google Scholar] [CrossRef]
- Farrow, S.C.; Kamileen, M.O.; Meades, J.; Ameyaw, B.; Xiao, Y.; O’Connor, S.E. Cytochrome P450 and O-methyltransferase catalyze the final steps in the biosynthesis of the anti-addictive alkaloid ibogaine from Tabernanthe iboga. J. Biol. Chem. 2018, 293, 13821–13833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krengel, F.; Herrera Santoyo, J.; Olivera Flores, T.D.J.; Chávez Ávila, V.M.; Pérez Flores, F.J.; Reyes Chilpa, R. Quantification of anti-addictive alkaloids ibogaine and voacangine in in vivo-and in vitro-grown plants of two Mexican Tabernaemontana species. Chem. Biodivers. 2016, 13, 1730–1737. [Google Scholar] [CrossRef] [PubMed]
- Bardiot, D.; Koukni, M.; Smets, W.; Carlens, G.; McNaughton, M.; Kaptein, S.; Dallmeier, K.; Chaltin, P.; Neyts, J.; Marchand, A. Discovery of Indole Derivatives as Novel and Potent Dengue Virus Inhibitors. J. Med. Chem. 2018, 61, 8390–8401. [Google Scholar] [CrossRef] [PubMed]
- Laura, G.F.M.; Njoya, E.M.; Jouda, J.-B.; Kweka, B.N.W.; Mbazoa, C.D.; Wang, F.; Seguin, E.; Wandji, J. A new cytotoxic indole alkaloid from Tabernaemontana inconspicua stapf. Nat. Prod. Res. 2021, 35, 1590–1595. [Google Scholar] [CrossRef] [PubMed]
- Sundar, L.; Chang, F.N. Antimicrobial activity and biosynthesis of indole antibiotics produced by Xenorhabdus nematophilus. Microbiology 1993, 139, 3139–3148. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, S.; Schiller, G.; Lister, J.; Damon, L.; Goldberg, S.; Aulitzky, W.; Ben-Yehuda, D.; Stock, W.; Coutre, S.; Douer, D.; et al. High-Dose Vincristine Sulfate Liposome Injection for Advanced, Relapsed, and Refractory Adult Philadelphia Chromosome–Negative Acute Lymphoblastic Leukemia. J. Clin. Oncol. 2013, 31, 676–683. [Google Scholar] [CrossRef] [Green Version]
- Diwaker, D.; Mishra, K.P.; Ganju, L.; Singh, S.B. Protein Disulfide Isomerase Mediates Dengue Virus Entry in Association with Lipid Rafts. Viral Immunol. 2015, 28, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Limjindaporn, T.; Wongwiwat, W.; Noisakran, S.; Srisawat, C.; Netsawang, J.; Puttikhunt, C.; Kasinrerk, W.; Avirutnan, P.; Thiemmeca, S.; Sriburi, R.; et al. Interaction of dengue virus envelope protein with endoplasmic reticulum-resident chaperones facilitates dengue virus production. Biochem. Biophys. Res. Commun. 2009, 379, 196–200. [Google Scholar] [CrossRef]
- Choy, M.M.; Zhang, S.L.; Costa, V.V.; Tan, H.C.; Horrevorts, S.; Ooi, E.E. Proteasome Inhibition Suppresses Dengue Virus Egress in Antibody Dependent Infection. PLoS Negl. Trop. Dis. 2015, 9, e0004058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teissier, E.; Zandomeneghi, G.; Loquet, A.; Lavillette, D.; Lavergne, J.-P.; Montserret, R.; Cosset, F.-L.; Böckmann, A.; Meier, B.H.; Penin, F.; et al. Mechanism of Inhibition of Enveloped Virus Membrane Fusion by the Antiviral Drug Arbidol. PLoS ONE 2011, 6, e15874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germi, R.; Crance, J.-M.; Garin, D.; Guimet, J.; Lortat-Jacob, H.; Ruigrok, R.W.; Zarski, J.-P.; Drouet, E. Heparan Sulfate-Mediated Binding of Infectious Dengue Virus Type 2 and Yellow Fever Virus. Virology 2002, 292, 162–168. [Google Scholar] [CrossRef]
- Nelson, J.; McFerran, N.V.; Pivato, G.; Chambers, E.; Doherty, C.; Steele, D.; Timson, D.J. The 67 kDa laminin receptor: Structure, function and role in disease. Biosci. Rep. 2008, 28, 33–48. [Google Scholar] [CrossRef] [Green Version]
- Tassaneetrithep, B.; Burgess, T.H.; Granelli-Piperno, A.; Trumpfheller, C.; Finke, J.; Sun, W.; Eller, M.A.; Pattanapanyasat, K.; Sarasombath, S.; Birx, D.L.; et al. DC-SIGN (CD209) Mediates Dengue Virus Infection of Human Dendritic Cells. J. Exp. Med. 2003, 197, 823–829. [Google Scholar] [CrossRef] [Green Version]
- Talarico, L.B.; Pujol, C.A.; Zibetti, R.G.M.; Faría, P.C.S.; Noseda, M.D.; Duarte, M.E.R.; Damonte, E.B. The antiviral activity of sulfated polysaccharides against dengue virus is dependent on virus serotype and host cell. Antivir. Res. 2005, 66, 103–110. [Google Scholar] [CrossRef]
- Goo, L.; VanBlargan, L.A.; Dowd, K.A.; Diamond, M.S.; Pierson, T.C. A single mutation in the envelope protein modulates flavivirus antigenicity, stability, and pathogenesis. PLoS Pathog. 2017, 13, e1006178. [Google Scholar] [CrossRef] [Green Version]
- Hishiki, T.; Kato, F.; Tajima, S.; Toume, K.; Umezaki, M.; Takasaki, T.; Miura, T. Hirsutine, an Indole Alkaloid of Uncaria rhynchophylla, Inhibits Late Step in Dengue Virus Lifecycle. Front. Microbiol. 2017, 8, 1674. [Google Scholar] [CrossRef] [PubMed]
- Hitakarun, A.; Khongwichit, S.; Wikan, N.; Roytrakul, S.; Yoksan, S.; Rajakam, S.; Davidson, A.D.; Smith, D.R. Evaluation of the antiviral activity of orlistat (tetrahydrolipstatin) against dengue virus, Japanese encephalitis virus, Zika virus and chikungunya virus. Sci. Rep. 2020, 10, 1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loaiza-Cano, V.; Monsalve-Escudero, L.M.; Quintero-Gil, C.; Pastrana, M.; Andres, P.M.S.; Galeano, E.; Wildeman, Z.; Martinez-Gutierrez, M. In Vitro and In Silico Anti-Arboviral Activities of Dihalogenated Phenolic Derivates of L-Tyrosine. Molecules 2021, 26, 3430. [Google Scholar] [CrossRef]
- Terstappen, G.C.; Reggiani, A. In silico research in drug discovery. Trends Pharm. Sci. 2001, 22, 23–26. [Google Scholar] [CrossRef]
- Velásquez, M.; Drosos, J.; Gueto, C.; Márquez, J.; Vivas-Reyes, R. Autodock-PM6 method to choose the better pose in molecular docking studies. Rev. Colomb. Quím. 2013, 42, 101–124. [Google Scholar]
- Murgueitio, M.S.; Bermudez, M.; Mortier, J.; Wolber, G. In silico virtual screening approaches for anti-viral drug discovery. Drug Discov. Today Technol. 2012, 9, e219–e225. [Google Scholar] [CrossRef]
- Ekins, S.; Mestres, J.; Testa, B. In silico pharmacology for drug discovery: Applications to targets and beyond. Br. J. Pharm. 2007, 152, 21–37. [Google Scholar] [CrossRef] [Green Version]
- Scheiner, S.; Kar, T.; Pattanayak, J. Comparison of Various Types of Hydrogen Bonds Involving Aromatic Amino Acids. J. Am. Chem. Soc. 2002, 124, 13257–13264. [Google Scholar] [CrossRef]
- Quintero-Gil, C.; Parra-Suescún, J.; Lopez-Herrera, A.; Orduz, S. In-silico design and molecular docking evaluation of peptides derivatives from bacteriocins and porcine beta defensin-2 as inhibitors of Hepatitis E virus capsid protein. Virusdisease 2017, 28, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Domingo, C.; Niedrig, M.; Teichmann, A.; Kaiser, M.; Rumer, L.; Jarman, R.G.; Donoso-Mantke, O. 2nd International External Quality Control Assessment for the Molecular Diagnosis of Dengue Infections. PLoS Negl. Trop. Dis. 2010, 4, e833. [Google Scholar] [CrossRef] [Green Version]
- Cockburn, J.; Sanchez, M.E.N.; Fretes, N.; Urvoas, A.; Staropoli, I.; Kikuti, C.M.; Coffey, L.L.; Seisdedos, F.A.; Bedouelle, H.; Rey, F.A. Mechanism of Dengue Virus Broad Cross-Neutralization by a Monoclonal Antibody. Structure 2012, 20, 303–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavi, A.; Ngan, C.H.; Movshovitz-Attias, D.; Bohnuud, T.; Yueh, C.; Beglov, D.; Schueler-Furman, O.; Kozakov, D. Detection of peptide-binding sites on protein surfaces: The first step toward the modeling and targeting of peptide-mediated interactions. Proteins Struct. Funct. Bioinform. 2013, 81, 2096–2105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berendsen, H.; Van Der Spoel, D.; Van Drunen, R. GROMACS: A message-passing parallel molecular dynamics implementation. Comput. Phys. Commun. 1995, 91, 43–56. [Google Scholar] [CrossRef]
- Lemkul, J. From Proteins to Perturbed Hamiltonians: A Suite of Tutorials for the GROMACS-2018 Molecular Simulation Package [Article v1.0]. Living J. Comput. Mol. Sci. 2019, 1, 5068. [Google Scholar] [CrossRef]
- MacKerell, A.D., Jr.; Brooks, B.; Brooks, C.L., III; Nilsson, L.; Roux, B.; Won, Y.; Karplus, M. CHARMM: The energy function and its parameterization. Encycl. Comput. Chem. 2002, 1, 271. [Google Scholar]
- Vanommeslaeghe, K.; Hatcher, E.; Acharya, C.; Kundu, S.; Zhong, S.; Shim, J.; Darian, E.; Guvench, O.; Lopes, P.; Vorobyov, I.; et al. CHARMM general force field: A force field for drug-like molecules compatible with the CHARMM all-atom additive biological force fields. J. Comput. Chem. 2009, 31, 671–690. [Google Scholar] [CrossRef] [Green Version]
- Selvaraj, C.; Dinesh, D.C.; Panwar, U.; Abhirami, R.; Boura, E.; Singh, S.K. Structure-based virtual screening and molecular dynamics simulation of SARS-CoV-2 Guanine-N7 methyltransferase (nsp14) for identifying antiviral inhibitors against COVID-19. J. Biomol. Struct. Dyn. 2020, 38, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.; Wang, S.M. Classification of VUS and unclassified variants in BRCA1 BRCT repeats by molecular dynamics simulation. Comput. Struct. Biotechnol. J. 2020, 18, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, A.C.; Sahi, S. Molecular modeling and molecular dynamics simulation-based structural analysis of GPR3. Netw. Model. Anal. Health Inform. Bioinform. 2017, 6, 9. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aligned Sequence | |||||||
|---|---|---|---|---|---|---|---|
| Position | 112 | 120 | 124 | 126 | 454 | 478 | |
| Reference sequence | AB609589 DENV-2 New Guinea C | G | T | N | E | T | S |
| Target sequence | DENV-2/NG (Sample 1) | S | T | I | K | A | S |
| Reference sequence | NC001474 DENV-2 16681 | G | R | N | E | T | T |
| Target sequence | DENV-2/16681 (Sample 2) | G | R | N | E | T | T |
| Ligand | Serotype | Free Binding Energy (Kcal/mol) | Hydrogen Bonds | Residues Forming H Bonds | Distance between H+ Bonds (Å) | Residues Participating in Hydrophobic Interactions |
|---|---|---|---|---|---|---|
| VOAC | DENV-1 | −5.90 ± 0.05 | 2 | Arg350 | 3.22 | Lys334-Thr339-Phe337-Ser338-Arg350-Gly349-Pro371-Glu370-Leu351 |
| DENV-2 | −6.03 ± 0.06 | 1 | Phe337 (×2) | 3.21−3.14 | Met301 (×2)-Lys334 (×3)-Ile335-Phe337-Glu338-Leu (×4)-Arg350(×2)-Gly349 | |
| DENV-3 | −5.60 ± 0.00 | 2 | Phe615 | 2.99−3.10 | Leu629-Lys612-Met579-Arg628-Gly627-Pro614-Ile613-Phe615 | |
| DENV-4 | −6.30 ± 0.05 | N/A | N/A | N/A | Ile616-Val626-Lys625-Val627-Gly628-Ile630-Ser632-Thr634-Pro635-Val614-Glu617 | |
| VOAC-OH | DENV-2 | −5.30 ± 0.10 | 2 | Lys334−Asn355 | 3.20−3.25 | Lys334 (×2)-Met301-Phe337-Ile335-Pro356-Leu351 (×2)-Val354 |
| Rupicoline | DENV-2 | −5.07 ± 0.06 | 1 | Phe337 | 3.07 | Pro356-Pro336(×3)-Val354-Leu351-Lys334-Asn355-Met301 (×2)-Phe337-Glu338 (×2)-Gly381 |
| OXO-VOAC | DENV-2 | −6.17 ± 0.15 | 3 | Asn355−Val354 −Phe337 | 3.08−3.07−3.27 | Met301-Ile335-Pro356-Phe337 (×4)-Lys334-Leu351 (×3)-Arg350 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monsalve-Escudero, L.M.; Loaiza-Cano, V.; Zapata-Cardona, M.I.; Quintero-Gil, D.C.; Hernández-Mira, E.; Pájaro-González, Y.; Oliveros-Díaz, A.F.; Diaz-Castillo, F.; Quiñones, W.; Robledo, S.; et al. The Antiviral and Virucidal Activities of Voacangine and Structural Analogs Extracted from Tabernaemontana cymosa Depend on the Dengue Virus Strain. Plants 2021, 10, 1280. https://doi.org/10.3390/plants10071280
Monsalve-Escudero LM, Loaiza-Cano V, Zapata-Cardona MI, Quintero-Gil DC, Hernández-Mira E, Pájaro-González Y, Oliveros-Díaz AF, Diaz-Castillo F, Quiñones W, Robledo S, et al. The Antiviral and Virucidal Activities of Voacangine and Structural Analogs Extracted from Tabernaemontana cymosa Depend on the Dengue Virus Strain. Plants. 2021; 10(7):1280. https://doi.org/10.3390/plants10071280
Chicago/Turabian StyleMonsalve-Escudero, Laura Milena, Vanessa Loaiza-Cano, Maria Isabel Zapata-Cardona, Diana Carolina Quintero-Gil, Estiven Hernández-Mira, Yina Pájaro-González, Andrés Felipe Oliveros-Díaz, Fredyc Diaz-Castillo, Wistón Quiñones, Sara Robledo, and et al. 2021. "The Antiviral and Virucidal Activities of Voacangine and Structural Analogs Extracted from Tabernaemontana cymosa Depend on the Dengue Virus Strain" Plants 10, no. 7: 1280. https://doi.org/10.3390/plants10071280
APA StyleMonsalve-Escudero, L. M., Loaiza-Cano, V., Zapata-Cardona, M. I., Quintero-Gil, D. C., Hernández-Mira, E., Pájaro-González, Y., Oliveros-Díaz, A. F., Diaz-Castillo, F., Quiñones, W., Robledo, S., & Martinez-Gutierrez, M. (2021). The Antiviral and Virucidal Activities of Voacangine and Structural Analogs Extracted from Tabernaemontana cymosa Depend on the Dengue Virus Strain. Plants, 10(7), 1280. https://doi.org/10.3390/plants10071280