
Recurrent Selection with Sub-Lethal Doses of Mesotrione Reduces Sensitivity in Amaranthus palmeri


Abstract
:1. Introduction
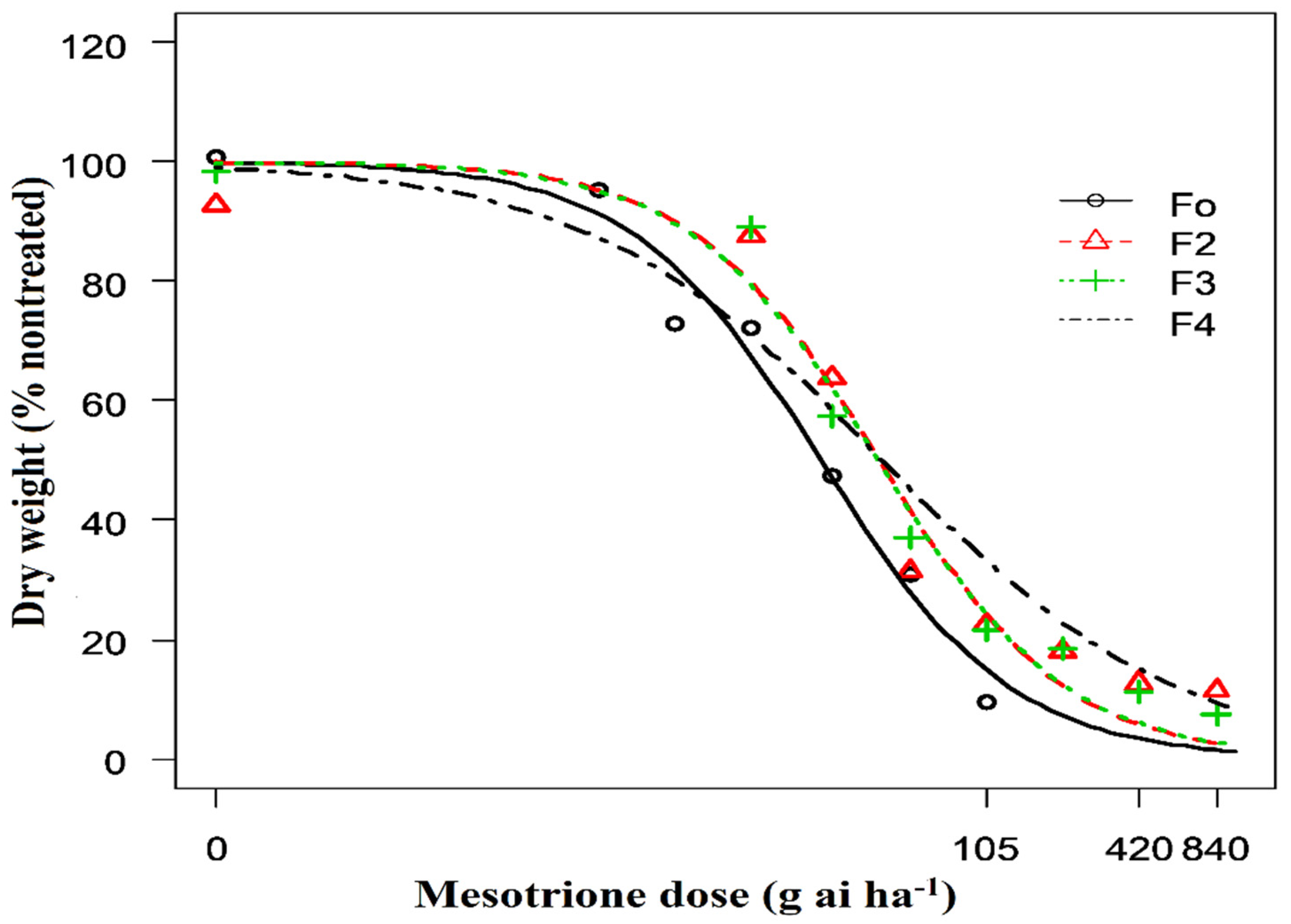
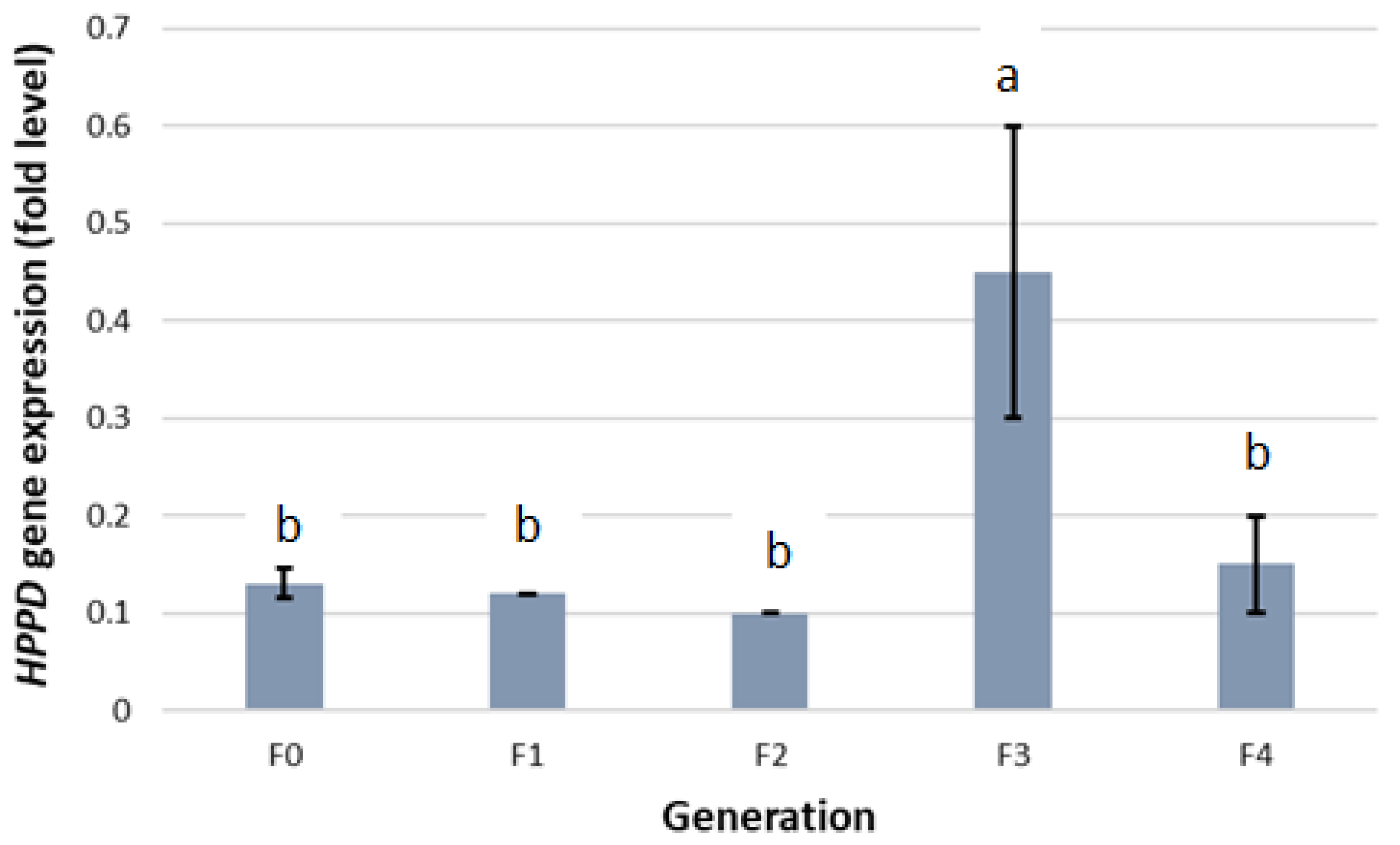
2. Results
2.1. Low-Dose Mesotrione Selection
2.2. HPPD Gene Expression
3. Discussion
4. Materials and Methods
4.1. Parental Populations (F0)
4.2. Generation of F1–F4
4.3. Dose–Response Experiments
4.4. Data Analysis
4.5. RNA Extraction, cDNA Synthesis, and Quantitative PCR
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van Wychen, L. A 2016 Survey of the Most Common and Troublesome Weeds in Broadleaf Crops, Fruits and Vegetables in the United States and Canada. 2016. Available online: http://wssa.net/wp-content/uploads/2016_Weed_Survey_Final.xlsx (accessed on 8 May 2021).
- Korres, N.E.; Norsworthy, J.K. Palmer amaranth (Amaranthus palmeri) demographic and biological characteristics in wide-row soybean. Weed Sci. 2017, 65, 491–503. [Google Scholar] [CrossRef]
- Heap, I. The International Survey of Herbicide Resistant Weeds. 2021. Available online: http://weedscience.com/Home.aspx (accessed on 8 May 2021).
- Ward, S.M.; Webster, T.; Steckel, L.E. Palmer amaranth (Amaranthus palmeri): A review. Weed Technol. 2013, 27, 12–27. [Google Scholar] [CrossRef] [Green Version]
- Hausman, N.E.; Singh, S.; Tranel, P.J.; Riechers, D.E.; Kaundun, S.S.; Polge, N.D.; Thomas, D.A.; Hager, A.G. Resistance to HPPD-inhibiting herbicides in a population of waterhemp (Amaranthus tuberculatus) from Illinois, United States. Pest Manag. Sci. 2011, 67, 258–261. [Google Scholar] [CrossRef]
- Thompson, C.R.; Peterson, D.E.; Lally, N.G. Characterization of HPPD-resistant Palmer amaranth. In Proceedings of the 52th Annual Conference of the Weed Science Society of America, Waikoloa, HI, USA, 6–9 February 2012. [Google Scholar]
- Sutton, P.; Richards, C.; Buren, L.; Glasgow, L. Activity of mesotrione on resistant weeds in maize. Pest Manag. Sci. 2002, 58, 981–984. [Google Scholar] [CrossRef] [PubMed]
- Starkey, C.E.; Norsworthy, J.K.; Schwartz, L.M. Use of HPPD-inhibiting herbicides for control of troublesome weeds in the Midsouthern United States. Adv. Crop Sci. Technol. 2015, 4. [Google Scholar] [CrossRef]
- Gottula, J.; Chapman, K.; Gao, Y.; Gillikin, N.; Beale, J.; Dharmasri, C.; Privalle, L. Agronomic performance and crop composition of genetically engineered cotton tolerant to HPPD inhibiting herbicides. J. Cotton Sci. 2018, 22, 75–85. [Google Scholar]
- Nandula, V.K. Herbicide Resistance Traits in Maize and Soybean: Current Status and Future Outlook. Plants 2019, 8, 337. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, G.; Bartlett, D.W.; Fraser, T.E.M.; Hawkes, T.R.; Holt, D.C.; Townson, J.K.; Wichert, R.A. Mesotrione: A new selective herbicide for use in maize. Pest Manag. Sci. 2001, 57, 120–128. [Google Scholar] [CrossRef]
- Norris, S.R.; Barrette, T.R.; DellaPenna, D. Genetic dissection of carotenoid synthesis in Arabidopsis defines plastoquinone as an essential component of phytoene desaturation. Plant Cell 1995, 7, 2139–2149. [Google Scholar]
- Ruiz-Sola, M.Á.; Rodríguez-Concepción, M. Carotenoid Biosynthesis in Arabidopsis: A Colorful Pathway. Arab. Book 2012, 10, e0158. [Google Scholar] [CrossRef] [Green Version]
- Doyle, P.; Stypa, M. Reduced herbicide rates—A Canadian perspective. Weed Technol. 2004, 18, 1157–1165. [Google Scholar] [CrossRef]
- Neve, P.; Powles, S. Recurrent selection with reduced herbicide rates results in the rapid evolution of herbicide resistance in Lolium rigidum. Theor. Appl. Genet. 2005, 110, 1154–1166. [Google Scholar] [CrossRef] [PubMed]
- Gressel, J. Creeping resistances: The outcome of using marginally effective or reduced rates of herbicide. Brighton Crop Prot. Conf. Weeds 1995, 2, 587. [Google Scholar]
- Gardner, S.N.; Gressel, J.; Mangel, M. A revolving dose strategy to delay the evolution of both quantitative vs major monogene resistances to pesticides and drugs. Int. J. Pest Manag. 1998, 44, 161–180. [Google Scholar] [CrossRef]
- Ma, R.; Kaundun, S.S.; Tranel, P.J.; Riggins, C.W.; McGinness, D.L.; Hager, A.G.; Hawkes, T.; McIndoe, E.; Riechers, D.E. Distinct detoxification mechanisms confer resistance to mesotrione and atrazine in a population of waterhemp. Plant Physiol. 2013, 163, 368–377. [Google Scholar] [CrossRef] [Green Version]
- Nakka, S.; Godar, A.S.; Wani, P.S.; Thompson, C.R.; Peterson, D.E.; Roelofs, J.; Jugulum, M. Physiological and molecular characterization of hydroxyphenylpyruvate dioxygenase (HPPD)-inhibitor resistance in Palmer amaranth (Amaranthus palmeri S. Wats.). Front. Plant Sci. 2017, 8, 555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neve, P.; Powles, S.B. High survival frequencies at low herbicide use rates in populations of Lolium rigidum result in rapid evolution of herbicide resistance. Heredity 2005, 95, 485–492. [Google Scholar] [CrossRef] [Green Version]
- Busi, R.; Neve, P.; Powles, S.B. Evolved polygenic herbicide resistance in Lolium rigidum by low-dose herbicide selection within standing genetic variation. Evol. Appl. 2013, 6, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Busi, R.; Powles, S.B. Evolution of glyphosate resistance in a Lolium rigidum population by glyphosate selection at sub-lethal doses. Heredity 2009, 103, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Renton, M.; Diggle, A.; Manalil, S.; Powles, S. Does cutting herbicide rates threaten the sustainability of weed management in cropping systems? J. Theor. Biol. 2011, 283, 14–27. [Google Scholar] [CrossRef]
- Norsworthy, J.K. Repeated sublethal rates of glyphosate lead to decreased sensitivity in Palmer amaranth. Crop Manag. 2012, 11, 1–6. [Google Scholar] [CrossRef]
- Ashworth, M.B.; Walsh, M.J.; Flower, K.C.; Powles, S.B. Recurrent selection with reduced 2,4-D amine doses results in the rapid evolution of 2,4-D herbicide resistance in wild radish (Raphanus raphanistrum L.). Pest Manag. Sci. 2016, 72, 2091–2098. [Google Scholar] [CrossRef] [PubMed]
- Tehranchian, P.; Norsworthy, J.K.; Powles, S.; Bararpour, M.T.; Bagavathiannan, M.V.; Barber, T.; Robert, C.S. Recurrent suble-thal-dose selection for reduced susceptibility of Palmer amaranth (Amaranthus palmeri) to dicamba. Weed Sci. 2017, 65, 206–212. [Google Scholar] [CrossRef]
- Yu, Q.; Han, H.P.; Cawthray, G.; Wang, S.F.; Powles, S.B. Enhanced rates of herbicide metabolism in low herbicide-dose selected resistant Lolium rigidum. Plant Cell Environ. 2013, 36, 818–827. [Google Scholar] [CrossRef] [PubMed]
- Godar, A.S.; Varanasi, V.K.; Nakka, S.; Prasad, P.V.V.; Thompson, C.R.; Mithila, J. Physiological and molecular mechanisms of differential sensitivity of Palmer amaranth (Amaranthus palmeri) to mesotrione at varying growth temperatures. PLoS ONE 2015, 10, e0126731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busi, R.; Girotto, M.; Powles, S.B. Response to low-dose herbicide selection in self-pollinated Avena fatua. Pest Manag. Sci. 2016, 72, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Jasieniuk, M.; Brûlé-Babel, A.L.; Morrison, I.N. The evolution and genetics of herbicide resistance in weeds. Weed Sci. 1996, 44, 176–193. [Google Scholar] [CrossRef]
- González-Torralva, F.; Norsworthy, J.K.; Piveta, L.B.; Varanasi, V.K.; Barber, T.; Brabham, C. Susceptibility of Arkansas A. palmeri accessions to common herbicide sites of action. Weed Technol. 2021, in press. [Google Scholar]
- Singh, S.; Roma-Burgos, N.; Singh, V.; Alcober, E.A.L.; Salas-Perez, R.; Shivrain, V. Differential response of Arkansas Palmer amaranth (Amaranthus palmeri) to glyphosate and mesotrione. Weed Technol. 2018, 32, 579–585. [Google Scholar] [CrossRef]
- Oliveira, M.C.; Gaines, T.A.; Dayan, F.E.; Patterson, E.; Jhala, A.J.; Knezevic, S.Z. Reversing resistance to tembotrione in an Amaranthus tuberculatus (var. rudis) population from Nebraska, USA with cytochrome P450 inhibitors. Pest Manag. Sci. 2018, 74, 2296–2305. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.C.; Gaines, T.A.; Jhala, A.J.; Knezevic, S.Z. Inheritance of mesotrione resistance in an Amaranthus tuberculatus (var. rudis) population from Nebraska, USA. Front. Plant Sci. 2018, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Norsworthy, J.K.; Oliver, L.R.; Purcell, L.C. Diurnal leaf movement effects on spray interception and glyphosate efficacy. Weed Technol. 1999, 13, 466–470. [Google Scholar] [CrossRef]
- Norsworthy, J.; Ward, S.M.; Shaw, D.R.; Llewellyn, R.S.; Nichols, R.L.; Webster, T.; Bradley, K.W.; Frisvold, G.; Powles, S.B.; Burgos, N.R.; et al. Reducing the risks of herbicide resistance: Best management practices and recommendations. Weed Sci. 2012, 60, 31–62. [Google Scholar] [CrossRef] [Green Version]
- Koger, C.H.; Poston, D.H.; Reddy, K.N. Effect of glyphosate spray coverage on control of pitted morningglory (Ipomoea lacunosa). Weed Technol. 2004, 18, 124–130. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing Computer Program, Version 3.6.1; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: http://www.r-project.org (accessed on 8 May 2021).
- Seefeldt, S.S.; Jensen, J.E.; Fuerst, E.P. Log-logistic analysis of herbicide dose-response relationships. Weed Technol. 1995, 9, 218–227. [Google Scholar] [CrossRef]
- Owczarzy, R.; Tataurov, A.V.; Wu, Y.; Manthey, J.A.; McQuisten, K.A.; Almabrazi, H.G.; Pedersen, K.F.; Lin, Y.; Garretson, J.; McEntaggart, N.O.; et al. IDT SciTools: A suite for analysis and design of nucleic acid oligomers. Nucleic Acids Res. 2008, 36, W163–W169. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Mesotrione Dose (x) (1× = 105 g ai ha−1) | # of Seedlings Treated | Survivors (%) | |
|---|---|---|---|
| F0 | 0.5 | 200 | 2.5 |
| 1.0 | 200 | 0 | |
| F1 | 0.75 | 275 | 2.5 |
| 2.0 | 275 | 1.4 | |
| F2 | 1.0 | 250 | 56 |
| 1.5 | 200 | 37 | |
| F3 | 1.5 | 115 | 31 |
| 2.0 | 100 | 10 |
| Generation | GR50 a | Sensitivity Index b |
|---|---|---|
| F0 | 23.57 (2.00) | - |
| F2 | 39.96 (3.33) | 1.5 (F2/F0) |
| F3 | 39.60 (3.33) | 1.5 (F3/F0) |
| F4 | 41.22 (5.07) | 1.7 (F4/F0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Norsworthy, J.K.; Varanasi, V.K.; Bagavathiannan, M.; Brabham, C. Recurrent Selection with Sub-Lethal Doses of Mesotrione Reduces Sensitivity in Amaranthus palmeri. Plants 2021, 10, 1293. https://doi.org/10.3390/plants10071293
Norsworthy JK, Varanasi VK, Bagavathiannan M, Brabham C. Recurrent Selection with Sub-Lethal Doses of Mesotrione Reduces Sensitivity in Amaranthus palmeri. Plants. 2021; 10(7):1293. https://doi.org/10.3390/plants10071293
Chicago/Turabian StyleNorsworthy, Jason K., Vijay K. Varanasi, Muthukumar Bagavathiannan, and Chad Brabham. 2021. "Recurrent Selection with Sub-Lethal Doses of Mesotrione Reduces Sensitivity in Amaranthus palmeri" Plants 10, no. 7: 1293. https://doi.org/10.3390/plants10071293