1. Introduction
Silicon (Si) is the second most abundant element after oxygen in the Earth’s crust, comprising approximately 29% (28.8% wt) of the Earth’s crust [
1,
2,
3]. Although Si is considered a quasi-essential element rather than a plant nutrient, it is increasingly being applied in agricultural systems worldwide [
2,
3,
4,
5]. Silicon content in the soil ranges from almost 1 to 45% depending on soil types [
6]. The potential of Si in improving crop yield has been demonstrated in many studies, especially under biotic (pathogens attack) and abiotic (e.g., salinity, drought, high temperature, heavy metals toxicity) stress conditions [
7,
8]. Silicon is known for its role in alleviating negative effects of stress on many plant species. Monocotyledons in general and
Poaceae species such as maize
(Zea mays L.) and wheat (
Triticum aestivum L.) are clearly favored due to an enhanced supply of Si [
3,
6,
9]. It has been reported that some
Poaceae species could accumulate Si to a level above 1% of total shoot biomass [
7].
The cultivation and harvest of Si-accumulating crops is responsible for a constant depletion of the Si reservoir in soils [
10,
11] which may decrease Si bioavailability [
12]. The enhanced Si removal by Si-accumulating crops, therefore, disrupts the recycling of Si by plants back into the soil [
13]. Liang et al. [
14] reported that Si content in Oxisols in the tropical zone can be less than 1% due to intense weathering processes. Highly weathered tropical and subtropical soils under continuous cropping systems are generally low in available Si content due to heavy desilication of primary silicate minerals, release and leaching of basic cations with decreased base saturation, and crop removal [
3,
7,
15,
16]. Therefore, the decrease in Si availability in tropical soils might have significant impacts on cropping systems if not properly managed.
A further problem in weathered and acidic soils is related to the occurrence of high levels of exchangeable Al
3+. Soil acidity and related Al
3+ toxicity have long been recognized as significant and alarming constraints in cereal production under tropical conditions. Nonetheless, Ca and Mg silicate could be a viable alternative to increase Si availability and neutralize Al toxicity as an alternative to conventional liming. Silicate application could reduce the time associated with liming reactions in the soil profile due to their higher solubility (6–7 times more soluble than carbonates) and alkalinity compared with limestone [
17,
18]. However, most studies related to Si-amendment sources application focused on soil chemical shifts instead of plant growth promotion, and nutrient uptake and imbalance. The application of various forms of Si (i.e., slag materials, silica powder, silicates, among others) could improve plant growth and development with greater nutrient uptake in different crops [
16,
19,
20,
21]. Haddad et al. [
22] indicated a beneficial effect of Si treatment in alleviating damage associated with nitrogen (N) deficiency in rapeseed (
Brassica napus L.). These authors showed that Si supply modulates the root expression of a large panel of genes [
23], promotes a stronger N uptake associated with the induction of root nitrate transporters, and delays leaf senescence in plants cultivated under N deficiency [
22]. Nonetheless, Si application has been reported to increase N availability in the soil by modifying physicochemical (e.g., soil exchange capacity) [
24,
25] and biological properties of soil (e.g., increasing the biomass of microbial N fixers) [
26]. Singh et al. [
27] reported that Si application in rice (
Oryza sativa L.) enhances N availability in soil and leads to an increase in N uptake by plants. Laîné et al. [
28] concluded that Si application (12 kg Si ha
−1 as silicic acid) may have increased N availability in the soil, improved N uptake, and led to an accumulation of N in rapeseed plants cultivated with high N rates (160 kg ha
−1). In this regard, Si application could affect N availability to plants and enhance nutrient use efficiency (also known as agronomic efficiency), expressed as grain produced per unit of N applied. Thereby, Si fertilization might improve N fertilization management while reducing the need for mineral N fertilization in agricultural systems [
19].
Nitrogen is considered one of the most limiting nutrients for crop growth and yield [
29]. However, excessive N supply could directly contribute to soil acidification, ammonia (NH
3) and nitrogen oxide (N
2O, NO
2 and NO) emissions, and N leaching with extended consequences on global warming [
30]. The application of N to maize and wheat using a side-dressing approach in the Brazilian Savannah has led to an increase of about 15% in the farmer’s total operational profit [
31,
32]. Over the last decades, several management practices have been developed to enable farmers to reduce the application of N-based fertilizers in cereal crops in order to limit potential negative environmental impacts and cost resulting from the heavy application of chemical N fertilizer [
33]. Integrating N management practices could contribute significantly to improved cereal production under the savannah conditions, particularly in a maize-wheat rotation, which has high N nutritional demands [
34,
35]. Moreover, developing management practices that enhance agronomic efficiency leading to a greater grain yield is required to foment sustainable agricultural practices in the tropical agriculture.
Therefore, we propose a novel approach to investigate the effect of Si-amendment application, combined with N levels in a maize-wheat cropping system under tropical conditions. This research could provide new clues on how Si application could be included as an important strategy for improving the sustainability of cereal production with an improved N fertilization management. We hypothesized that Si application would improve the agronomic efficiency and N accumulation in maize and wheat crops. The increased agronomic efficiency and N accumulation may provide increased shoot and root development which may lead to greater grain yield when compared to plots that did not receive Si application. Therefore, the objective of this study was to investigate the combined effects of Si-amendment source (Ca and Mg silicate) and N levels (control, low, medium and high levels) application on: (1) leaf chlorophyll index (LCI), inorganic (N-NH4+ and N-NO3−) and total N and Si content in maize and wheat plants; (2) plant biomass, grain yield and agronomic efficiency of individual treatments.
3. Discussion
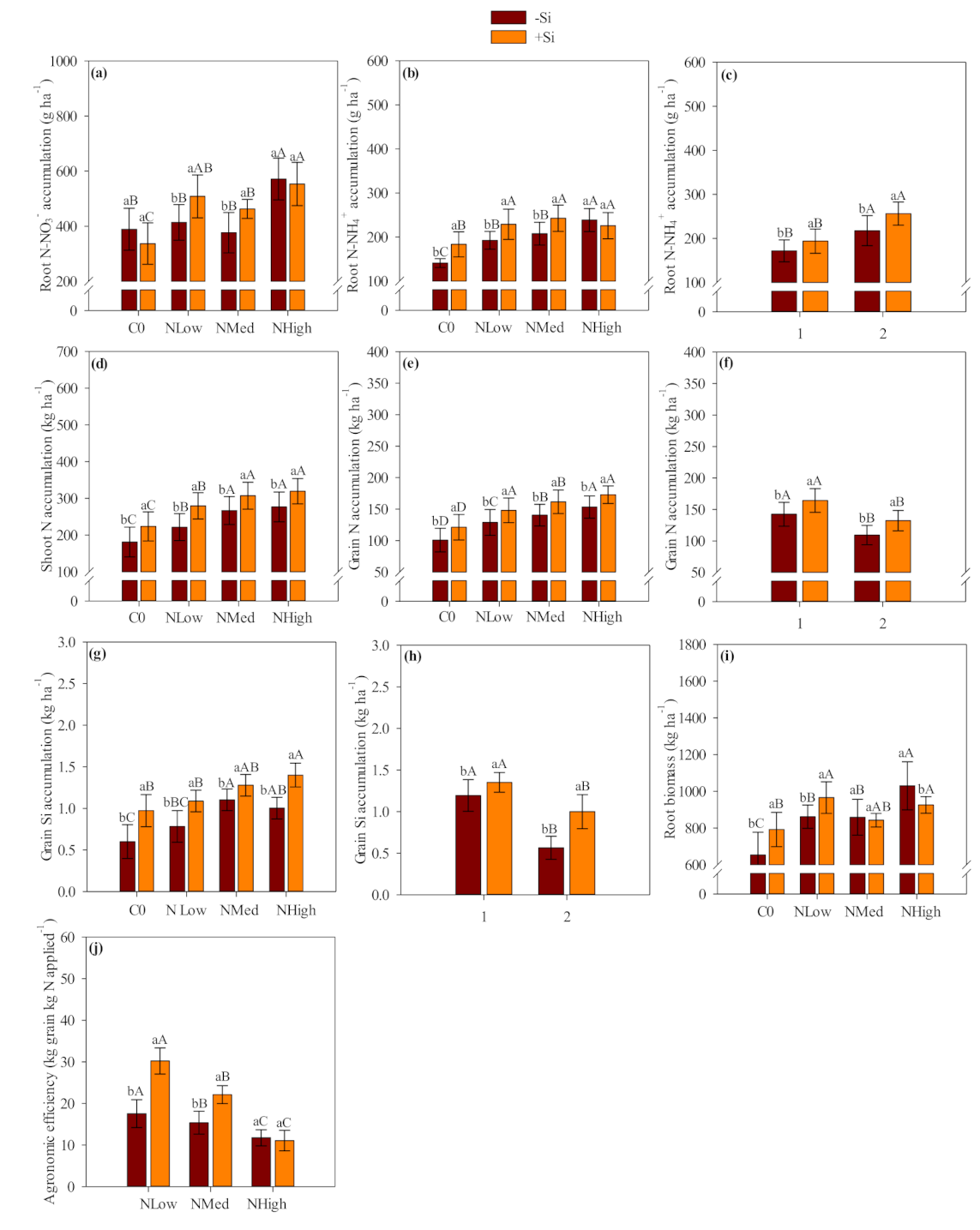
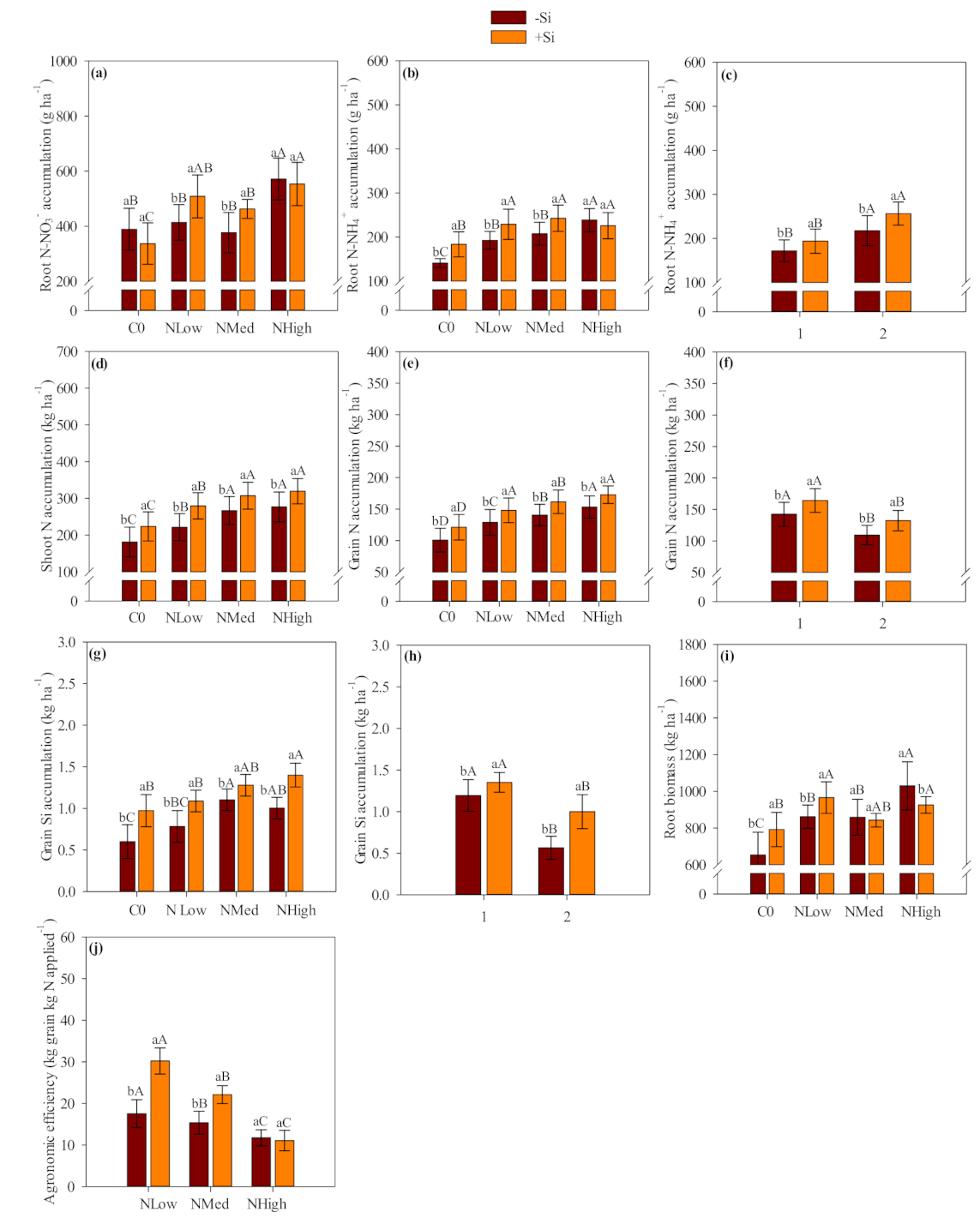
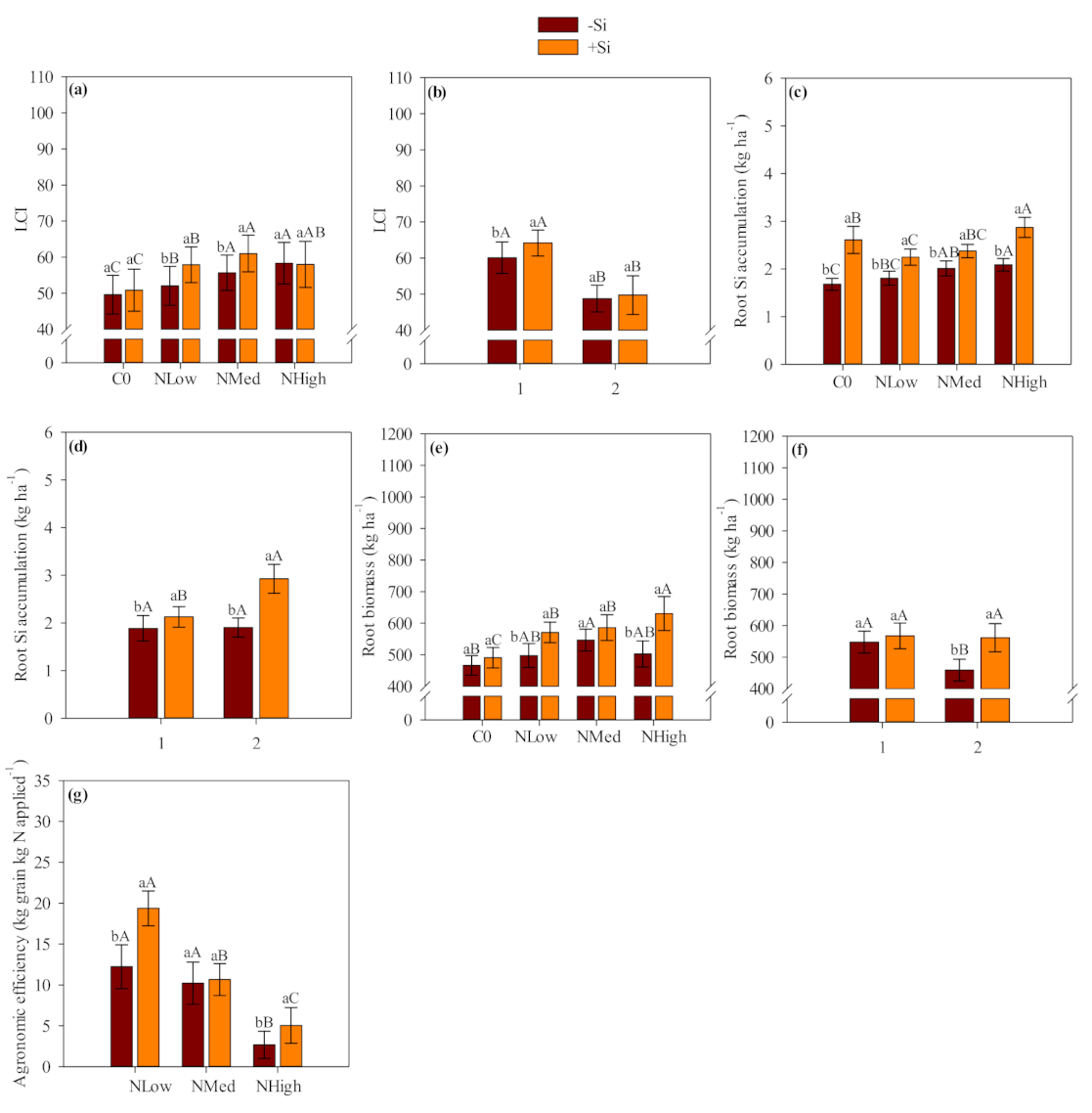
Our results showed that Ca and Mg silicate was an effective source of Si which increased Si accumulation in the shoots and roots of field-grown maize and wheat crops. Most of the absorbed Si was accumulated in the shoot tissue, where it is deposited within the leaf epidermis through plant transpiration flux. The Si inside plant epidermis was condensed into a polymerized silica gel (SiO
2 nH
2O) known as a phytolith that is immobile and developed a protective structural layer in the plant cell walls [
36,
37,
38]. Interestingly, we verified that grain Si accumulation in maize and wheat also increased with silicate application. Some plant species can take up and translocate large quantities of Si in the aboveground tissue due to specific Si transporters [
39]. For example, Ma et al. [
40] reported that the transport of Si from soil to the rice panicles is mediated by different transporters. According to these authors, Lsi1, belonging to a NIP group of the aquaporin family, is responsible for the uptake of Si from soil into the root cells in both dicots and monocots although its expression patterns and cellular localization differ with plant species. The subsequent transport of Si out of the root cells towards the stele is medicated by an active efflux transporter, Lsi2 [
40]. Silicon in the xylem sap is presented in the form of monosilicic acid and is unloaded by Lsi6, a homolog of Lsi1 in rice [
40]. Lsi6 is also involved in the inter-vascular transfer of Si at the node, which is necessary for preferential Si distribution to the panicles [
40].
Silicon-applied treatments showed a total Si accumulation (shoot, root and grain) of 43 kg Si ha
−1 for maize and 36 kg Si ha
−1 for the wheat crop. Some previous studies reported that sugarcane (
Saccharum officinarum L.) has the potential to take up the largest amount of silicon (300–700 kg of Si ha
−1), followed by rice (150–300 kg of Si ha
−1), and wheat (30–150 kg of Si ha
−1) [
41]. On an average, plants can absorb anywhere from 30 to 300 kg Si ha
−1 [
38,
42]. Such values of absorbed silicon cannot be fully explained by passive absorption because the upper 0.20 m soil layer contains only an average of 0.1–1.6 kg Si ha
−1 as monosilicic acid [
41]. The above high Si accumulation observed in maize plants indicated that this crop could be considered one of the most efficient Si accumulators in
Poaceae, after sugarcane and rice.
Leaf chlorophyll index (LCI) was benefited by Si application in maize. The Si application also benefited LCI in wheat, however the positive response was more evident in the first year than in the second wheat crop season when coupled with low and medium N levels. Silicon fertilization can benefit the foliar architecture of plants by improving erectness of leaves which leads to greater light interception, reducing self-shading and lodging, postponing senescence and improving photosynthesis [
39,
43]. Additionally, Si application had greater influence on the agronomic efficiency of the N fertilizer applied to the maize and wheat crops. Specifically, under low and medium N levels, agronomic efficiency was increased by 73 and 44% with Si application in maize, respectively. Similarly, under low and high N levels, agronomic efficiency was increased by 58 and 85% with Si application in wheat. Silicon application can enhance primary metabolism by decreasing the transpiration rate [
44] and improving photosynthesis [
45,
46] and nutrient uptake [
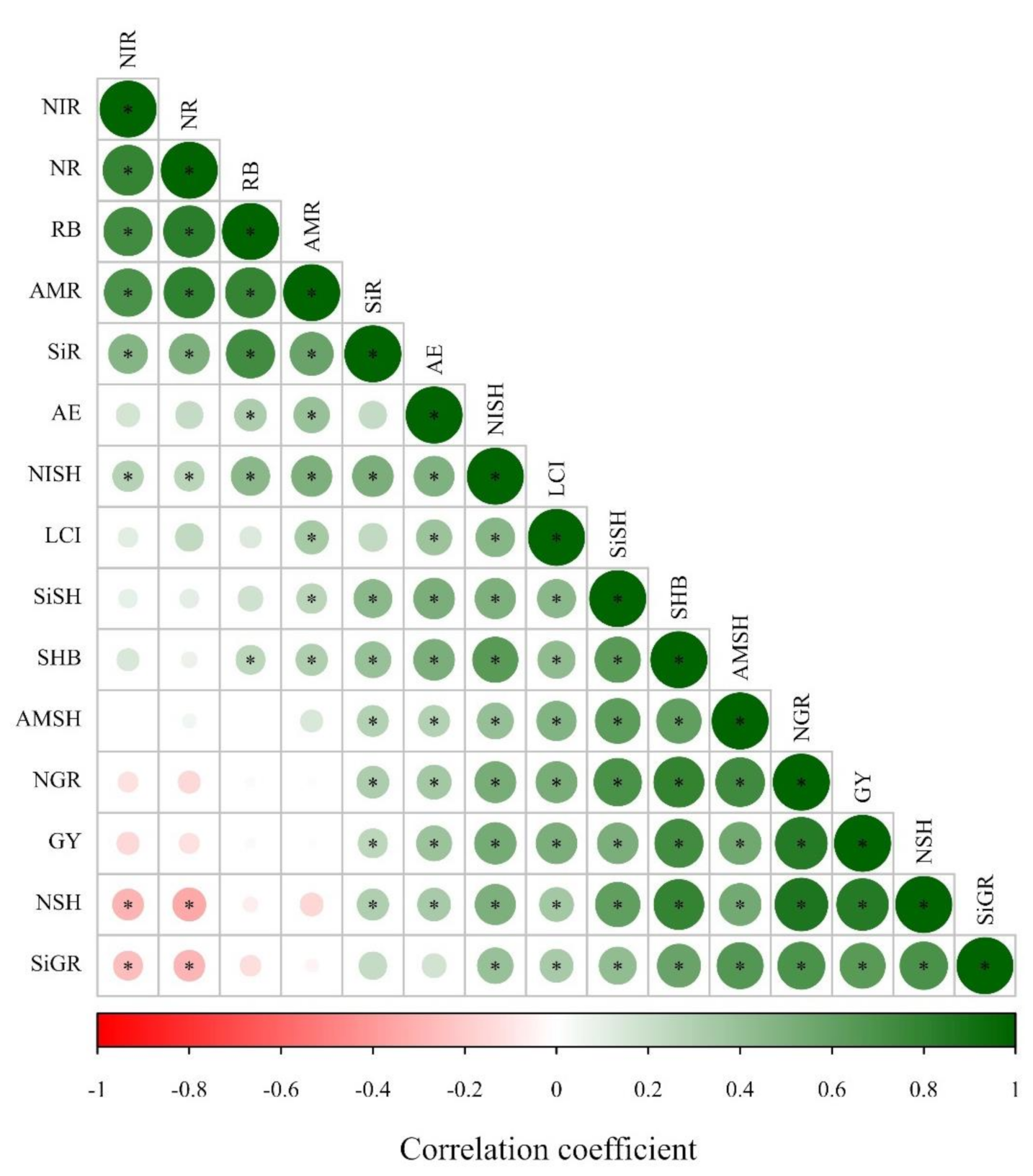
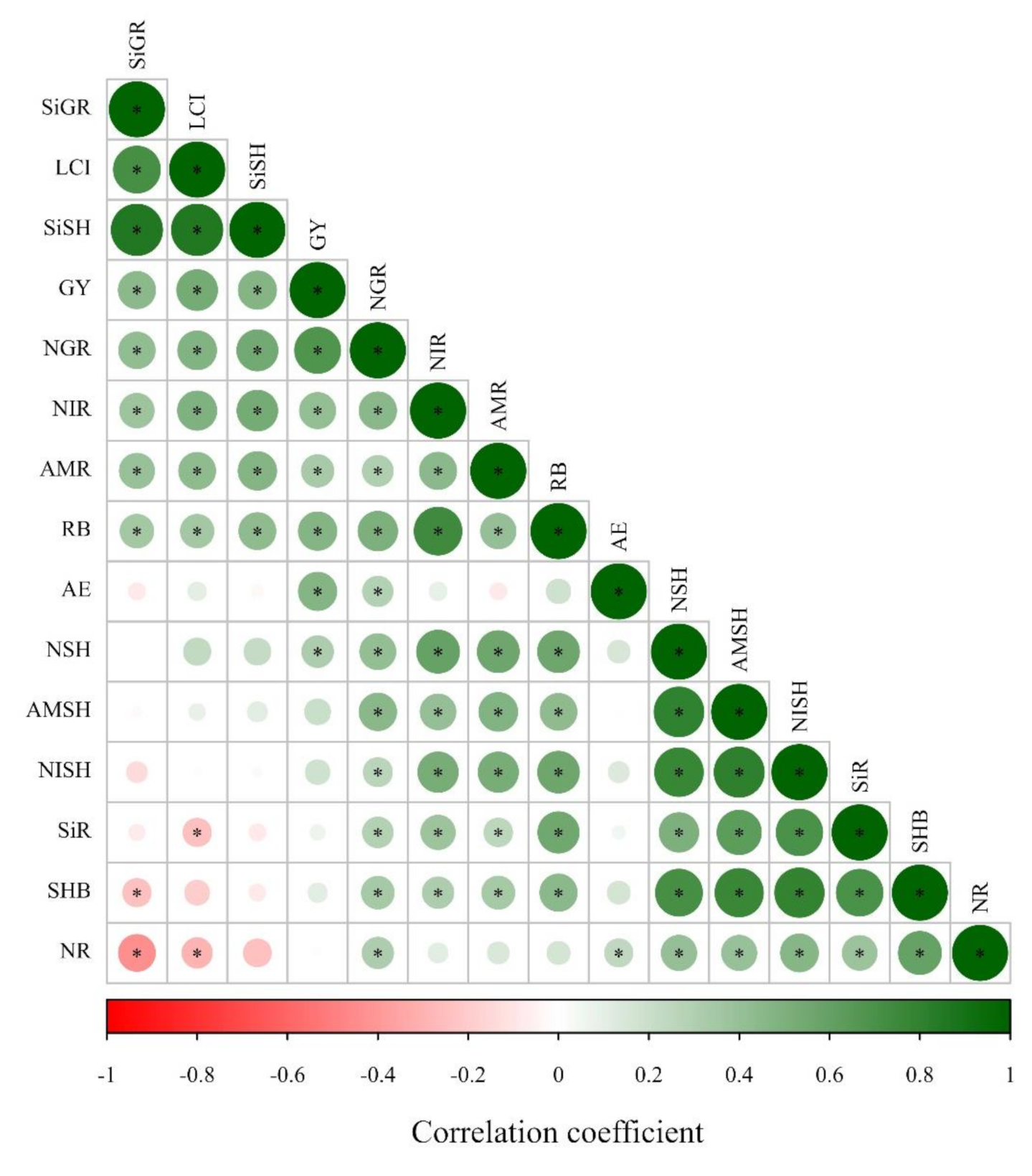
47]. Thereby, the greater Si uptake and LCI verified in our study together with enhanced agronomic efficiency of N-applied levels due to silicate application positively influenced the inorganic N accumulation in maize and wheat root and shoot, and grain N accumulation, leading to a greater shoot biomass and grain yield. The positive Pearson’s correlation along with Si accumulation in maize and wheat plants, LCI, and N accumulation in plant and biomass components support this hypothesis (
Figure 2 and
Figure 4).
The results suggested that Si application increases N agronomic efficiency and grain yield, so presumably less N would need to be applied when used in combination with Si. Based on the verified maize and wheat grain yield, linear interpolation was accessed (the equations were presented as
Appendix C). It would be possible to reduce N application from 180–185 to 100 kg N ha
−1 (for maize) and from 195–200 to 100 kg N ha
−1 (for wheat) when silicate was applied, with similar grain yield (maize: −Si 180–185N = 10223–10264 kg ha
−1 vs. +Si 100N = 10252 kg ha
−1; wheat: −Si 195–200N = 4385–4405 kg ha
−1 vs. +Si 100N = 4392 kg ha
−1) (
Appendix C). Positive responses to Si application, mainly under abiotic and biotic environments, were reported elsewhere for different plant species [
48,
49,
50,
51,
52]. Here, we have focused on the positive effects of Si application on N agronomic efficiency under harsh field growing conditions, such as drought and high temperatures which are common under tropical agriculture conditions. In our study, although total rainfall was relatively well distributed during maize growing seasons, high temperatures were observed during the field trial (maximum and minimum temperature above 35 °C and 20 °C in most of the cases, respectively) (
Figure 5). Regarding wheat crop, supplementary irrigation was performed, although total rainfall was poor and not well distributed during the wheat growing seasons (
Figure 5). In addition, high temperatures were observed during the field trial (maximum and minimum temperature above 30 °C and 15 °C in most of the cases, respectively). Thus, plant growth response to Si application under tropical conditions would be more noticeable, as verified in our study, especially for Si accumulator crops such as maize and wheat, as we discussed above.
Although verifying a positive residual effect of Si application, our study indicated that the influence of residual Si-amendment source was less evident in each successive crop season. For example, shoot and grain Si accumulation was greater in the first than second maize cropping season. In addition, the shoot and grain N accumulation, and the shoot biomass and grain yield followed the same trend as observed for the shoot and grain Si accumulation. Our study observed the same behavior in wheat crop, with a reduced shoot and grain Si accumulation followed by reduced LCI, grain N accumulation, shoot biomass and grain yield. We could expect this behavior once Si uptakes by maize-wheat crops are higher and coupled with no other nutrient application would reduce Si availability along successive crop seasons. Thus, adequate Si replenishment along continuous successive cropping seasons are required to avoid this element depletion and impaired cereal growth and yield.


 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}