
Genetic Similarity of Avena sativa L. Varieties as an Example of a Narrow Genetic Pool of Contemporary Cereal Species



Abstract

1. Introduction
2. Materials and Methods
2.1. Plant Material and DNA Extraction
2.2. Molecular Analyses Using ISSR and SCoT Primer
2.3. Data Analysis
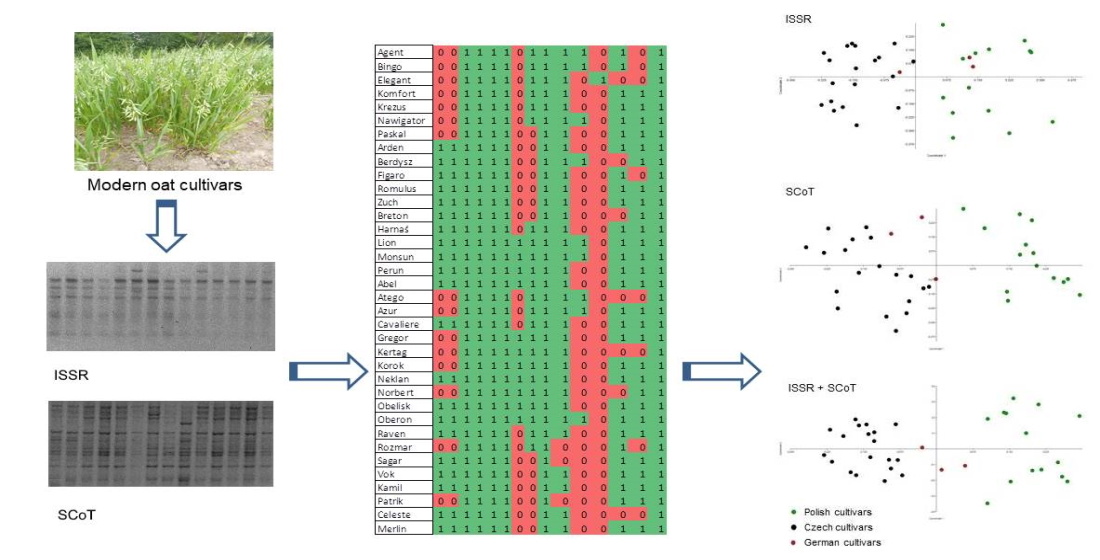
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Warburton, M.L.; Reif, J.C.; Frisch, M.; Bohn, M.; Bedoya, C.; Xia, X.C.; Crossa, J.; Franco, J.; Hoisington, D.; Pixley, K.; et al. Genetic Diversity in CIMMYT Nontemperate Maize Germplasm: Landraces, Open Pollinated Varieties, and Inbred Lines. Crop Sci. 2008, 48, 617–624. [Google Scholar] [CrossRef]
- Roussel, V.; Koenig, J.; Beckert, M.; Balfourier, F. Molecular diversity in French bread wheat accessions related to temporal trends and breeding programmes. Theor. Appl. Genet. 2004, 108, 920–930. [Google Scholar] [CrossRef]
- Reif, J.C.; Zhang, P.; Dreisigacker, S.; Warburton, M.L.; Van Ginkel, M.; Hoisington, D.; Bohn, M.; Melchinger, A.E. Wheat genetic diversity trends during domestication and breeding. Theor. Appl. Genet. 2005, 110, 859–864. [Google Scholar] [CrossRef]
- Christiansen, M.J.; Andersen, S.B.; Ortiz, R. Diversity changes in an intensively bred wheat germplasm during the 20th century. Mol. Breed. 2002, 9, 1–11. [Google Scholar] [CrossRef]
- Donini, P.; Law, J.R.; Koebner, R.M.D.; Reeves, J.C.; Cooke, R.J. Temporal trends in the diversity of UK wheat. Theor. Appl. Genet. 2000, 100, 912–917. [Google Scholar] [CrossRef]
- Koebner, R.M.D.; Donini, P.; Reeves, J.C.; Cooke, R.J.; Law, J.R. Temporal flux in the morphological and molecular diversity of UK barley. Theor. Appl. Genet. 2003, 106, 550–558. [Google Scholar] [CrossRef]
- Montilla-Bascón, G.; Sánchez-Martín, J.; Rispail, N.; Rubiales, D.; Mur, L.; Langdon, T.; Griffiths, I.; Howarth, C.; Prats, E. Genetic Diversity and Population Structure Among Oat Cultivars and Landraces. Plant Mol. Biol. Report. 2013, 31, 1305–1314. [Google Scholar] [CrossRef]
- Frankel, O.; Bennett, E. Genetic resources in plants: Their exploration and conservation. In Genetic Resources in Plants: Their Exploration and Conservation; Blackwell Scientific: Oxford, UK, 1970; p. 575. ISBN 0632057300. [Google Scholar]
- Boczkowska, M.; Onyśk, A. Unused genetic resources: A case study of Polish common oat germplasm. Ann. Appl. Biol. 2016, 169, 155–165. [Google Scholar] [CrossRef]
- Fu, Y.B.; Peterson, G.W.; Williams, D.; Richards, K.W.; Fetch, J.M. Patterns of AFLP variation in a core subset of cultivated hexaploid oat germplasm. Theor. Appl. Genet. 2005, 111, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Semagn, K.; Bjornstad, A.; Ndjiondjop, M.N. An overview of molecular marker methods for plants. African J. Biotechnol. 2006, 5, 2540–2568. [Google Scholar]
- Agarwal, M.; Shrivastava, N.; Padh, H. Advances in molecular marker techniques and their applications in plant sciences. Plant Cell Rep. 2008, 27, 617–631. [Google Scholar] [CrossRef]
- Okoń, S.; Surmacz-Magdziak, A.; Paczos-Grzeda, E.; Oko, S.; Surmacz-Magdziak, A.; Paczos-Grzda, E. Genetic diversity among cultivated and wild chamomile germplasm based on ISSR analysis. Acta Sci. Pol. Hortorum Cultus 2013, 12, 43–50. [Google Scholar]
- Kiełtyka-Dadasiewicz, A.; Okoń, S.; Ociepa, T.; Król, B. Morphological and genetic diversity among peppermint (Mentha × piperita L.) cultivars. Acta Sci. Pol. Hortorum Cultus 2017, 16, 151–161. [Google Scholar] [CrossRef]
- Kołodziej, B.; Okoń, S.; Nucia, A.; Ociepa, T.; Luchowska, K.; Sugier, D.; Gevrenova, R.; Henry, M. Morphological, chemical, and genetic diversity of Gypsophila L. (Caryophyllaceae) species and their potential use in the pharmaceutical industry. Turk. J. Botany 2018, 42, 257–270. [Google Scholar]
- Zietkiewicz, E.; Rafalski, A.; Labuda, D. Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification. Genomics 1994, 20, 176–183. [Google Scholar] [CrossRef]
- Reddy, M.P.; Sarla, N.; Siddiq, E.A. Inter simple sequence repeat (ISSR) polymorphism and its application in plant breeding. Euphytica 2002, 128, 9–17. [Google Scholar] [CrossRef]
- Sawant, S.V.; Singh, P.K.; Gupta, S.K.; Madnala, R.; Tuli, R. Conserved nucleotide sequences in highly expressed genes in plants. J. Genet. 1999, 78, 123–131. [Google Scholar] [CrossRef]
- Collard, B.C.Y.; Mackill, D.J. Start Codon Targeted (SCoT) Polymorphism: A Simple, Novel DNA Marker Technique for Generating Gene-Targeted Markers in Plants. Plant Mol. Biol. Report. 2009, 27, 86–93. [Google Scholar]
- Biosci, I.J.; Iqbal, A.; Ahmad, I.; Ahmad, H.; Nadeem, M.S.; Nisar, M.; Riaz, H. An efficient DNA extraction protocol for medicinal plants. Int. J. Biosci. 2013, 3, 30–35. [Google Scholar]
- Anderson, J.A.; Churchill, G.A.; Autrique, J.E.; Tanksley, S.D.; Sorrells, M.E. Optimizing parental selection for genetic linkage maps. Genome 1993, 36, 181–186. [Google Scholar] [CrossRef]
- Belaj, A.; Trujillo, I.; De La Rosa, R.; Rallo, L.; Giménez, M.J. Polymorphism and Discrimination Capacity of Randomly Amplified Polymorphic Markers in an Olive Germplasm Bank. J. Am. Soc. Hort. Sci. 2001, 126, 64–7164. [Google Scholar] [CrossRef]
- Prevost, A.; Wilkinson, M.J. A new system of comparing PCR primers applied to ISSR fingerprinting of potato cultivars. Theor. Appl. Genet. 1999, 98, 107–112. [Google Scholar] [CrossRef]
- Dice, L.R. Measures of the Amount of Ecologic Association Between Species. Ecology 1945, 26, 297–302. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Struss, D.; Plieske, J. The use of microsatellite markers for detection of genetic diversity in barley populations. Theor. Appl. Genet. 1998, 97, 308–315. [Google Scholar] [CrossRef]
- Chen, Z.W.; Lu, R.J.; Zou, L.; Du, Z.Z.; Gao, R.H.; He, T.; Huang, J.H. Genetic diversity analysis of barley landraces and cultivars in the Shanghai region of China. Genet. Mol. Res. 2012, 11, 644–650. [Google Scholar] [CrossRef]
- Soriano, J.M.; Villegas, D.; Aranzana, M.J.; García Del Moral, L.F.; Royo, C. Genetic structure of modern durum wheat cultivars and mediterranean landraces matches with their agronomic performance. PLoS ONE 2016, 11, e0160983. [Google Scholar] [CrossRef]
- Alipour, H.; Bihamta, M.R.; Mohammadi, V.; Peyghambari, S.A.; Bai, G.; Zhang, G. Genotyping-by-sequencing (GBS) revealed molecular genetic diversity of Iranian wheat landraces and cultivars. Front. Plant Sci. 2017, 8, 1293. [Google Scholar] [CrossRef] [PubMed]
- Nersting, L.G.; Andersen, S.B.; Von Bothmer, R.; Gullord, M.; Bagger Jørgensen, R. Morphological and molecular diversity of Nordic oat through one hundred years of breeding. Euphytica 2006, 150, 327–337. [Google Scholar] [CrossRef]
- Fu, Y.B.; Peterson, G.W.; Scoles, G.; Rossnagel, B.; Schoen, D.J.; Richards, K.W. Allelic Diversity Changes in 96 Canadian Oat Cultivars Released from 1886 to 2001. Crop Sci. 2003, 43, 1989–1995. [Google Scholar] [CrossRef]
- Achleitner, A.; Tinker, N.A.; Zechner, E.; Buerstmayr, H. Genetic diversity among oat varieties of worldwide origin and associations of AFLP markers with quantitative traits. Theor. Appl. Genet. 2008, 117, 1041–1053. [Google Scholar] [CrossRef] [PubMed]
- Paczos-Grzęda, E. Pedigree, RAPD and simplified AFLP-based assessment of genetic relationships among Avena sativa L. cultivars. Euphytica 2004, 138, 13–22. [Google Scholar] [CrossRef]
- Boczkowska, M.; Tarczyk, E. Genetic diversity among Polish landraces of common oat (Avena sativa L.). Genet. Resour. Crop Evol. 2013, 60, 2157–2169. [Google Scholar] [CrossRef]
- Paczos-Grzeda, E. Wykorzystanie metod ISSR i RAPD oraz analizy rodowodow do oceny podobienstwa miedzyodmianowego Avena sativa L. Zesz. Probl. Postępów Nauk Rol. 2007, 517, 547–558. [Google Scholar]
- Tinker, N.A.; Kilian, A.; Wight, C.P.; Heller-Uszynska, K.; Wenzl, P.; Rines, H.W.; Bjørnstad, Å.; Howarth, C.J.; Jannink, J.-L.; Anderson, J.M.; et al. New DArT markers for oat provide enhanced map coverage and global germplasm characterization. BMC Genomics 2009, 10, 39. [Google Scholar] [CrossRef]
- Newell, M.A.; Cook, D.; Tinker, N.A.; Jannink, J.L. Population structure and linkage disequilibrium in oat (Avena sativa L.): Implications for genome-wide association studies. Theor. Appl. Genet. 2011, 122, 623–632. [Google Scholar] [CrossRef]
- Paczos-Grzęda, E.M.; Bednarek, P.T.; Koroluk, A. Zastosowanie markerów silicoDArT do oceny polimorfizmu międzyodmianowego. Folia Pomeranae Univ. Technol. Stetin. Agric. Aliment. Piscaria Zootech. 2014, 30, 75–84. [Google Scholar]
- Nagaoka, T.; Ogihara, Y. Applicability of inter-simple sequence repeat polymorphisms in wheat for use as DNA markers in comparison to RFLP and RAPD markers. Theor. Appl. Genet. 1997, 94, 597–602. [Google Scholar] [CrossRef]
- Sofalian, O.; Chaparzadeh, N.; Javanmard, A.; Hejazi, M.S. Study the genetic diversity of wheat landraces from northwest of Iran based on ISSR molecular markers. Int. J. Agric. Biol. 2008, 10, 466–468. [Google Scholar]
- Virk, P.S.; Zhu, J.; Newbury, H.J.; Bryan, G.J.; Jackson, M.T.; Ford-Lloyd, B.V. Effectiveness of different classes of molecular marker for classifying and revealing variation in rice (Orzyza sativa) germplasm. Euphytica 2000, 112, 275–284. [Google Scholar] [CrossRef]
- Kolodinska Brantestam, A.; von Bothmer, R.; Dayteg, C.; Rashal, I.; Tuvesson, S.; Weibull, J. Inter simple sequence repeat analysis of genetic diversity and relationships in cultivated barley of Nordic and Baltic origin. Hereditas 2004, 141, 186–192. [Google Scholar] [CrossRef]
- Hou, Y.-C.; Yan, Z.; Wei, Y.-M.; Zheng, Y.-L. Genetic diversity in barley from West China based on RAPD and ISSR analysis. Barley Genet. Newslet. 2005, 35, 9–22. [Google Scholar]
- Fernández, M.E.; Figueiras, A.M.; Benito, C. The use of ISSR and RAPD markers for detecting DNA polymorphism, genotype identification and genetic diversity among barley cultivars with known origin. Theor. Appl. Genet. 2002, 104, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Pujar, S.; Tamhankar, S.; Gupta, V.; Rao, V. Diversity analysis of Indian tetraploid wheat using intersimple sequence repeat markers reveals their superiority over random amplified polymorphic DNA markers. Biochem. Genet. 2002, 40, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Etminan, A.; Pour-Aboughadareh, A.; Mohammadi, R.; Ahmadi-Rad, A.; Noori, A.; Mahdavian, Z.; Moradi, Z. Applicability of start codon targeted (SCoT) and inter-simple sequence repeat (ISSR) markers for genetic diversity analysis in durum wheat genotypes. Biotechnol. Biotechnol. Equip. 2016, 30, 1075–1081. [Google Scholar] [CrossRef]
- Nosair, H. Genetic diversity studies on seven Egyptian wheat (Triticum aestivum) cultivars using Scot and ISSR polymorphism markers. Taeckholmia 2020, 40, 143–151. [Google Scholar]
- Abdel-Lateif, K.S.; Hewedy, O.A. Genetic diversity among egyptian wheat cultivars using SCoT and ISSR markers. Sabrao J. Breed. Genet. 2018, 50, 36–45. [Google Scholar]
- Blair, M.W.; Garris, A.J.; Iyer, A.S.; Chapman, B.; Kresovich, S.; McCouch, S.R. High resolution genetic mapping and candidate gene identification at the xa5 locus for bacterial blight resistance in rice (Oryza sativa L.). Theor. Appl. Genet. 2003, 107, 62–73. [Google Scholar] [CrossRef]
- Al-Turki, T.A.; Basahi, M.A. Assessment of ISSR based molecular genetic diversity of Hassawi rice in Saudi Arabia. Saudi J. Biol. Sci. 2015, 22, 591–599. [Google Scholar] [CrossRef]
- Tripathi, S.; Singh, S.K.; Srivashtav, V.; Khaire, A.R.; Vennela, P.; Singh, D.K. Molecular diversity analysis in rice (Oryza sativa L.) using SSR markers. Electron. J. Plant Breed. 2020, 11, 776–782. [Google Scholar]
- Blair, M.W.; McCouch, S.R.; Panaud, O. Inter-simple sequence repeat (ISSR) amplification for analysis of microsatellite motif frequency and fingerprinting in rice (Oryza sativa L.). Theor. Appl. Genet. 1999, 98, 780–792. [Google Scholar] [CrossRef]
- Petrovicová, L.; Balážová, Ž.; Vivodík, M.; Gálová, Z. Detection genetic variability Of Secale cereale L. by scot markers. Potravin. Slovak J. Food Sci. 2017, 11, 197–202. [Google Scholar] [CrossRef][Green Version]
- Aboulila, A.A.; Mansour, M. Efficiency of Triple-SCoT Primer in Characterization of Genetic Diversity and Genotype-Specific Markers against SSR Fingerprint in Some Egyptian Barley Genotypes. Am. J. Mol. Biol. 2017, 07, 123–137. [Google Scholar] [CrossRef]
- He, X.; Bjørnstad, Å. Diversity of North European oat analyzed by SSR, AFLP and DArT markers. Theor. Appl. Genet. 2012, 125, 57–70. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Number | Variety | Country | Breeder |
|---|---|---|---|
| 1 | Agent | Poland | HR–Strzelce |
| 2 | Arden | Poland | DANKO |
| 3 | Berdysz | Poland | DANKO |
| 4 | Bingo | Poland | HR–Strzelce |
| 5 | Breton | Poland | MHR |
| 6 | Elegant | Poland | HR–Strzelce |
| 7 | Figaro | Poland | DANKO |
| 8 | Harnaś | Poland | MHR |
| 9 | Komfort | Poland | HR–Strzelce |
| 10 | Krezus | Poland | HR–Strzelce |
| 11 | Navigator | Poland | HR–Strzelce |
| 12 | Paskal | Poland | HR–Strzelce |
| 13 | Romulus | Poland | DANKO |
| 14 | Zuch | Poland | DANKO |
| 15 | Lion | Germany | Saaten–Union |
| 16 | Monsoon | Germany | Saaten–Union |
| 17 | Perun | Germany | Saaten–Union |
| 18 | Abel | Czech Republic | SELGEN |
| 19 | Atego | Czech Republic | SELGEN |
| 20 | Azur | Czech Republic | SELGEN |
| 21 | Cavaliere | Czech Republic | SELGEN |
| 22 | Gregor | Czech Republic | SELGEN |
| 23 | Kertag, New | Czech Republic | SELGEN |
| 24 | Korok | Czech Republic | SELGEN |
| 25 | Neclan | Czech Republic | SELGEN |
| 26 | Norbert | Czech Republic | SELGEN |
| 27 | Obelisk | Czech Republic | SELGEN |
| 28 | Oberon | Czech Republic | SELGEN |
| 29 | Raven | Czech Republic | SELGEN |
| 30 | Rosemary | Czech Republic | SELGEN |
| 31 | Sagar | Czech Republic | SELGEN |
| 32 | Vok | Czech Republic | SELGEN |
| 33 | Kamil | Czech Republic | SELGEN |
| 34 | Patrik | Czech Republic | SELGEN |
| 35 | Celeste | Czech Republic | SELGEN |
| 36 | Merlin | Czech Republic | SELGEN |
| Primer | Sequence | Number of Products | Primer Diversity % | Frequency of Polymorphic Products | Resolving Power of the Primer (Rp) | Polymorphic Information Content (PIC) | |
|---|---|---|---|---|---|---|---|
| Total | Total | ||||||
| SR 6 | (GT)8 C | 11 | 6 | 54.55 | 0.55 | 2.23 | 0.22 |
| SR 17 | (GA)8 YC | 11 | 7 | 63.64 | 0.43 | 4.99 | 0.14 |
| SR 32 | (AG)8 YT | 12 | 7 | 58.33 | 0.32 | 9.81 | 0.13 |
| SR 39 | (GA)8 GG | 16 | 9 | 56.25 | 0.76 | 7.83 | 0.17 |
| SR 42 | (AG)8 YA | 24 | 11 | 45.83 | 0.41 | 2.39 | 0.17 |
| SR 50 | (TC)9 C | 10 | 6 | 60.00 | 0.42 | 2.64 | 0.22 |
| SR 54 | (CT)9 T | 10 | 3 | 30.00 | 0.36 | 0.49 | 0.10 |
| SR 59 | (GATA)6 T | 9 | 5 | 55.56 | 0.73 | 3.35 | 0.20 |
| Total | 103 | 54 | 52.43 | ||||
| Average/primer | 12.87 | 6.75 | |||||
| Primer | Sequence 5′->3′ | Number of Products | Primer Diversity % | Frequency of Polymorphic Products | Resolving Power of the Primer (RP) | Polymorphic Information Content (PIC) | |
|---|---|---|---|---|---|---|---|
| Total | Polymorphic | ||||||
| SCoT 3 | CAA-CAA-TGG-CTA-CCA-CCG | 13 | 6 | 46.15 | 0.61 | 16.0 | 0.21 |
| SCoT 4 | CAA-CAA-TGG-CTA-CCA-CCT | 9 | 5 | 55.56 | 0.38 | 4.49 | 0.15 |
| SCoT 12 | ACG-ACA-TGG-CGA-CCA-ACG | 9 | 4 | 44.44 | 0.61 | 3.14 | 0.12 |
| SCoT 33 | CCA-TGG-CTA-CCA-CCG-CCA | 13 | 4 | 37.77 | 0.29 | 0.49 | 0.08 |
| SCoT 34 | ACC-ATG-GCT-ACC-ACC-GCA | 14 | 8 | 57.14 | 0.40 | 0.67 | 0.21 |
| Total | 58 | 27 | 46.55 | ||||
| Average/primer | 11.6 | 5.4 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cieplak, M.; Okoń, S.; Werwińska, K. Genetic Similarity of Avena sativa L. Varieties as an Example of a Narrow Genetic Pool of Contemporary Cereal Species. Plants 2021, 10, 1424. https://doi.org/10.3390/plants10071424
Cieplak M, Okoń S, Werwińska K. Genetic Similarity of Avena sativa L. Varieties as an Example of a Narrow Genetic Pool of Contemporary Cereal Species. Plants. 2021; 10(7):1424. https://doi.org/10.3390/plants10071424
Chicago/Turabian StyleCieplak, Magdalena, Sylwia Okoń, and Krystyna Werwińska. 2021. "Genetic Similarity of Avena sativa L. Varieties as an Example of a Narrow Genetic Pool of Contemporary Cereal Species" Plants 10, no. 7: 1424. https://doi.org/10.3390/plants10071424
APA StyleCieplak, M., Okoń, S., & Werwińska, K. (2021). Genetic Similarity of Avena sativa L. Varieties as an Example of a Narrow Genetic Pool of Contemporary Cereal Species. Plants, 10(7), 1424. https://doi.org/10.3390/plants10071424