
Selenium Fortification Alters the Growth, Antioxidant Characteristics and Secondary Metabolite Profiles of Cauliflower (Brassica oleracea var. botrytis) Cultivars in Hydroponic Culture

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Growth Parameters
2.2. Changes in Photosynthetic Pigments, Antioxidant Capacity, Phenolic Compounds and Selenium Content in the Floret and the Leaves
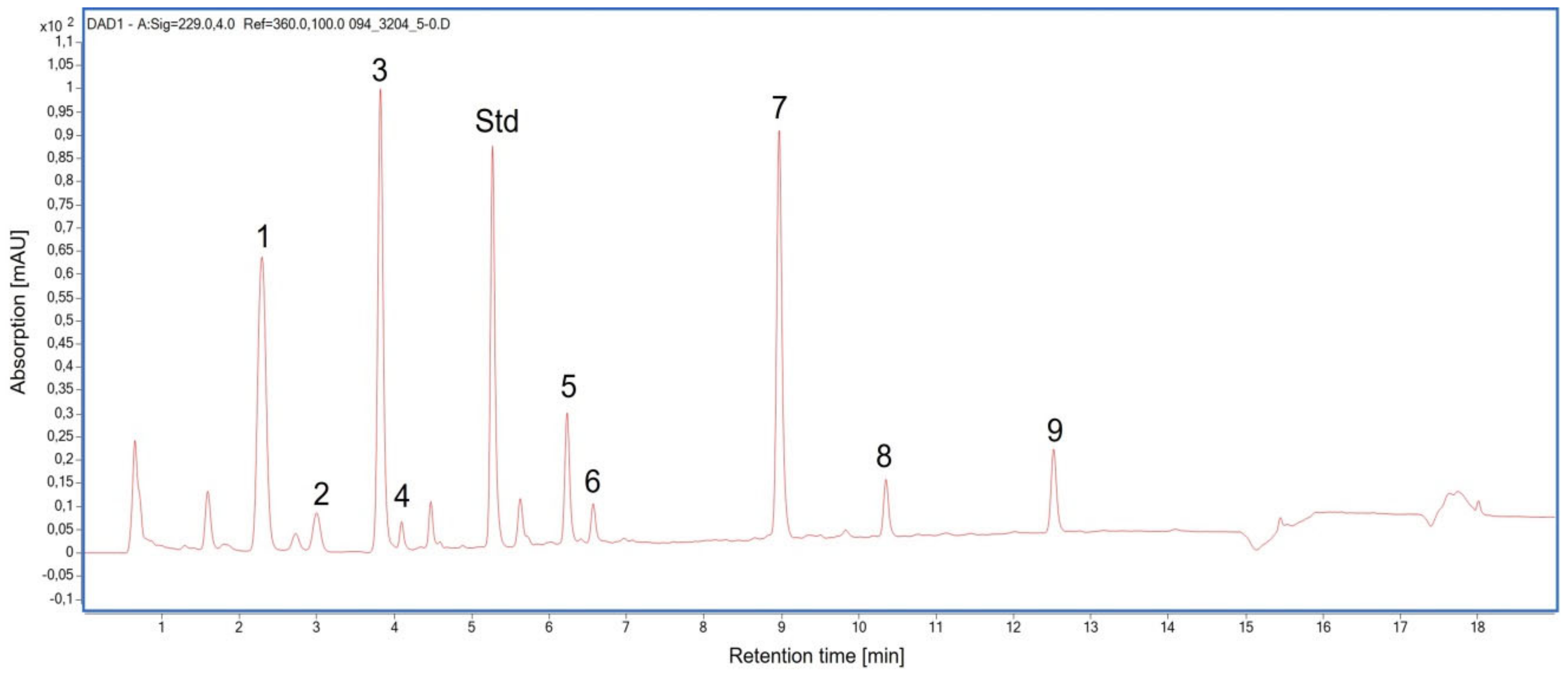
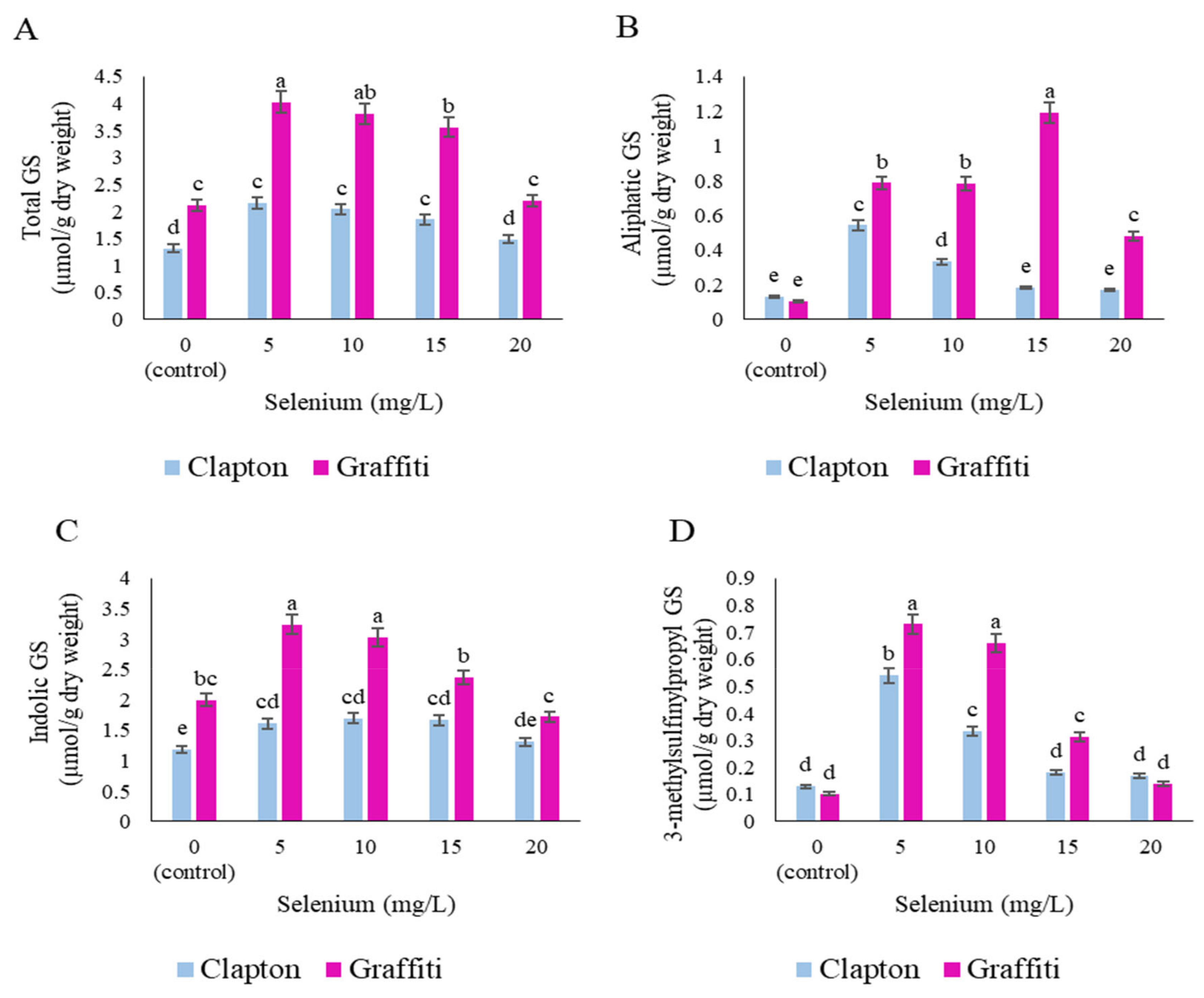
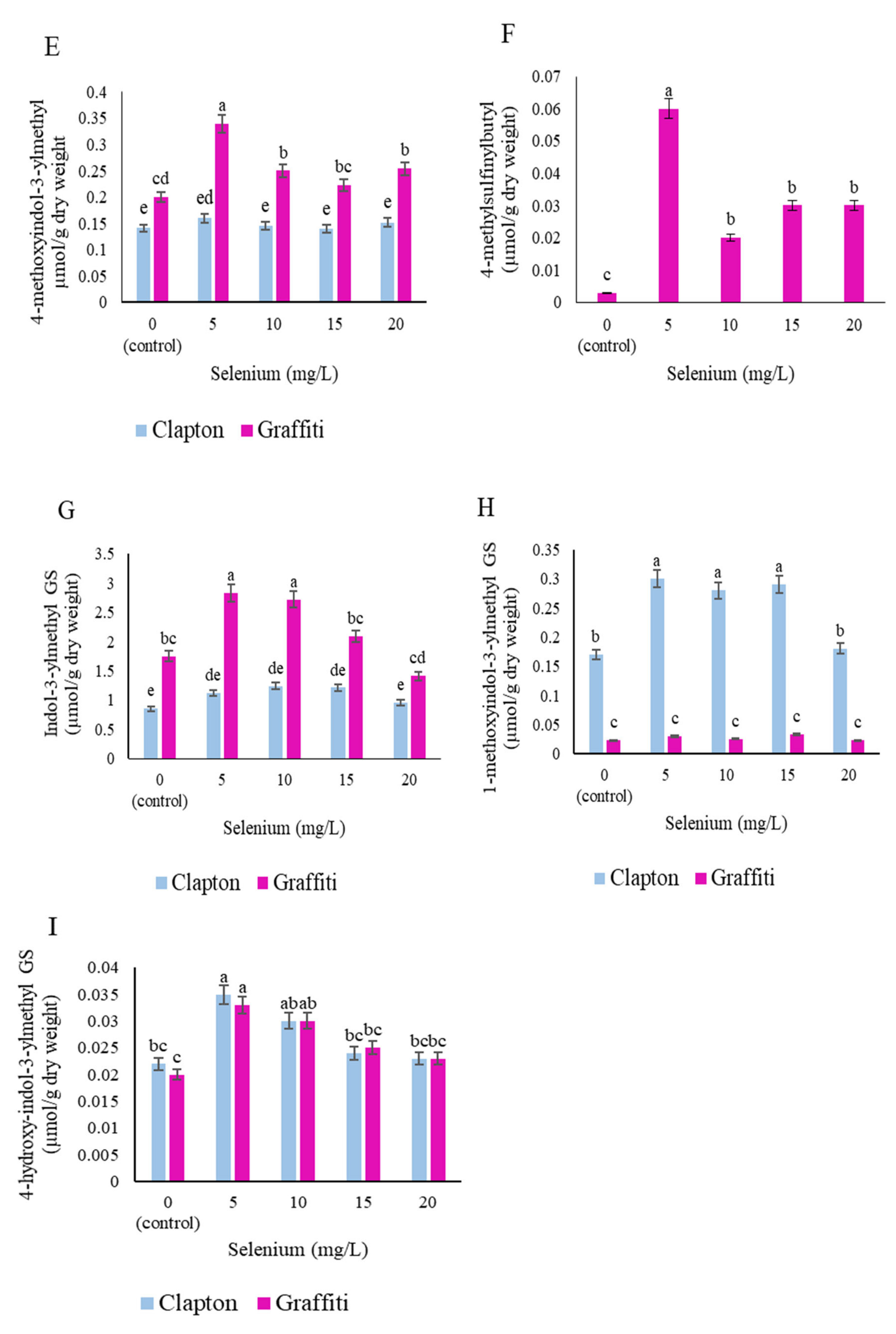
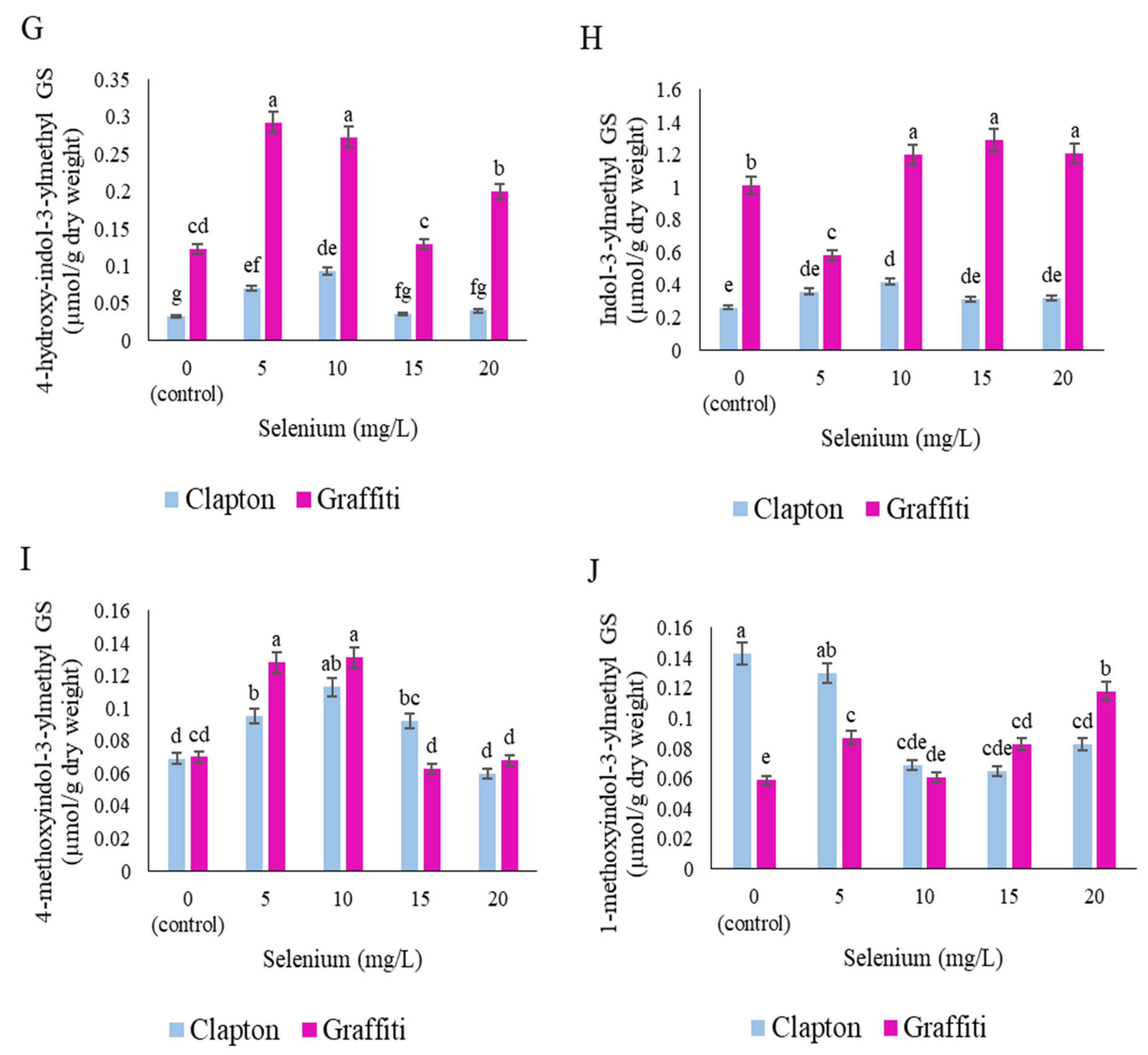
2.3. Glucosinolate Compounds in Leaves
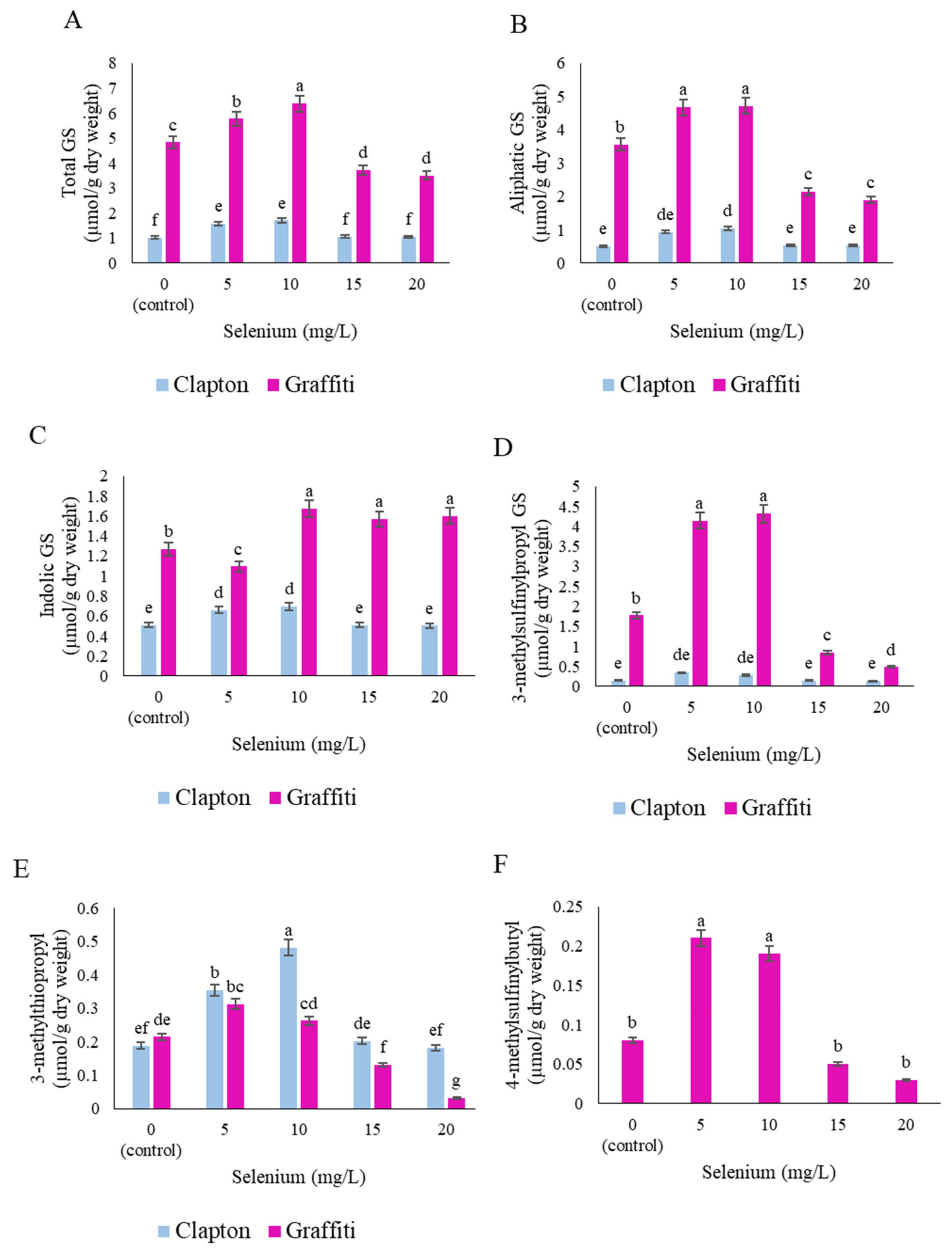
2.4. Glucosinolate Compounds in the Floret
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Materials
5.2. Determination of Photosynthetic Pigments, Polyphenolic Compounds and Antioxidant Capacity
5.3. Determination of the Total Selenium Content in the Leaves and Florets
5.4. Glucosinolate Compounds
5.5. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Longchamp, M.; Angeli, N.; Castrec-Rouelle, M. Selenium uptake in Zea mays supplied with selenate or selenite under hydroponic conditions. J. Plant Soil. 2013, 362, 107–117. [Google Scholar] [CrossRef]
- Jóźwiak, W.; Mleczek, M.; Politycka, B. The effect of exogenous selenium on the growth and photosynthetic pigments content of cucumber seedlings. J. Fresenius Environ. Bull. 2016, 25, 142–152. [Google Scholar]
- Irmak, S. Effects of selenium application on plant growth and some quality parameters in peanut (Arachis hypogaea). Pak. J. Biol. Sci. 2017, 20, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Cao, F.; Cheng, W.; Zhang, G.; Wu, F. Modulation of exogenous glutathione in phytochelatins and photosynthetic performance against Cd stress in the two rice genotypes differing in Cd tolerance. J. Biol. Trace Elem. Res. 2011, 143, 1159–1173. [Google Scholar] [CrossRef]
- Schiavon, M.; dall’Acqua, S.; Mietto, A.; Pilon-Smits, E.A.; Sambo, P.; Masi, A.; Malagoli, M. Selenium fertilization alters the chemical composition and antioxidant constituents of tomato (Solanum lycopersicon L.). J. Agric. Food Chem. 2013, 61, 10542–10554. [Google Scholar] [CrossRef]
- Chomchan, R.; Siripongvutikorn, S.; Puttarak, P. Selenium bio-fortification: An alternative to improve phytochemicals and bioactivities of plant foods. J. Funct. Foods Health Dis. 2017, 7, 263–279. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, S.R.; Kwak, J.H. Chemical composition and antioxidant activity in different tissues of Brassica vegetables. J. Mol. 2015, 20, 1228–1243. [Google Scholar] [CrossRef] [Green Version]
- Malagoli, M.; Schiavon, M.; dall’Acqua, S.; Pilon-Smits, E.A.H. Effects of selenium biofortification on crop nutritional quality. Front. J. Plant Sci. 2015, 6, 280. [Google Scholar] [CrossRef] [Green Version]
- Wijaya, H.; Rouw, R.H.; Kadir, A.R. Brassica box food products as a healthy local food innovation in The Covid-19 pandemic period. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2020. [Google Scholar]
- Wiesner-Reinhold, M.; Schreiner, M.; Baldermann, S.; Schwarz, D.; Hanschen, F.S.; Kipp, A.P.; McKenzie, M.J. Mechanisms of selenium enrichment and measurement in brassicaceous vegetables, and their application to human health. J. Front. Plant Sci. 2017, 8, 1365. [Google Scholar] [CrossRef] [Green Version]
- Kieliszek, M.; Błażejak, S. Current knowledge on the importance of selenium in food for living organisms: A review. J. Mol. 2016, 21, 609. [Google Scholar] [CrossRef] [Green Version]
- Blažević, I.; Montaut, S.; Burčul, F.; Rollin, P. Glucosinolates: Novel Sources and Biological Potential; Ramawat, K.G., Mérillon, J.-M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 3–60. [Google Scholar]
- Kieliszek, M. Selenium–fascinating microelement, properties and sources in food. J. Mol. 2019, 24, 1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezzarossa, B.; Remorini, D.; Gentile, M.L.; Massai, R. Effects of foliar and fruit addition of sodium selenate on selenium accumulation and fruit quality. J. Sci. Food Agric. 2012, 92, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.W.; Ha, J.; Liang, S.X.; Kang, W.J. Protective role of selenium on garlic growth under cadmium stress. Commun. Soil Sci. Plant Anal. 2010, 41, 1195–1204. [Google Scholar] [CrossRef]
- Abdel, C.G.; Ali, H.O. Selenium enrichments of Cauliflower (Brassica oleracea L. var. Botrytis) and Broccoli (Brassica oleracea L. var. Italica) grown under drip-hydroponic system hybrid 704. J. Farming Allied Sci. 2016, 5, 126–167. [Google Scholar]
- Dziubinska, H.; Filek, M.; Krol, E.; Trebacz, K. Cadmium and selenium modulate slow vacuolar channels in rape (Brassica napus) vacuoles. J. Plant Physiol. 2010, 167, 1566–1570. [Google Scholar] [CrossRef] [PubMed]
- Ulhassan, Z.; Gill, R.A.; Ali, S.; Mwamba, T.M.; Ali, B.; Wang, J.; Zhou, W. Dual behavior of selenium: Insights into physio-biochemical, anatomical and molecular analyses of four Brassica napus cultivars. J. Chemosphere 2019, 225, 329–341. [Google Scholar] [CrossRef]
- Podsędek, A. Natural antioxidants and antioxidant capacity of Brassica vegetables: A review. J. LWT-Food Sci. Technol. 2007, 40, 1–11. [Google Scholar] [CrossRef]
- Cömert, E.D.; Mogol, B.A.; Gökmen, V. Relationship between color and antioxidant capacity of fruits and vegetables. J. Food Sci. 2020, 2, 1–10. [Google Scholar] [CrossRef]
- Golob, A.; Novak, T.; Maršić, N.K.; Šircelj, H.; Stibilj, V.; Jerše, A.; Germ, M. Biofortification with selenium and iodine changes morphological properties of (Brassica oleracea L. var. gongylodes) and increases their contents in tubers. J. Plant Physiol. Biochem. 2020, 150, 234–243. [Google Scholar] [CrossRef]
- Chu, J.; Yao, X.; Zhang, Z. Responses of wheat seedlings to exogenous selenium supply under cold stress. J. Biol. Trace Elem. Res. 2010, 136, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Finley, J.W.; Sigrid-Keck, A.; Robbins, R.J.; Hintze, K.J. Selenium enrichment of broccoli: Interactions between selenium and secondary plant compounds. J. Nutr. 2005, 135, 1236–1238. [Google Scholar] [CrossRef] [Green Version]
- Ríos, J.J.; Rosales, M.A.; Blasco, B.; Cervilla, L.M.; Romero, L.; Ruiz, J.M. Biofortification of Se and induction of the antioxidant capacity in lettuce plants. J. Sci. Hortic. 2008, 116, 248–255. [Google Scholar] [CrossRef]
- Zhong, Y.; Cheng, J.J. Effects of selenite on unicellular green microalga Chlorella pyrenoidosa: Bioaccumulation of selenium, enhancement of photosynthetic pigments, and amino acid production. J. Agric. Food Chem. 2017, 65, 10875–10883. [Google Scholar] [CrossRef]
- Bañuelos, G.S.; Arroyo, I.; Pickering, I.J.; Yang, S.I.; Freeman, J.L. Selenium biofortification of broccoli and carrots grown in soil amended with Se-enriched hyperaccumulator Stanleya pinnata. J. Food Chem. 2015, 166, 603–608. [Google Scholar] [CrossRef]
- Ávila, F.W.; Yang, Y.; Faquin, V.; Ramos, S.J.; Guilherme, L.R.G.; Thannhauser, T.W.; Li, L. Impact of selenium supply on Se-methylselenocysteine and glucosinolate accumulation in selenium-biofortified Brassica sprouts. J. Food Chem. 2014, 165, 578–586. [Google Scholar] [CrossRef]
- Matich, A.J.; McKenzie, M.J.; Lill, R.E.; McGhie, T.K.; Chen, R.K.Y.; Rowan, D.D. Distribution of selenoglucosinolates and their metabolites in Brassica treated with sodium selenate. J. Agric. Food Chem. 2015, 63, 1896–1905. [Google Scholar] [CrossRef]
- Bhandari, S.R.; Jo, J.S.; Lee, J.G. Comparison of glucosinolate profiles in different tissues of nine Brassica crops. J. Mol. 2015, 20, 15827–15841. [Google Scholar] [CrossRef] [PubMed]
- Bellostas, N.; Kachlicki, P.; Sørensen, J.C.; Sørensen, H. Glucosinolate profiling of seeds and sprouts of B. oleracea varieties used for food. J. Sci. Hortic. 2007, 114, 234–242. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Juvik, J.A. Effect of selenium fertilization and methyl jasmonate treatment on glucosinolate accumulation in broccoli florets. J. Am. Soc. Hortic. Sci. 2011, 136, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyhenoloxidases in Beta vulgari. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Singleton, V.L.; Rossi, J.A., Jr. Colorimetric of total phenols with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Koleva, I.I.; van Beek, T.A.; Linssen, J.P.H.; de Groot, A.; Evstatieva, L.N. Screening of plant extracts for antioxidant activity: A comparative study on three testing methods. Phytochem. Anal. 2002, 13, 8–17. [Google Scholar] [CrossRef]
- Szeles, E. Studying of Change of Selenium Species in Soil and Plant Samples from a Long-Term Field Experiment. Ph.D. Thesis, University of Debercen, Debrecen, Hungary, 2007. [Google Scholar]
- Wiesner, M.; Zrenner, R.; Krumbein, A.; Glatt, H.; Schreiner, M. Genotypic variation of the glucosinolate profile in pak choi (Brassica rapa ssp. chinensis). J. Agric. Food Chem. 2013, 61, 1943–1953. [Google Scholar] [CrossRef] [PubMed]
- Hanschen, F.S. Domestic boiling and salad preparation habits affect glucosinolate degradation in red cabbage (Brassica oleracea var. capitata f. rubra). J. Food Chem. 2020, 321, 126694. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Treatment (mg/L) | Floret Fresh Weight (g) | Floret Diameter (cm) | Root Fresh Weight (g) | Root Dry Weight (g) | Stem Diameter (cm) | Stem Height (cm) |
|---|---|---|---|---|---|---|---|
| Clapton | Control | 448.49 ± 8.10c | 14.41 ± 0.31e | 95.66 ± 0.63e | 12.33 ± 0.31d | 3.18 ± 0.02ed | 22.16 ± 1.10c |
| 5 Na2SeO4 | 503.99 ± 8.65ab | 15.27 ± 0.16ed | 113.83 ± 5.89cde | 15.79 ± 0.41cd | 3.27 ± 0.02cde | 23.58 ± 0.67c | |
| 10 Na2SeO4 | 527.45 ± 9.02a | 16.24 ± 0.15cd | 132.50 ± 2.12c | 19.84 ± 0.36c | 3.58 ± 0.06b | 22.33 ± 0.47c | |
| 15 Na2SeO4 | 512.02 ± 27.10ab | 15.33 ± 0.27de | 119.99 ± 3.14cd | 15.49 ± 1.02cd | 3.50 ± 0.03bc | 21.72 ± 0.31c | |
| 20 Na2SeO4 | 469.22 ±5.34bc | 15.29 ± 0.33de | 108.05 ± 3.54ed | 14.49 ± 0.07d | 3.35 ± 0.02bcde | 21.22 ± 0.17c | |
| Graffiti | Control | 241.33 ± 6.76e | 15.71 ± 0.01cd | 210.22 ± 13.85b | 25.55 ± 2.54b | 3.12 ± 0.05e | 30.97 ± 1.89b |
| 5 Na2SeO4 | 286.89 ± 18.48ed | 17.84 ± 0.15a | 241.25 ± 3.65a | 29.32 ± 2.06b | 3.30 ± 0.02bcde | 37.10 ± 2.56a | |
| 10 Na2SeO4 | 318.11 ± 2.36d | 18.33 ± 0.65a | 295.78 ± 3.74a | 35.65 ± 0.41a | 3.92 ± 0.23a | 36.00 ± 2.35ab | |
| 15 Na2SeO4 | 316.77 ± 7.61d | 17.54 ± 0.09ab | 248.16 ± 5.11a | 30.32 ± 1.36b | 3.47 ± 0.02bcd | 35.61 ± 1.74ab | |
| 20 Na2SeO4 | 273.33 ± 9.03ed | 16.47 ± 0.28bc | 241.66 ± 2.20a | 26.79 ± 2.37b | 3.39 ± 0.05bcd | 31.71 ± 1.55ab |
| Cultivar | Treatment (mg/L) | Fresh Weight (g) | Leave Area (cm2) | Dry Weight (g) | Root Length (cm) | ||
|---|---|---|---|---|---|---|---|
| Leave | Stem | Leave | Stem | ||||
| Clapton | Control | 329.99 ± 0.64h | 140.99 ± 0.16g | 423.82 ± 2.48f | 18.33 ± 0.42g | 9.16 ± 0.07g | 34.58 ± 0.92de |
| 5 Na2SeO4 | 359.41 ± 2.60g | 156.24 ± 0.76ef | 440.35 ± 0.48e | 22.85 ± 0.33f | 11.22 ± 0.40f | 36.75 ± 1.32cd | |
| 10 Na2SeO4 | 405.08 ± 6.42f | 160.66 ± 2.56e | 473.59 ± 1.03bc | 27.22 ± 0.74e | 14.08 ± 0.24e | 40.66 ± 0.31ab | |
| 15 Na2SeO4 | 366.35 ± 4.36g | 160.41 ± 1.69e | 452.96 ± 1.09ed | 23.06 ± 0.12f | 10.61 ± 0.20gf | 32.5 ± 0.72e | |
| 20 Na2SeO4 | 341.94 ± 1.16h | 147.91 ± 0.52gf | 448.37 ± 1.11e | 22.4 ± 0.17f | 9.7 ± 0.15gf | 32.74 ± 0.67e | |
| Graffiti | Control | 575.75 ± 10.52e | 197.66 ± 0.79d | 455.33 ± 1.33ed | 41.4 ± 0.44d | 20.33 ± 0.16d | 36.88 ± 1.93bcd |
| 5 Na2SeO4 | 607.33 ± 3.52d | 213.66 ± 3.20c | 466.54 ± 3.62cd | 49.78 ± 1.17b | 22.78 ± 0.78c | 38.27 ± 0.96bcd | |
| 10 Na2SeO4 | 728.93 ± 3.05a | 288.83 ± 7.44a | 488.60 ± 3.55b | 55.21 ± 0.45a | 28.38 ± 0.54a | 42.22 ± 0.80a | |
| 15 Na2SeO4 | 708.16 ± 2.80b | 261.5 ± 2.16b | 535.17 ± 14.01a | 46.55 ± 0.67c | 25.33 ± 0.44b | 40.44 ± 1.84abc | |
| 20 Na2SeO4 | 691.01 ± 4.99c | 216.16 ± 3.76c | 472.13 ± 3.24c | 42.63 ± 0.98d | 21.74 ± 0.54cd | 40.22 ± 0.75abc | |
| Cultivar | Treatment (mg/L) | Chlorophyll (mg/g FW) | Carotenoid (mg/g FW) | Antioxidants (DPPH %) | Total Polyphenolic (mg Gallic Acid 100 g−1) | Se Content (mg/kg DW) | ||
|---|---|---|---|---|---|---|---|---|
| a | b | Total | ||||||
| Clapton | Control | 0.41 ± 0.00e | 0.11 ± 0.00de | 0.52 ± 0.01f | 0.12 ± 0.00f | 14 ± 1.92f | 170.6 ± 2.57f | 0.03 ± 0.00g |
| 5 Na2SeO4 | 0.54 ± 0.00c | 0.14 ± 0.00c | 0.68 ± 0.00d | 0.15 ± 0.00e | 18 ± 0.96ef | 178.32 ± 4.62ef | 0.12 ± 0.00f | |
| 10 Na2SeO4 | 0.70 ± 0.00a | 0.16 ± 0.00b | 0.87 ± 0.01b | 0.21 ± 0.00b | 28.5 ± 2.46e | 192.48 ± 2.36d | 0.23 ± 0.02de | |
| 15 Na2SeO4 | 0.74 ± 0.00a | 0.20 ± 0.00a | 0.95 ± 0.00a | 0.25 ± 0.00a | 25 ± 0.48ef | 202.31 ± 3.30d | 0.27 ± 0.03d | |
| 20 Na2SeO4 | 0.61 ± 0.00b | 0.14 ± 0.00bc | 0.76 ± 0.00 | 0.18 ± 0.00c | 16 ± 0.96f | 191.63 ± 5.00ed | 0.57 ± 0.01a | |
| Graffiti | Control | 0.34 ± 0.01f | 0.09 ± 0.00e | 0.44 ± 0.01g | 0.08 ± 0.00g | 147 ± 6.72c | 249.52 ± 0.66c | 0.04 ± 0.00g |
| 5 Na2SeO4 | 0.45 ± 0.01ed | 0.10 ± 0.00de | 0.55 ± 0.01f | 0.14 ± 0.00e | 205.66 ± 1.94a | 256.02 ± 2.26c | 0.18 ± 0.01e | |
| 10 Na2SeO4 | 0.60 ± 0.00b | 0.14 ± 0.00bc | 0.63 ± 0.00e | 0.21 ± 0.00b | 170.33 ± 0.27b | 323.29 ± 4.15a | 0.36 ± 0.00c | |
| 15 Na2SeO4 | 0.48 ± 0.01d | 0.14 ± 0.00bc | 0.74 ± 0.00c | 0.17 ± 0.00d | 145 ± 2.88c | 324.66 ± 1.98a | 0.40 ± 0.00bc | |
| 20 Na2SeO4 | 0.48 ± 0.00d | 0.11 ± 0.00d | 0.60 ± 0.00e | 0.14 ± 0.00e | 129 ± 4.82d | 305.19 ± 6.23b | 0.44 ± 0.00b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saeedi, M.; Soltani, F.; Babalar, M.; Izadpanah, F.; Wiesner-Reinhold, M.; Baldermann, S. Selenium Fortification Alters the Growth, Antioxidant Characteristics and Secondary Metabolite Profiles of Cauliflower (Brassica oleracea var. botrytis) Cultivars in Hydroponic Culture. Plants 2021, 10, 1537. https://doi.org/10.3390/plants10081537
Saeedi M, Soltani F, Babalar M, Izadpanah F, Wiesner-Reinhold M, Baldermann S. Selenium Fortification Alters the Growth, Antioxidant Characteristics and Secondary Metabolite Profiles of Cauliflower (Brassica oleracea var. botrytis) Cultivars in Hydroponic Culture. Plants. 2021; 10(8):1537. https://doi.org/10.3390/plants10081537
Chicago/Turabian StyleSaeedi, Mahboobeh, Forouzandeh Soltani, Mesbah Babalar, Fatemeh Izadpanah, Melanie Wiesner-Reinhold, and Susanne Baldermann. 2021. "Selenium Fortification Alters the Growth, Antioxidant Characteristics and Secondary Metabolite Profiles of Cauliflower (Brassica oleracea var. botrytis) Cultivars in Hydroponic Culture" Plants 10, no. 8: 1537. https://doi.org/10.3390/plants10081537
APA StyleSaeedi, M., Soltani, F., Babalar, M., Izadpanah, F., Wiesner-Reinhold, M., & Baldermann, S. (2021). Selenium Fortification Alters the Growth, Antioxidant Characteristics and Secondary Metabolite Profiles of Cauliflower (Brassica oleracea var. botrytis) Cultivars in Hydroponic Culture. Plants, 10(8), 1537. https://doi.org/10.3390/plants10081537