
Mechanistic Association of Quantitative Trait Locus with Malate Secretion in Lentil (Lens culinaris Medikus) Seedlings under Aluminium Stress


 ,
,
Abstract
:1. Introduction
2. Results
2.1. Phenotyping of Morpho-Physiological Traits in Response to Al Stress
2.1.1. Root Elongation Rate
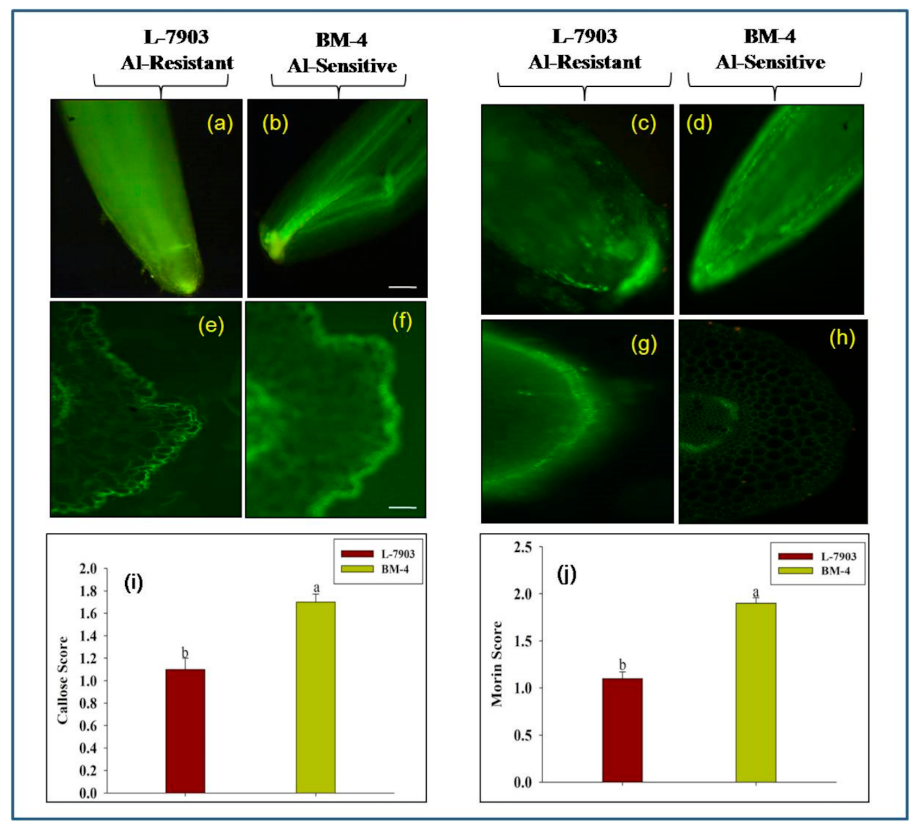
2.1.2. Callose Accumulation in Roots
2.1.3. Al Localization in Root Using Morin Assay
2.1.4. Al Content in Roots
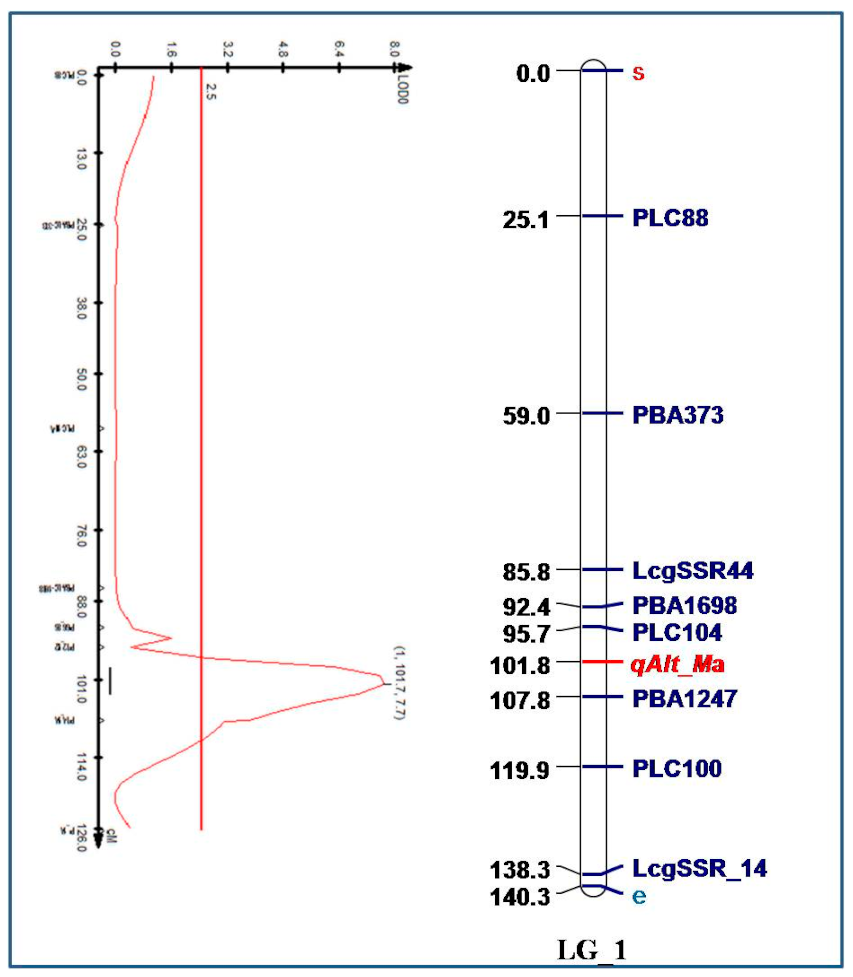
2.2. Mapping a Major QTL for Secretion of Malate
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Estimation of Relative Root Elongation in Response to Al stress
4.3. Determination of Al-induced Callose Formation
4.4. Determination of Al Accumulation using Morin Assay
4.5. Determination of Al Content
4.6. Estimation of Organic Acids in Roots in Response to Al Stress
4.7. Mapping of Molecular Markers
4.8. Construction of Linkage Map
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, D.; Raje, R.S.; Choudhary, A.K. Genetic control of aluminium tolerance in pigeonpea (Cajanus cajan L.). Crop Pasture Sci. 2011, 62, 761–764. [Google Scholar] [CrossRef]
- Panda, S.K.; Baluska, F.; Matsumoto, H. Aluminum stress signaling in plants. Plant Signal. Behav. 2009, 7, 592–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eticha, D.; Thé, C.; Welcker, C.; Narro, L.; Stab, A.; Horst, W.J. Aluminium-induced callose formation in root apices: Inheritance and selection trait for adaptation of tropical maize to acid soils. Field Crop. Res. 2005, 93, 252–263. [Google Scholar] [CrossRef]
- Scott, B.J.; Fisher, J.A.; Cullis, B.R. Aluminium tolerance and lime increase wheat yield on the acidic soils of central and southern New South Wales. Aust. J. Exp. Agric. Int. 2001, 41, 523–532. [Google Scholar] [CrossRef]
- Tang, C.; Rengel, Z.; Diatloff, E.; Gazey, C. Responses of wheat and barley to liming on a sandy soil with subsoil acidity. Field Crop. Res. 2003, 80, 235–244. [Google Scholar] [CrossRef]
- Singh, D.; Raje, R.S. Genetics of aluminium tolerance in chickpea (Cicer arietinum). Plant Breed. 2011, 130, 563–568. [Google Scholar] [CrossRef]
- Singh, D.; Dikshit, H.K.; Kumar, A. Aluminium tolerance in lentil (Lens culinaris Medik.) with monogenic inheritance pattern. Plant Breed. 2015, 134, 105–110. [Google Scholar] [CrossRef]
- Ma, J.F.; Ryan, P.R.; Delhaize, E. Aluminium tolerance in plants and the complexing role of organic acids. Trends Plant Sci. 2001, 6, 273–278. [Google Scholar] [CrossRef]
- Zheng, S.J.; Ma, J.F.; Matsumoto, H. High aluminum resistance in buckwheat I Al-induced specific secretion of oxalic acid from root tips. Plant Physiol. 1998, 117, 745–751. [Google Scholar] [CrossRef] [Green Version]
- Guo, P.; Bai, G.; Carve, B.; Li, R.; Bernardo, A.; Baum, M. Transcriptional analysis between two wheat near-isogenic lines contrasting in aluminum tolerance under aluminum stress. Mol. Genet. Genom. 2007, 277, 1–12. [Google Scholar] [CrossRef]
- Sasaki, T.; Yamamoto, Y.; Ezaki, B.; Katsuhara, M.; Ahn, S.J.; Ryan, P.R.; Delhaize, E.; Matsumoto, H. A wheat gene encoding an aluminum-activated malate transporter. Plant J. 2004, 37, 645–653. [Google Scholar] [CrossRef]
- Furukawa, J.; Yamaji, N.; Wang, H.; Mitani, N.; Murata, Y.; Sato, K.; Katsuhara, M.; Takeda, K.; Ma, J.F. An aluminum-activated citrate transporter in barley. Plant Cell Physiol. 2007, 48, 1081–1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magalhaes, J.V.; Liu, J.; Guimara˜es, C.T.; Lana, U.G.; Alves, V.M.; Wang, Y.H.; Schaffert, R.E.; Hoekenga, O.A.; Pineros, M.A.; Shaff, J.E.; et al. A gene in the multidrug and toxic compound extrusion (MATE) family confers aluminum tolerance in sorghum. Nat. Genet. 2007, 39, 1156–1161. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Delhaize, E.; Zhou, M.; Ryan, P.R. The barley MATE gene, HvAACT1, increases citrate efflux and Al3+ tolerance when expressed in wheat and barley. Ann. Bot. 2013, 112, 603–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, D.; Dikshit, H.K.; Singh, R. Variation of aluminium tolerance in lentil (Lens culinaris Medik.). Plant Breed. 2012, 1, 751–761. [Google Scholar] [CrossRef]
- Singh, D.; Pal, M.; Singh, C.K.; Taunk, J.; Jain, P.; Chaturvedi, A.K.; Maurya, S.; Karwa, S.; Singh, R.; Tomar, R.S.S.; et al. Molecular scanning and morpho-physiological dissection of component mechanism in Lens species in response to aluminium stress. PLoS ONE 2016, 11, e0160073. [Google Scholar] [CrossRef] [PubMed]
- Singh, C.K.; Singh, D.; Tomar, R.S.S.; Karwa, S.; Upadhyaya, K.C.; Pal, M. Molecular mapping of aluminium resistance loci based on root re-growth and Al-induced fluorescent signals (callose accumulation) in lentil (Lens culinaris Medikus). Mol. Biol. Rep. 2018, 45, 2103–2113. [Google Scholar] [CrossRef]
- Yusuf, M.; Khan, T.A.; Fariduddin, Q. Responses of photosynthesis, stress markers and antioxidants under aluminium, salt and combined stresses in wheat cultivars. Cogent Food Agric. 2016, 2, 1216246. [Google Scholar] [CrossRef]
- Carcamo, M.P.; Reyes-Díaz, M.; Rengel, Z.; Alberdi, M.; Omena-Garcia, R.P.; Nunes-Nesi, A.; Inostroza-Blancheteau, C. Aluminum stress differentially affects physiological performance and metabolic compounds in cultivars of highbush blueberry. Sci. Rep. 2019, 9, 11275. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Choudhary, A.K. Inheritance of aluminium tolerance in pea. Plant Breed. 2010, 129, 688–692. [Google Scholar] [CrossRef]
- Raman, H.; Zhang, K.; Cakir, M.; Appels, R.; Garvin, D.F.; Maron, L.G.; Kochian, L.V.; Moroni, J.S.; Raman, R.; Imtiaz, M.; et al. Molecular characterization and mapping of ALMT1, the aluminium-tolerance gene of bread wheat (Triticumaestivum L.). Genome 2005, 48, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Raman, H.; Moroni, J.S.; Sato, K.; Read, B.J.; Scott, B.J. Identification of AFLP and microsatellite markers linked with an aluminium tolerance gene in barley (Hordeum vulgare L.). Theor. Appl. Genet. 2002, 105, 458–464. [Google Scholar] [CrossRef]
- Ma, J.F.; Shen, R.; Zhao, Z.; Wissuwa, M.; Takeuchi, Y.; Ebitani, T.; Yano, M. Response of rice to Al stress and identification of quantitative trait loci for Al tolerance. Plant Cell Physiol. 2002, 43, 652–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.L.; Bai, G.H.; Ma, H.X.; Carver, B.F. Quantitative trait loci for aluminium resistance in wheat. Mol. Breed. 2007, 19, 153–161. [Google Scholar] [CrossRef]
- Guimaraes, C.T.; Simoes, C.C.; Pastina, M.M.; Maron, L.G.; Magalhaes, J.V.; Vasconcellos, R.C.; Guimaraes, L.J.; Lana, U.G.; Tinoco, C.F.; Noda, R.W.; et al. Genetic dissection of Al tolerance QTLs in the maize genome by high density SNP scan. BMC Genom. 2014, 15, 153. [Google Scholar] [CrossRef] [Green Version]
- Miller, T.E.; Iqbal, N.; Reader, S.M.; Mahmood, A.; Cant, K.A.; King, I.P. A cytogenetic approach to the improvement of aluminium tolerance in wheat. New Phytol. 1997, 137, 93–98. [Google Scholar] [CrossRef]
- Bian, M.; Zhou, M.; Sun, D.; Li, C. Molecular approaches unravel the mechanism of acid soil tolerance in plants. Crop J. 2013, 1, 91–104. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.; Wu, D.; Jabeen, Z.; Huang, Y.; Huang, Y.; Zhang, G. Genome-wide association analysis of aluminum tolerance in cultivated and Tibetan wild barley. PLoS ONE 2013, 8, e69776. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Raman, H.; Zhou, M.; Ryan, P.R.; Delhaize, E.; Hebb, D.M.; Coombes, N.; Mendham, N. High-resolution mapping of the Alp locus and identification of a candidate gene HvMATE controlling aluminium tolerance in barley (Hordeum vulgare L.). Theor. Appl. Genet. 2007, 115, 265–276. [Google Scholar] [CrossRef]
- Wang, X.; Cheng, Y.; Yang, C.; Yang, C.; Mu, Y.; Xia, Q.; Ma, Q. QTL mapping for aluminum tolerance in RIL population of soybean (Glycine max L.) by RAD sequencing. PLoS ONE 2019, 14, e0223674. [Google Scholar] [CrossRef]
- Maron, L.G.; Piñeros, M.A.; Guimarães, C.T.; Magalhaes, J.V.; Pleiman, J.K.; Mao, C.; Shaff, J.; Belicuas, S.N.; Kochian, L.V. Two functionally distinct members of the MATE (multi-drug and toxic compound extrusion) family of transporters potentially underlie two major aluminum tolerance QTLs in maize. Plant J. 2010, 61, 728–740. [Google Scholar] [CrossRef] [PubMed]
- FAO (Food and Agricultural Organization). 2018. Available online: http://www.fao.org/faostat/en/#data (accessed on 7 October 2020).
- Ma, J.F.; Nagao, S.; Sato, K.; Ito, H.; Furukawa, J.; Takeda, K. Molecular mapping of a gene responsible for Al-activated secretion of citrate in barley. J. Exp. Bot. 2004, 55, 1335–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, Y.; Jiang, L.; Su, N.; Wang, J.K.; Deng, P.; Ma, J.F.; Zhai, H.Q.; Wan, J.M. The genetic basic and fine-mapping of a stable quantitative-trait loci for aluminium tolerance in rice. Planta 2007, 227, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Alvim, M.N.; Ramos, F.T.; Oliveira, D.C.; Isaias, R.M.S.; Franca, M.G.C. Aluminium localization and toxicity symptoms related to root growth inhibition in rice (Oryza sativa L.) seedlings. J. Biosci. 2012, 37, 1079–1088. [Google Scholar] [CrossRef]
- Too, E.J.; Carlsson, A.S.; Onkware, A.O.; Were, B.A.; Geleta, M.; Bryngelsson, T.; Gudu, S. Cell membrane integrity, callose accumulation, and root growth in aluminum-stressed sorghum seedlings. Biol. Plant. 2014, 58, 768–772. [Google Scholar] [CrossRef]
- Radmer, L.; Tesfaye, M.; Somers, D.A.; Temple, S.J.; Vance, C.P.; Samac, D.A. Aluminum resistance mechanisms in oat (Avena sativa L.). Plant Soil 2012, 351, 121–134. [Google Scholar] [CrossRef]
- Ryan, P.R.; Delhaize, E.; Randall, P.J. Malate efflux from root apices and tolerance to aluminium are highly correlated in wheat. Funct. Plant Biol. 1995, 22, 531–536. [Google Scholar] [CrossRef]
- Ryan, P.R.; Raman, H.; Gupta, S.; Horst, W.J.; Delhaize, E. A second mechanism for aluminum resistance in wheat relies on the constitutive efflux of citrate from roots. Plant Physiol. 2009, 149, 340–351. [Google Scholar] [CrossRef] [Green Version]
- Kochian, L.V.; Pineros, M.A.; Hoekenga, O.A. The physiology, genetics and molecular biology of plant aluminum resistance and toxicity. In Root Physiology: From Gene to Function; Springer: Dordrecht, The Netherlands, 2005; pp. 175–195. [Google Scholar]
- Yang, Z.M.; Nian, H.; Sivaguru, M.; Tanakamaru, S.; Matsumoto, H. Characterization of aluminium-induced citrate secretion in aluminium-tolerant soybean (Glycine max) plants. Physiol. Plant. 2001, 113, 64–71. [Google Scholar] [CrossRef]
- Dong, D.; Peng, X.; Yan, X. Organic acid exudation induced by phosphorus deficiency and/or aluminium toxicity in two contrasting soybean genotypes. Physiol. Plant. 2004, 122, 90–99. [Google Scholar] [CrossRef]
- Eticha, D.; Zahn, M.; Bremer, M.; Yang, Z.; Rangel, A.F.; Rao, I.M.; Horst, W.J. Transcriptomic analysis reveals differential gene expression in response to aluminium in common bean (Phaseolus vulgaris) genotypes. Ann. Bot. 2010, 105, 1119–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangel, A.F.; Rao, I.M.; Braun, H.P.; Horst, W.J. Aluminum resistance in common bean (Phaseolus vulgaris) involves induction and maintenance of citrate exudation from root apices. Physiol. Plant. 2010, 138, 176–190. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, J.V.; Garvin, D.F.; Wang, Y.; Sorrells, M.E.; Klein, P.E.; Schaffert, R.E.; Li, L.; Kochian, L.V. Comparative mapping of a major aluminum tolerance gene in sorghum and other species in the Poaceae. Genetics 2004, 167, 1905–1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raman, H.; Stodart, B.; Ryan, P.R.; Delhaize, E.; Emebiri, L.; Raman, R.; Coombes, N.; Milgate, A. Genome-wide association analyses of common wheat (Triticum aestivum L.) germplasm identifies multiple loci for aluminium resistance. Genome 2010, 53, 957–966. [Google Scholar] [CrossRef]
- Wang, Q.F.; Zhao, Y.; Yi, Q.; Li, K.Z.; Yu, Y.X.; Chen, L.M. Over expression of malate dehydrogenase in transgenic tobacco leaves: Enhanced malate synthesis and augmented Al-resistance. Acta Physiol. 2010, 32, 1209–1220. [Google Scholar] [CrossRef]
- Ma, J.F.; Chen, Z.C.; Shen, R.F. Molecular mechanisms of Al resistance in gramineous plants. Plant Soil 2014, 381, 1–12. [Google Scholar] [CrossRef]
- Hoekenga, O.A.; Maron, L.G.; Piñeros, M.A.; Cançado, G.M.; Shaff, J.; Kobayashi, Y.; Ryan, P.R.; Dong, B.; Delhaize, E.; Sasaki, T.; et al. AtALMT1, which encodes a malate transporter, is identified as one of several genes critical for aluminum resistance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 9738–9743. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Magalhaes, J.V.; Shaff, J.; Kochian, L.V. Aluminum activated citrate and malate transporters from the MATE and ALMT families function independently to confer Arabidopsis aluminum tolerance. Plant J. 2009, 57, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Houde, M.; Diallo, A.O. Identification of genes and pathways associated with aluminum stress and tolerance using transcriptome profiling of wheat near-isogenic lines. BMC Genom. 2008, 9, 400. [Google Scholar] [CrossRef] [Green Version]
- You, J.; Zhang, H.; Liu, N.; Gao, L.; Kong, L.; Yang, Z. Transcriptomic responses to aluminum stress in soybean roots. Genome 2011, 54, 923–933. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Lander, E.S.; Green, P.; Abrahamson, J.; Barlow, A.; Daly, M.J.; Lincol, S.E.; Newberg, L.A. MAPMAKER: An interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics 1987, 1, 174–181. [Google Scholar] [CrossRef]
- Kosambi, D.D. The estimation of map distances from recombination values. In DD Kosambi; Springer: New Delhi, India, 2016; pp. 125–130. [Google Scholar]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Porter, A.H. An AFLP-based interspecific linkage map of sympatric Hybrid Colias Butterflies. Genetics 2004, 168, 215–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Parents | RILs (BM-4 X L-7903) | χ2 | p-Value | ||
|---|---|---|---|---|---|---|
| L-7903 (Al-Resistant) | BM-4 (Al-Sensitive) | Resistant Lines | Sensitive Lines | |||
| Malate content (nmol h−1 g−1 FW) | 10.2 | 5.4 | 8.27–12.1 | 3.85–6.23 | - | - |
| Number of plants | - | - | 82 | 64 | 2.22 | 0.136 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, C.K.; Singh, D.; Sharma, S.; Chandra, S.; Tomar, R.S.S.; Kumar, A.; Upadhyaya, K.C.; Pal, M. Mechanistic Association of Quantitative Trait Locus with Malate Secretion in Lentil (Lens culinaris Medikus) Seedlings under Aluminium Stress. Plants 2021, 10, 1541. https://doi.org/10.3390/plants10081541
Singh CK, Singh D, Sharma S, Chandra S, Tomar RSS, Kumar A, Upadhyaya KC, Pal M. Mechanistic Association of Quantitative Trait Locus with Malate Secretion in Lentil (Lens culinaris Medikus) Seedlings under Aluminium Stress. Plants. 2021; 10(8):1541. https://doi.org/10.3390/plants10081541
Chicago/Turabian StyleSingh, Chandan Kumar, Dharmendra Singh, Shristi Sharma, Shivani Chandra, Ram Sewak Singh Tomar, Arun Kumar, K. C. Upadhyaya, and Madan Pal. 2021. "Mechanistic Association of Quantitative Trait Locus with Malate Secretion in Lentil (Lens culinaris Medikus) Seedlings under Aluminium Stress" Plants 10, no. 8: 1541. https://doi.org/10.3390/plants10081541