
Validation of Reference Genes for Quantitative PCR in Johnsongrass (Sorghum halepense L.) under Glyphosate Stress

 ,
,  and
and
Abstract
:1. Introduction
2. Results
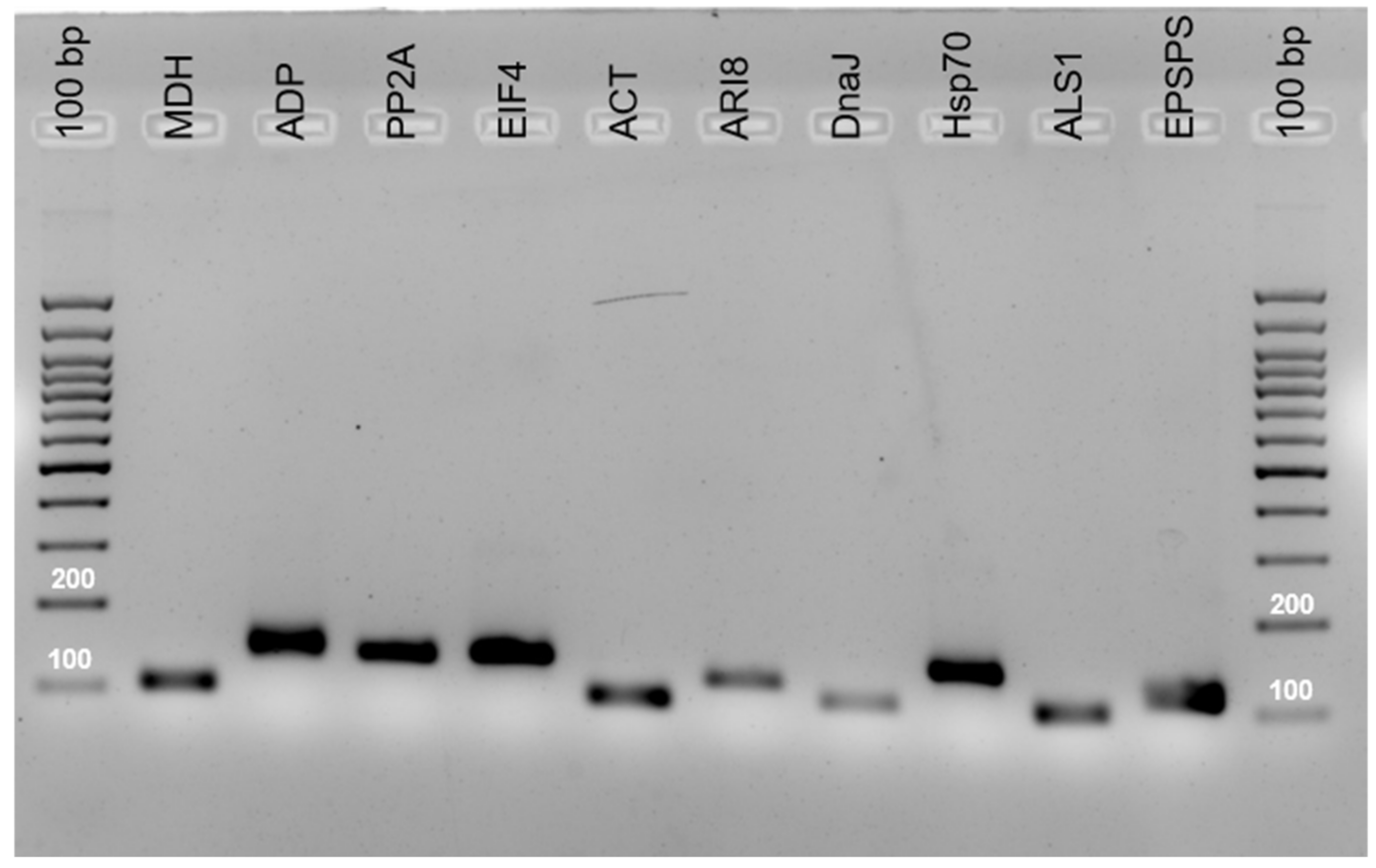
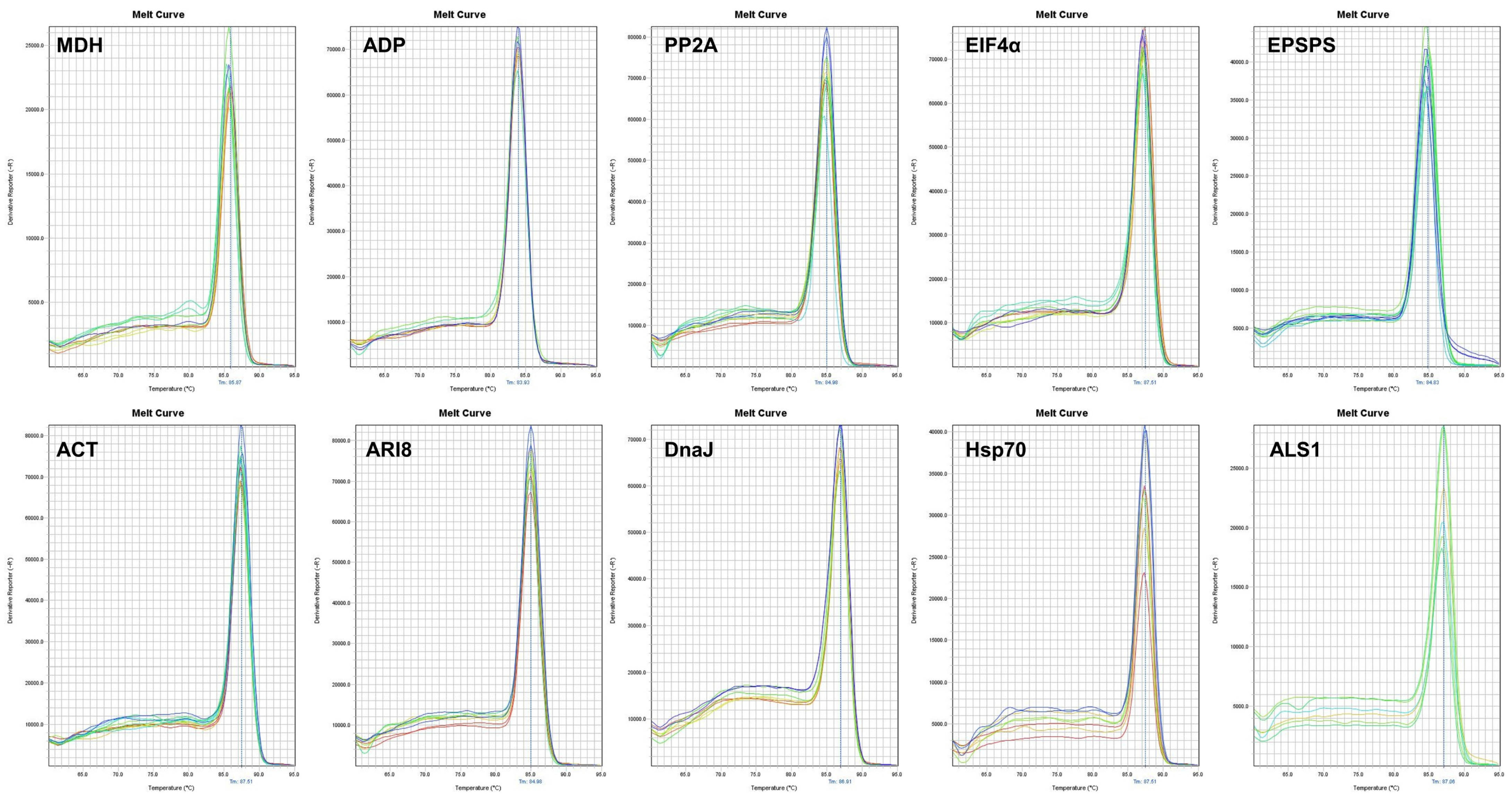
2.1. Primer Specificity and PCR Amplification Efficiency
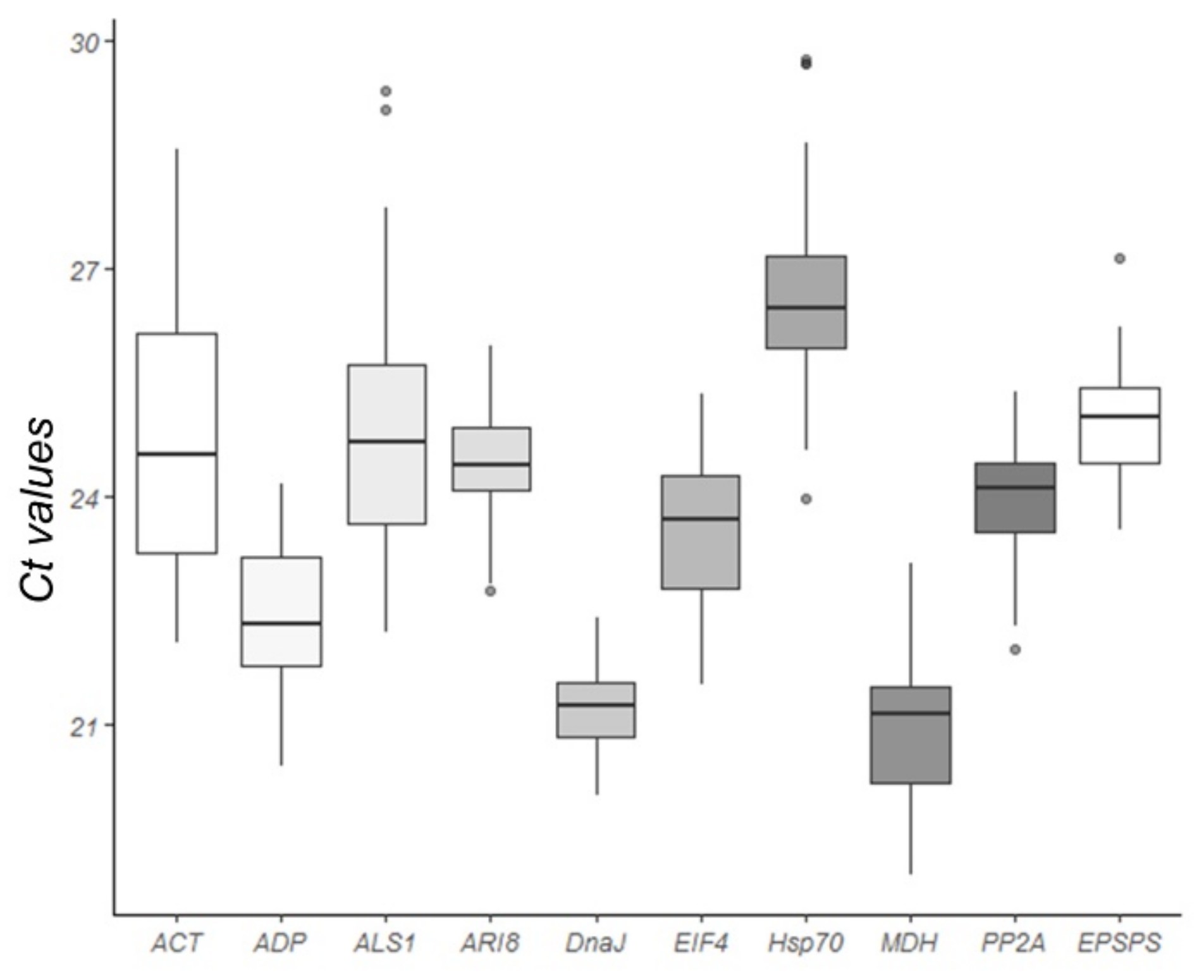
2.2. Analysis of Gene Expression Stability Using Different Software Programs
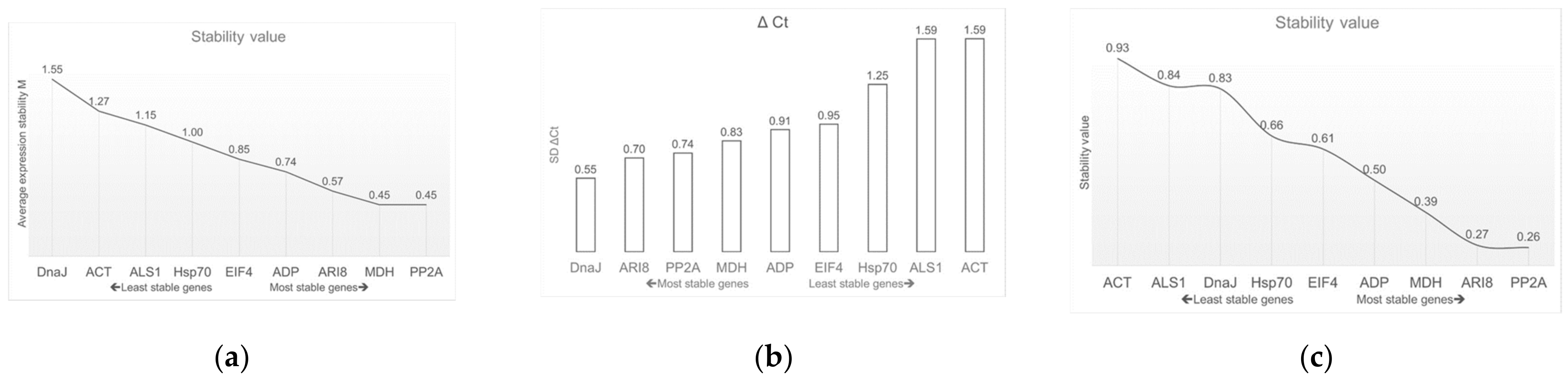
2.2.1. BestKeeper
2.2.2. geNorm
2.2.3. NormFinder
2.2.4. ΔCt Method
2.2.5. RefFinder Tool
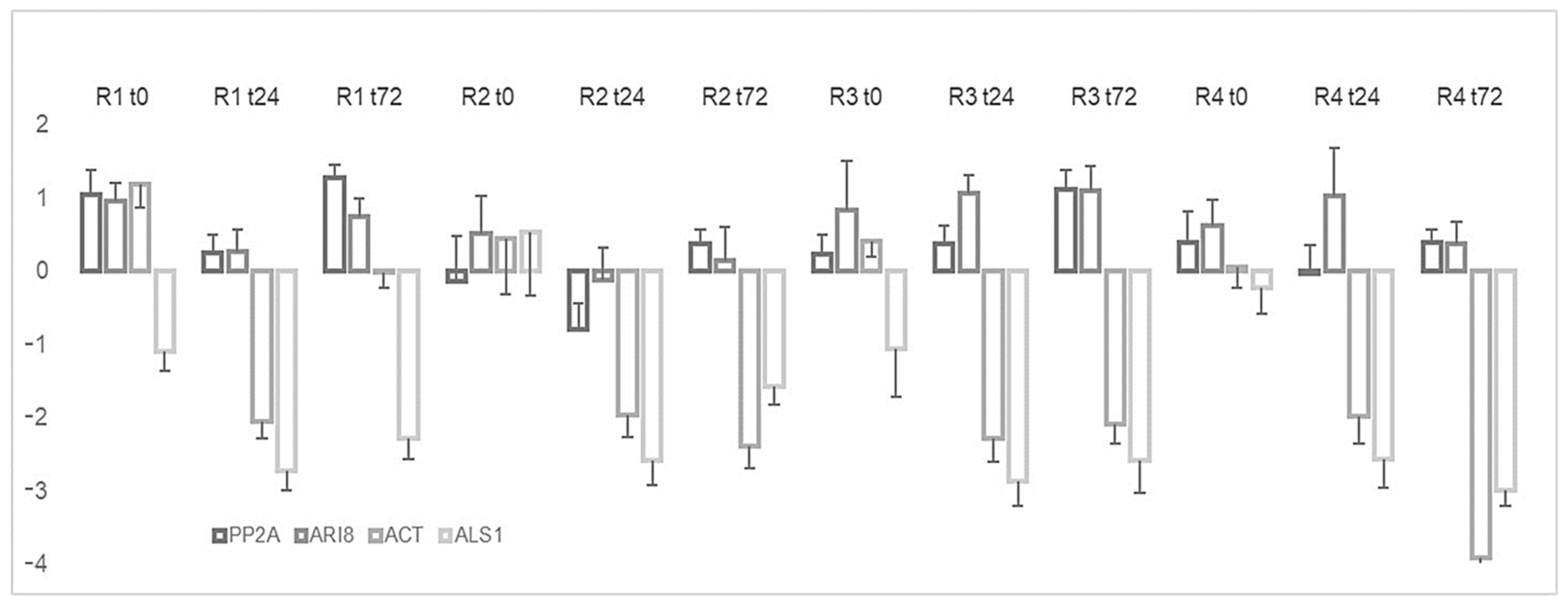
2.3. Expression Level of EPSPS
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. RNA Isolation and cDNA Synthesis
4.3. Reference Gene Selection and Primer Design
4.4. qPCR Assay
4.5. Data Analysis for Expression Stability
4.6. Validation of Reference Genes
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oerke, E.-C. Crop losses to pests. J. Agric. Sci. 2005, 144, 31–43. [Google Scholar] [CrossRef]
- Heap, I. Global perspective of herbicide-resistant weeds. Pest. Manag. Sci. 2014, 70, 1306–1315. [Google Scholar] [CrossRef] [PubMed]
- Heap, I. The International Herbicide-Resistant Weed Database. 2010. Available online: http://www.weedscience.org/Home.aspx (accessed on 26 June 2021).
- Holm, L.G.; Plucknett, D.L.; Pancho, J.V.; Herberger, J.P. The World’s Worst Weeds. Distribution and Biology; University Press of Hawaii: Honolulu, HI, USA, 1977. [Google Scholar]
- Vila-Aiub, M.M.; Balbi, M.C.; Gundel, P.E.; Ghersa, C.M.; Powles, S.B. Evolution of Glyphosate-Resistant Johnsongrass (Sorghum halepense) in Glyphosate-Resistant Soybean. Weed Sci. 2007, 55, 566–571. [Google Scholar] [CrossRef]
- Paterson, A.H.; Kong, W.; Johnston, R.; Nabukalu, P.; Wu, G.; Poehlman, W.L.; Goff, V.H.; Isaacs, K.; Lee, T.-H.; Guo, H. The Evolution of an Invasive Plant, Sorghum halepense L. (Johnsongrass). Front. Genet. 2020, 11, 1–10. [Google Scholar] [CrossRef]
- Leguizamón, E.S. Manejo de malezas problema. In Sorgo de Alepo. Bases Para su Manejo y Control en Sistemas de Producción; REM-AAPRESID: Rosario, Argentina, 2012; Volumen II. [Google Scholar]
- Aapresid. Top 10 de Las Malezas Difíciles Que Más Avanzaron. 16 January 2018. Available online: https://www.aapresid.org.ar/rem/top-10-de-las-malezas-dificiles-que-mas-avanzaron/# (accessed on 13 December 2020).
- Duke, S.O.; Powles, S.B. Glyphosate: A once-in-a-century herbicide. Pest Manag. Sci. 2008, 64, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Gaines, T.A.; Patterson, E.L.; Neve, P. Molecular mechanisms of adaptive evolution revealed by global selection for glyphosate resistance. New Phytol. 2019, 223, 1770–1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binimelis, R.; Pengue, W.; Monterroso, I. Transgenic treadmill: Responses to the emergence and spread of glyphosate-resistant johnsongrass in Argentina. Geoforum 2009, 40, 623–633. [Google Scholar] [CrossRef]
- Ustarroz, D. Informe Técnico Estación Experimental Agropecuaria Manfredi Año: 2015 Resistencia Múltiple de. 2015. Available online: http://weedscience.org/documents/showdocuments.aspx?DocumentID=1372 (accessed on 1 July 2021).
- Scursoni, J.; Morello, J.P.; Fantin, G.; Gatica, J.; Medina Herrera, D.; Pinton, S.; Seco, B.; Rodriguez, M.L. Resistencia a Herbicidas Inhibidores de ACCasa (Haloxifop Metil y Cletodim) y Glifosato en Sorgo de Alepo (Sorghum halepense). 2021. Available online: http://www.asacim.org.ar/wp-content/uploads/2021/01/Resistencia-a-herbicidas-inhibidores-de-ACCasa-en-Sorgo-de-Alepo-1-1.pdf (accessed on 1 July 2021).
- Vazquez-Garcia, J.G.; Palma-Bautista, C.; Rojano-Delgado, A.M.; De Prado, R.; Menendez, J. The First Case of Glyphosate Resistance in Johnsongrass (Sorghum halepense (L.) Pers.) in Europe. Plants 2020, 9, 313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palau, H.; Senesi, S.; Mogni, L.; Ordóñez, I. Impacto Económico Macro y Micro de Malezas Resistentes en el Agro Argentino; ADAMA-FAUBA: Ciudad Autónoma de Buenos Aires, Argentina, 2015. [Google Scholar]
- Délye, C.; Jasieniuk, M.; Le Corre, V. Deciphering the evolution of herbicide resistance in weeds. Trends Genet. 2013, 29, 649–658. [Google Scholar] [CrossRef]
- Gaines, T.A.; Duke, S.O.; Morran, S.; Rigon, C.A.G.; Tranel, P.J.; Küpper, A.; Dayan, F.E. Mechanisms of evolved herbicide resistance. J. Biol. Chem. 2020, 295, 10307–10330. [Google Scholar] [CrossRef]
- Chauhan, B.S. Grand Challenges in Weed Management. Front. Agron. 2020, 1, 1–4. [Google Scholar] [CrossRef]
- Délye, C. Unravelling the genetic bases of non-target-site-based resistance (NTSR) to herbicides: A major challenge for weed science in the forthcoming decade. Pest Manag. Sci. 2012, 69, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Tani, E.; Chachalis, D.; Travlos, I.S. A Glyphosate Resistance Mechanism in Conyza canadensis Involves Synchronization of EPSPS and ABC-transporter Genes. Plant Mol. Biol. Rep. 2015, 33, 1721–1730. [Google Scholar] [CrossRef]
- Chen, J.; Huang, Z.; Huang, H.; Wei, S.; Liu, Y.; Jiang, C.; Zhang, J.; Zhang, C. Selection of relatively exact reference genes for gene expression studies in goosegrass (Eleusine indica) under herbicide stress. Sci. Rep. 2017, 7, 46494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, P.E.; Reddy, D.E.; Sivasakthi, K.; Ebhatnagar-Mathur, P.; Evadez, V.; Sharma, K.K. Evaluation of Sorghum [Sorghum bicolor (L.)] Reference Genes in Various Tissues and under Abiotic Stress Conditions for Quantitative Real-Time PCR Data Normalization. Front. Plant Sci. 2016, 7, 529. [Google Scholar] [CrossRef]
- De Spiegelaere, W.; Dern-Wieloch, J.; Weigel, R.; Schumacher, V.; Schorle, H.; Nettersheim, D.; Bergmann, M.; Brehm, R.; Kliesch, S.; Vandekerckhove, L. Reference Gene Validation for RT-qPCR, a Note on Different Available Software Packages. PLoS ONE 2015, 10, e0122515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.; Shipley, G.L. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Sundaram, V.K.; Sampathkumar, N.K.; Massaad, C.; Grenier, J. Optimal use of statistical methods to validate reference gene stability in longitudinal studies. PLoS ONE 2019, 14, e0219440. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Tian, C.; Wang, Y.; Wan, F.; Hu, L.; Xiong, A.; Tian, J. Selection of reliable reference genes for quantitative RT-PCR in garlic under salt stress. PeerJ 2019, 7, e7319. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Xu, H.; Cao, Y.; Yang, P.; Feng, Y.; Tang, Y.; Yuan, S.; Ming, J. Validation of Reference Genes for Quantitative Real-Time PCR during Bicolor Tepal Development in Asiatic Hybrid Lilies (Lilium spp.). Front. Plant Sci. 2017, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Liu, X.; Chen, S.; Li, B.; Wang, X.; Fan, C.; Wang, G.; Ni, H. Selection of relatively exact reference genes for gene expression studies in flixweed (Descurainia sophia) by quantitative real-time polymerase chain reaction. Pestic. Biochem. Physiol. 2016, 127, 59–66. [Google Scholar] [CrossRef]
- Zhang, Y.; Han, X.; Chen, S.; Zheng, L.; He, X.; Liu, M.; Qiao, G.; Wang, Y.; Zhuo, R. Selection of suitable reference genes for quantitative real-time PCR gene expression analysis in Salix matsudana under different abiotic stresses. Sci. Rep. 2017, 7, 40290. [Google Scholar] [CrossRef] [Green Version]
- Petit, C.; Pernin, F.; Heydel, J.-M.; Délye, C. Validation of a set of reference genes to study response to herbicide stress in grasses. BMC Res. Notes 2012, 5, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrzesińska, B.; Kierzek, R.; Obrępalska-Stęplowska, A. Evaluation of six commonly used reference genes for gene expression studies in herbicide-resistantAvena fatuabiotypes. Weed Res. 2016, 56, 284–292. [Google Scholar] [CrossRef]
- Moretti, M.L.; Alarcón-Reverte, R.; Pearce, S.; Morran, S.; Hanson, B.D. Transcription of putative tonoplast transporters in response to glyphosate and paraquat stress in Conyza bonariensis and Conyza canadensis and selection of reference genes for qRT-PCR. PLoS ONE 2017, 12, e0180794. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Wang, X.; Chen, J.; Huang, Z.; Huo, H.; Jiang, C.; Huang, H.; Zhang, C.; Wei, S. Selection of reference genes for qPCR normalization in buffalobur (Solanum rostratum Dunal). Sci. Rep. 2019, 9, 6948. [Google Scholar] [CrossRef] [Green Version]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Gao, Y.-Y.; Huang, Y.-Q.; Fan, Q.; Lu, X.-T.; Wang, C.-K. Selection of housekeeping genes for quantitative gene expression analysis in yellow-feathered broilers. Ital. J. Anim. Sci. 2017, 17, 540–546. [Google Scholar] [CrossRef] [Green Version]
- Guénin, S.; Mauriat, M.; Pelloux, J.; Van Wuytswinkel, O.; Bellini, C.; Gutierrez, L. Normalization of qRT-PCR data: The necessity of adopting a systematic, experimental conditions-specific, validation of references. J. Exp. Bot. 2009, 60, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S. The continuing problem of poor transparency of reporting and use of inappropriate methods for RT-qPCR. Biomol. Detect. Quantif. 2017, 12, 7–9. [Google Scholar] [CrossRef]
- Su, X.; Lu, L.; Li, Y.; Zhen, C.; Hu, G.; Jiang, K.; Yan, Y.; Xu, Y.; Wang, G.; Shi, M. Reference gene selection for quantitative real-time PCR (qRT-PCR) expression analysis in Galium aparine L. PLoS ONE 2020, 15, e0226668. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, Y.; Fang, H.; Shi, H.; Chen, K.; Zhang, Z.; Tan, X. Selection of reference genes for quantitative reverse-transcription polymerase chain reaction normalization in Brassica napus under various stress conditions. Mol. Genet. Genom. 2014, 289, 1023–1035. [Google Scholar] [CrossRef] [PubMed]
- Yanniccari, M.; Gómez-Lobato, M.E.; Istilart, C.; Natalucci, C.; Giménez, D.O.; Castro, A.M. Mechanism of Resistance to Glyphosate in Lolium perenne from Argentina. Front. Ecol. Evol. 2017, 5, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Schneider, T.; Rizzardi, M.A.; Grando, M.F.; Ceccon, C.C.; Nunes, A.L.; DiDone, D. Expression of genes related to the mechanism of resistance of Conyza sumatrensis to glyphosate. Acta Sci. Agron. 2020, 43, e44824. [Google Scholar] [CrossRef]
- Nandula, V.K.; Ray, J.D.; Ribeiro, D.N.; Pan, Z.; Reddy, K. Glyphosate Resistance in Tall Waterhemp (Amaranthus tuberculatus) from Mississippi is due to both Altered Target-Site and Nontarget-Site Mechanisms. Weed Sci. 2013, 61, 374–383. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of Real-Time Quantitative Reverse Transcription-PCR Data: A Model-Based Variance Estimation Approach to Identify Genes Suited for Normalization, Applied to Bladder and Colon Cancer Data Sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Ruijter, J.M.; Ramakers, C.; Hoogaars, W.M.H.; Karlen, Y.; Bakker, O.; Hoff, M.J.B.V.D.; Moorman, A.F.M. Amplification efficiency: Linking baseline and bias in the analysis of quantitative PCR data. Nucleic Acids Res. 2009, 37, e45. [Google Scholar] [CrossRef] [Green Version]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat Versión 2011. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. Available online: http://www.infostat.com.ar (accessed on 1 June 2021).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Cellular Function | Accession Number | Primers (F/R) (5″–3″) | Amplicon Length (bp) | Standard Curve Parameters | |||
|---|---|---|---|---|---|---|---|---|
| Slope | R2 | Eff% | E | |||||
| MDH | Malate dehydrogenase | XM_002467034 | TGCAGTGGTGGTGAATGGAA | 103 | −3.277 | 0.985 | 104.133 | 2.019 |
| GCGTCTTCTCTTCCGACAGC | ||||||||
| ADP | ADP-Ribosylation Factor | XM_002441244 | GTCTGTCGGATGTGGGGATGT | 136 | −3.287 | 0.996 | 101.179 | 2.015 |
| CACAGCACACAGTCGGACATG | ||||||||
| PP2A | Serine/threonine Protein Phosphatase | XM_002453490 | AACCCGCAAAACCCCAGACTA | 138 | −3.188 | 0.998 | 105.919 | 2.059 |
| TACAGGTCGGGCTCATGGAAC | ||||||||
| EIF4α | Eukaryotic Initiation Factor 4A | XM_002451491 | CAACTTTGTCACCCGCGATGA | 144 | −3.316 | 0.995 | 100.265 | 2.002 |
| TCCAGAAACCTTAGCAGCCCA | ||||||||
| ACT | Actin | Sobic.009G005900.1 | TCGAGACACTTGTGGCAGATT | 100 | −3.396 | 0.997 | 97.003 | 1.970 |
| CGCACATGGAGCCACAACAT | ||||||||
| ARI8 | E3 ubiquitin protein ligase ARI8 | Sobic.006G131000.2 | CGGGCTCTGGAAACTGGATT | 121 | −3.327 | 0.999 | 99.772 | 1.998 |
| TTGATGCCCTGTTCTTGCCA | ||||||||
| DnaJ | Chaperone protein DnaJ 49 | Sobic.003G185200.1 | TTTCAGGACTGGTGGGATGC | 103 | −3.363 | 1 | 98.3 | 1.983 |
| GAGCAACAGCAGCAGTAGGA | ||||||||
| Hsp70 | Heat shock 70 kDa protein | Sobic.002G249800.1 | ACCTGCTGAAGTCACCAAGG | 150 | −3.178 | 0.996 | 106.392 | 2.064 |
| CCACCACCTTGTTGCATGTG | ||||||||
| ALS1 | Acetolactate Synthase | Sobic.004G155800.2 | TGGGCCTTGGCAATTTCC | 100 | −3.168 | 0.934 | 106.827 | 2.089 |
| AGATCCGCCTTATCCACTGCAT | ||||||||
| EPSPS | 5-enolpyruvylshikimate-3-phosphate synthase | Sobic.010G023800.1 | CATGGACCGAGACTAGCGTAACTG | 113 | −3.309 | 0.979 | 100.538 | 2.005 |
| AGTCATGGCAACATCAGGCATT | ||||||||
| Rank | BestKeeper | geNorm | NormFinder | ΔCt Method | RefFinder | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene | SD (±Cq) | CV (% Cq) | Gene | Stability | Gene | Stability | Gene | Stability | Gene | Stability | |
| 1 | DnaJ | 0.43 | 2.04 | PP2A | 0.45 | PP2A | 0.26 | DnaJ | 0.55 | PP2A | 1.32 |
| 2 | ARI8 | 0.55 | 2.26 | MDH | 0.45 | ARI8 | 0.27 | ARI8 | 0.70 | ARI8 | 2.21 |
| 3 | PP2A | 0.60 | 2.53 | ARI8 | 0.57 | MDH | 0.39 | PP2A | 0.74 | DnaJ | 2.63 |
| 4 | MDH | 0.69 | 3.28 | ADP | 0.74 | ADP | 0.50 | MDH | 0.83 | MDH | 2.63 |
| 5 | ADP | 0.75 | 3.34 | EIF4 | 0.85 | EIF4 | 0.61 | ADP | 0.91 | ADP | 5.00 |
| 6 | EIF4 | 0.82 | 3.47 | Hsp70 | 1.00 | Hsp70 | 0.66 | EIF4 | 0.95 | EIF4 | 6.00 |
| 7 | Hsp70 | 0.93 | 3.50 | ALS1 | 1.15 | DnaJ | 0.83 | Hsp70 | 1.25 | Hsp70 | 7.00 |
| 8 | ALS1 | 1.23 | 4.96 | ACT | 1.27 | ALS1 | 0.84 | ALS1 | 1.59 | ALS1 | 8.00 |
| 9 | ACT | 1.36 | 5.49 | DnaJ | 1.55 | ACT | 0.93 | ACT | 1.59 | ACT | 9.00 |
| Rank | BestKeeper | geNorm | NormFinder | ΔCt Method | RefFinder | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gene | SD (±Cq) | Gene | Stability | Gene | Stability | Gene | Stability | Gene | Stability | |
| 1 | DnaJ | 0.36 | PP2A | 0.49 | PP2A | 0.22 | PP2A | 0.94 | PP2A | 1.32 |
| 2 | ARI8 | 0.48 | MDH | 0.49 | ARI8 | 0.35 | ARI8 | 1.00 | ARI8 | 2.21 |
| 3 | PP2A | 0.56 | ARI8 | 0.59 | MDH | 0.50 | DnaJ | 1.01 | DnaJ | 2.63 |
| 4 | MDH | 0.64 | DnaJ | 0.64 | DnaJ | 0.51 | MDH | 1.04 | MDH | 2.63 |
| 5 | ADP | 0.66 | ADP | 0.73 | ADP | 0.82 | ADP | 1.17 | ADP | 5.00 |
| 6 | EIF4 | 0.69 | EIF4 | 0.80 | EIF4 | 0.95 | EIF4 | 1.23 | EIF4 | 6.00 |
| 7 | Hsp70 | 0.89 | Hsp70 | 0.95 | Hsp70 | 1.28 | Hsp70 | 1.51 | Hsp70 | 7.00 |
| 8 | ALS1 | 1.16 | ALS1 | 1.11 | ALS1 | 1.31 | ALS1 | 1.54 | ALS1 | 8.00 |
| 9 | ACT | 1.35 | ACT | 1.2 | ACT | 1.46 | ACT | 1.64 | ACT | 9.00 |
| ID | Collection Site | Latitude | Longitude |
|---|---|---|---|
| S1 | Oro Verde, Entre Ríos | 31°49′58″ S | 60°31′27″W |
| S2 | Facultad; Entre Ríos | 31°49′59″ S | 60°31′ 28″ W |
| R1 | Soresi; Entre Ríos | 31°19′56″ S | 60°01′16″ W |
| R2 | Pavioti; Entre Ríos | 31°18′48″ S | 59°46′30″ W |
| R3 | Hasenkamp; Entre Ríos | 31°27′20″ S | 59°52′54″ W |
| R4 | Hernandarias; Entre Ríos | 31°17′11″ S | 59°46′42″ W |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ulrich, M.N.; Muñiz-Padilla, E.; Corach, A.; Hopp, E.; Tosto, D. Validation of Reference Genes for Quantitative PCR in Johnsongrass (Sorghum halepense L.) under Glyphosate Stress. Plants 2021, 10, 1555. https://doi.org/10.3390/plants10081555
Ulrich MN, Muñiz-Padilla E, Corach A, Hopp E, Tosto D. Validation of Reference Genes for Quantitative PCR in Johnsongrass (Sorghum halepense L.) under Glyphosate Stress. Plants. 2021; 10(8):1555. https://doi.org/10.3390/plants10081555
Chicago/Turabian StyleUlrich, María Noelia, Esteban Muñiz-Padilla, Alejandra Corach, Esteban Hopp, and Daniela Tosto. 2021. "Validation of Reference Genes for Quantitative PCR in Johnsongrass (Sorghum halepense L.) under Glyphosate Stress" Plants 10, no. 8: 1555. https://doi.org/10.3390/plants10081555
APA StyleUlrich, M. N., Muñiz-Padilla, E., Corach, A., Hopp, E., & Tosto, D. (2021). Validation of Reference Genes for Quantitative PCR in Johnsongrass (Sorghum halepense L.) under Glyphosate Stress. Plants, 10(8), 1555. https://doi.org/10.3390/plants10081555