
Auxin and Cytokinin Interplay during Leaf Morphogenesis and Phyllotaxy

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. General Leaf Development Mechanism
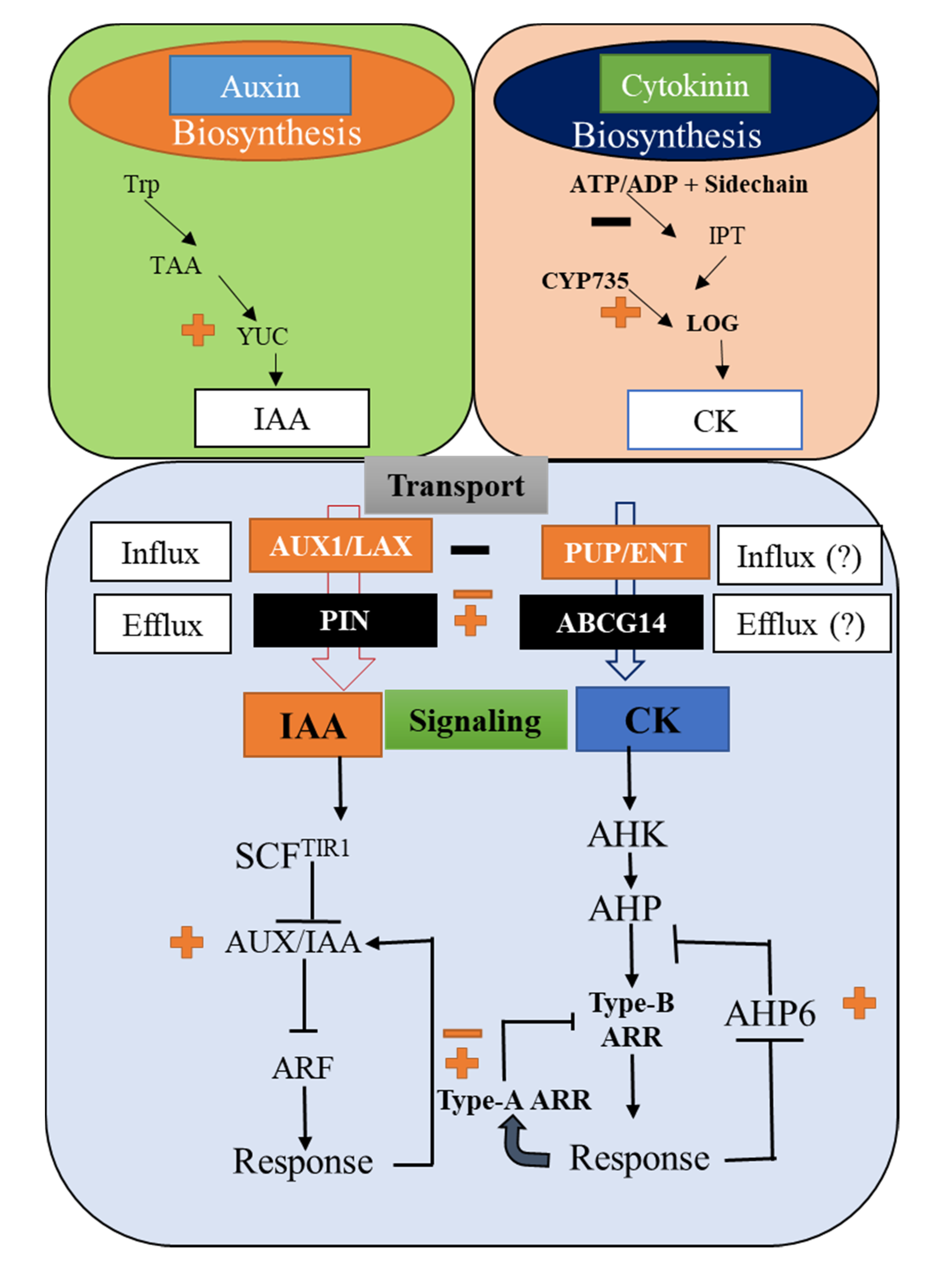
3. Auxin and Cytokinin Biosynthesis: Convergence and Divergence
3.1. Auxin Biosynthesis in Plant
3.2. Cytokinin Biosynthesis in Plant
4. Effects of Auxins and CKs in Regulating Their Signaling Pathways: An Inspection
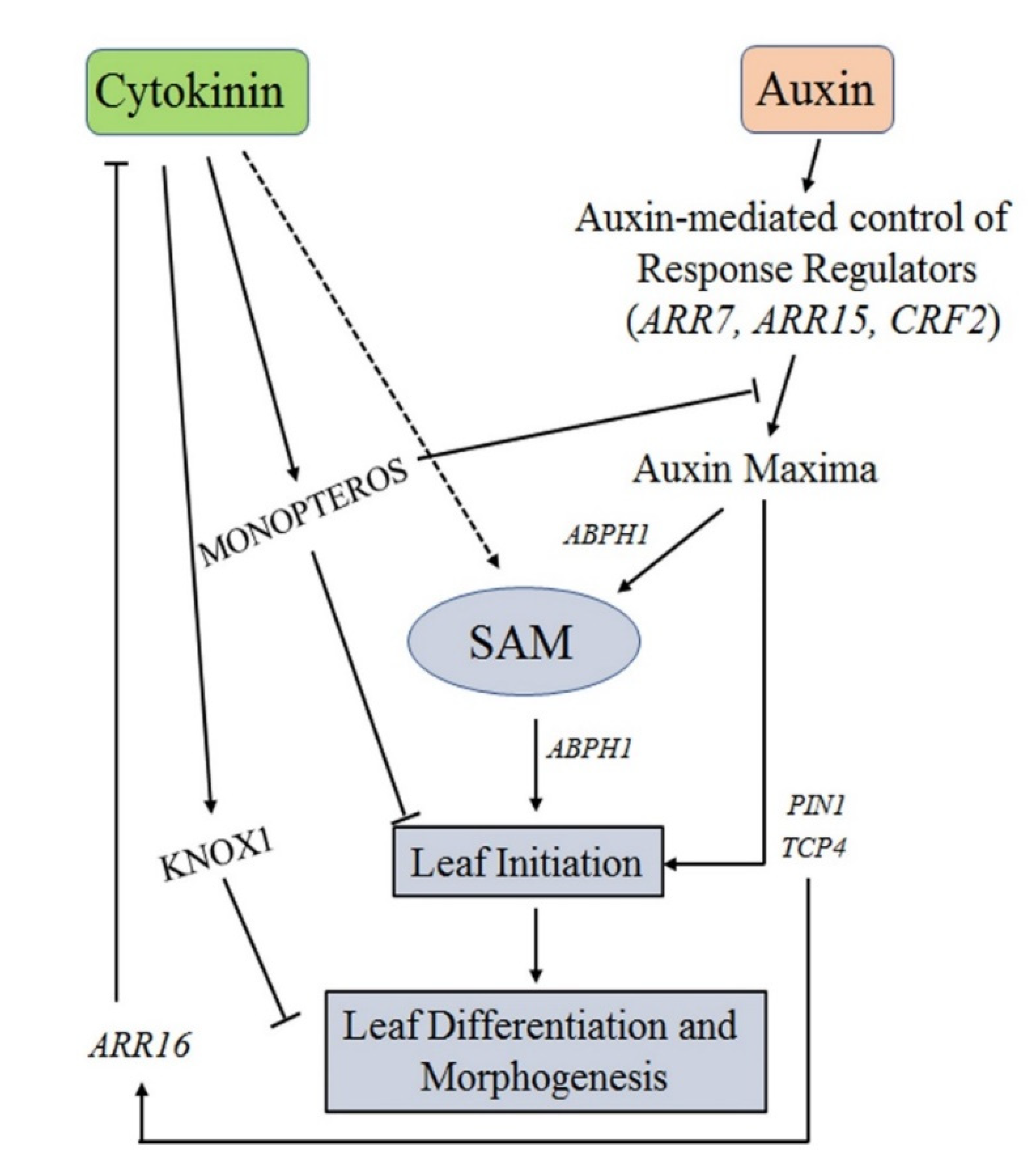
5. Role of Auxins-CKs Interactions on Leaf Development
6. Effects of Auxin-Cytokinin Interplay in Phyllotaxy
7. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ANTHRANILATE SYNTHASE α1 | ASA1 |
| ARABIDOPSIS HISTIDINE KINASE 2 | AHK2 |
| ARABIDOPSIS HISTIDINE PHOSPHOTRANSFERASE PROTEINS | AHP |
| ARABIDOPSIS RESPONSE REGULATOR 7 | ARR7 |
| AUXIN RESPONSE FACTOR 19 | ARF19 |
| AUXIN/LIKE AUX | AUX/LAX |
| AUXIN RESISTANT 1, 3 | AXR1, AXR3 |
| Chromatin Immunoprecipitation | CHIP |
| class 1 KNOTTEDLIKE HOMEOBOX | KNOX1 |
| CYTOCHROME P450 MONOOXYGENASE, FAMILY 735, SUBFAMILY A | CYP735A |
| CYTOKININ OXIDASE/DEHYDROGENASE | CKX |
| CYTOKININ RESPONSE FACTOR 2 | CRF2 |
| Cytokinins | CKs |
| Gibberellin | GA |
| GRETCHEN HAGEN 3.17, 3.9 | GH3.17, H3.9 |
| Ground meristem | GM |
| INDOLE-3-GLYCEROL PHOSPHATE SYNTHASE | IGPS |
| Indole-3-pyruvate acid | IPA |
| Indole-3-acetic acid | IAA |
| ISOPENTENYLTRANSFERASE | IPT |
| LONELY GUY | LOG |
| MONOPTEROS | MP |
| 1-naphthylphthalamic acid | NPA |
| N6-isopentenyladenine | iP |
| Naphthaleneacetic acid | NAA |
| NITRILASE3 | NIT3 |
| PHOSPHORIBOSYLANTHRANILATE TRANSFERASE1 | PAT1 |
| PIN-FORMED | PIN |
| Shoot apical meristem | SAM |
| SHORT HYPOCOTYL2/Indole-3-acetic acid3 | SHY2/IAA3 |
| trans-zeatin | tZ |
| tryptophan | Trp |
| TRYPTOPHAN AMINOTRANFERASE OF ARABIDOPSIS | TAA |
| YUCCA | YUC |
References
- Roth-Nebelsick, A.; Uhl, D.; Mosbrugger, V.; Kerp, H. Evolution and Function of Leaf Venation Architecture: A Review. Ann. Bot. 2001, 87, 553–566. [Google Scholar] [CrossRef]
- Schaller, G.E.; Bishopp, A.; Kieber, J.J. The Yin-Yang of Hormones: Cytokinin and Auxin Interactions in Plant Development. Plant Cell 2015, 27, 44–63. [Google Scholar] [CrossRef] [Green Version]
- Danilova, M.N.; Doroshenko, A.S.; Kudryakova, N.V.; Klepikova, A.V.; Shtratnikova, V.Y.; Kusnetsov, V.V. The Crosstalk Between Cytokinin and Auxin Signaling Pathways in the Control of Natural Senescence of Arabidopsis thaliana Leaves. Russ. J. Plant Physiol. 2020, 67, 1028–1035. [Google Scholar] [CrossRef]
- Zhao, Z.; Andersen, S.U.; Ljung, K.; Dolezal, K.; Miotk, A.; Schultheiss, S.J.; Lohmann, J.U. Hormonal Control of the Shoot Stem-Cell Niche. Nat. Cell Biol. 2010, 465, 1089–1092. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liu, G.; Liu, J.; Zhang, B.; Meng, W.; Müller, B.; Hayashi, K.; Zhang, X.; Zhao, Z.; De Smet, I.; et al. Synergistic Action of Auxin and Cytokinin Mediates aluminum-induced Root Growth Inhibition in Arabidopsis. EMBO Rep. 2017, 18, 1213–1230. [Google Scholar] [CrossRef]
- Hurný, A.; Cuesta, C.; Cavallari, N.; Ötvös, K.; Duclercq, J.; Dokládal, L.; Montesinos, J.C.; Gallemí, M.; Semerádová, H.; Rauter, T.; et al. Synergistic on Auxin and Cytokinin 1 Positively Regulates Growth and Attenuates Soil Pathogen Resistance. Nat. Commun. 2020, 11, 2170. [Google Scholar] [CrossRef] [PubMed]
- Schlereth, A.; Möller, B.; Liu, W.; Kientz, M.; Flipse, J.; Rademacher, E.H.; Schmid, M.; Jürgens, G.; Weijers, D. Monopteros Controls Embryonic Root Initiation by Regulating a Mobile Transcription Factor. Nat. Cell Biol. 2010, 464, 913–916. [Google Scholar] [CrossRef]
- Ruzicka, K.; Simaskova, M.; Duclercq, J.; Petrasek, J.; Zazimalova, E.; Simon, S.; Friml, J.; Van Montagu, M.C.E.; Benkova, E. Cytokinin Regulates Root Meristem Activity via Modulation of the Polar Auxin Transport. Proc. Natl. Acad. Sci. USA 2009, 106, 4284–4289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šimášková, M.; O’Brien, J.A.; Khan-Djamei, M.; Van Noorden, G.; Ötvös, K.; Vieten, A.; De Clercq, I.; Van Haperen, J.M.A.; Cuesta, C.; Hoyerova, K.; et al. Cytokinin Response Factors Regulate PIN-FORMED Auxin Transporters. Nat. Commun. 2015, 6, 8717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurný, A.; Benková, E. Methodological Advances in Auxin and Cytokinin Biology; Springer Science and Business Media LLC: Berlin, Germany, 2017; Volume 1569, pp. 1–29. [Google Scholar] [CrossRef]
- Lee, Z.H.; Hirakawa, T.; Yamaguchi, N.; Ito, T. The Roles of Plant Hormones and Their Interactions with Regulatory Genes in Determining Meristem Activity. Int. J. Mol. Sci. 2019, 20, 4065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; Jiao, Y. The Diverse Roles of Auxin in Regulating Leaf Development. Plants 2019, 8, 243. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.-H.; Liu, Y.-B.; Zhang, X.-S. Auxin–Cytokinin Interaction Regulates Meristem Development. Mol. Plant 2011, 4, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Sablowski, R. The Dynamic Plant Stem Cell Niches. Curr. Opin. Plant Biol. 2007, 10, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.V. Live-Imaging Stem-Cell Homeostasis in the Arabidopsis Shoot Apex. Curr. Opin. Plant Biol. 2008, 11, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Skalák, J.; Vercruyssen, L.; Claeys, H.; Hradilová, J.; Cerny, M.; Novák, O.; Plačková, L.; Saiz-Fernández, I.; Skaláková, P.; Coppens, F.; et al. Multifaceted Activity of Cytokinin in Leaf Development Shapes Its Size and Structure in Arabidopsis. Plant J. 2019, 97, 805–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarpella, E.; Meijer, A.H. Pattern Formation in the Vascular System of Monocot and Dicot Plant Species. New Phytol. 2004, 164, 209–242. [Google Scholar] [CrossRef] [PubMed]
- Tsukaya, H. Leaf Development. Arab. Book 2002, 1, e0072. [Google Scholar] [CrossRef]
- Kang, J.; Dengler, N. Vein Pattern Development in Adult Leaves of Arabidopsis thaliana. Int. J. Plant Sci. 2004, 165, 231–242. [Google Scholar] [CrossRef]
- Donnelly, P.M.; Bonettaa, D.; Tsukaya, H.; Dengler, R.E.; Dengler, N.G. Cell Cycling and Cell Enlargement in Developing Leaves of Arabidopsis. Dev. Biol. 1999, 215, 407–419. [Google Scholar] [CrossRef] [Green Version]
- Baima, S.; Possenti, M.; Matteucci, A.; Wisman, E.; Altamura, M.M.; Ruberti, I.; Morelli, G. The Arabidopsis ATHB-8 HD-Zip Protein Acts As a Differentiation-Promoting Transcription Factor of the Vascular Meristems. Plant Physiol. 2001, 126, 643–655. [Google Scholar] [CrossRef] [Green Version]
- Candela, H.; Martínez-Laborda, A.; Micol, J.L. Venation Pattern Formation in Arabidopsis thaliana Vegetative Leaves. Dev. Biol. 1999, 205, 205–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y. Auxin Biosynthesis. The Arabidopsis Book. Am. Soc. Plant 2014, 12, e0173. [Google Scholar]
- Zhao, Y. Auxin Bionsynthesis and Its Role in Plant Development. Annu. Rev. Plant Biol. 2010, 2, 49–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, Y.; Men, S.; Fischer, U.; Stepanova, A.; Alonso, J.; Ljung, K.; Grebe, M. Local Auxin Biosynthesis Modulates Gradient-Directed Planar Polarity in Arabidopsis. Nat. Cell Biol. 2009, 11, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Ferrer, J.-L.; Ljung, K.; Pojer, F.; Hong, F.; Long, J.A.; Li, L.; Moreno, J.; Bowman, M.E.; Ivans, L.J.; et al. Rapid Synthesis of Auxin via a New Tryptophan-Dependent Pathway Is Required for Shade Avoidance in Plants. Cell 2008, 133, 164–176. [Google Scholar] [CrossRef] [Green Version]
- Frébort, I.; Kowalska, M.; Hluska, T.; Frébortová, J.; Galuszka, P. Evolution of Cytokinin Biosynthesis and Degradation. J. Exp. Bot. 2011, 62, 2431–2452. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, H. CYTOKININS: Activity, Biosynthesis, and Translocation. Annu. Rev. Plant Biol. 2006, 57, 431–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takei, K.; Yamaya, T.; Sakakibara, H. Arabidopsis CYP735A1 and CYP735A2 Encode Cytokinin Hydroxylases That Catalyze the Biosynthesis of Trans-Zeatin. J. Biol. Chem. 2004, 279, 41866–41872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroha, T.; Tokunaga, H.; Kojima, M.; Ueda, N.; Ishida, T.; Nagawa, S.; Fukuda, H.; Sugimoto, K.; Sakakibara, H. Functional Analyses of LONELY GUY Cytokinin-Activating Enzymes Reveal the Importance of the Direct Activation Pathway in Ara-Bidopsis. Plant Cell 2009, 21, 3152–3169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajguz, A.; Piotrowska, A. Conjugates of Auxin and Cytokinin. Phytochemistry 2009, 70, 957–969. [Google Scholar] [CrossRef]
- Hou, B.; Lim, E.-K.; Higgins, G.; Bowles, D.J. N-Glucosylation of Cytokinins by Glycosyltransferases of Arabidopsis thaliana. J. Biol. Chem. 2004, 279, 47822–47832. [Google Scholar] [CrossRef] [Green Version]
- Mok, D.W.; Mok, M.C. Cytokinin Metabolism and Action. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 89–118. [Google Scholar] [CrossRef]
- Schmülling, T.; Werner, T.; Riefler, M.; Krupková, E.; Bartrina y Manns, I. Structure and Function of Cytokinin Oxidase/De-Hydrogenase Genes of Maize, Rice, Arabidopsis and Other Species. J. Plant Res. 2003, 116, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Werner, T.; Köllmer, I.; Bartrina, I.; Holst, K.; Schmülling, T. New Insights into the Biology of Cytokinin Degradation. Plant Biol. 2006, 8, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, M.; Galuszka, P.; Frébortová, J.; Šebela, M.; Béres, T.; Hluska, T.; Šmehilová, M.; Bilyeu, K.D.; Frébort, I. Vacuolar and Cytosolic Cytokinin Dehydrogenases of Arabidopsis thaliana: Heterologous Expression, Purification and Properties. Phytochemistry 2010, 71, 1970–1978. [Google Scholar] [CrossRef]
- Gao, S.; Fang, J.; Xu, F.; Wang, W.; Sun, X.; Chu, J.; Cai, B.; Feng, Y.-Q.; Chu, C. Cytokinin Oxidase/Dehydrogenase4 Integrates Cytokinin and Auxin Signaling to Control Rice Crown Root Formation. Plant Physiol. 2014, 165, 1035–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, L.; Jiao, F.; Chu, J.; Jin, G.; Chen, M.; Wu, P. The Two-Component Signal System in Rice (Oryza Sativa L.): A Genome-Wide Study of Cytokinin Signal Perception and Transduction. Genomics 2007, 89, 697–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pils, B.; Heyl, A. Unraveling the Evolution of Cytokinin Signaling. Plant Physiol. 2009, 151, 782–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higuchi, M.; Pischke, M.S.; Mähönen, A.P.; Miyawaki, K.; Hashimoto, Y.; Seki, M.; Kobayashi, M.; Shinozaki, K.; Kato, T.; Tabata, S.; et al. In Planta Functions of the Arabidopsis Cytokinin Receptor Family. Proc. Natl. Acad. Sci. USA 2004, 101, 8821–8826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, C.; Ohashi, Y.; Sato, S.; Kato, T.; Tabata, S.; Ueguchi, C. Histidine Kinase Homologs That Act As Cytokinin Receptors Possess Overlapping Functions in the Regulation of Shoot and Root Growth in Arabidopsis. Plant Cell 2004, 16, 1365–1377. [Google Scholar] [CrossRef] [Green Version]
- Caesar, K.; Thamm, A.M.K.; Witthöft, J.; Elgass, K.; Huppenberger, P.; Grefen, C.; Horak, J.; Harter, K. Evidence for the Lo-Calization of the Arabidopsis Cytokinin Receptors AHK3 and AHK4 in the Endoplasmic Reticulum. J. Exp. Bot. 2011, 62, 5571–5580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wulfetange, K.; Lomin, S.N.; Romanov, G.A.; Stolz, A.; Heyl, A.; Schmülling, T. The Cytokinin Receptors of Arabidopsis Are Located Mainly to the Endoplasmic Reticulum. Plant Physiol. 2011, 156, 1808–1818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirose, N.; Takei, K.; Kuroha, T.; Kamada-Nobusada, T.; Hayashi, H.; Sakakibara, H. Regulation of Cytokinin Biosynthesis, Compartmentalization and Translocation. J. Exp. Bot. 2007, 59, 75–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, D.; Kang, J.; Kiba, T.; Park, J.; Kojima, M.; Do, J.; Kim, K.Y.; Kwon, M.; Endler, A.; Song, W.; et al. Arabidopsis ABCG14 Is Essential for the Root-Toshoot Translocation of Cytokinin. Proc. Natl. Acad. Sci. USA 2014, 111, 7150–7155. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Novak, O.; Wei, Z.; Gou, M.; Zhang, X.; Yu, Y.; Yang, H.; Cai, Y.; Strnad, M.; Liu, C.-J. Arabidopsis ABCG14 Protein Controls the Acropetal Translocation of Root-Synthesized Cytokinins. Nat. Commun. 2014, 5, 3274. [Google Scholar] [CrossRef] [PubMed]
- Miyawaki, K.; Matsumoto-Kitano, M.; Kakimoto, T. Expression of Cytokinin Biosynthetic Isopentenyltransferase Genes in Arabidopsis: Tissue Specificity and Regulation by Auxin, Cytokinin, and Nitrate. Plant J. 2004, 37, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Ioio, R.D.; Nakamura, K.; Moubayidin, L.; Perilli, S.; Taniguchi, M.; Morita, M.T.; Aoyama, T.; Costantino, P.; Sabatini, S. A Genetic Framework for the Control of Cell Division and Differentiation in the Root Meristem. Science 2008, 322, 1380–1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, M.; Takei, K.; Kojima, M.; Sakakibara, H.; Mori, H. Auxin Controls Local Cytokinin Biosynthesis in the Nodal Stem in Apical Dominance. Plant J. 2006, 45, 1028–1036. [Google Scholar] [CrossRef] [PubMed]
- Nordström, A.; Tarkowski, P.; Tarkowska, D.; Norbaek, R.; Åstot, C.; Dolezal, K.; Sandberg, G. Auxin Regulation of Cytokinin Biosynthesis in Arabidopsis thaliana: A Factor of Potential Importance for Auxin-Cytokininregulated Development. Proc. Natl. Acad. Sci. USA 2004, 101, 8039–8044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, B.; Gunneras, S.A.; Petersson, S.V.; Tarkowski, P.; Graham, N.; May, S.; Dolezal, K.; Sandhberg, G.; Ljung, K. Cytokinin Regulation of Auxin Synthesis in Arabidposis Involves a Homeostatic Feedback Loop Regulated via Auxin and Cytokinin Signal Transduction. Plant Cell 2010, 22, 2956–2969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Showk, S.; Ruonala, R.; Helariutta, Y. Crossing Paths: Cytokinin Signalling and Crosstalk. Development 2013, 140, 1373–1383. [Google Scholar] [CrossRef] [Green Version]
- Laplaze, L.; Benkova, E.; Casimiro, I.; Maes, L.; Vanneste, S.; Swarup, R.; Weijers, D.; Calvo, V.; Parizot, B.; Herrera-Rodriguez, M.B.; et al. Cytokinins Act Directly on Lateral Root Founder Cells to Inhibit Root Initiation. Plant Cell 2008, 19, 3889–3900. [Google Scholar] [CrossRef] [Green Version]
- Hou, H.; Jessica, E.; James, M.; Elizbath, A.S. FORKED1 Encodes a PH Domain Protein That Is Required for PIN1 Localization in Developming Leaf Veins. Plant J. 2010, 63, 960–973. [Google Scholar] [CrossRef]
- Kalousek, P.; Buchtová, D.; Balla, J. Cytokinin and Polar Transport of Auxin in Axillary Pea Buds. Acta Univ. Agri Cult. Et Silvic. Mendel. Brun. LVIII 2010, 58, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; To, J.P.C.; Cheng, C.-Y.; Schaller, G.E.; Kieber, J.J. Type-A Response Regulators Are Required for Proper Root Apical Meristem Function through Post-Transcriptional Regulation of PIN Auxin Efflux Carriers. Plant J. 2011, 68, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Marhavy, P.; Bielach, A.; Abas, L.; Abuzeineh, A.; Duclercq, J.; Tanaka, H.; Pařezová, M.; Petrasek, J.; Friml, J.; Kleine-Vehn, J.; et al. Cytokinin Modulates Endocytic Trafficking of PIN1 Auxin Efflux Carrier to Control Plant Organogenesis. Dev. Cell 2011, 21, 796–804. [Google Scholar] [CrossRef] [PubMed]
- Bishopp, A.; Help, H.; El-Showk, S.; Weijers, D.; Scheres, B.; Friml, J.; Benkova, E.; Mahonen, A.P.; Helariutta, Y. A Mutually Inhibitory Interaction between Auixin and Cytokinin Specifies Vascular Pattern in Roots. Curr. Biol. 2011, 21, 917–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenner, W.G.; Schmülling, T. Transcript Profiling of Cytokinin Action in Arabidopsis Roots and Shoots Discovers Largely Similar But Also Organ-Specific Responses. BMC Plant Biol. 2012, 12, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biedroń, M.; Banasiak, A. Auxin-Mediated Regulation of Vascular Patterning in Arabidopsis thaliana Leaves. Plant Cell Rep. 2018, 37, 1215–1229. [Google Scholar] [CrossRef] [Green Version]
- Heisler, M.G.; Byrne, M.E. Progress in Understanding the Role of Auxin in Lateral Organ Development in Plants. Curr. Opin. Plant Biol. 2020, 53, 73–79. [Google Scholar] [CrossRef]
- Lee, B.-H.; Yu, S.-I.; Jackson, D. Control of Plant Architecture: The Role of Phyllotaxy and Plastochron. J. Plant Biol. 2009, 52, 277–282. [Google Scholar] [CrossRef]
- Kieber, J.J.; Schaller, G.E. Cytokinins. Arab. Book 2014, 12, e0168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besnard, F.; Refahi, Y.; Morin, V.; Marteaux, B.; Brunoud, G.; Chambrier, P.; Rozier, F.; Mirabet, V.; Legrand, J.; Lainé, S.; et al. Cytokinin Signalling Inhibitory Fields Provide Robustness to Phyllotaxis. Nat. Cell Biol. 2014, 505, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Bilsborough, G.D.; Runions, A.; Barkoulas, M.; Jenkins, H.W.; Hasson, A.; Galinha, C.; Laufs, P.; Hay, A.; Prusinkiewicz, P.; Tsiantis, M. Model for the Regulation of Arabidopsis thaliana Leaf Margin Development. Proc. Natl. Acad. Sci. USA 2011, 108, 3424–3429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeMason, D.A.; Polowick, P.L. Patterns of DR5::GUS Expression in Organs of Pea (Pisum sativum). Int. J. Plant Sci. 2009, 170, 1–11. [Google Scholar] [CrossRef]
- Koenig, D.; Bayer, E.; Kang, J.; Kuhlemeier, C.; Sinha, N. Auxin Patterns Solanum Lycopersicum Leaf Morphogenesis. Development 2009, 136, 2997–3006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Gera, H.; Shwartz, I.; Shao, M.-R.; Shani, E.; Estelle, M.; Ori, N. ENTIRE and GOBLET Promote Leaflet Development in Tomato by Modulating Auxin Response. Plant J. 2012, 70, 903–915. [Google Scholar] [CrossRef] [PubMed]
- Holst, K.; Schmülling, T.; Werner, T. Enhanced Cytokinin Degradation in Leaf Primordia of Transgenic Arabidopsis Plants Reduces Leaf Size and Shoot Organ Primordia Formation. J. Plant Physiol. 2011, 168, 1328–1334. [Google Scholar] [CrossRef] [PubMed]
- Bharathan, G.; Goliber, T.E.; Moore, C.; Kessler, S.; Pham, T.; Sinha, N.R. Homologies in Leaf Form Inferred from KNOXI Gene Expression during Development. Science 2002, 296, 1858–1860. [Google Scholar] [CrossRef]
- Bar, M.; Ori, N. Leaf Development and Morphogenesis. Development 2014, 141, 4219–4230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shani, E.; Ben-Gera, H.; Shleizer-Burko, S.; Burko, Y.; Weiss, D.; Ori, N. Cytokinin Regulates Compound Leaf Development in Tomato. Plant Cell 2010, 22, 3206–3217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Hu, S.; Zhao, X.; Kumar, S.; Li, Y.; Yang, J.; Hou, H. Mechanisms of the Morphological Plasticity Induced by Phytohormones and the Environment in Plants. Int. J. Mol. Sci. 2021, 22, 765. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Hu, S.; Yang, J.; Schultz, E.A.; Clarke, K.; Hou, H. Water-Wisteria As an Ideal Plant to Study Heterophylly in Higher Aquatic Plants. Plant Cell Rep. 2017, 36, 1225–1236. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Hu, S.; Hou, H.; Kimura, S. Heterophylly: Phenotypic Plasticity of Leaf Shape in Aquatic and Amphibious Plants. Plants 2019, 8, 420. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Hu, S.; Yang, J.; Zhao, X.; Kimura, S.; Schultz, E.A.; Hou, H. Establishment of an Agrobacterium Mediated Trans-Formation Protocol for the Detection of Cytokinin in the Heterophyllous Plant Hygrophila difformis (Acanthaceae). Plant Cell Rep. 2020, 39, 737–750. [Google Scholar] [CrossRef]
- Nakayama, H.; Nakayama, N.; Seiki, S.; Kojima, M.; Sakakibara, H.; Sinha, N.; Kimura, S. Regulation of the KNOX-GA Gene Module Induces Heterophyllic Alteration in North American Lake Cress. Plant Cell 2014, 26, 4733–4748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, H.; Sinha, N.R.; Kimura, S. How Do Plants and Phytohormones Accomplish Heterophylly, Leaf Phenotypic Plasticity, in Response to Environmental Cues. Front. Plant Sci. 2017, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Lee, S.; Park, G.; Cho, H.; Choi, D.; Umeda, M.; Choi, Y.; Hwang, D.; Hwang, I. Cytokinin-Responsive Growth Regulator Regulates Cell Expansion and Cytokinin-Mediated Cell Cycle Progression. Plant Physiol. 2021, 186, 1734–1746. [Google Scholar] [CrossRef]
- Das Gupta, M.; Aggarwal, P.; Nath, U. CINCINNATA in Antirrhinum majus Directly Modulates Genes Involved in Cytokinin and Auxin Signaling. New Phytol. 2014, 204, 901–912. [Google Scholar] [CrossRef] [PubMed]
- Efroni, I.; Han, S.-K.; Kim, H.J.; Wu, M.-F.; Steiner, E.; Birnbaum, K.D.; Hong, J.C.; Eshed, Y.; Wagner, D. Regulation of Leaf Maturation by Chromatin-Mediated Modulation of Cytokinin Responses. Dev. Cell 2013, 24, 438–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muszynski, M.G.; Moss-Taylor, L.; Chudalayandi, S.; Cahill, J.; Del Valle-Echevarria, A.R.; Alvarez-Castro, I.; Petefish, A.; Sakakibara, H.; Krivosheev, D.M.; Lomin, S.N.; et al. The Maize Hairy Sheath Frayed1 (Hsf1) Mutation Alters Leaf Patterning through Increased Cytokinin Signaling. Plant Cell 2020, 32, 1501–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurepa, J.; Shull, T.E.; Smalle, J.A. Antagonistic Activity of Auxin and Cytokinin in Shoot and Root Organs. Plant Direct 2019, 3, e00121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, J.; Jones, A.; Godin, C.; Traas, J. Systems Analysis of Shoot Apical Meristem Growth and Development: Integrating Hormonal and Mechanical Signaling. Plant Cell 2012, 24, 3907–3919. [Google Scholar] [CrossRef] [Green Version]
- Kierzkowski, D.; Lenhard, M.; Smith, R.; Kuhlemeier, C. Interaction between Meristem Tissue Layers Controls Phyllotaxis. Dev. Cell 2013, 26, 616–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furutani, M.; Nakano, Y.; Tasaka, M. MAB4-Induced Auxin Sink Generates Local Auxin Gradients in Arabidopsis Organ for-Mation. Proc. Natl. Acad. Sci. USA 2014, 111, 1198–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoma, S.; Lucas, M.; Chopard, J.; Schaedel, M.; Traas, J.; Godin, C. Flux-Based Transport Enhancement As a Plausible Unifying Mechanism for Auxin Transport in Meristem Development. PLoS Comput. Biol. 2008, 4, e1000207. [Google Scholar] [CrossRef]
- Bartrina, I.; Otto, E.; Strnad, M.; Werner, T.; Schmülling, T. Cytokinin Regulates the Activity of Reproductive Meristems, Flower Organ Size, Ovule Formation, and Thus Seed Yield in Arabidopsis thaliana. Plant Cell 2011, 23, 69–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, J.I.; Hibara, K.I.; Kojima, M.; Sakakibara, H.; Nagato, Y. Rice DECUSSATE Controls Phyllotaxy by Affecting the Cy-Tokinin Signaling Pathway. Plant J. 2012, 72, 869–881. [Google Scholar] [CrossRef]
- Uchida, N.; Shimada, M.; Tasaka, M. ERECTA-Family Receptor Kinases Regulate Stem Cell Homeostasis via Buffering Its Cytokinin Responsiveness in the Shoot Apical Meristem. Plant Cell Physiol. 2012, 54, 343–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.-K.; Wilson, R.L.; Palme, K.; Ditengou, F.A.; Shpak, E.D. ERECTA Family Genes Regulate Auxin Transport in the Shoot Apical Meristem and Forming Leaf Primordia. Plant Physiol. 2013, 162, 1978–1991. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.H.; Johnston, R.; Yang, Y.; Gallavotti, A.; Kojima, M.; Travençolo, B.A.N.; da, F. Costa, L.; Sakakibara, H.; Jackson, D. Studies of Aberrant Phyllotaxy1 Mutants of Maize Indicate Complex Interactions between Auxin and Cytokinin Signaling in the Shoot Apical Meristem. Plant Physiol. 2009, 150, 205–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamport, D.T.A.; Tan, L.; Held, M.; Kieliszewski, M.J. Phyllotaxis Turns Over a New Leaf—A New Hypothesis. Int. J. Mol. Sci. 2020, 21, 1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
| Gene Name | Biological Function | Regulation Effect | Reference |
|---|---|---|---|
| Auxin Signaling Pathway Genes Regulated by CKs | |||
| AUX1 | Auxin influx transport | Upregulation | [57] |
| PIN1, PIN2, PIN3, PIN4, PIN6, PIN7 | Auxin efflux transport | Lateralization, Upregulation, Tissue-specific downregulation | [54,59,60] |
| IAA7, IAA13, IAA17, AMI1 | Auxin signaling | Downregulation | [61] |
| AUXIN RESISTANT 1, 31 (AXR1, AXR3) | Auxin signaling | Upregulation | [53,62] |
| CYP79B2 | Auxin biosynthesis | Upregulation | [53] |
| YUCCA5, YUCCA6 | Auxin biosynthesis | Temporal expression regulation | [53] |
| ANTHRANILATE SYNTHASE α1 (ASA1/WEI2) | Auxin precursor synthesis | Up-regulation | [53] |
| GH3.17, GH3.9 | Auxin conjugation | Up-regulation | [53] |
| PHOSPHORIBOSYLANTHRANILATE TRANSFERASE1 (PAT1)/TRYPTOPHAN BIOSYNTHESIS1 (TRP1), INDOLE-3-GLYCEROL PHOSPHATE SYNTHASE(IGPS), NITRILASE 3 (NIT3) | Auxin biosynthesis | Up-regulation | [53] |
| CK Signaling Pathways Genes Regulated by Auxin | |||
| ARR7, ARR15 | Cytokinin signaling | Spatial expression regulation | [4,52] |
| IPT1, IPT2 | Cytokinin biosynthesis | Downregulation | [49] |
| IPT5, IPT7 | Cytokinin biosynthesis | Up-regulation | [47] |
| CRF2, AHP6 | Cytokinin signaling | Up-regulation | [7,60] |
| CKX2, CKX4, CKX7 | Cytokinin degradation | Downregulation | [51] |
| CYP735A | Cytokinin biosynthesis | Downregulation | [29] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussain, S.; Nanda, S.; Zhang, J.; Rehmani, M.I.A.; Suleman, M.; Li, G.; Hou, H. Auxin and Cytokinin Interplay during Leaf Morphogenesis and Phyllotaxy. Plants 2021, 10, 1732. https://doi.org/10.3390/plants10081732
Hussain S, Nanda S, Zhang J, Rehmani MIA, Suleman M, Li G, Hou H. Auxin and Cytokinin Interplay during Leaf Morphogenesis and Phyllotaxy. Plants. 2021; 10(8):1732. https://doi.org/10.3390/plants10081732
Chicago/Turabian StyleHussain, Sajid, Satyabrata Nanda, Junhua Zhang, Muhammad Ishaq Asif Rehmani, Muhammad Suleman, Gaojie Li, and Hongwei Hou. 2021. "Auxin and Cytokinin Interplay during Leaf Morphogenesis and Phyllotaxy" Plants 10, no. 8: 1732. https://doi.org/10.3390/plants10081732