
In Vitro Propagation Method for Production of Phenolic-Rich Planting Material of Culinary Rhubarb ‘Malinowy’


Abstract
:1. Introduction
2. Results
2.1. Culture Initiation
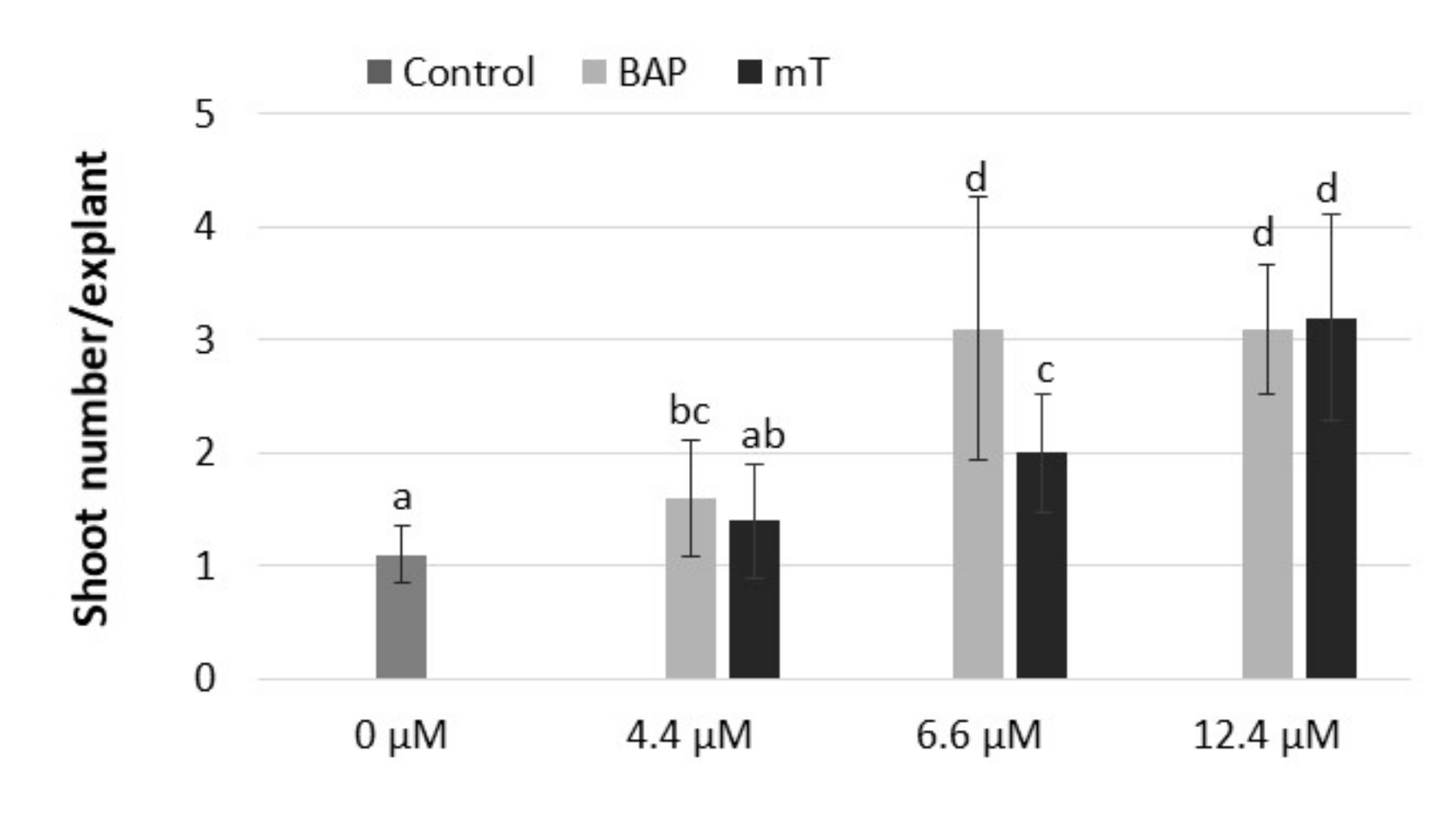
2.2. Shoot Multiplication
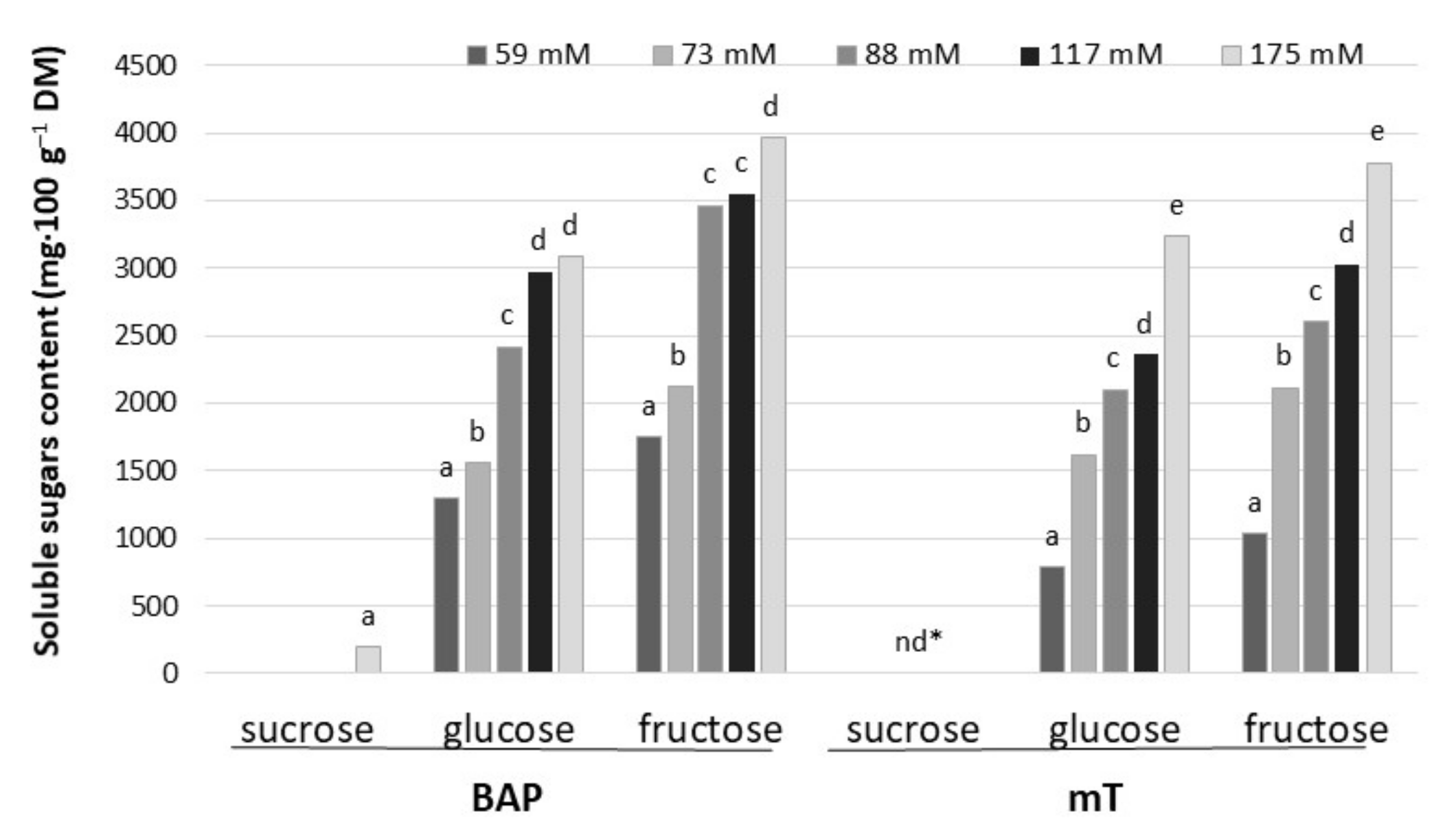
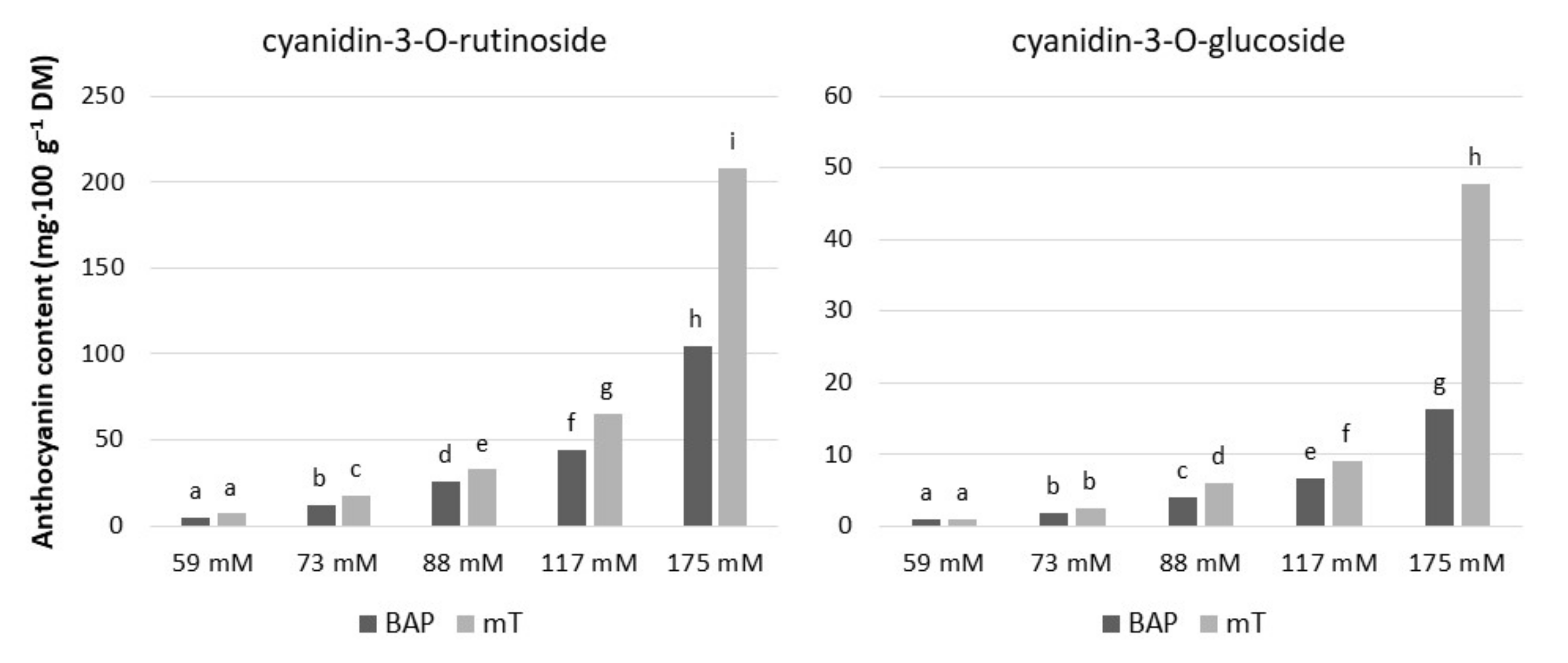
2.3. Soluble Sugar and Anthocyanin Contents in Leaf petioles
2.4. Rooting and Acclimatization
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Culture Initiation
4.3. Shoot Multiplication
4.4. Rooting and Acclimatization
4.5. Soluble Sugar Analysis
4.6. Estimation of Anthocyanins by HPLC
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, A.; Yang, M.; Liu, J. Molecular phylogeny, recent radiation and evolution of gross morphology of the rhubarb genus Rheum (Polygonaceae) inferred from chloroplast DNA trnL-F sequences. Ann. Bot. 2005, 96, 489–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Q.; Wu, H.F.; Guo, J.; Nan, H.J.; Chen, S.L.; Yang, J.S.; Xu, X.D. Review of rhubarbs: Chemistry and pharmacology. Chin. Herb. Med. 2013, 5, 9–32. [Google Scholar]
- Xiang, H.; Zuo, J.; Guo, F.; Dong, D. What we already know about rhubarb: A comprehensive review. Chin. Med. 2020, 15, 88. [Google Scholar] [CrossRef] [PubMed]
- Bratsch, A.; Mainville, D. Specialty crop profile: Rhubarb; Virginia Tech: Blacksburg, VA, USA, 2009; Publication 438–110. [Google Scholar]
- Foust, C.M.; Marshall, D.E. Culinary rhubarb production in North America: History and recent statistics. HortScience 1991, 26, 1360–1363. [Google Scholar] [CrossRef] [Green Version]
- Roberfroid, M.B. Concepts and strategy of functional food science: The European perspective. Am. J. Clin. Nutr. 2000, 71, 1660S–1664S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oszmiański, J.; Wojdyło, A. Polyphenol content and antioxidative activity in apple purées with rhubarb juice supplement. Int. J. Food Sci. Technol. 2008, 43, 501–508. [Google Scholar] [CrossRef]
- Krafczyk, N.; Kötke, M.; Lehnert, N.; Glomb, M.A. Phenolic composition of rhubarb. Eur. Food Reseach Technol. 2008, 228, 187–196. [Google Scholar] [CrossRef]
- Takeoka, G.R.; Dao, L.; Harden, L.; Pantoja, A.; Kuhl, J.C. Antioxidant activity, phenolic and anthocyanin contents of various rhubarb (Rheum spp.) varieties. Int. J. Food Sci. Technol. 2013, 48, 172–178. [Google Scholar] [CrossRef]
- Kołodziejczyk-Czepas, J.; Liudvytska, O. Rheum rhaponticum and Rheum rhabarbarum: A review of phytochemistry, biological activities and therapeutic potential. Phytochem. Rev. 2021, 20, 589–607. [Google Scholar] [CrossRef]
- Kalisz, S.; Oszmiański, J.; Kolniak-Ostek, J.; Grobelna, A.; Kieliszek, M.; Cendrowski, A. Effect of a variety of polyphenols compounds and antioxidant properties of rhubarb (Rheum rhabarbarum). Food Sci. Technol. 2020, 118, 108775. [Google Scholar] [CrossRef]
- Kuhl, J.C.; DeBoer, V.L. Genetic diversity of rhubarb cultivars. J. Am. Soc. Hortic. Sci. 2008, 133, 587–592. [Google Scholar] [CrossRef]
- Maynard, D.N. Evaluation of propagating stock and practices for annual rhubarb production. HortScience 1990, 25, 1399–1401. [Google Scholar] [CrossRef]
- Tomlinson, J.A.; Walkey, D.G.A. The isolation and identification of rhubarb viruses occurring in Britain. Ann. Appl. Biol. 1976, 59, 415–427. [Google Scholar] [CrossRef]
- Robertson, N.L.; Ianson, D.C. First report of Turnip mosaic virus in rhubarb in Alaska. Plant Dis. 2005, 89, 430. [Google Scholar] [CrossRef]
- Thomas, J. Virus Identification and Development of Long-Term Management Strategies for the Rhubarb Industry; Final report Hal Project VG05053; Horticulture Australia: Sydney, NSW, Australia, 2011; p. 93. [Google Scholar]
- Komorowska, B.; Ptaszek, M.; Jarecka-Boncela, A.; Hasiów-Jaroszewska, B. First report of Arabis Mosaic Virus in Rhubarb in Poland. Plant Dis. 2018, 102, 1863. [Google Scholar] [CrossRef] [PubMed]
- Clapa, D.; Borsai, O.; Hârta, M.; Bonta, V.; Szabo, K.; Coman, V.; Bobis, O. Micropropagation, genetic fidelity and phenolic compound of Rheum rhabarbarum L. Plants 2020, 9, 656. [Google Scholar] [CrossRef] [PubMed]
- Lepse, L. Adapting technology for local rhubarb (Rheum rhaponticum L.) clones propagation In Vitro. Latv. J. Agron. 2005, 8, 324–327. [Google Scholar]
- Lassus, C.; Voipio, I. Micropropagation of rhubarb with special reference to weaning stage and subsequent growth. Agric. Sci. Finl. 1994, 3, 189–194. [Google Scholar] [CrossRef] [Green Version]
- Kozak, D.; Sałata, A. Effect of cytokinins on In Vitro multiplication of rhubarb (Rheum rhaponticum L.) ‘Karpov Lipskiego’ shoots and ex vitro acclimatization and growth. Acta Sci. Pol. Hortorum Cultus 2011, 10, 75–87. [Google Scholar]
- Beretta, D.; Eccherr, T. Determination of optimal level of BAP, GA3 and NAA in micropropagation of Camellia by an orthogonal composite design. Acta Hortic. 1987, 212, 151–154. [Google Scholar] [CrossRef]
- Wojtania, A.; Skrzypek, E.; Gabryszewska, E. Morphological and biochemical responses to gibberellic acid in Magnolia × ‘Spectrum’ In Vitro. Acta Biol. Crac. Ser. Bot. 2016, 58, 103–111. [Google Scholar] [CrossRef]
- Gabryszewska, E.A. Effect of different sucrose and nitrogen salt levels in the medium and temperature on In Vitro propagation of Helleborus niger L. Acta Agrobotanica 2015, 68, 161–171. [Google Scholar] [CrossRef] [Green Version]
- El-Dawayati, M.M.; Zayed, Z.E. Controlling hyperhydricity in date palm In Vitro culture by reduced concentration of nitrate nutrients. In Date Palm Biotechnology Protocols Volume I: Tissue Culture Applications; Al-Khayri, J., Jain, S., Johnson, D., Eds.; Humana Press: New York, NY, USA, 2017; Volume 1637. [Google Scholar] [CrossRef]
- Ogura-Tsujita, Y.; Okuba, H. Effects of low nitrogen medium on endogenous changes in ethylene, auxins, and cytokinins in In Vitro shoot formation from rhizomes of Cymbidium kanran. Vitr. Cell. Dev. Biol.-Plant 2006, 42, 614–616. [Google Scholar] [CrossRef]
- Gabryszewska, E. The effect of glucose and growth regulators on the organogenesis of Paeonia lactiflora Pall. in vitro. J. Fruit Ornam. Plants Res. 2010, 18, 309–320. [Google Scholar]
- Wojtania, A.; Węgrzynowicz-Lesiak, E.; Dziurka, M.; Waligórski, P. Sucrose and cytokinin interactions in relations to ethylene and abscisic acid production in the regulation of morphogenesis in Pelargonium × hortorum L.H. BAILEY In Vitro. Acta Biol. Crac. Ser. Bot. 2015, 57, 62–69. [Google Scholar] [CrossRef]
- Wojtania, A.; Matysiak, B. In vitro propagation of Rosa ‘Konstancin’ (R. rugosa × R. beggeriana), a plant with high nutritional and pro-health value. Folia Hortic. 2018, 30, 259–267. [Google Scholar] [CrossRef] [Green Version]
- Smeekens, S.; Ma, J.; Hanson, J.; Rolland, F. Sugar signal and molecular networks controlling plant growth. Curr. Opin. Plant Biol. 2010, 13, 274–279. [Google Scholar] [CrossRef]
- Ciereszko, I. Regulatory roles of sugars in plant growth and development. Acta Soc. Bot. Pol. 2018, 87, 3583. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, F.M.; Visser, R.G.F.; Bovy, A. Antocyanin biosynthesis and degradation mechanisms in Solanaceous vegetables: A review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef] [PubMed]
- Weiss, D. Regulation of flower pigmentation and growth: Multiple signalling pathways control anthocyanin synthesis in expanding petals. Physiol. Plant. 2000, 110, 152–157. [Google Scholar] [CrossRef]
- Gallop, R.; Farhi, S.; Peri, A. Regulation of the leucoanthocyanindin dioxygenase gene expression in Vitis vinifera. Plant Sci. 2001, 161, 579–588. [Google Scholar] [CrossRef]
- See, K.S.; Bhatt, A.; Keng, C.L. Effect of sucrose and methyl jasmonate on biomass and anthocyanin production in cell suspension culture of Melastoma malabathricum (Melastomaceae). Rev. Biol. Trop. 2011, 59, 597–606. [Google Scholar] [PubMed]
- Kawa-Miszczak, L.; Węgrzynowicz-Lesiak, E.; Gabryszewska, E.; Saniewski, M. Effect of different sucrose and nitrogen levels in the medium on chlorophyll and anthocyanin content i clematis pitcher shoots cultured In Vitro at different temperatures. J. Fruit Ornam. Plant Res. 2009, 17, 113–121. [Google Scholar]
- Ai, T.N.; Naing, A.H.; Lim, S.-H.; Kim, C.K. Sucrose-induced anthocyanin accumulation in vegetative tissue of Petunia plants requires anthocyanin regulatory transcription factor. Plant Sci. 2016, 252, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Das, P.K.; Shin, D.H.; Choi, S.B.; Park, Y.I. Sugar-hormone cross-talk in anthocyanin biosynthesis. Malecules Cells 2012, 34, 501–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Góraj-Koniarska, J.; Saniewski, M. The effect of sugars in relations to methyl jasmonate on anthocyanin formation in the roots of Kolanchoe blossfeldiana (Poelln.). Acta Agrobot. 2014, 68, 173–178. [Google Scholar]
- Aremu, A.O.; Bairu, M.W.; Szüčová, L.; Doležal, K.; Finnie, J.F.; Van Staden, J. Assessment of the role of meta-topolins on In Vitro produced phenolics and acclimatization competence of micropropagated ‘Williams’ banana. Acta Physiol. Plant. 2012, 34, 2265–2273. [Google Scholar] [CrossRef]
- Wojtania, A. Effect of meta-topolin on In Vitro propagation of Pelargonium × hortorum and Pelargonium × hederaefolium cultivars. Acta Soc. Bot. Pol. 2010, 79, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Podwyszyńska, M.; Cieślińska, M. Rooting shoots of apple varieties and their tetraploids obtained by the In Vitro technique. Acta Sci. Pol. Hortorum Cultus 2018, 17, 49–62. [Google Scholar] [CrossRef]
- Podwyszyńska, M.; Pluta, S. In vitro tetraploid induction of the blackcurrant (Ribes nigrum L.) and preliminary phenotypic observations. Zemdirb.-Agric. 2019, 106, 151–158. [Google Scholar] [CrossRef]
- Clark, M.F.; Adams, A.N. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol. 1977, 34, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Nielsen, I.L.; Haren, G.R.; Magnussen, E.L.; Dragsted, L.O.; Rasmussen, S.E. Quantification of anthocyanins in commercial black currant juices by simple high-performance liquid chromatography. Investigation of their pH stability and antioxidative potency. J. Agric. Food Chem. 2003, 51, 5861–5866. [Google Scholar] [CrossRef] [PubMed]
- Dogan, Y.; Baslar, S.; Ay, G.; Mert, H.H. The use of wild edible plants in western and central Anatolia (Turkey). Econ. Bot. 2004, 58, 684–690. [Google Scholar] [CrossRef]
- Penafiel, D.; Vanhove, W.; Espinel, R.L.; Van Damme, P. Food biodiversity includes both locally cultivated and wild food species in Guasaganda, Central Ecuador. J. Ethn. Foods 2019, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Perrino, E.V.; Valerio, F.; Gannouchi, A.; Trani, A.; Mezzapesa, G. Ecological and Plant Community Implication on Essential Oils Composition in Useful Wild Officinal Species: A Pilot Case Study in Apulia (Italy). Plants 2021, 10, 574. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donor Plants | Total Number of Explants | Contaminated Explants (%) | Uncontaminated Explants that Did Not Developed Shoots (%) | Uncontaminated Explants that Developed Shoots (%) | |
|---|---|---|---|---|---|
| Bacteria | Fungi | ||||
| M1 * | 34 | 29.4 | - | 3.0 | 67.6 |
| M2 | 56 | 28.6 | 10.7 | 8.8 | 51.9 |
| Propagation Medium | Survival Rate (%) | Shoot Number/Explant |
|---|---|---|
| 100% MS control (hormone free) | 0 | - |
| 100% MS + 4.4 µM BAP + 0.3 µM GA3 | 95.3 | 1.69 bc 1 |
| 100% MS + 4.4 µM BAP + 0.1 µM NAA | 58.7 | 1.07 a |
| 75% MS + 4.4 µM BAP + 0.3 µM GA3 | 94.1 | 1.92 c |
| 75% MS + 4.4 µM BAP + 0.1 µM NAA | 72.0 | 1.31 ab |
| Cytokinin Type | Sucrose Concentration (mM) | Fresh Mass (g) | Shoot Number | Shoot Length (mm) | Leaf Number/ Clump | ||
|---|---|---|---|---|---|---|---|
| Shoot Clump | Callus | Single Shoot | |||||
| BAP | 59 | 1.90 bc 1 | 0.170 a | 0.395 a | 4.8 e | 31.6 a | 25.1 d |
| 73 | 2.37 cd | 0.162 a | 0.566 b–c | 4.3 de | 37.5 bc | 21.5 d | |
| 88 | 1.68 ab | 0.261 a–c | 0.586 bc | 3.1 bc | 38.5 bc | 16.9 c | |
| 117 | 1.28 a | 0.362 b–d | 0.653 c–d | 2.2 ab | 42.4 cd | 11.5 b | |
| 175 | 1.41 a | 0.532 e | 0.907 e | 1.5 a | 49.0 e | 8.9 ab | |
| mT | 59 | 2.51 d | 0.256 a–c | 0.446 ab | 4.9 e | 30.9 a | 32.1 e |
| 73 | 2.31 d | 0.237 ab | 0.522 a–c | 4.7 e | 36.6 b | 24.6 d | |
| 88 | 1.61 ab | 0.423 de | 0.494 a–c | 3.7 cd | 40.1 b–d | 12.7 b | |
| 117 | 1.31 a | 0.402 c–e | 0.576 b–c | 2.3 ab | 38.9 bc | 11.7 b | |
| 175 | 1.28 a | 0.430 de | 0.768 de | 1.7 a | 44.8 de | 6.9 a | |
| Treatment | Auxin Concentrations (µM) | Rooting Frequency (%) | Root Number/Shoot | Root Length (mm) | Shoot Length (mm) |
|---|---|---|---|---|---|
| Control | 0.0 | 40.0 a 1 | 2.1 a | 29.2 b | 75.2 a |
| IBA | 0.49 | 55.9 b | 2.5 a | 28.8 b | 72.0 a |
| 2.5 | 82.0 c | 2.6 a | 30.3 b | 77.8 a | |
| 4.9 | 94.9 d | 10.7 b | 13.84 a | 78.5 a |
| Type of Microcutting | Survival Frequency (%) | Cutting Length (mm) | Rooting Index |
|---|---|---|---|
| Unrooted | 83 | 72.8 a 1 | 1.9 a |
| Rooted | 100 | 107.0 b | 2.9 b |
| Growth Duration | Length of Leaf Petioles (mm) | Leaf Area (cm2) | Soluble Sugar Contents (mg·100 g−1 DM) | Anthocyanin Contents (mg·100 g−1 DM) |
|---|---|---|---|---|
| 1 week | 60.5 | 20.2 | Sucrose-t.a. Glucose-762 Fructose-858 | cyanidin-3-O-glucoside-2.58 cyanidin-3-O-rutinoside-45.5 |
| 6 weeks | 71.1 | 38.8 | Sucrose-2196 Glucose-4958 Fructose-4790 | cyanidin-3-O-glucoside-11.3 cyanidin-3-O-rutinoside-176.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojtania, A.; Mieszczakowska-Frąc, M. In Vitro Propagation Method for Production of Phenolic-Rich Planting Material of Culinary Rhubarb ‘Malinowy’. Plants 2021, 10, 1768. https://doi.org/10.3390/plants10091768
Wojtania A, Mieszczakowska-Frąc M. In Vitro Propagation Method for Production of Phenolic-Rich Planting Material of Culinary Rhubarb ‘Malinowy’. Plants. 2021; 10(9):1768. https://doi.org/10.3390/plants10091768
Chicago/Turabian StyleWojtania, Agnieszka, and Monika Mieszczakowska-Frąc. 2021. "In Vitro Propagation Method for Production of Phenolic-Rich Planting Material of Culinary Rhubarb ‘Malinowy’" Plants 10, no. 9: 1768. https://doi.org/10.3390/plants10091768
APA StyleWojtania, A., & Mieszczakowska-Frąc, M. (2021). In Vitro Propagation Method for Production of Phenolic-Rich Planting Material of Culinary Rhubarb ‘Malinowy’. Plants, 10(9), 1768. https://doi.org/10.3390/plants10091768