
Stem and Leaf Anatomy of Aragoa (Plantaginaceae): In Search of Lost Rays



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
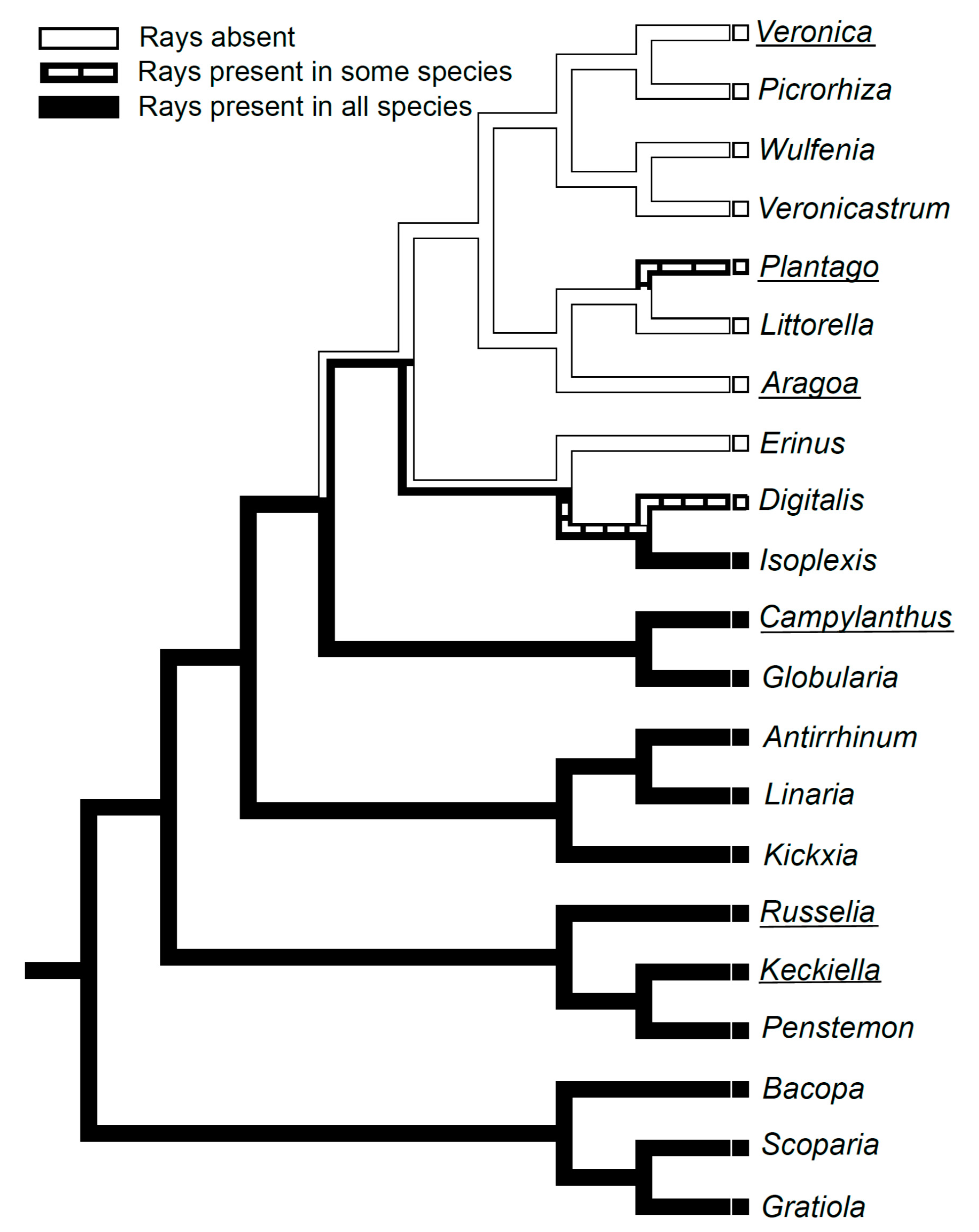
1. Introduction
2. Results
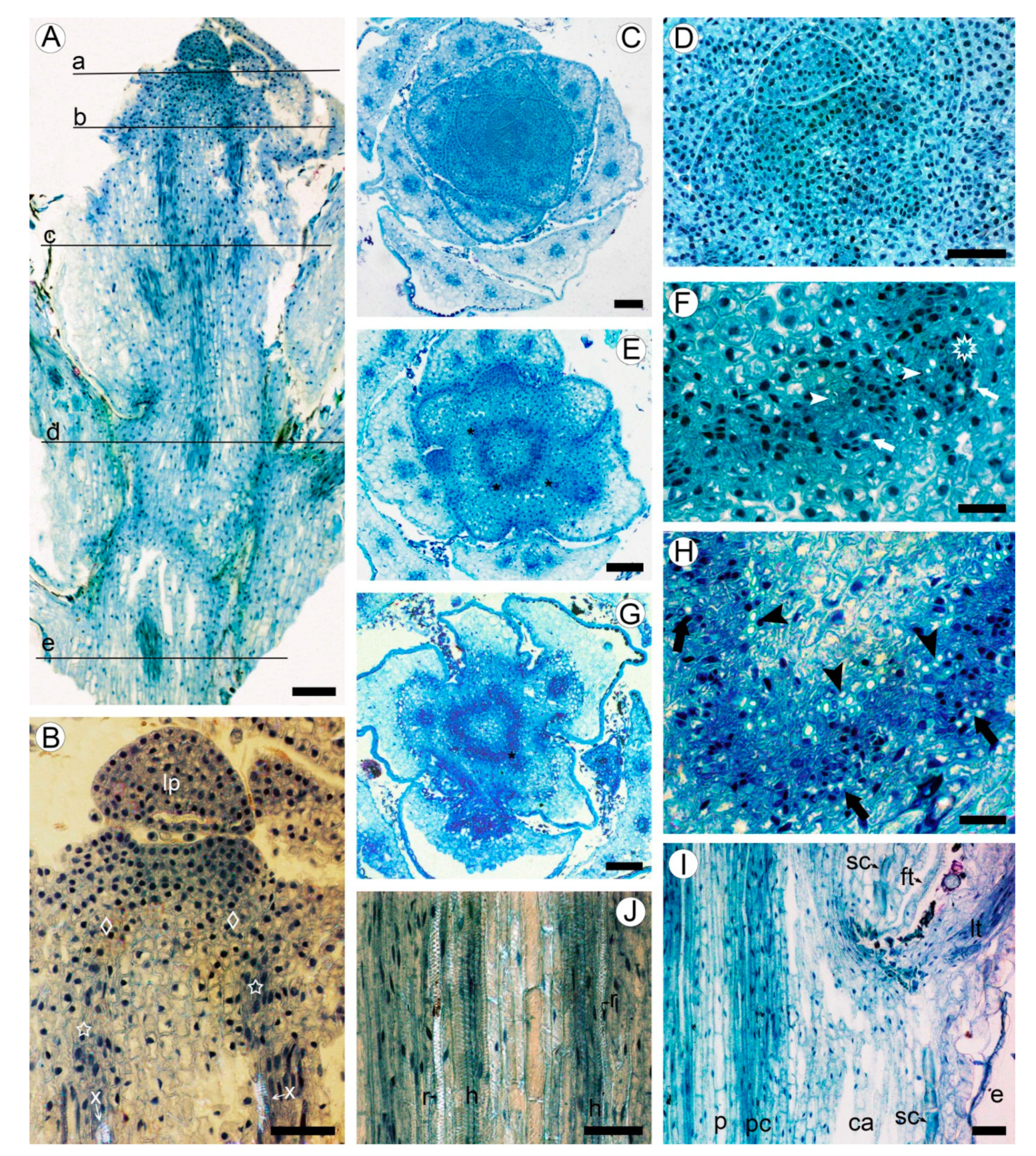
2.1. Development of Meristematic and Conductive Tissues in Stem
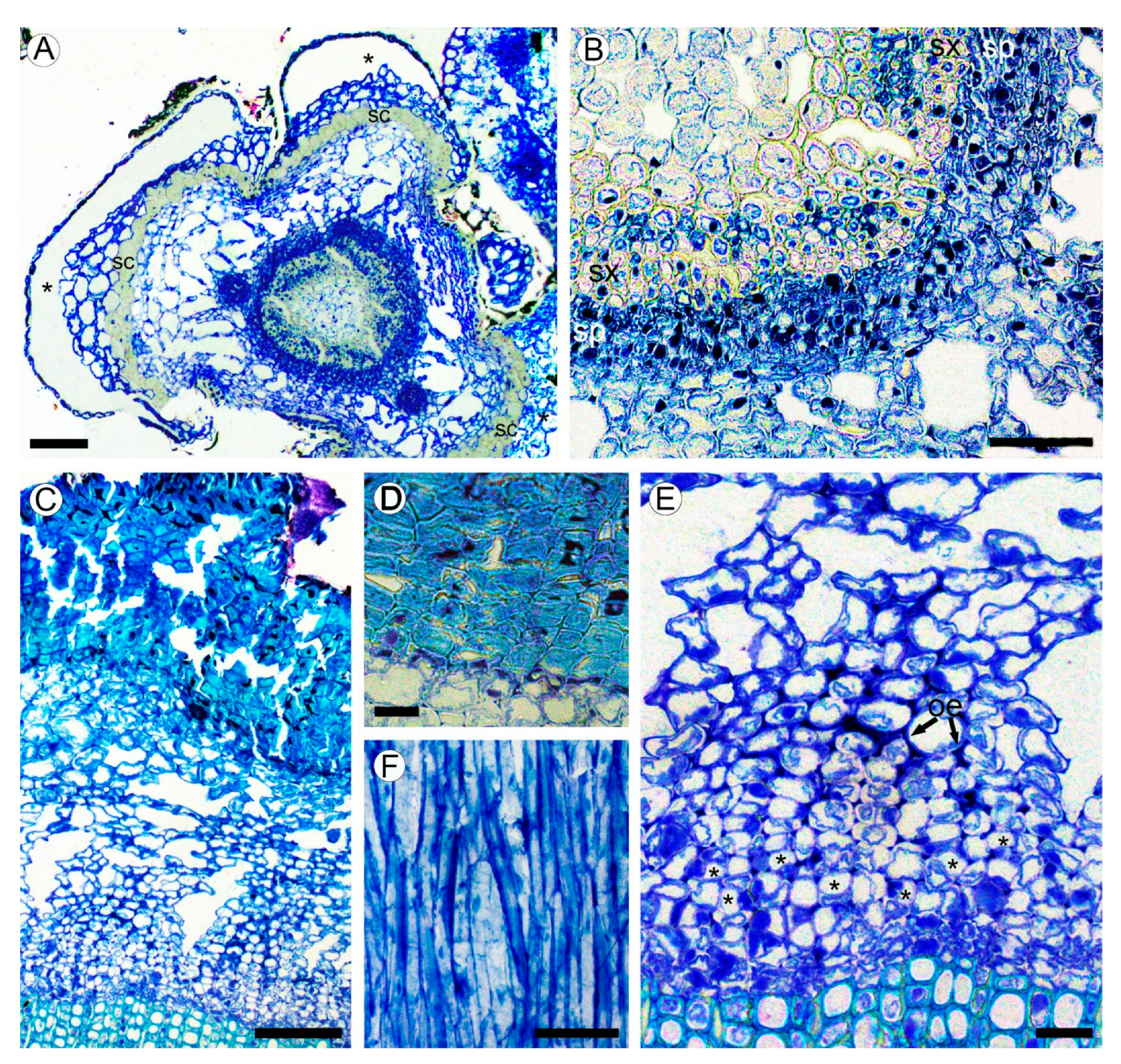
2.2. Bark Structure
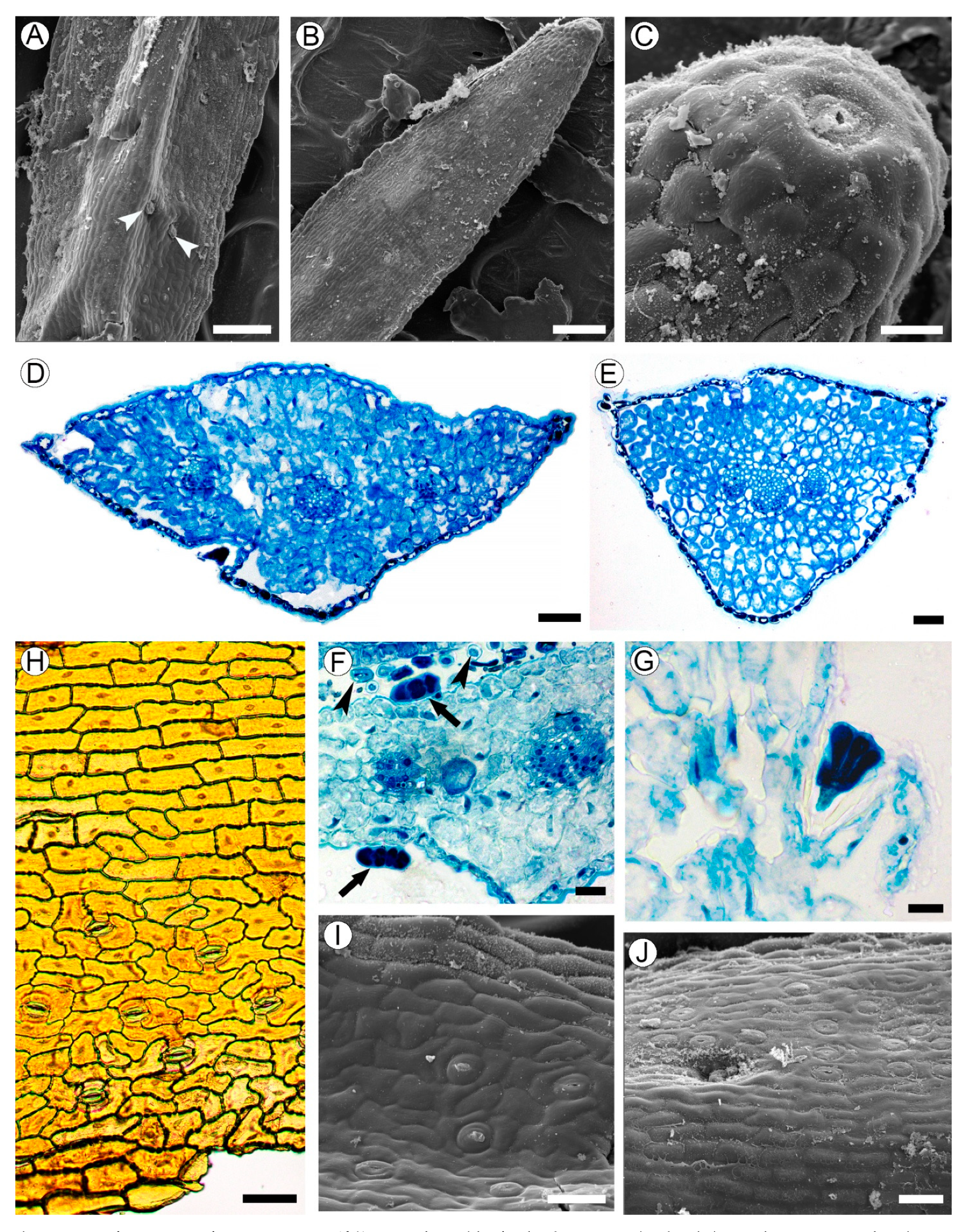
2.3. Leaf Structure
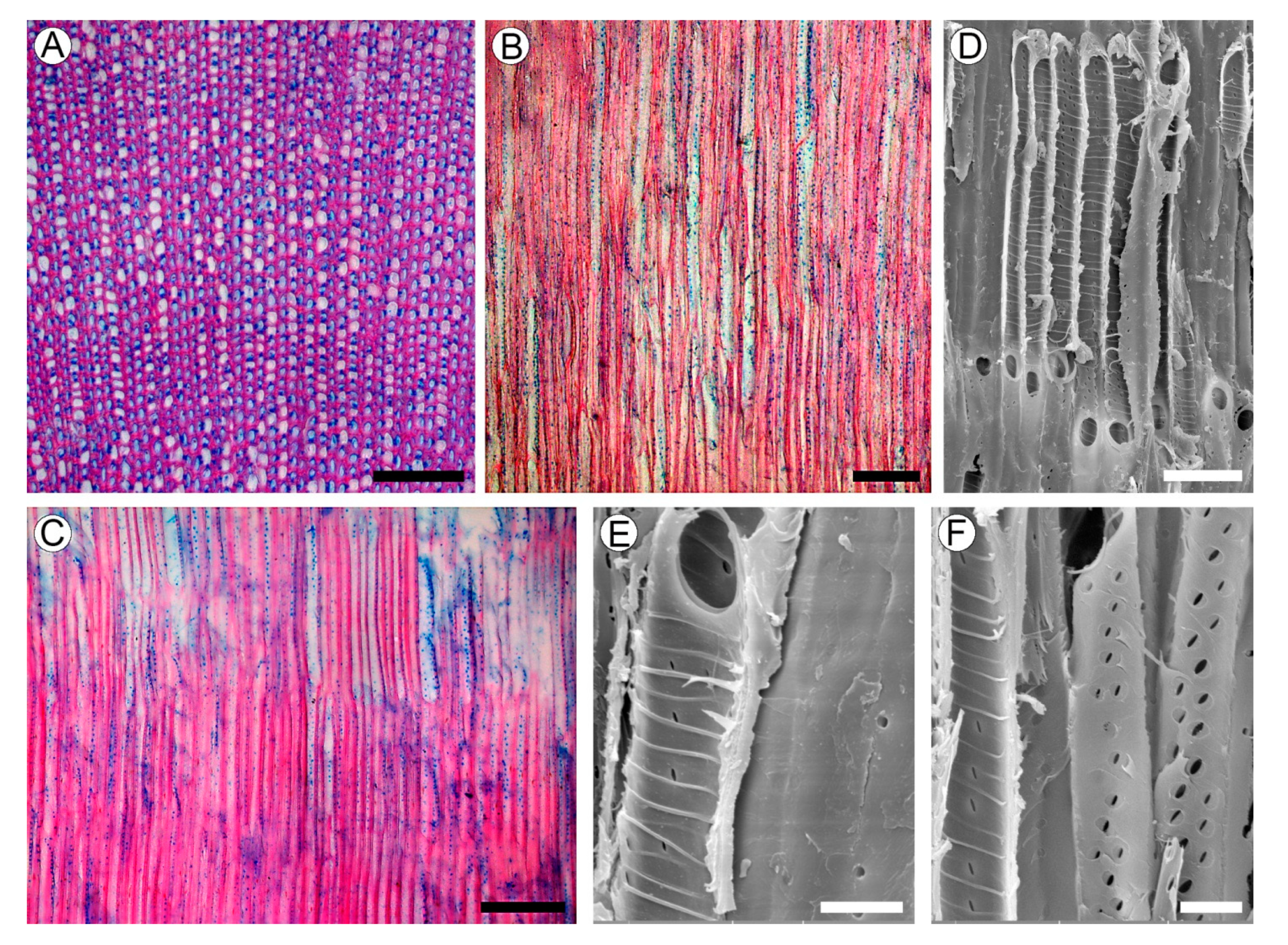
2.4. Wood Structure
2.5. Evolution of Rayless Wood within Plantaginaceae
3. Discussion
4. Materials and Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pennell, F.W. Taxonomy and distribution of Aragoa, and its bearing on the geological history of the Northern Andes. Proc. Acad. Nat. Sci. USA 1937, 89, 425–432. [Google Scholar]
- Fernández-Alonso, J.L. Novedades taxonómicas en Aragoa H.B.K. (Scrophulariaceae) y sinopsis del género. Anales Jard. Bot. 1993, 51, 73–96. [Google Scholar]
- Fernández-Alonso, J.L. Scrophulariaceae-Aragoeae. Flora 1995, 16, 1–224. [Google Scholar]
- Bello, M.A.; Rudall, P.J.; González, F.; Fernández-Alonso, J.L. Floral morphology and development in Aragoa (Plantaginaceae) and related members of the order Lamiales. Int. J. Plant Sci. 2004, 165, 723–738. [Google Scholar] [CrossRef]
- Sklenář, P.; Dušková, E.; Balslev, H. Tropical and temperate: Evolutionary history of páramo flora. Bot. Rev. 2011, 77, 71–108. [Google Scholar] [CrossRef]
- Albach, D.C.; Meudt, H.M.; Oxelman, B. Piecing together the “new” Plantaginaceae. Am. J. Bot. 2005, 92, 297–315. [Google Scholar] [CrossRef] [PubMed]
- Bello, M.A.; Chase, M.W.; Olmstead, R.; Rønsted, N.; Albach, D. The páramo endemic Aragoa is the sister genus of Plantago (Plantaginaceae; Lamiales): Evidence from plastid rbcL and nuclear ribosomal ITS sequence data. Kew Bull. 2002, 57, 585–597. [Google Scholar] [CrossRef]
- Mower, J.P.; Guo, W.; Partha, R.; Fan, W.; Levsen, N.; Wolff, K.; Nugent, J.M.; Pabón-Mora, N.; González, F. Plastomes from tribe Plantagineae (Plantaginaceae) reveal infrageneric structural synapormorphies and localized hypermutation for Plantago and functional loss of ndh genes from Littorella. Mol. Phylogenet. Evol. 2021, 162, 107217. [Google Scholar] [CrossRef]
- Hoggard, R.K.; Kores, P.J.; Molvray, M.; Hoggard, G.D.; Broughton, D.A. Molecular systematics and biogeography of the amphibious genus Littorella (Plantaginaceae). Am. J. Bot. 2003, 90, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Mennega, A.M.W. On unusual wood structures in Scrophulariaceae. Acta Bot. Neerl. 1975, 24, 359–360. [Google Scholar]
- InsideWood. 2004-Onwards. Available online: http://insidewood.lib.ncsu.edu/ (accessed on 10 April 2021).
- Lersten, N.R.; Curtis, J.D. Idioblasts and other unusual internal foliar secretory structures in Scrophulariaceae. Plant Syst. Evol. 2001, 227, 63–73. [Google Scholar] [CrossRef]
- Dill, F.E. Morphology of Veronicastrum virginicum. Trans. Kansas Acad. Sci. 1941, 44, 158–163. [Google Scholar] [CrossRef]
- Metcalfe, C.R.; Chalk, L. Anatomy of the Dicotyledons; Clarendon Press: Oxford, UK, 1950; Volume 2, pp. 1052–1059. [Google Scholar]
- Carlquist, S. Wood anatomy of insular species of Plantago and the problem of raylessness. Bull. Torrey Bot. Club 1970, 97, 353–361. [Google Scholar] [CrossRef]
- Carlquist, S. Living cells in wood. 2. Raylessness: Histology and evolutionary significance. Bot. J. Linn. Soc. 2015, 178, 529–555. [Google Scholar] [CrossRef]
- Schweingruber, F.H.; Börner, A.; Schulze, E.-D. Atlas of Stem Anatomy in Herbs, Shrubs und Trees; Springer: Berlin/Heidelberg, Germany, 2011; Volume 2, pp. 1–415. [Google Scholar]
- Schweingruber, F.; Landolt, W. The Xylem Database. Swiss Federal Institute for Forest, Snow and Landscape Research. 2010. Available online: https://www.wsl.ch/dendropro/xylemdb/ (accessed on 10 April 2021).
- Kaplan, A.; Hasanoğlu, A.; Ikbal, A.I. Morphological, anatomical and palynological properties of some Turkish Veronica L. species (Scrphulariaceae). Int. J. Bot. 2007, 3, 23–32. [Google Scholar] [CrossRef]
- Ida Christi, V.E.; Senthamarai, R. Qualitative and quantitative pharmacognostical studies on Scoparia dulcis Linn leaf. Int. J. Pharm. Pharmaceut. Res. 2015, 3, 57–74. [Google Scholar]
- Doležal, J.; Dvorský, M.; Börner, A.; Wild, J.; Schweingruber, F.H. Anatomy, Age and Ecology of High Mountain Plants in Ladakh, the Western Himalaya; Springer: Cham, Switzerland, 2018; p. 616. [Google Scholar]
- Carlquist, S. A theory of paedomorphosis in dicotyledonous woods. Phytomorphology 1962, 12, 30–45. [Google Scholar]
- Kostytschew, S. Der Bau und das Dickenwachstum der Dikotylenstämme. Ber. Dtsch. Bot. Ges. 1922, 40, 297–305. [Google Scholar]
- Kostytschew, S. Der Bau und das Dickenwachstum der Dikotylenstämme. Beih. Bot. Zentralbl. 1924, 40, 295–350. [Google Scholar]
- Helm, J. Untersuchungen über die Differenzierung der Sprossscheitelmeristeme von Dikotylen, unter besonderer Berücksichtig- ung des Prokambiums. Planta 1931, 15, 105–191. [Google Scholar] [CrossRef]
- Krumbiegel, A.; Kästner, A. Sekundäres Dickenwachstum von Sproß und Wurzel bei annuellen Dicotylen. Biosyst. Ecol. 1993, 4, 1–49. [Google Scholar]
- Hussain, K. Bacopa monnieri (L.) Lennell—A good biomarker of water pollution/contamination. J. Stress Physiol. Biochem. 2010, 6, 91–101. [Google Scholar]
- Hamed, K.A.; Hassan, S.A.; Mohamed, A.-S.H.; Hosney, N.K. Morphological and anatomical study on Plantaginaceae Juss. and some related taxa of Scrophulariaceae Juss. Egypt. J. Exp. Biol. Bot. 2014, 10, 135–146. [Google Scholar]
- Ahmed, E.M.; Desoukey, S.Y.; Fouad, M.A.; Kamel, M.S. A pharmacognostical study of Russelia equisetiformis Sch. & Cham. Int. J. Pharmacog. Phytochem. Res. 2016, 8, 174–192. [Google Scholar]
- Beck, C.B. An Introduction to Plant Structure and Development; Plant Anatomy for the Twenty-First Century; Cambridge University Press: Cambridge, CA, USA, 2010; pp. 1–464. [Google Scholar]
- Claßen-Bockhoff, R.; Franke, D.; Krämer, H. Early ontogeny defines the diversification of primary vascular bundle systems in angiosperms. Bot. J. Linn. Soc. 2021, 195, 281–307. [Google Scholar] [CrossRef]
- Cumbie, B.G. Development and structure of the xylem in Canavalia (Leguminosae). Bull. Torrey Bot. Club 1967, 94, 162–175. [Google Scholar] [CrossRef]
- Cumbie, B.G. Developmental changes in the xylem and vascular cambium of Apocynum sibiricum. Bull. Torrey Bot. Club 1969, 96, 629–640. [Google Scholar] [CrossRef]
- Soh, W.Y. Early ontogeny of vascular cambium II. Aucuba japonica and Weigela coraeensis. Bot. Mag. 1974, 87, 17–32. [Google Scholar]
- Soh, W.Y. Early ontogeny of vascular cambium III. Robinia pseudo-acacia and Syringa oblata. Bot. Mag. 1974, 87, 99–112. [Google Scholar] [CrossRef]
- Myśkow, E. Procambium–cambium transition during vascular meristem development in Diospyros lotus. Botany 2010, 88, 985–993. [Google Scholar] [CrossRef]
- Michener, D.S. Wood and leaf anatomy of Keckiella (Scrophulariaceae): Ecological considerations. Aliso 1981, 10, 39–57. [Google Scholar] [CrossRef][Green Version]
- Kalashnikova, O.A.; Bildanova, L.I.; Ryzhov, V.M.; Tarasenko, L.V. Morphological and anatomical analysis of Gratiola officinalis L. In Pharmaceutical Botany: Its Present State and Perspectives; Kurkin, V.A., Ed.; SamGMU: Samara, Russia, 2017; pp. 92–103. (In Russian) [Google Scholar]
- Pilger, R. Vergleichende Anatomie der Gattung Plantago mit Rücksicht auf die Existenzbedingungen. Bot. Jahrb. 1898, 25, 296–351. [Google Scholar]
- Filippa, E.; Barboza, G.E.; Luján, M.C.; Ariza Espinar, L. Anatomía foliar de las especies centro-argentinas de Plantago (Plantaginaceae). Darwiniana 1999, 37, 1–13. [Google Scholar]
- Esau, K. Origin and development of primary vascular tissues in seed plants. Bot. Rev. 1943, 9, 125–206. [Google Scholar] [CrossRef]
- Carlquist, S. Living cells in wood. 1. Absence, scarcity, and histology of axial parenchyma as keys to function. Bot. J. Linn. Soc. 2015, 177, 291–321. [Google Scholar] [CrossRef]
- Wang, J.C.; Pan, B.R.; Albach, D.C. Evolution of morphological and climatic adaptations in Veronica L. (Plantaginaceae). PeerJ 2016, 4, e2333. [Google Scholar] [CrossRef]
- Carlquist, S. Comparative Wood Anatomy. Systematic, Ecological, and Evolutionary Aspects of Dicotyledon Wood, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2001; pp. 1–448. [Google Scholar]
- Padrón, R.S.; Wilcox, B.P.; Crespo, P.; Célleri, R. Rainfall in the Andean Páramo: New insights from high-resolution monitoring in southern Ecuador. J. Hydrometeorol. 2015, 16, 985–996. [Google Scholar] [CrossRef]
- Cárdenas, M.F.; Tobón, C.; Rock, B.N.; del Valle, J.I. Ecophysiology of frailejones (Espeletia spp.), and its contribution to the hydrological functioning of páramo ecosystems. Plant Ecol. 2018, 219, 185–198. [Google Scholar] [CrossRef]
- Rada, F.; Azócar, A.; García-Núñez, C. Plant functional diversity in tropical Andean páramos. Plant Ecol. Divers. 2019, 12, 1–15. [Google Scholar] [CrossRef]
- Alves, E.S.; Angyalossy-Alfonso, V. Ecological trends in the wood anatomy of some Brazilian species 1. Growth rings and vessels. IAWA J. 2000, 21, 3–30. [Google Scholar] [CrossRef]
- Lindorf, H. Eco-anatomical wood features of species from a very dry tropical forest. IAWA J. 1994, 15, 361–376. [Google Scholar] [CrossRef]
- Baas, P.; Ewers, F.W.; Davies, S.D.; Wheeler, E.A. The evolution of xylem physiology. In Evolution of Plant Physiology from Whole Plants to Ecosystems; Hemsley, A.R., Poole, I., Eds.; Linnean Society Symposium Series No. 21; Elsevier Academic Press: London, UK, 2004; pp. 273–296. [Google Scholar]
- Wheeler, E.A.; Baas, P.; Rodgers, S. Variations in dicot wood anatomy: A global analysis based on the Insidewood database. IAWA J. 2007, 28, 229–258. [Google Scholar] [CrossRef]
- Hargrave, K.R.; Kolb, K.J.; Ewers, F.W.; Davis, S.D. Conduit diameter and drought-induced embolism in Salvia mellifera Greene (Labiatae). New Phytol. 1994, 126, 695–705. [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S.; Wheeler, J.K.; Castro, L. Scaling of angiosperm xylem structure with safety and efficiency. Tree Physiol. 2006, 26, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Tyree, M.T. The impact of vessel size in vulnerability curves: Data and models for within-species variability in saplings of aspen, Populus tremuloides Michx. Plant Cell Environ. 2010, 33, 1059–1069. [Google Scholar] [CrossRef]
- Lens, F.; Sperry, J.S.; Christman, M.A.; Choat, B.; Rabaey, D.; Jansen, S. Testing hypotheses that link wood anatomy to cavitation resistance and hydraulic conductivity in the genus Acer. New Phytol. 2011, 190, 709–723. [Google Scholar] [CrossRef]
- Jacobsen, A.L.; Pratt, R.B.; Venturas, M.D.; Hacke, U.G. Large volume vessels are vulnerable to water-stress-induced embolism in stems of poplar. IAWA J. 2019, 40, 4–22. [Google Scholar] [CrossRef]
- Roth-Nebelsick, A.; Speck, T. Mechanical and hydrodynamic properties of vessels with tertiary helical thickenings: New information about possible functional relationships. In Plant Biomechanics; Spatz, H.-C., Speck, T., Eds.; Thieme: Stuttgart, Germany, 2000; pp. 265–271. [Google Scholar]
- Kedrov, G.B. Functioning wood. Wulfenia 2012, 19, 57–95. [Google Scholar]
- Morris, H.; Plavcová, L.; Cvecko, P.; Fichtler, E.; Mark, A.F.; Gillingham, M.A.F.; Martínez-Cabrera, H.I.; McGlinn, D.J.; Wheeler, E.; Zheng, J.M.; et al. A global analysis of parenchyma tissue fractions in secondary xylem of seed plants. New Phytol. 2016, 209, 1553–1565. [Google Scholar] [CrossRef]
- Evans, D.E. Aerenchyma formation. New Phytol. 2003, 161, 35–49. [Google Scholar] [CrossRef]
- Andrzejewska, E.; Świętosławski, I. The morphology of hairs in species of Plantago L. sections: Leucopsyllium Decne. and Hymenopsyllium Pilger. Acta Soc. Bot. Poloniae 1988, 57, 9–19. [Google Scholar] [CrossRef][Green Version]
- Andrzejewska, E.; Świętosławski, I. The morphology of hairs in species of Plantago L. sections: Bauphula Decne. and Arnoslossum Decne. Acta Soc. Bot. Poloniae 1989, 58, 15–45. [Google Scholar] [CrossRef][Green Version]
- Andrzejewska, E.; Świętosławski, I. Hair anatomy in Plantago subg. Psyllium (Plantaginaceae). Plant Syst. Evol. 1993, 184, 113–123. [Google Scholar] [CrossRef]
- Johansen, D.A. Plant Microtechnique; McGraw-Hill: New York, NY, USA, 1940; pp. 1–525. [Google Scholar]
- Exley, R.R.; Meylan, B.A.; Butterfield, B.G. A technique for obtaining clear cut surfaces on wood samples prepared for the scanning electron microscope. J. Microsc. 1977, 110, 75–78. [Google Scholar] [CrossRef]
- IAWA Committee. IAWA list of microscopic features for hardwood identification. IAWA Bull. 1989, 10, 219–332. [Google Scholar]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 2.75. 2011. Available online: http://mesquiteproject.org (accessed on 12 February 2021).
- Feder, N.; O’Brien, T.P. Plant microtechnique: Some principles and new methods. Am. J. Bot. 1968, 55, 123–142. [Google Scholar] [CrossRef]
- Cheadle, V.I.; Gifford, E.M.; Esau, K. A staining combination for phloem and contiguous tissues. Stain Technol. 1953, 28, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Angyalossy, V.; Pace, M.R.; Evert, R.F.; Marcati, C.R.; Oskolski, A.A.; Terrazas, T.; Kotina, E.; Lens, F.; Mazzoni-Viveiros, S.C.; Angeles, G.; et al. IAWA list of microscopic bark features. IAWA J. 2016, 37, 517–615. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oskolski, A.; Vuza, N.; Shipunov, A. Stem and Leaf Anatomy of Aragoa (Plantaginaceae): In Search of Lost Rays. Plants 2021, 10, 1773. https://doi.org/10.3390/plants10091773
Oskolski A, Vuza N, Shipunov A. Stem and Leaf Anatomy of Aragoa (Plantaginaceae): In Search of Lost Rays. Plants. 2021; 10(9):1773. https://doi.org/10.3390/plants10091773
Chicago/Turabian StyleOskolski, Alexei, Nathi Vuza, and Alexey Shipunov. 2021. "Stem and Leaf Anatomy of Aragoa (Plantaginaceae): In Search of Lost Rays" Plants 10, no. 9: 1773. https://doi.org/10.3390/plants10091773
APA StyleOskolski, A., Vuza, N., & Shipunov, A. (2021). Stem and Leaf Anatomy of Aragoa (Plantaginaceae): In Search of Lost Rays. Plants, 10(9), 1773. https://doi.org/10.3390/plants10091773