

The Application of Phytohormones as Biostimulants in Corn Smut Infected Hungarian Sweet and Fodder Corn Hybrids

 ,
,  ,
,  ,
,  ,
, 
Abstract
:1. Introduction
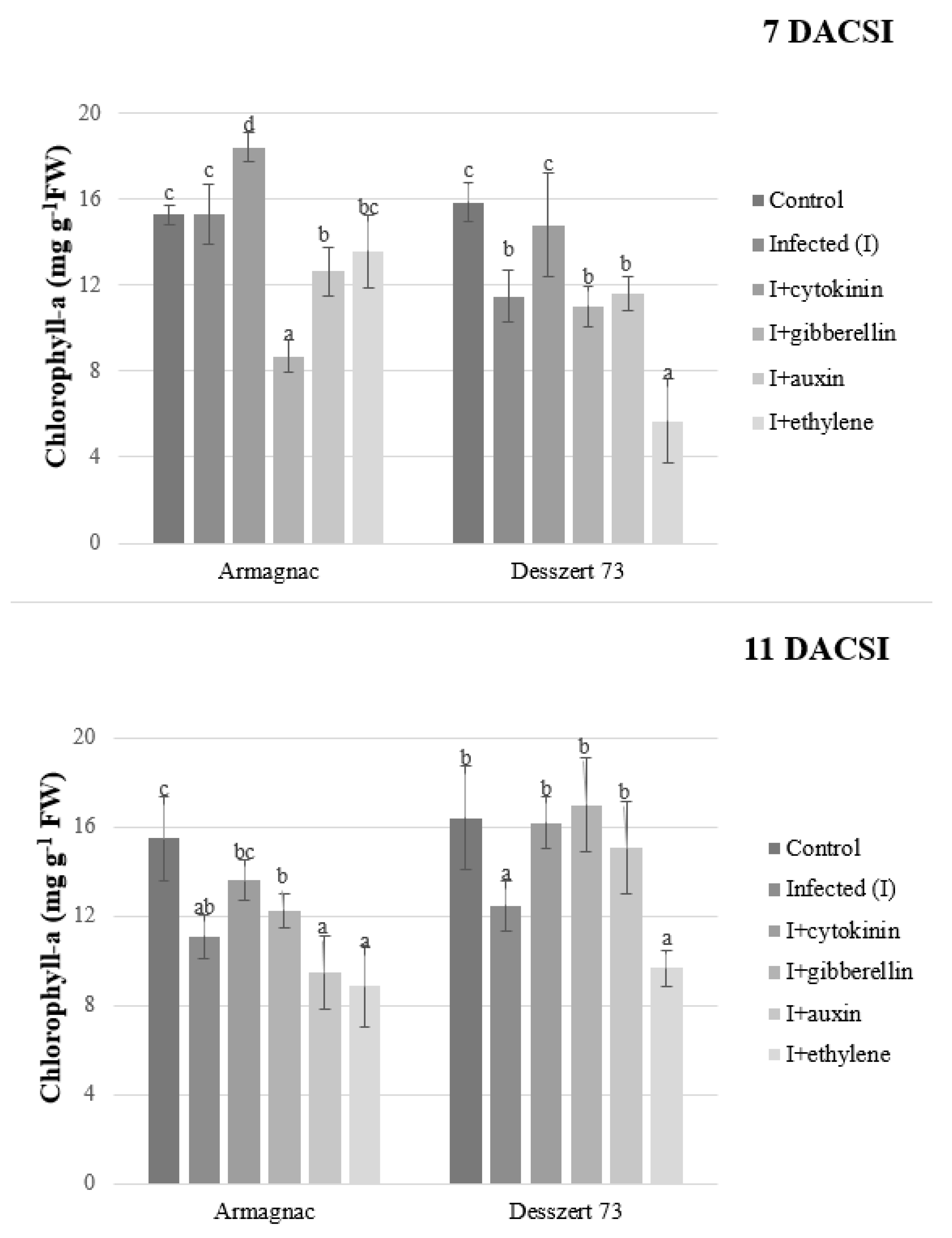
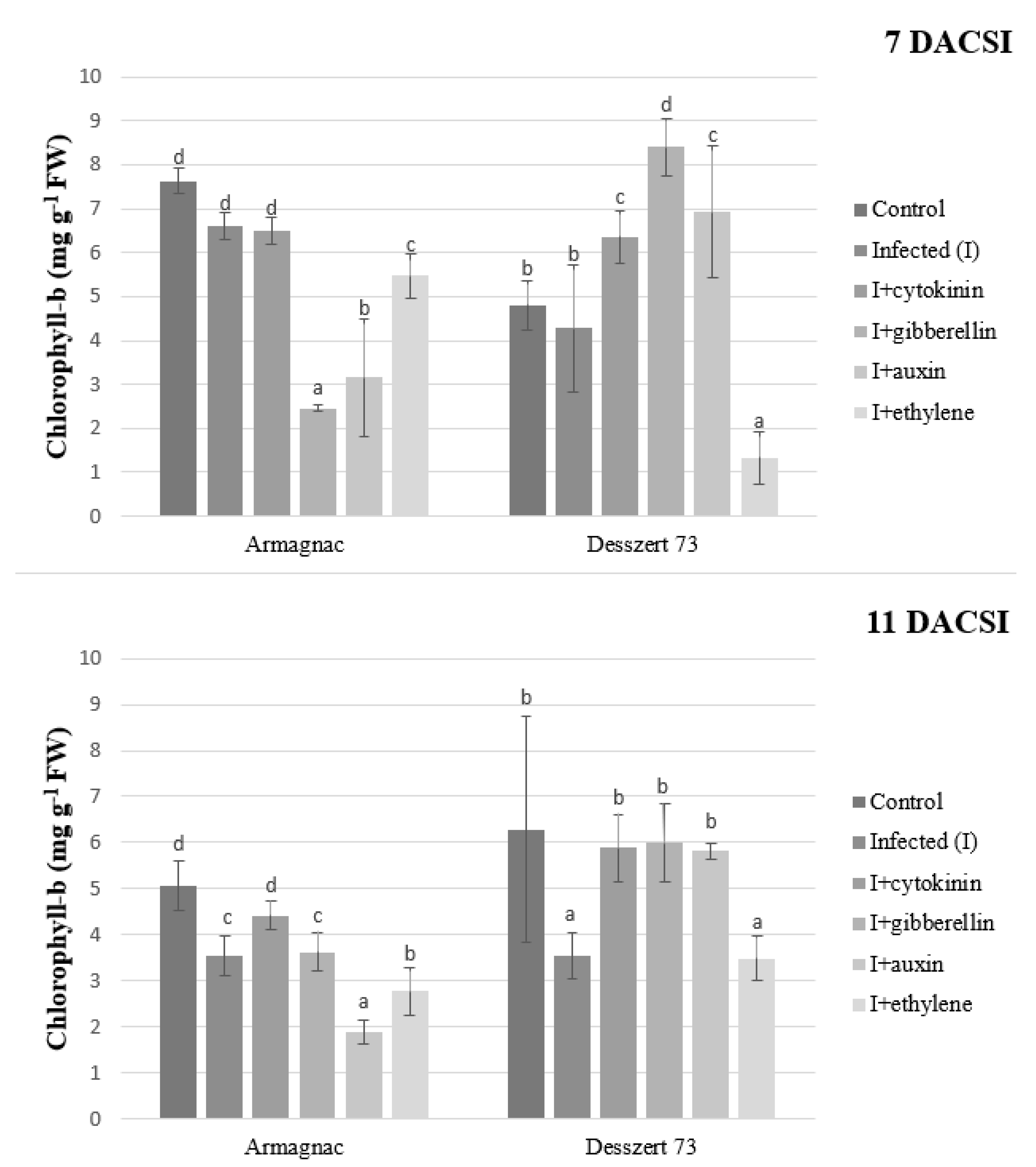
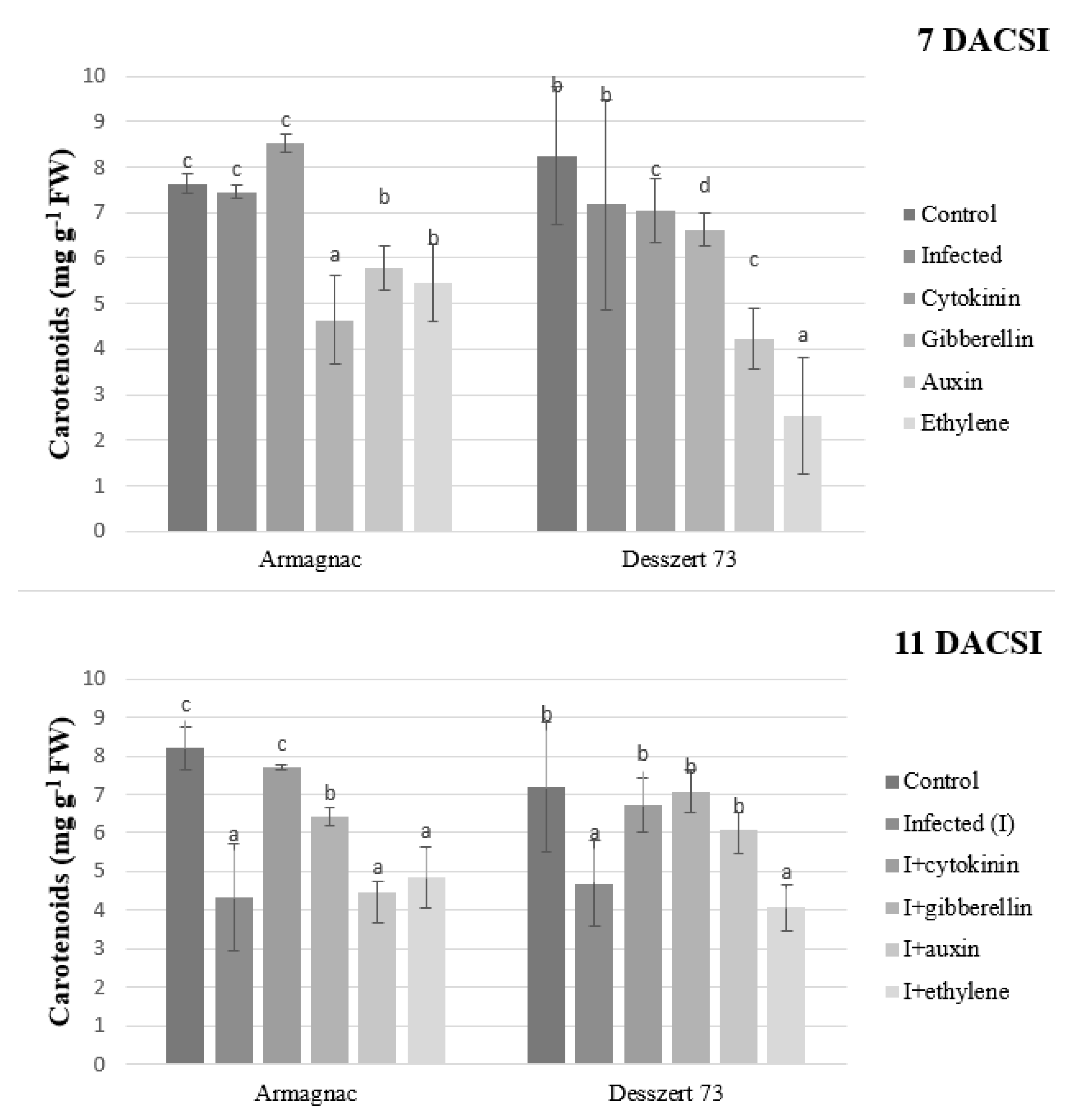
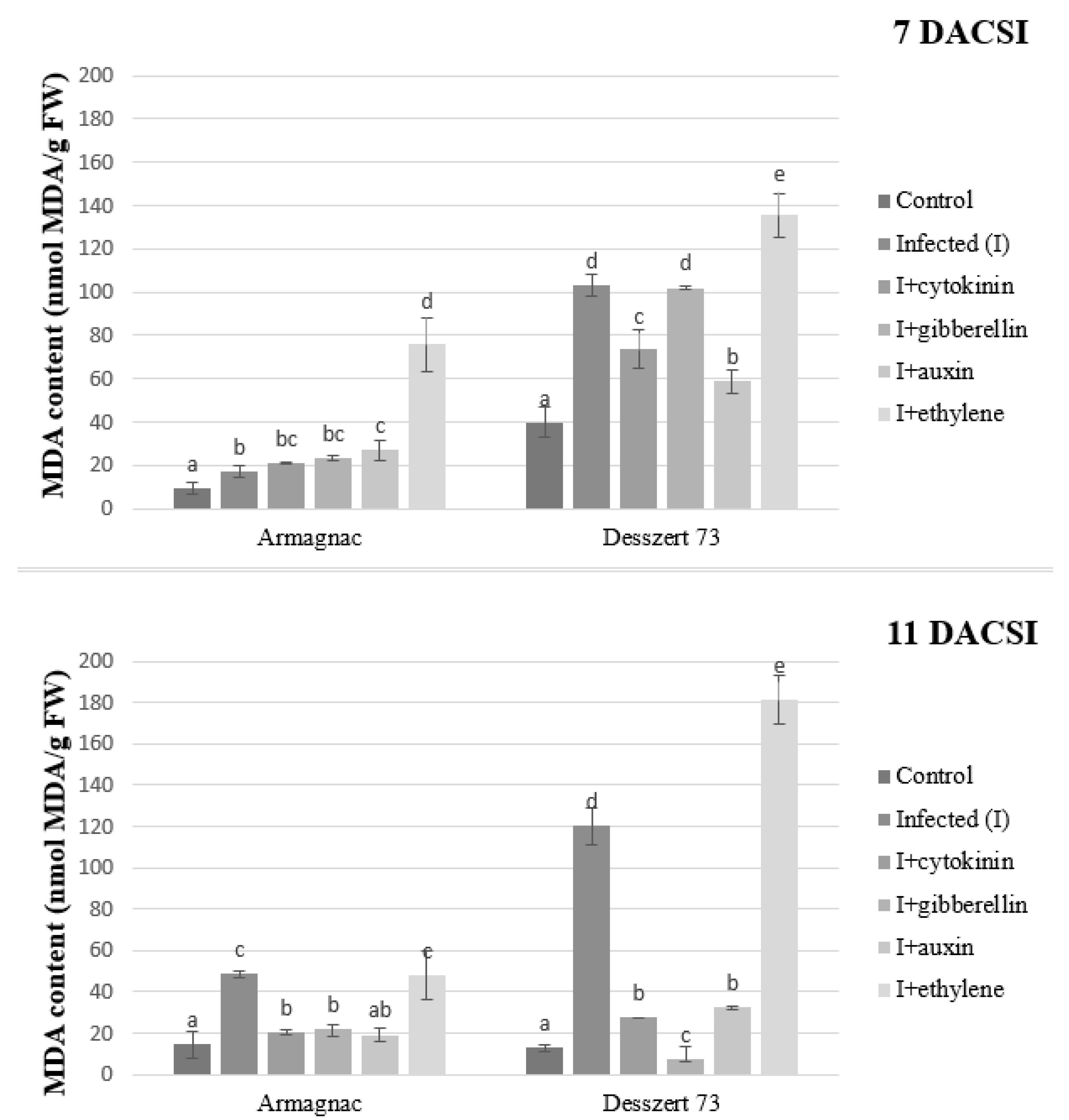
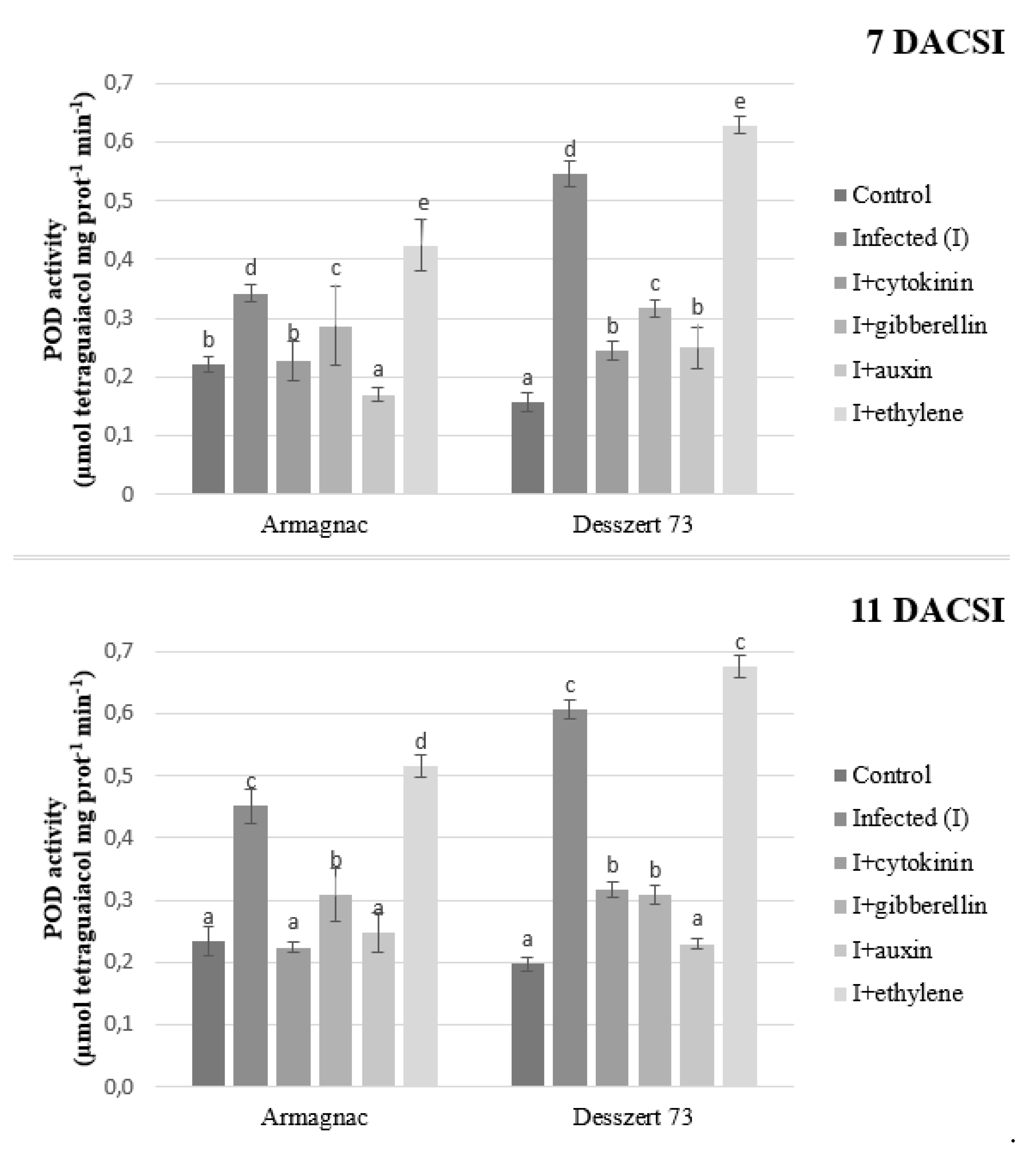
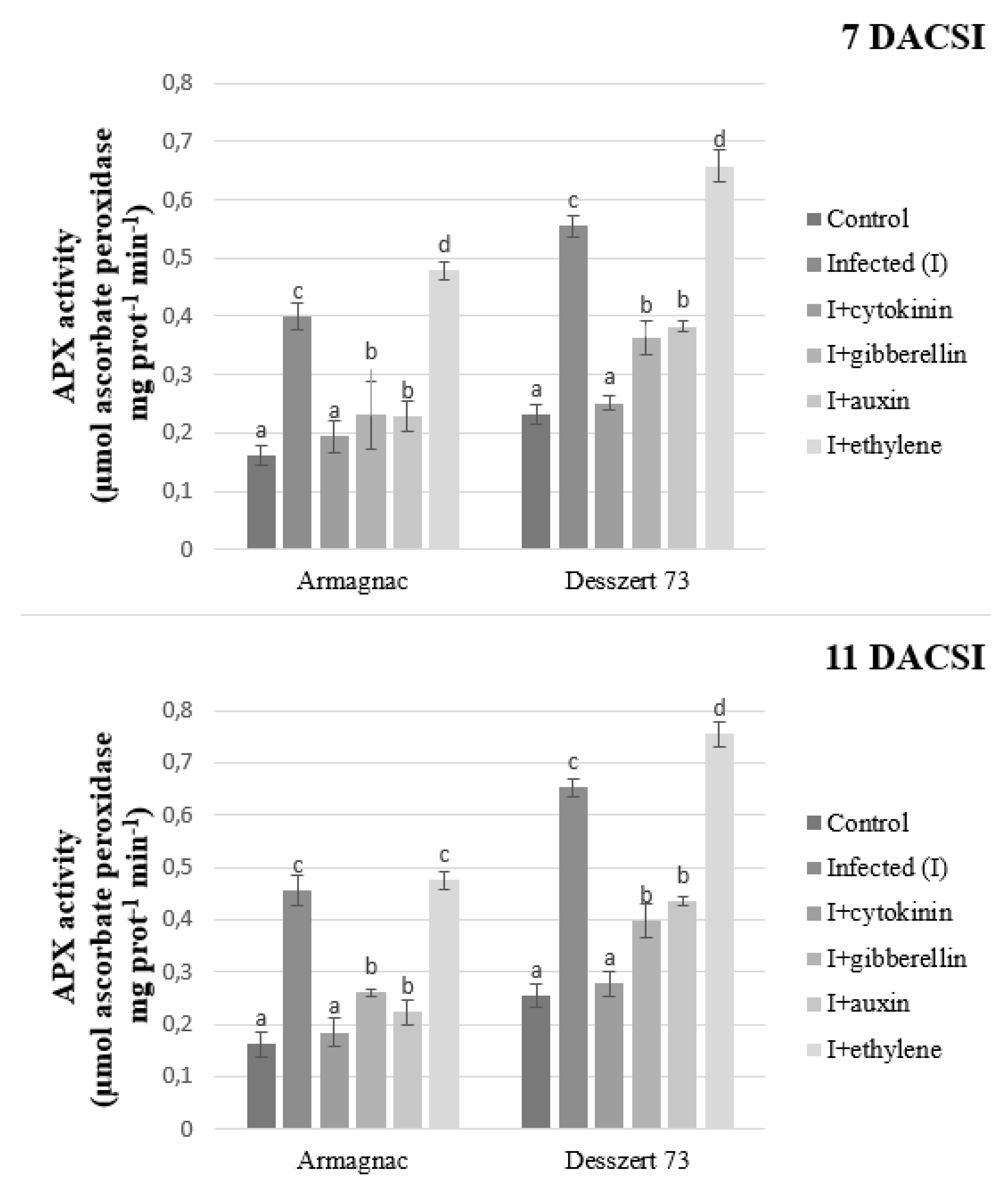
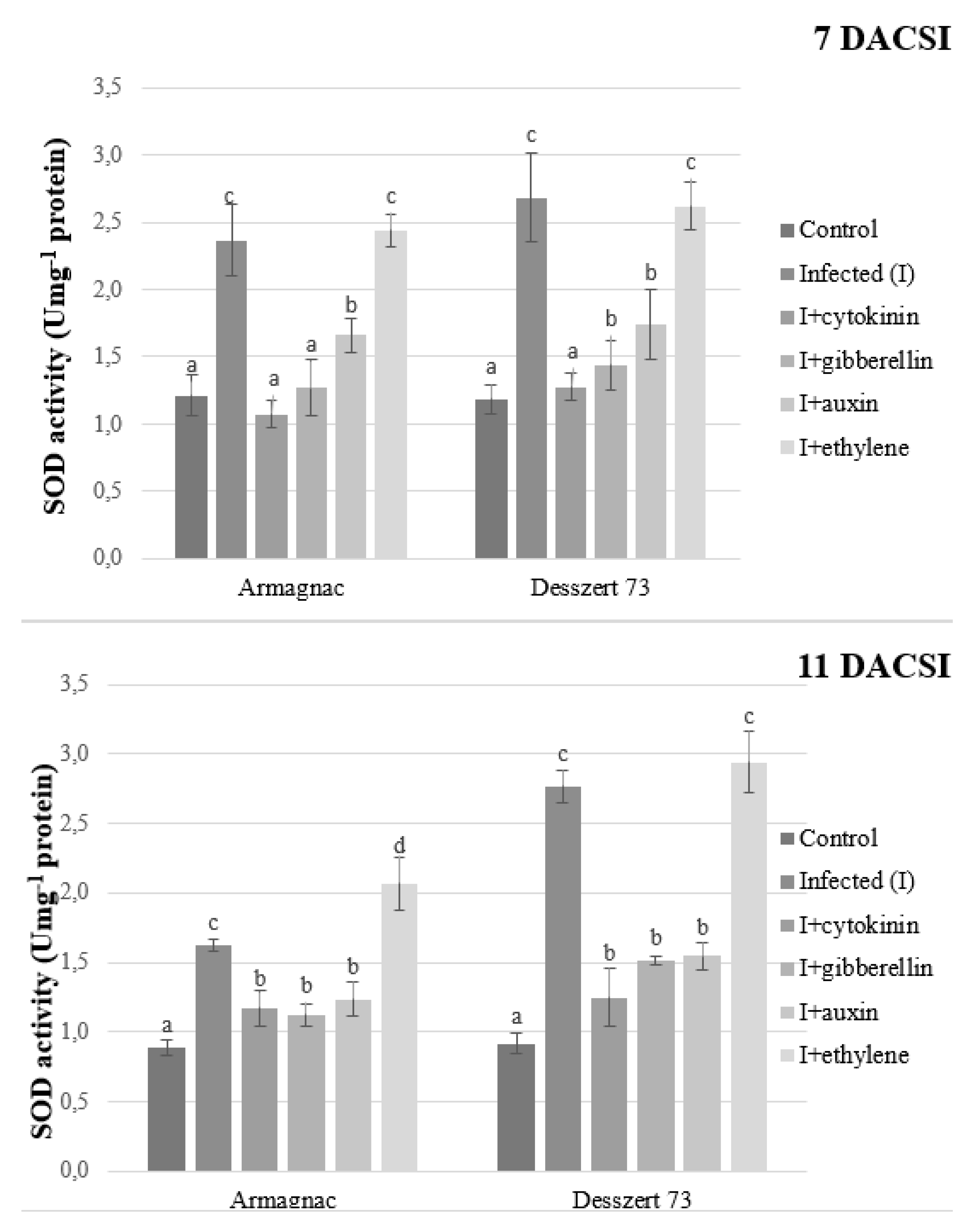
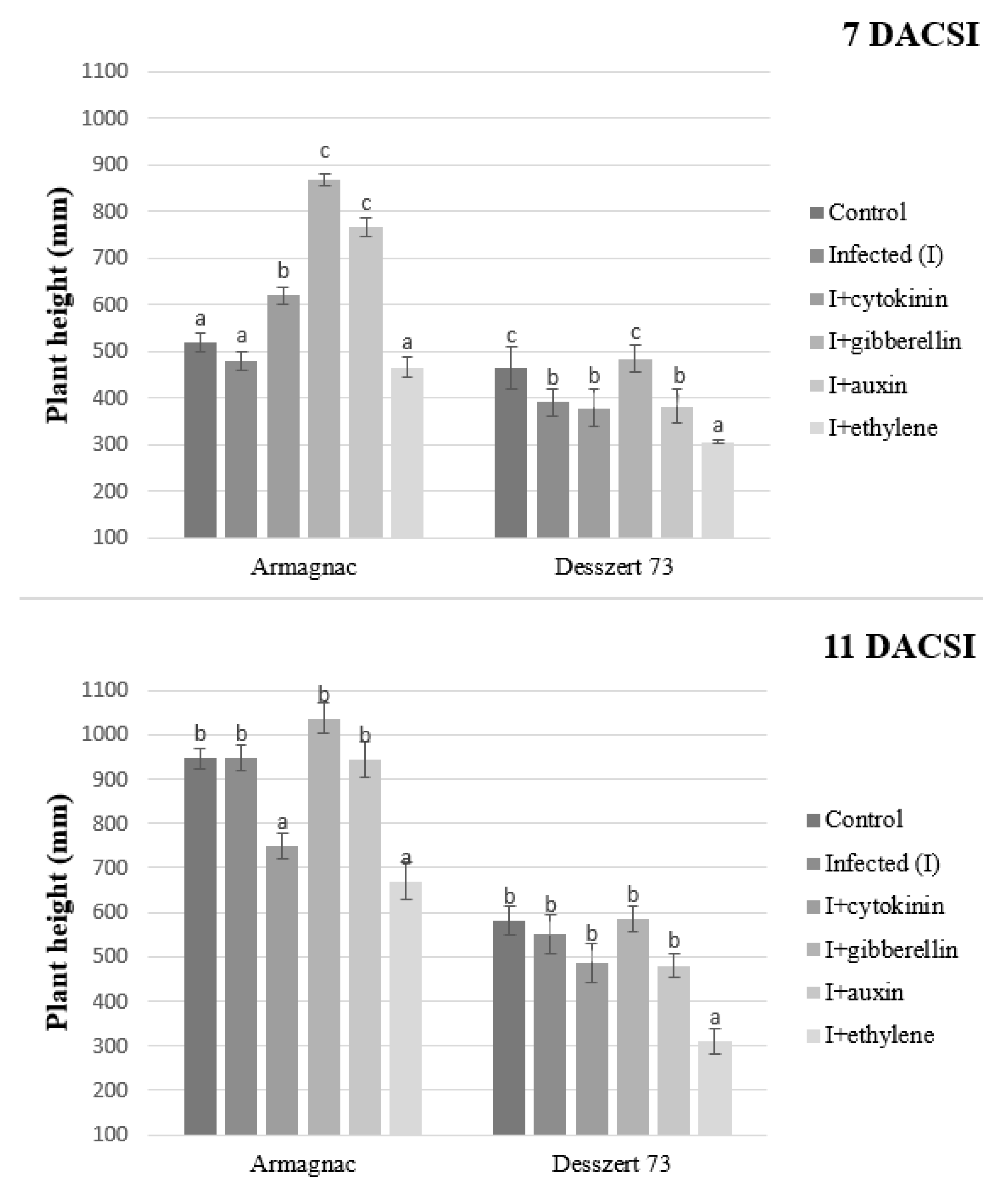
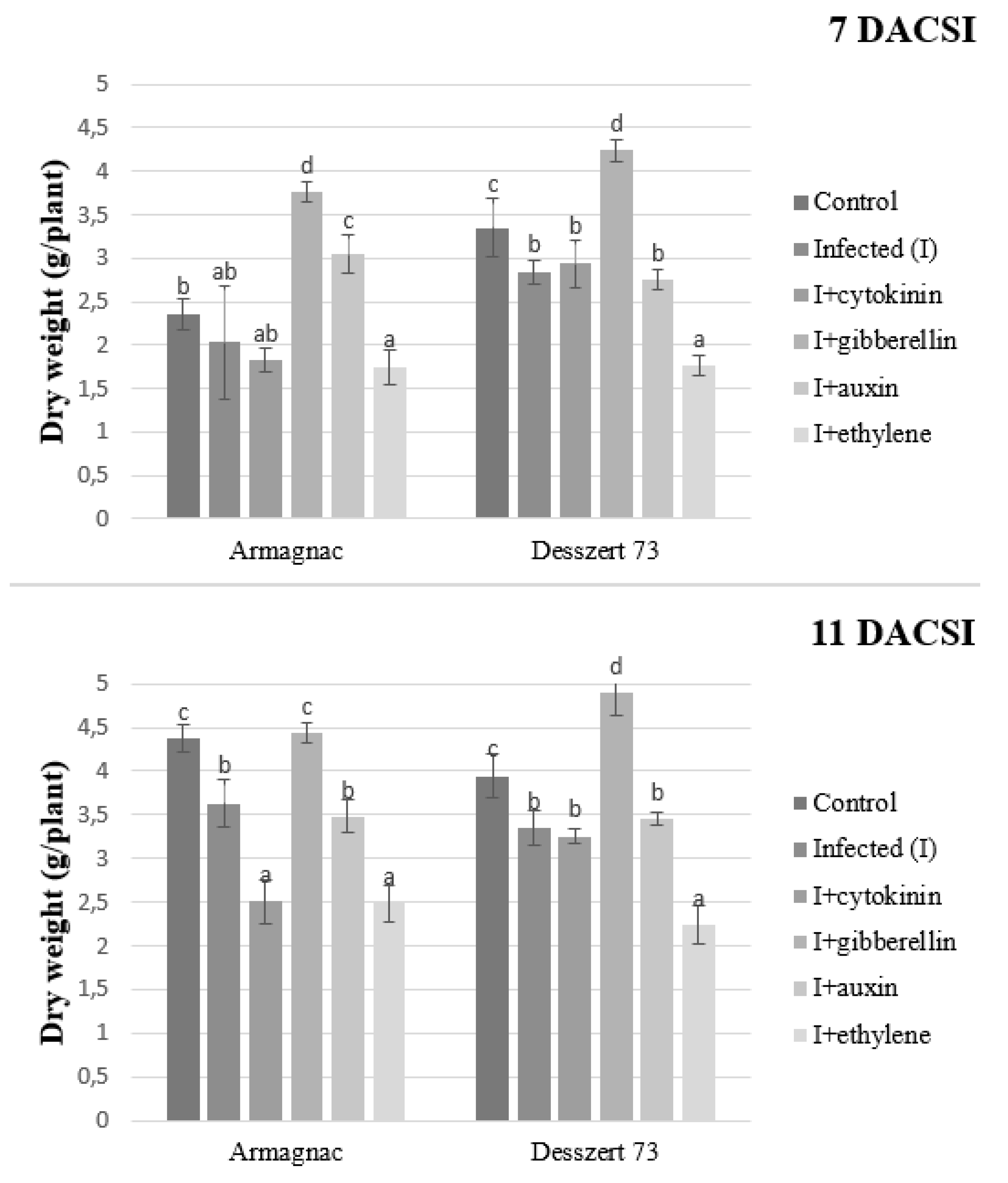
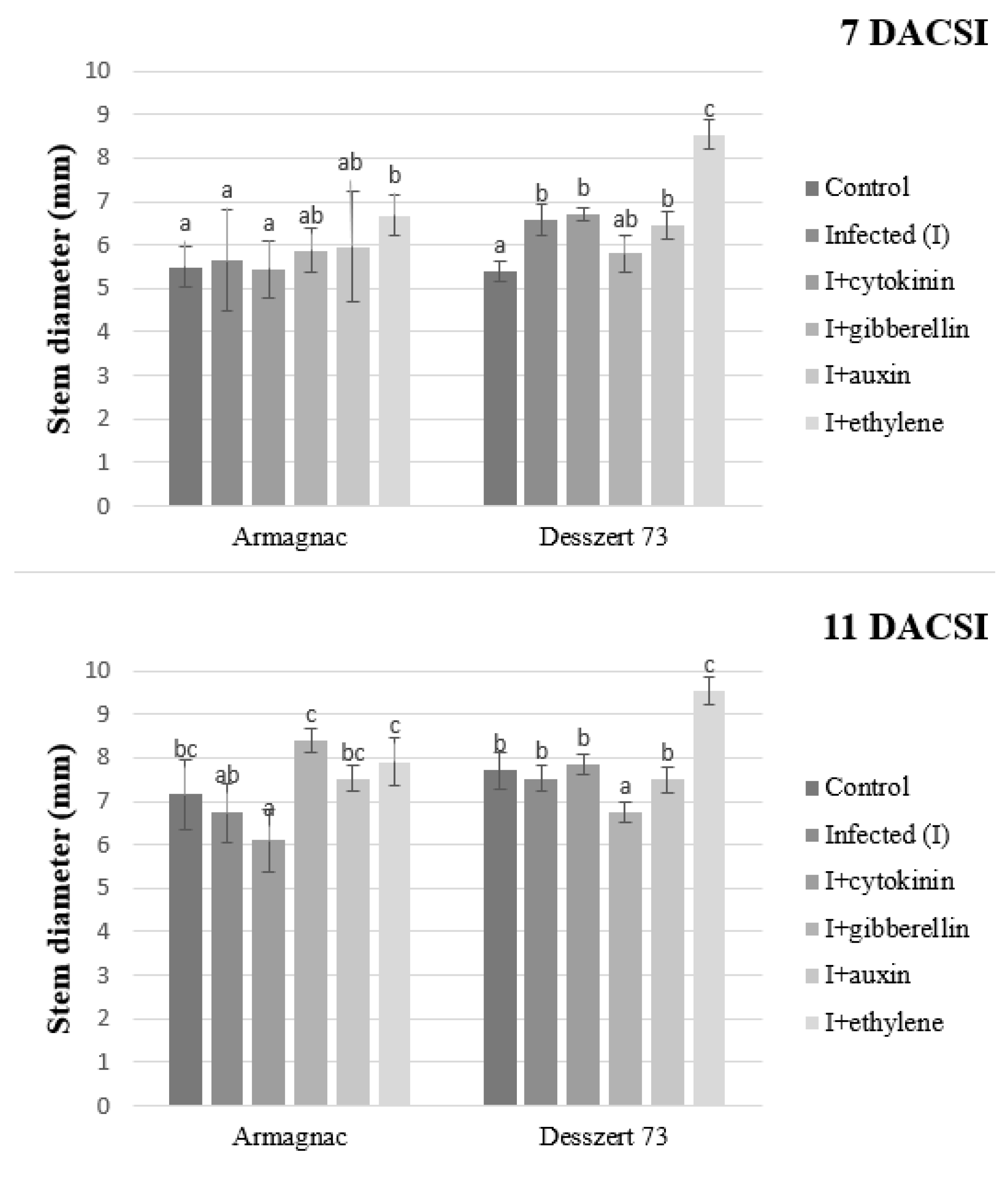
2. Results
3. Discussion
4. Materials and Methods
4.1. Growing of Plants and Applied Treatments
4.2. Photosynthetic Pigment Quantification
4.3. Morphological Parameters
4.4. Determination of the Level of Lipid Peroxidation
4.5. Activity of Antioxidant Enzymes
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Knutson, T.; Zhang, R.; Horovitz, L.W. Prospects for a prolonged slowdown in global warming in the early 21st century. Nat. Commun. 2016, 7, 13676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malhi, G.S.; Kaur, M.; Kaushik, P. Impact of Climate Change on Agriculture and Its Mitigation Strategies: A Review. Sustainability 2021, 13, 1318. [Google Scholar] [CrossRef]
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; Senthil-Kumar, M. Impact of Combined Abiotic and Biotic Stresses on Plant Growth and Avenues for Crop Improvement by Exploiting Physio-morphological Traits. Front. Plant Sci. 2017, 8, 537. [Google Scholar] [CrossRef] [Green Version]
- Hough, A.R.; Nechols, J.R.; McCornack, B.P.; Margolies, D.C.; Sandercock, B.K.; Yan, D.; Murray, L. The Effect of Temperature and Host Plant Resistance on Population Growth of the Soybean Aphid Biotype 1 (Hemiptera: Aphididae). Environ. Entomol. 2016, 46, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.P.; Leach, J.E. High temperature-induced plant disease susceptibility: More than the sum of its parts. Curr.Opin. Plant Biol. 2020, 56, 235–241. [Google Scholar] [CrossRef]
- Gagnon, A.È.; Bourgeois, G.; Bourdages, L.; Grenier, P.; Blondlot, A. Impact of climate change on Ostrinia nubilalis (Lepidoptera: Crambidae) phenology and its implications on pest management. Agric. For. Entomol. 2019, 21, 253–264. [Google Scholar] [CrossRef]
- Sharma, H.C.; Sullivan, D.; Bhatnagar, V.S. Population dynamics and natural mortality factors of the Oriental armyworm, Mythimna separata (Lepidoptera: Noctuidae), in South-Central India. Crop Prot. 2002, 21, 721–732. [Google Scholar] [CrossRef] [Green Version]
- Nazarov, P.A.; Baleev, D.N.; Ivanova, M.I.; Sokolova, L.M.; Karakozova, M.V. Infectious Plant Diseases: Etiology, Current Status, Problems and Prospects in Plant Protection. Actanaturae 2020, 12, 46–59. [Google Scholar] [CrossRef]
- Váry, Z.; Mullins, E.; McElwain, J.C.; Doohan, F.M. The severity of wheat diseases increases when plants and pathogens are acclimatized to elevated carbon dioxide. Glob. Change Biol. 2015, 21, 2661–2669. [Google Scholar] [CrossRef]
- Niu, C.; Yu, D.; Wang, Y.; Ren, H.; Jin, Y.; Zhou, W.; Li, B.; Cheng, Y.; Yue, J.; Gao, Z.; et al. Common and pathogen-specific virulence factors are different in function and structure. Virulence 2013, 4, 473–482. [Google Scholar] [CrossRef]
- Elsharkawy, M.M.; El-Khateeb, N.M.M. Antifungal activity and resistance induction against Sclerotium cepivorum by plant growth-promoting fungi in onion plants. Egypt. J. Biol. Pest Co. 2019, 29, 1–9. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Yang, Y.; Zhong, C.; Notaguchi, M.; Yu, W. A Susceptible Scion Reduces Rootstock Tolerance to Ralstonia solanacearum in Grafted Eggplant. Horticulturae 2019, 5, 78. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Montiel, L.G.; Chiquito-Contreras, C.J.; Murillo-Amador, B.; Vida- Hernández, L.; Quiñones-Aguilar, E.E.; Chiquito-Contreras, R.G. Efficiency of two inoculation methods of Pseudomonas putida on growth and yield of tomato plants. J. Soil Sci.Plant Nutr. 2017, 17, 1003–1012. [Google Scholar] [CrossRef] [Green Version]
- Meena, M.; Zehra, A.; Dubey, M.K.; Aamir, M.; Gupta, V.K.; Upadhyay, R.S. Comparative Evaluation of Biochemical Changes in Tomato (Lycopersicon esculentum Mill.). Infected by Alternaria alternata and Its Toxic Metabolites (TeA, AOH, and AME). Front. Plant Sci. 2016, 7, 1408. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Yao, S.; Ming, J.; Deng, L.; Zeng, K. Variations in chlorophyll and carotenoid contents and expression of genes involved in pigment metabolism response to oleocellosis in citrus fruits. Food Chem. 2019, 272, 49–57. [Google Scholar] [CrossRef]
- Debona, D.; Rodrigues, F.Á.; Rios, J.A.; Nascimento, K.J. Biochemical changes in the leaves of wheat plants infected by Pyricularia oryzae. Biochem. Cell Biol. 2012, 102, 1121–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Wan, X.; Shah, F.; Fahad, S.; Huang, J. The role of antioxidant enzymes in adaptive responses to sheath blight infestation under different fertilization rates and hill densities. Sci. World J. 2014, 2014, 502134. [Google Scholar] [CrossRef]
- Barna, B.; Fodor, J.; Harrach, B.D.; Pogány, M.; Király, Z. The Janus face of reactive oxygen species in resistance and susceptibility of plants to necrotrophic and biotrophic pathogens. Plant Physiol. Biochem. 2012, 59, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqur, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Monteiro, A.; Resende, M.; Valente, T.; Junior, P.; Pereira, V.; Costa, J.; Silva, J. Manganese Phosphite in Coffee Defence against Hemileia vastatrix, the Coffee Rust Fungus: Biochemical and Molecular Analyses. J. Phytopathol. 2016, 164, 1043–1053. [Google Scholar] [CrossRef] [Green Version]
- Faize, M.; Burgos, L.; Faize, L.; Petri, C.; Barba-Espin, G.; Díaz-Vivancos, P.; Clemente-Moreno, M.J.N.; Alburquerque, N.; Hernández, J.A. Modulation of tobacco bacterial disease resistance using cytosolic ascorbate peroxidase and Cu, Zn-superoxide dismutase. Plant Pathol. 2011, 61, 858–866. [Google Scholar] [CrossRef]
- Nowogórska, A.; Patykowski, J. Selected reactive oxygen species and antioxidant enzymes in common bean after Pseudomonas syringae pv. phaseolicola and Botrytis cinerea infection. Acta Physiol. Plant. 2015, 37, 1725. [Google Scholar] [CrossRef] [Green Version]
- Hameed, S.; Akhtar, K.P.; Hammed, A.; Gulzar, T.; Kiran, S.; Yousaf, S.; Abbas, G.; Asghar, M.J.; Sarwar, N. Biochemical changes in the leaves of mung bean (Vigna radiate) plants infected by phytoplasma. Turkish J. Biochem. 2017, 42, 591–599. [Google Scholar] [CrossRef]
- Dong, X. SA, JA ethylene, and disease resistance in plants. Curr. Opin. Plant Biol. 1998, 1, 316–323. [Google Scholar] [CrossRef]
- Díaz, J.; Ten Have, A.; van Kan, J.A.L. The role of ethylene and wound signaling in resistance of tomato to Botrytis cinerea. Plant Physiol. 2002, 129, 1341–1351. [Google Scholar] [CrossRef] [Green Version]
- Van Loon, L.C.; Geraats, B.P.J.; Linthorst, H.J.M. Ethylene as a modulator of disease resistance in plants. Trends Plant Sci. 2006, 11, 184–191. [Google Scholar] [CrossRef]
- Yang, C.; Li, W.; Cao, J.; Meng, F.; Yu, Y.; Huang, J.; Jiang, L.; Liu, M.; Zhang, Z.; Chen, X.; et al. Activation of ethylene signaling pathways enhances disease resistance by regulating ROS and phytoalexin production in rice. Plant J. 2016, 89, 338–353. [Google Scholar] [CrossRef] [Green Version]
- Xiong, X.P.; Sun, S.C.; Zhang, X.Y.; Li, Y.J.; Liu, F.; Zhu, Q.H.; Xue, F.; Sun, J. GhWRKY70D13 regulates resistance to Verticillium dahliae in cotton through the ethylene and jasmonic acid signaling pathways. Front. Plant Sci. 2020, 11, 69. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; van der Does, D.; Zamiousdis, C.; Leon-Reyes, A.; van Wees, S.C.M. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [Green Version]
- Denancé, N.; Sánchez-Vallet, A.; Goffner, D.; Molina, A. Disease resistance or growth: The role of plant hormones in balancing immune responses and fitness costs. Front. Plant Sci. 2013, 4, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurny, A.; Cuesta, C.; Cavallari, N.; Ötvös, K.; Duclercq, J.; Dokládal, L.; Montesinos, J.C.; Gallemí, M.; Semerádová, H.; Rauter, T.; et al. Synergistic on auxin and cytokinin1 positively regulates growth and attenuates soil pathogen resistance. Nat. Commun. 2020, 11, 2170. [Google Scholar] [CrossRef]
- Schepetilnikov, M.; Ryabova, L.A. Auxin signaling in regulation of plant translation reinitiation. Front. Plant Sci. 2017, 8, 1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, J.; Wang, S. Insights into auxin signaling in plant-pathogen interactions. Front. Plant Sci. 2011, 2, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esquerré-Tugayé, M.T.; Lafitte, C.; Mazau, D.; Toppan, A.; Touzé, A. Cell surfaces in plant-microorganism interactions: II. Evidence for the accumulation of hydroxyproline-rich glycoproteins in the cell wall of diseased plants as a defense mechanism. Plant Physiol. 1979, 64, 320–326. [Google Scholar] [CrossRef] [Green Version]
- Van Loon, L.C.; Pennings, G.G.H. Involvement of ethylene in the induction of systemic acquired resistance in tobacco. In Mechanisms of Plant Defense Responses; Fritig, B., Legrand, M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1993; pp. 156–159. [Google Scholar]
- Schaller, G.E.; Street, I.H.; Kieber, J.J. Cytokinin and the cell cycle. Curr. Opin. Plant Biol. 2014, 12, 7–15. [Google Scholar] [CrossRef]
- Hönig, M.; Plíhalová, L.; Husicková, A.; Nisler, J.; Dolezal, K. Role of cytokinins in senescence, antioxidant defense and photosynthesis. Int. J. Mol. Sci. 2018, 19, 4045. [Google Scholar] [CrossRef] [Green Version]
- Sakakibara, H. Cytokinins: Activity biosynthesis, and translocation. Annu. Rev. Plant Biol. 2006, 57, 431–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhtar, S.S.; Mekureyaw, M.F.; Pandey, C.; Roitsch, T. Role of cytokinins for interactions of plants with microbial pathogens and pest insects. Front. Plant Sci. 2020, 10, 1777. [Google Scholar] [CrossRef] [Green Version]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defense responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef]
- Galicia-García, P.R.; Silva-Rojas, H.V.; Mendoza-Onofre, L.E.; Zavaleta-Mancera, H.A.; Córdova-Téllez, L.; Espinosa-Calderón, A. Selection of aggressive pathogenic and solopathogenic strains of Ustilago maydis to improve Huitlacoche production. Acta Bot. Bras. 2016, 30, 683–692. [Google Scholar] [CrossRef] [Green Version]
- Frommer, D.; Veres, S.; Radócz, L. Susceptibility of stem infected sweet corn hybrids to common smut disease. Acta Agr. Debr. 2018, 74, 55–57. [Google Scholar] [CrossRef] [PubMed]
- Morrison, E.; Emery, R.J.; Saville, B. Fungal derived cytokinins are necessary for normal Ustilago maydis infection of corn. Plant Pathol. 2017, 66, 726–742. [Google Scholar] [CrossRef]
- Moura, R.M.; Pedrosa, E.M.; Guimares, L.M.P. A rare syndrome of corn smut. Fitopatol. Bras. 2001, 26, 782. [Google Scholar] [CrossRef]
- Lambie, S.C.; Kretschmer, M.; Croll, D.; Haslam, T.M.; Kunst, L.; Klose, J.; Kronstad, J.W. The putative phospholipase Lip2 counteracts oxidative damage and influences the virulence of Ustilago maydis. Mol. Plant Pathol. 2017, 18, 210–221. [Google Scholar] [CrossRef] [Green Version]
- Hemetsberger, C.; Herrberger, C.; Zechmann, B.; Hillmer, M.; Doehlemann, G. The Ustilago maydis effector Pep1 suppresses plant immunity by inhibition of host peroxidase activity. PLoS Pathog. 2012, 8, e1002684. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Choi, D.; Lee, S.; Ryu, C.M.; Hwang, I. Cytokinins and plant immunity: Old foes or new friends? Trends Plant Sci. 2011, 16, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Bölker, M.; Basse, C.W.; Schirawski, J. Ustilago maydis secondary metabolism-from genomics to biochemistry. Fungal Genet Biol. 2008, 45, S88–S93. [Google Scholar] [CrossRef]
- Morrison, E.N.; Emery, R.J.; Saville, B.J. Phytohormone involvement in the Ustilago maydis—Zea mays pathosystem: Relationships between Abscisic acid and cytokinin levels and strain virulence in infected cob tissue. PLoS ONE 2015, 10, e0130945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reineke, G.; Heinze, B.; Schirawski, J.; Buettner, H.; Kahmann, R.; Basse, C.W. Indole-3-acetic acid (IAA) biosynthesis in the smut fungus Ustilago maydis and its relevance for increased IAA levels in infected tissue and host tumor formation. Mol. Plant Pathol. 2008, 9, 339–355. [Google Scholar] [CrossRef]
- Snetselaar, K.M.; Mims, C.W. Light and electron-microscopy of Ustilago maydis hyphae in maize. Mycol. Res. 1994, 98, 347–355. [Google Scholar] [CrossRef]
- Chávez-Arias, C.C.; Gómez-Caro, S.; Restrepo-Díaz, H. Physiological, Biochemical and Chlorophyll Fluorescence Parameters of Physalis Peruviana L. Seedlings Exposed to Different Short-Term, Waterlogging Periods and Fusarium Wilt Infection. Agronomy 2019, 9, 213. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, M.C.; Vega, J.; Oliveira, J.G.; Gomes, M.M.A. Sugarcane yellow leaf virus infection leads to alterations in photosynthetic efficiency and carbohydrate accumulation in sugarcane leaves. Fitopatol. Bras. 2005, 30, 10–16. [Google Scholar] [CrossRef]
- Kyrychenko, A.M. Influence of bean yellow mosaic virus on metabolism of photosynthetic pigments, proteins and carbohydrates in Glycine soja L. Mikrobiol. Zh. 2014, 76, 47–52. [Google Scholar]
- Cortleven, A.; Schmülling, T. Regulation of chloroplast development and function by cytokinin. J. Exp. Bot. 2015, 66, 4999–5013. [Google Scholar] [CrossRef] [Green Version]
- Dobránszki, J.; Mendler-Drienyovszki, N. Cytokinin-induced changes in the chlorophyll content and fluorescence of in vitro apple leaves. J. Plant Phyiol. 2014, 171, 1472–1478. [Google Scholar] [CrossRef]
- Czerpak, R.; Dobrzyn, P.; Krotke, A.; Kicińska, E. The effect of auxins and salicylic acid on chlorophyll and carotenoid contents in Wolffia Arrhiza (L.) Wimm. (Lemnaceae) growing on media of various trophiticities. Pol. J. Environ. Stud. 2002, 11, 231–235. [Google Scholar]
- Misra, A.N.; Biswal, U.C. Effect of phytohormones on chlorophyll degradation during aging of chloroplast in vivo and in vitro. Protoplasma 1980, 105, 1–8. [Google Scholar] [CrossRef]
- Even-Chen, Z.; Atsmon, D.; Itai, C. Hormonal aspects of senescence in detached Tobacco leaves. Physiol. Plant. 1978, 44, 377–382. [Google Scholar] [CrossRef]
- Aharoni, N. Interrelationship between ethylene and growth regulators in the senescence of lettuce leaf discs. J. Plant Grow. Regul. 1989, 8, 309–317. [Google Scholar] [CrossRef]
- Ceusters, J.; van de Poel, B. Ethylene exerts species-species and age-dependent control of photosynthesis. Plant Physiol. 2018, 176, 2601–2612. [Google Scholar] [CrossRef] [Green Version]
- Giordano, M.; Petropoulos, S.A.; Cirillo, C.; Rouphael, Y. Biochemical, Physiological, and Molecular Aspects of Ornamental Plants Adaptation to Deficit Irrigation. Horticulturae 2021, 7, 107. [Google Scholar] [CrossRef]
- Mylonas, C.; Kouretas, D. Lipid peroxidation and tissue damage. In Vivo 1999, 13, 295–309. [Google Scholar]
- Monazzah, M.; Rabiei, Z.; Enferadi, S.T. The Effect of Oxalic Acid, the Pathogenicity Factor of Sclerotinia sclerotiorum on the Two Susceptible and Moderately Resistant Lines of Sunflower. Iran. J. Biotechnol. 2018, 16, 20–30. [Google Scholar] [CrossRef] [Green Version]
- Eloy, Y.R.; Vasconcelos, I.M.; Barreto, A.L.; Freire-Filho, F.R.; Oliveira, J.T. H2O2 plays an important role in the lifestyle of Colletotrichum gloeosporioides during interaction with cowpea (Vigna unguiculata (L.) Walp). Fungal Biol. 2015, 119, 747–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, H.; Ahmed, F.; Lin, B.; Li, Z.; Huang, Y.; Sun, G.; Ding, H.; Wang, C.; Meng, C.; Gao, Z. The effects of plant growth regulators on cell growth, protein, carotenoid, PUFAs and lipid production of Chlorella pyrenoidosa ZF Strain. Energies 2017, 10, 1696. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Xu, J. Phytohormones in microalgae: A new opportunity for microalgal biotechnology? Trends Plant Sci. 2015, 20, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Sivaramakrishnan, R.; Incharoernsakdi, A. Plant hormone induced enrichment of Chlorella sp omega-3 fatty acids. Biotech. Biofuels 2020, 13, 7. [Google Scholar] [CrossRef] [Green Version]
- Nazir, Y.; Halim, H.; Prabhakaran, P.; Ren, X.; Naz, T.; Mohamed, H.; Nosheen, S.; Mustafa, K.; Yang, W.; Abdul Hamid, A.; et al. Different Classes of Phytohormones Act Synergistically to Enhance the Growth, Lipid and DHA Biosynthetic Capacity of Aurantiochytrium sp. SW1. Biomolecules 2020, 10, 755. [Google Scholar] [CrossRef]
- Yu, X.J.; Sun, J.; Sun, Y.Q.; Zheng, J.Y.; Wang, Z. Metabolomics analysis of phytohormone gibberellin improving lipid and DHA accumulation in Aurantiochytrium sp. Biochem. Eng. J. 2016, 112, 258–268. [Google Scholar] [CrossRef]
- Caverzan, A.; Casassola, A.; Brammer, S.P. Antioxidant responses of wheat plants under stress. Gen. Mol. Biol. 2016, 39, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Vanacker, H.; Carver, T.L.; Foyer, C.H. Pathogen-induced changes in the antioxidant status of the apoplast in barley leaves. Plant Physiol. 1998, 117, 1103–1114. [Google Scholar] [CrossRef] [Green Version]
- Raseetha, S.; Oey, I.; Burritt, D.; Heenan, S.; Hamid, N. Evolution of antioxidant enzymes activity and volatile release during storage of processed broccoli (Brassica oleracea L. italica). LWT Food Sci. Technol. 2013, 54, 216–223. [Google Scholar] [CrossRef]
- Mahmud, J.A.; Bhuyan, M.H.M.B.; Anee, T.I.; Nahar, K.; Fujita, M.; Hasanuzzaman, M. Reactive oxygen species metabolism and antioxidant defense in plants under metal/metalloid stress. In Plant Abiotic Stress Tolerance; Hasanuzzaman, M., Hakeem, K., Nahar, K., Alharby, H., Eds.; Springer: Cham, Switzerland, 2019; pp. 221–257. [Google Scholar]
- Zehra, A.; Meena, M.; Dubey, M.K.; Aamir, M.; Upadhyay, R.S. Synergistic effects of plant defense elicitors and Trichoderma harzianum on enhanced induction of antioxidant defense system in tomato against Fusarium wilt disease. Bot. Stud. 2017, 58, 44. [Google Scholar] [CrossRef]
- Fimognari, L.; Dölker, R.; Kaselyte, G.; Jensen, C.; Akhtar, S.S.; Großkinsky, D.K.; Roitsch, T. Simple semi-high throughput determination of activity signatures of key antioxidant enzymes for physiological phenotyping. Plant Methods 2020, 16, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzniak, E.; Skłodowska, M. Fungal pathogen-induced changes in the antioxidant systems of leaf peroxisomes from infected tomato plants. Planta 2005, 222, 192–200. [Google Scholar] [CrossRef]
- Paranidharan, V.; Palaniswami, A.; Vidhyasekaran, P.; Velazhahan, R. A host-specific toxin of Rhizoctonia solani triggers superoxide dismutase (SOD) activity in rice. Arch. Phytopathol. Pflanzenschutz. 2005, 38, 151–157. [Google Scholar] [CrossRef]
- Lee, A.; Trinh, C.S.; Lee, W.J.; Kim, M.; Lee, H.; Pathiraja, D.; Choi, I.G.; Chung, N.; Choi, C.; Lee, B.C.; et al. Characterization of two leaf rust-resistant Aegilops tauschii accessions for the synthetic wheat development. Appl. Biol. Chem. 2020, 63, 13. [Google Scholar] [CrossRef] [Green Version]
- Naz, R.; Bano, A.; Nosheen, A.; Yasmin, H.; Keyani, R.; Shah, S.M.; Anwar, Z.; Roberts, T.H. Induction of defense-related enzymes and enhanced disease resistance in maize against Fusarium verticillioides by seed treatment with Jacaranda mimosifolia formulations. Sci. Rep. 2021, 11, 59. [Google Scholar] [CrossRef] [PubMed]
- Gulati, S.; Ballhausen, M.B.; Kulkarni, P.; Grosch, R.; Garbeva, P. A non-invasive soil-based setup to study tomato root volatiles released by healthy and infected roots. Sci. Rep. 2020, 10, 12704. [Google Scholar] [CrossRef]
- Aydi-Ben-Abdallah, R.; Jabnoun-Khiareddine, H.; Daami-Remadi, M. Fusarium wilt biocontrol and tomato growth stimulation, using endophytic bacteria naturally associated with Solanum sodomaeum and S. bonariense plants. Egypt. J. Biol. Pest Co. 2020, 30, 113. [Google Scholar] [CrossRef]
- Robert, C.; Bancal, M.O.; Nicolas, P.; Lannou, C.; Ney, B. Analysis and modelling of effects of leaf rust and Septoria tritici blotch on wheat growth. J. Exp. Bot. 2004, 55, 1079–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahir, H.A.S.; Gu, Q.; Wu, H.; Raza, W.; Safdar, A.; Huang, Z.; Rajer, F.U.; Gao, X. Effect of volatile compounds produced by Ralstonia solanacearum on plant growth promoting and systemic resistance inducing potential of Bacillus volatiles. BMC Plant Biol. 2017, 17, 133. [Google Scholar] [CrossRef]
- Prasch, C.M.; Sonnewald, U. Simultaneous application of heat, drought, and virus to Arabidopsis plants reveals significant shifts in signaling networks. Plant Physiol. 2013, 162, 1849–1966. [Google Scholar] [CrossRef] [PubMed]
- Szőke, L.; Tóth, B. Examination of the effect of corn smut infection on plant physiological parameters in a small scale field experiment. In Proceedings of the 44th Conference for Students of Agriculture and Veterinary Medicine with International Participation, Online, University of Novi Sad, Faculty of Agriculture, Novi Sad, Serbia, 15 December 2020; pp. 37–43. [Google Scholar]
- Zhu, S.; Gao, F.; Cao, X.; Chen, M.; Ye, G.; Wei, C.; Li, Y. The rice dwarf virus P2 protein interacts with ent-kaurene oxidases in vivo, leading to reduced biosynthesis of gibberellins and rice dwarf symptoms. Plant Physiol. 2005, 139, 1935–1945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, N.; Matsuoka, M.; Kitano, H.; Asano, T.; Kaku, H.; Komatsu, S. gid1, a gibberellin-insensitive dwarf mutant, shows altered regulation of probenazole-inducible protein (PBZ1) in response to cold stress and pathogen attack. Plant Cell Environ. 2006, 29, 619–631. [Google Scholar] [CrossRef]
- Argueso, C.T.; Ferreira, F.J.; Epple, P.; To, J.P.; Hutchison, C.E.; Schaller, G.E.; Dang, J.L.; Kieber, J.J. Two-component elements mediate interactions between cytokinin and salicylic acid in plant immunity. PLoS Genet. 2012, 8, e1002448. [Google Scholar] [CrossRef] [Green Version]
- Sardesai, N.; Lee, L.Y.; Chen, H.; Yi, H.; Olbricht, G.R.; Stirnberg, A.; Jeffries, J.; Xiong, K.; Doerge, R.W.; Gelvin, S.B. Cytokinins Secreted by Agrobacterium Promote Transformation by Repressing a Plant Myb Transcription Factor. Sci. Signal. 2013, 6, ra100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, N.; Goodwin, P.H.; Hsiang, T. The role of ethylene during the infection of Nicotiana tabacum by Colletotrichum destructivum. J. Exp. Bot. 2003, 54, 2449–2456. [Google Scholar] [CrossRef]
- Pennell, R.I.; Lamb, C. Programmed cell death in plants. Plant Cell 1997, 9, 1157–1168. [Google Scholar] [CrossRef] [Green Version]
- Veselova, S.V.; Nuzhnaya, T.V.; Burkhanova, G.F.; Rumyantsev, S.D.; Khusnutdinova, E.K.; Maksimov, I.V. Ethylene-Cytokinin Interaction Determines Early Defense Response of Wheat against Stagonospora nodorum Berk. Biomolecules 2021, 11, 174. [Google Scholar] [CrossRef]
- Dong, T.; Zheng, T.; Fu, W.; Guan, L.; Jia, H.; Fang, J. The Effect of Ethylene on the Color Change and Resistance to Botrytiscinerea Infection in ‘Kyoho’ GrapeFruits. Foods 2020, 9, 892. [Google Scholar] [CrossRef]
- Couto, D.; Zipfel, C. Regulation of pattern recognition receptor signaling in plants. Nat. Rev. Immunol. 2016, 16, 537–552. [Google Scholar] [CrossRef] [PubMed]
- Macho, A.P.; Zipfel, C. Plant PRRs and the activation of innate immune signaling. Mol. Cell. 2014, 54, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, K.; Katagiri, F. Comparing signaling mechanisms engaged in pattern-triggered and effector-triggered immunity. Curr. Opin. Plant Biol. 2010, 13, 459–465. [Google Scholar] [CrossRef]
- Berens, M.L.; Berry, H.M.; Mine, A.; Argueso, C.T.; Tsuda, K. Evolution of hormone signaling networks in plant defense. Annu. Rev. Phytopathol. 2017, 55, 401–425. [Google Scholar] [CrossRef]
- Chanclud, E.; Morel, J.B. Plant hormones: A fungal point of view. Mol. Plant Pathol. 2016, 17, 1289–1297. [Google Scholar] [CrossRef]
- Turian, G.; Hamilton, R.H. Chemical detection of 3-indolyl acetic acid in Ustilago zeae tumors. Biochim. Biophys. Acta 1960, 41, 148–150. [Google Scholar] [CrossRef]
- Sorensen, J.L.; Benfield, A.H.; Wollenberg, R.D.; Westphal, K.; Wimmer, R.; Nielsen, M.R.; Nielsen, K.F.; Carere, J.; Covarelli, L.; Beccari, G.; et al. The cereal pathogen Fusarium pseudograminearum produces a new class of active cytokinins during infection. Mol. Plant Pathol. 2018, 19, 1140–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruce, S.A.; Saville, B.J.; Emery, R.J.N. Ustilago maydis produces cytokinins and abscisic acid for potential regulation of tumor formation in maize. J. Plant Growth 2011, 30, 51–63. [Google Scholar] [CrossRef]
- Yang, D.L.; Li, Q.; Deng, Y.W.; Lou, Y.G.; Wang, M.Y.; Zhou, G.X.; Zhang, Y.Y.; He, Z.H. Altered disease development in the eui mutants and Eui overexpressors indicates that gibberellins negatively regulate rice basal disease resistance. Mol. Plant 2008, 1, 528–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, X.; Liu, J.H.; Zhao, W.S.; Chen, X.J.; Guo, Z.J.; Peng, Y.L. Gibberellin 20-oxidase gene OsGA20ox3 regulates plant stature and disease development in rice. Mol. Plant-Microbe Interact. 2013, 26, 227–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vleesschauwer, D.; Yang, Y.; Cruz, C.V.; Hofte, M. Abscisic acid-induced resistance against the brown spot pathogen Cochliobolus miyabeanus in rice involves MAP kinase-mediated repression of ethylene signaling. Plant Physiol. 2010, 152, 2036–2052. [Google Scholar] [CrossRef] [Green Version]
- Van Bockhaven, J.; Spíchal, L.; Novák, O.; Strnad, M.; Asano, T.; Kikuchi, S.; Höfte, M.; de Vleesschauwer, D. Silicon induces resistance to the brown spot fungus Cochliobolus miyabeanus by preventing the pathogen from hijacking the rice ethylene pathway. New Phytol. 2015, 206, 761–773. [Google Scholar] [CrossRef]
- Han, X.; Kahmann, R. Manipulation of phytohormone pathways by effectors of filamentous plant pathogens. Fron. Plant Sci. 2019, 10, 822. [Google Scholar] [CrossRef] [Green Version]
- Moran, R.; Porath, D. Chlorophyll Determination in intact tissues using N, N-dimethylformamide. Plant Physiol. 1980, 65, 478–479. [Google Scholar] [CrossRef] [Green Version]
- Wellburn, R.A. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvent with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Pukacka, S.; Ratajczak, E. Production and scavenging of reactive oxygen species in Fagus sylvatica seeds during storage at varied temperature and humidity. J. Plant Physiol. 2005, 162, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.P.; Mishra, R.K.; Singhal, G.S. Changes in the activities of antioxidant enzymes during exposure of intact wheat leaves to strong visible light at different temperatures in the presence of protein synthesis inhibitors. Plant Physiol. 1993, 102, 903–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeislin, N.; Ben-Zaken, R. Peroxidases, phenylalanine ammonia-lyase and lignification in peduncles of rose flowers. Plant Physiol. Biochem. 1991, 29, 147–151. [Google Scholar]
- Giannopolities, C.H.; Ries, S.K. Superoxide dismutase I. Occurrence in higher plant. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Beyer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ghasemi, A.; Zahediasl, S. Normality tests for statistical analysis: A guide for non-statisticians. Int. J. Endocrinol. Metab. 2012, 10, 486–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahapoonyanont, N.; Mahapoonyanont, T.; Pengkaew, N.; Kamhangkit, R. Power of the test of One-Way Anova after transforming with large sample size data. Procedia Soc. Behav.Sci. 2010, 9, 933–937. [Google Scholar] [CrossRef] [Green Version]
- Nanda, A.; Mohapatra, B.B.; Mahapatra, A.P.K.; Abiresh Prasad Kumar Mahapatra, A.P.K.; Mahapatra, A.P.K. Multiple comparison test by Tukey’s honestly significant difference (HSD): Do the confident level control type I error. IJAMS 2021, 6, 59–65. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Mean ± SD | Infected Mean ± SD | Cytokinin Mean ± SD | Gibberellin Mean ± SD | Auxin Mean ± SD | Ethylene Mean ± SD | |
|---|---|---|---|---|---|---|
| Chlorophyll-a | 15.75 ± 0.55 c | 12.58 ± 1.92 b | 15.735 ± 2.07 c | 12.23 ± 3.49 b | 12.175 ± 2.32 b | 9.44 ± 3.25 a |
| Chlorophyll-b | 5.95 ± 1.29 c | 4.49 ± 1.45 b | 5.79 ± 0.95 c | 5.12 ± 2.64 bc | 4.45 ± 2.64 b | 3.27 ± 1.72 a |
| Carotenoid | 3.62 ± 3.29 bc | 2.97 ± 2.15 ab | 3.37 ± 3.42 b | 3.88 ± 1.74 c | 3.39 ± 1.51 b | 2.49 ± 1.09 a |
| MDA | 19.1 ± 3.97 a | 72.35 ± 27.91 c | 35.68 ± 5.46 b | 38.64 ± 12.89 b | 34.31 ± 17.22 b | 110.23 ± 29.82 d |
| POD | 0.125 ± 0.29 a | 0.314 ± 0.12 c | 0.136 ± 0.45 a | 0.197 ± 0.65 b | 0.212 ± 0.48 b | 0.365 ± 0.021 c |
| APX | 0.202 ± 0.05 a | 0.515 ± 0.11 d | 0.227 ± 0.05 b | 0.313 ± 0.08 c | 0.317 ± 0.11 c | 0.592 ± 0.24 e |
| SOD | 0.072 ± 0.001 a | 0.105 ± 0.003 c | 0.081 ± 0.003 b | 0.073 ± 0.004 a | 0.082 ± 0.002 b | 0.108 ± 0.004 c |
| Plant height | 627 ± 217.74 d | 592 ± 246.20 c | 559 ± 161.55 b | 743 ± 254.19 e | 643 ± 258.47 c | 438 ± 171.67 a |
| Stem Diameter | 3.45 ± 0.23 bc | 2.56 ± 0.58 b | 3.40 ± 0.04 bc | 2.82 ± 1.50 bc | 2.45 ± 1.33 b | 1.79 ± 0.99 a |
| Shoot dry weight | 3.51 ± 0.76 c | 2.96 ± 0.61 bc | 2.63 ± 0.53 b | 4.33 ± 0.41 d | 3.18 ± 0.31 bc | 2.06 ± 0.32 a |
| Armagnac Mean ± SD | Desszert 73 Mean ± SD | LSD | |
|---|---|---|---|
| Chlorophyll-a | 12.88 ± 2.99 a | 13.09 ± 3.39 a | 0.378 |
| Chlorophyll-b | 4.43 ± 1.83 b | 1.88 ± 1.88 a | 0.038 |
| Carotenoids | 6.29 ± 1.56 a | 5.98 ± 1.70 a | 0.121 |
| MDA | 28.88 ± 9.07 a | 74.56 ± 24.45 b | 0.025 |
| POD | 0.30 ± 0.11 a | 0.37 ± 0.19 a | 0.216 |
| APX | 0.29 ± 0.13 a | 0.43 ± 0.18 b | 0.014 |
| SOD | 0.0815 ± 0.0006 a | 0.08901 ± 0.0018 a | 0.189 |
| Plant height | 751.00 ± 80.43 b | 449.67 ± 96.61 a | 0.030 |
| Stem diameter | 6.57 ± 0.99 a | 7.20 ± 1.16 b | 0.036 |
| Shoot dry weight | 2.98 ± 0.95 a | 3.25 ± 0.85 a | 0.176 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szőke, L.; Moloi, M.J.; Kovács, G.E.; Biró, G.; Radócz, L.; Hájos, M.T.; Kovács, B.; Rácz, D.; Danter, M.; Tóth, B. The Application of Phytohormones as Biostimulants in Corn Smut Infected Hungarian Sweet and Fodder Corn Hybrids. Plants 2021, 10, 1822. https://doi.org/10.3390/plants10091822
Szőke L, Moloi MJ, Kovács GE, Biró G, Radócz L, Hájos MT, Kovács B, Rácz D, Danter M, Tóth B. The Application of Phytohormones as Biostimulants in Corn Smut Infected Hungarian Sweet and Fodder Corn Hybrids. Plants. 2021; 10(9):1822. https://doi.org/10.3390/plants10091822
Chicago/Turabian StyleSzőke, Lóránt, Makoena Joyce Moloi, Gabriella Enikő Kovács, Györgyi Biró, László Radócz, Mária Takácsné Hájos, Béla Kovács, Dalma Rácz, Mátyás Danter, and Brigitta Tóth. 2021. "The Application of Phytohormones as Biostimulants in Corn Smut Infected Hungarian Sweet and Fodder Corn Hybrids" Plants 10, no. 9: 1822. https://doi.org/10.3390/plants10091822
APA StyleSzőke, L., Moloi, M. J., Kovács, G. E., Biró, G., Radócz, L., Hájos, M. T., Kovács, B., Rácz, D., Danter, M., & Tóth, B. (2021). The Application of Phytohormones as Biostimulants in Corn Smut Infected Hungarian Sweet and Fodder Corn Hybrids. Plants, 10(9), 1822. https://doi.org/10.3390/plants10091822