
Tree Growth Conditions Are Demanded When Optimal, Are Unwanted When Limited, but When Are They Suboptimal?




{kind=link}
Abstract
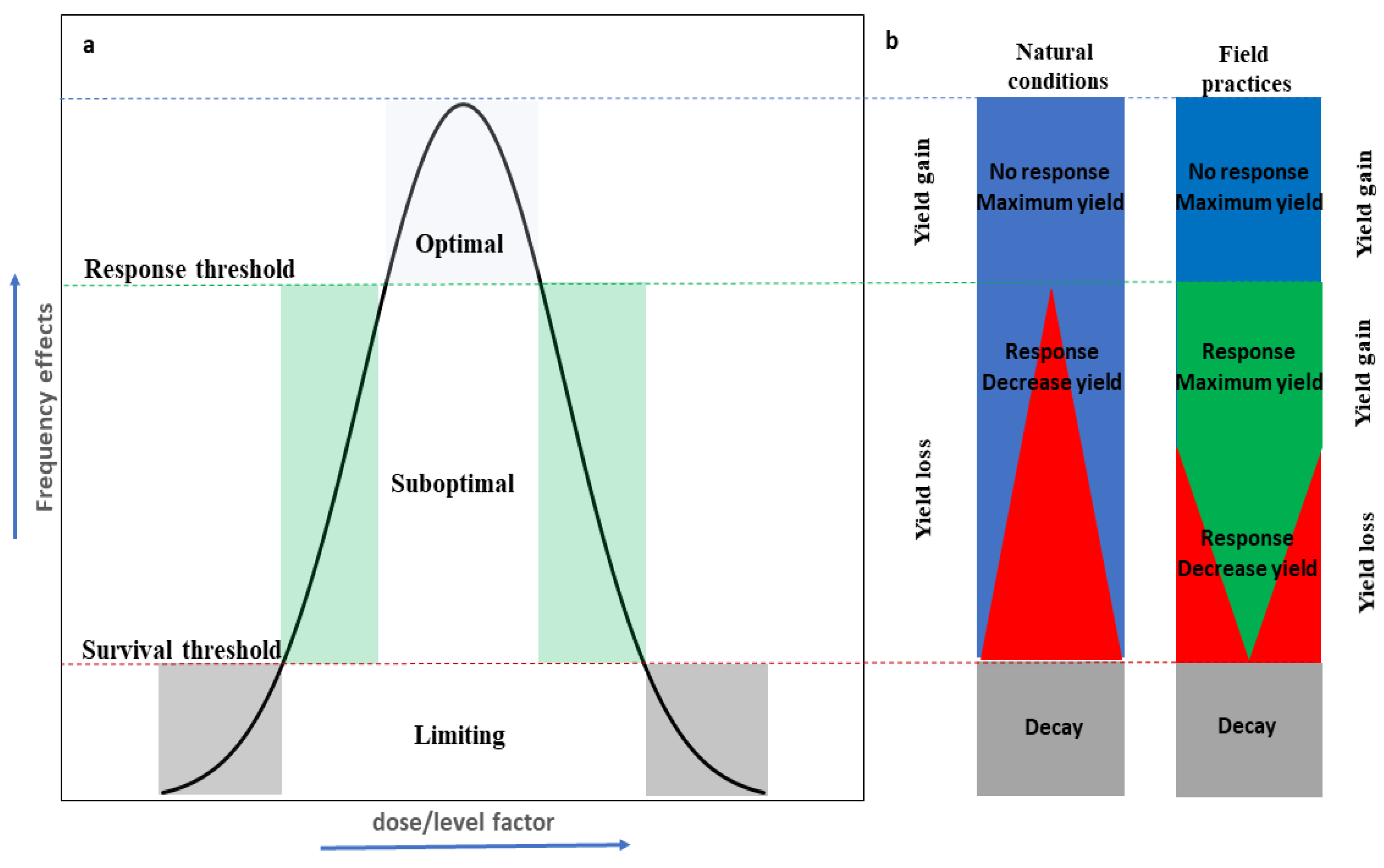
:1. Toward a Wider Definition of ‘Suboptimal Growth Conditions’
2. How to Study the Effects of Suboptimal Conditions in Trees under Natural Conditions
3. How Does Global Change Affect Suboptimal Conditions at Large Scale?
4. Might Suboptimal Conditions Drive Tree Acclimation and Adaptation?
5. General Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.D.; Schroeder, J.I. Genetic strategies for improving crop yields. Nature 2019, 575, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Inzé, D. Plant growth under suboptimal water conditions: Early responses and methods to study them. J. Exp. Bot. 2020, 7, 1706–1722. [Google Scholar]
- Poorter, H.; Bühler, J.; van Dusschoten, D.; Climent, J.; Postma, J.A. Pot size matters: A meta-analysis of the effects of rooting volume on plant growth. Funct. Plant Biol. 2012, 39, 839–850. [Google Scholar] [CrossRef] [Green Version]
- Brodribb, T.J.; Powers, J.; Cochard, H.; Choat, B. Hanging by a thread? Forests and drought. Science 2020, 368, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Mouheb, H.A.; Kadik, L.; Albert, C.H.; Berrached, R.; Prinzing, A. How do steppe plants follow their optimal environmental conditions or persist under suboptimal conditions? The differing strategies of annuals and perennials. Ecol. Evol. 2018, 8, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Keddy, P. Plant Ecology Origins, Processes, Consequences; Cambridge University Press: New York, NY, USA, 2017. [Google Scholar] [CrossRef]
- Di Matteo, G.; Nardi, P.; Verani, S.; Sperandio, G. Physiological adaptability of poplar clones selected for bioenergy purposes under non-irrigated and suboptimal site conditions: A case study in Central Italy. Biomass Bioenerg. 2015, 81, 183–189. [Google Scholar] [CrossRef]
- Alexander, P.D.; Williams, R.H.; Nevison, I.M. Improving gardener’s understanding of water management in peat and peat-free multi-purpose growing media: An assessment with fucsia. Acta Hortic. 2013, 1013, 257–263. [Google Scholar] [CrossRef]
- Scheiber, S.M.; Beeson, R. Petunia growth and maintenance in the landscape as influenced by alternative irrigation strategies. Hort. Sci. 2006, 41, 235–238. [Google Scholar] [CrossRef] [Green Version]
- Herrera, F.; Castillo, J.E.; Chica, A.F.; López Bellido, L. Use of municipal solid waste compost (MSWC) as a growing medium in the nursery production of tomato plants. Bioresour. Technol. 2008, 99, 287–296. [Google Scholar] [CrossRef]
- Hernández-Apaolaza, L.; Gascó, A.M.; Gascó, J.M.; Guerrero, F. Reuse of waste materials as growing media for ornamental plants. Bioresour. Technol. 2015, 96, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Korner, C.; Basler, D.; Hoch, G.; Kollas, C.; Lenz, A.; Randin, C.F.; Vitasse, F.; Zimmermann, N.E. Where, why and how? Explaining the low-temperaturerange limits of temperate tree species. J. Ecol. 2016, 104, 1076–1088. [Google Scholar] [CrossRef]
- Disney, M. Terrestrial LiDAR: A three-dimensional revolution in how we look at trees. New Phytol. 2019, 222, 1736–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pause, M.; Schweitzer, C.; Rosenthal, M.; Keuck, V.; Bumberger, J.; Dietrich, P.; Heurich, M.; Jung, A.; Lausch, A. In Situ/Remote Sensing Integration to Assess Forest Health—A Review. Remote Sens. 2016, 8, 471. [Google Scholar] [CrossRef] [Green Version]
- Amaral, J.; Ribeyre, Z.; Vigneaud, J.; Sow, M.D.; Fichot, R.; Messier, C.; Pinto, G.; Nolet, P.; Maury, S. Advances and Promises of Epigenetics for Forest Trees. Forests 2020, 11, 976. [Google Scholar] [CrossRef]
- Bewg, W.P.; Ci, D.; Tsai, C.-J. Genome Editing in Trees: From Multiple Repair Pathways to Long-Term Stability. Front. Plant Sci. 2018, 9, 9. [Google Scholar] [CrossRef]
- IPCC. Global warming of 1.5 °C. In An IPCC Special Report on the Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Masson-Delmotte, V., Zhai, P., Pörtner, H.O., Roberts, D., Skea, J., Shukla, P.R., Pirani, A., Moufouma-Okia, W., Péan, C., Pidcock, R., et al., Eds.; IPCC: Geneva, Switzerland, 2018. [Google Scholar]
- Vincent, L.A.; Zhang, X.; Brown, R.D.; Feng, Y.; Mekis, E.; Milewska, E.J.; Wan, H.; Wang, X.L. Observed trends in Canada’s climate and influence of low-frequency variability modes. J. Clim. 2015, 28, 4545–4560. [Google Scholar] [CrossRef]
- Wang, C.; Wang, B.; Cao, J. Unprecedented Northern Hemisphere tropical cyclone genesis in 2018 shaped by subtropical warming in the North Pacific and the North Atlantic. Geophys. Res. Lett. 2019, 46, 13327–13337. [Google Scholar] [CrossRef]
- Tabari, H.; Madani, K.; Willems, P. The contribution of anthropogenic influence to more anomalous extreme precipitation in Europe. Environ. Res. Lett. 2020, 15, 104077. [Google Scholar] [CrossRef]
- Price, D.; Alfaro, R.; Brown, K.; Flannigan, M.; Fleming, R.; Hogg, E.; Girardin, M.; Lakusta, T.; Johnston, M.; McKenney, D.; et al. Anticipating the consequences of climate change for Canada’s boreal forest ecosystems. Environ. Rev. 2013, 21, 322–365. [Google Scholar] [CrossRef]
- Huang, C.; Asner, G.; Barger, N.; Neff, J.; Floyd, M. Regional aboveground live carbon losses due to drought-induced tree dieback in pinon-juniper ecosystems. Remote Sens. Environ. 2010, 114, 1471–1479. [Google Scholar] [CrossRef]
- van Zanten, M.; Ai, H.; Quint, M. Plant thermotropism: An underexplored thermal engagement and avoidance strategy. J. Exp. Bot. 2021, erab209. [Google Scholar] [CrossRef] [PubMed]
- Coops, N.C.; Coggins, S.B.; Kurz, W.A. Mapping the environmental limitations to growth of coastal Douglas-fir stands on Vancouver Island, British Columbia. Tree Physiol. 2007, 27, 805–815. [Google Scholar] [CrossRef] [Green Version]
- Escandón, A.B.; Rojas, R.; Morales, L.V.; Corcuera, L.J.; Coopman, R.E.; Paula, S. Physiological differences between root suckers and saplings enlarge the regeneration niche in Eucryphia cordifolia Cav. Tree Physiol. 2018, 38, 129–138. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cocozza, C.; Traversi, M.L.; Giovannelli, A. Tree Growth Conditions Are Demanded When Optimal, Are Unwanted When Limited, but When Are They Suboptimal? Plants 2021, 10, 1943. https://doi.org/10.3390/plants10091943
Cocozza C, Traversi ML, Giovannelli A. Tree Growth Conditions Are Demanded When Optimal, Are Unwanted When Limited, but When Are They Suboptimal? Plants. 2021; 10(9):1943. https://doi.org/10.3390/plants10091943
Chicago/Turabian StyleCocozza, Claudia, Maria Laura Traversi, and Alessio Giovannelli. 2021. "Tree Growth Conditions Are Demanded When Optimal, Are Unwanted When Limited, but When Are They Suboptimal?" Plants 10, no. 9: 1943. https://doi.org/10.3390/plants10091943
APA StyleCocozza, C., Traversi, M. L., & Giovannelli, A. (2021). Tree Growth Conditions Are Demanded When Optimal, Are Unwanted When Limited, but When Are They Suboptimal? Plants, 10(9), 1943. https://doi.org/10.3390/plants10091943