
Above- and Below-Ground Carbon Storage of Hydrologically Altered Mangrove Wetlands in Puerto Rico after a Hurricane


Abstract
:1. Introduction
2. Results
2.1. Mangrove Forest Community Structure
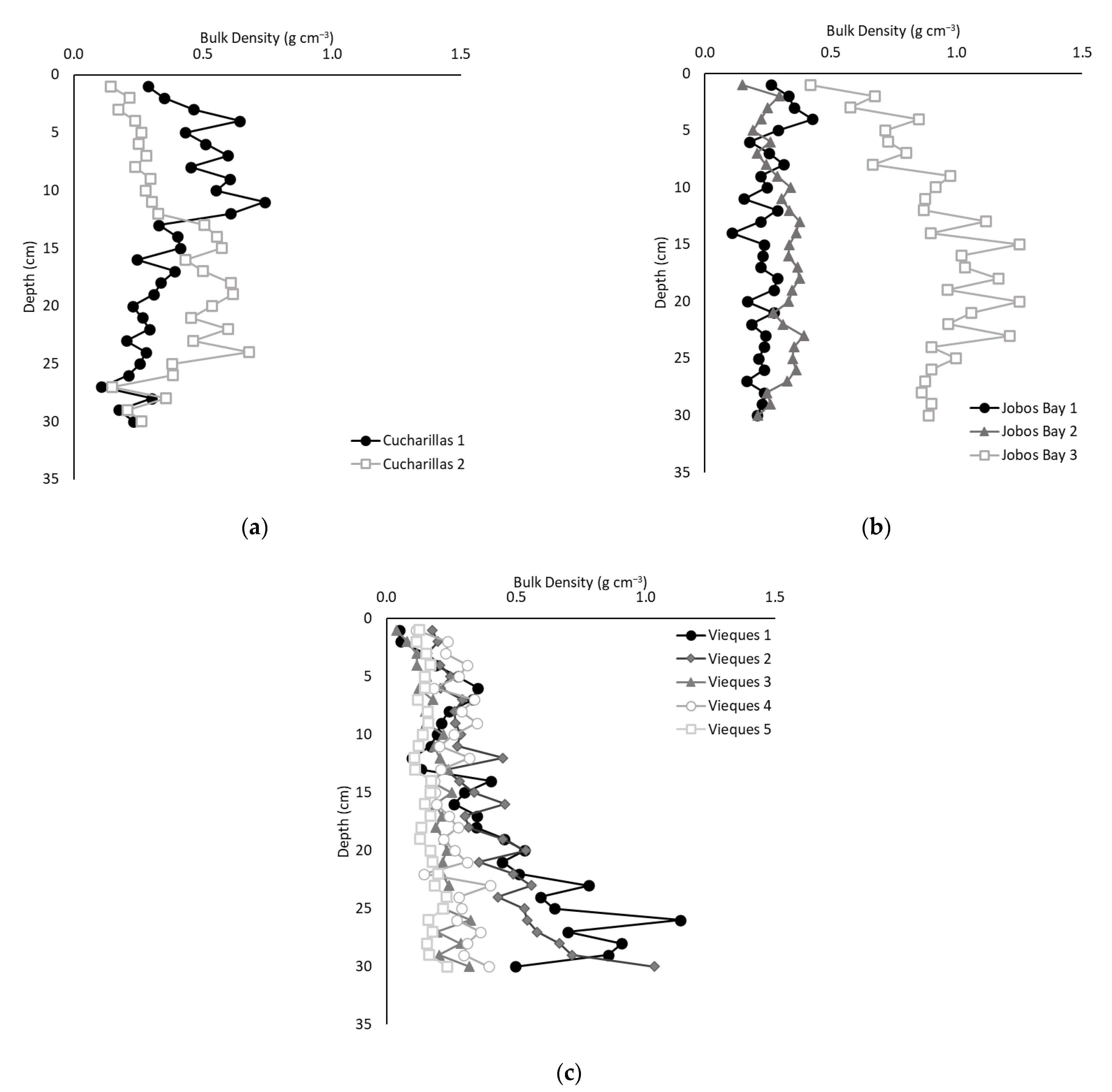
2.2. Soil Bulk Density
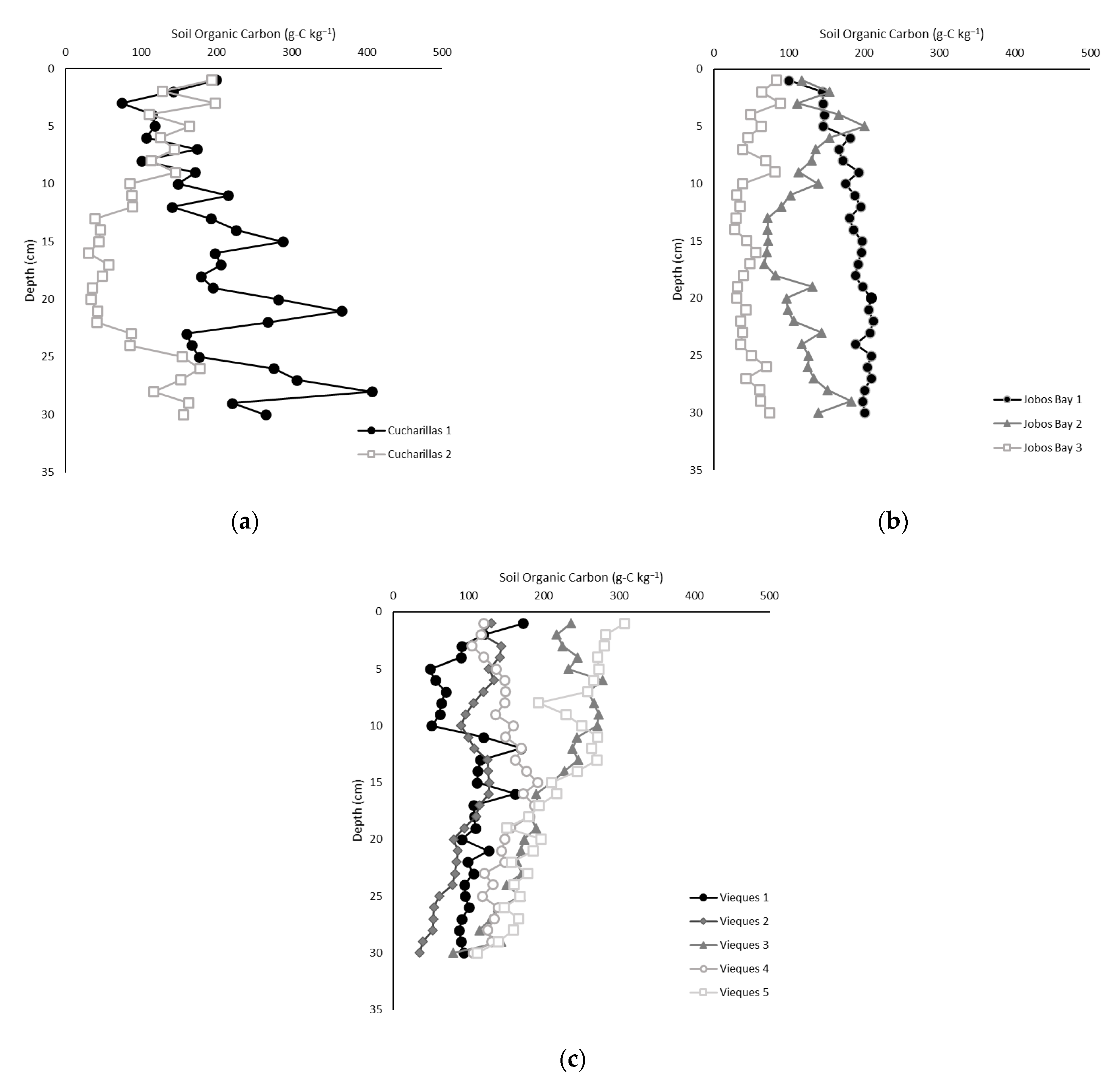
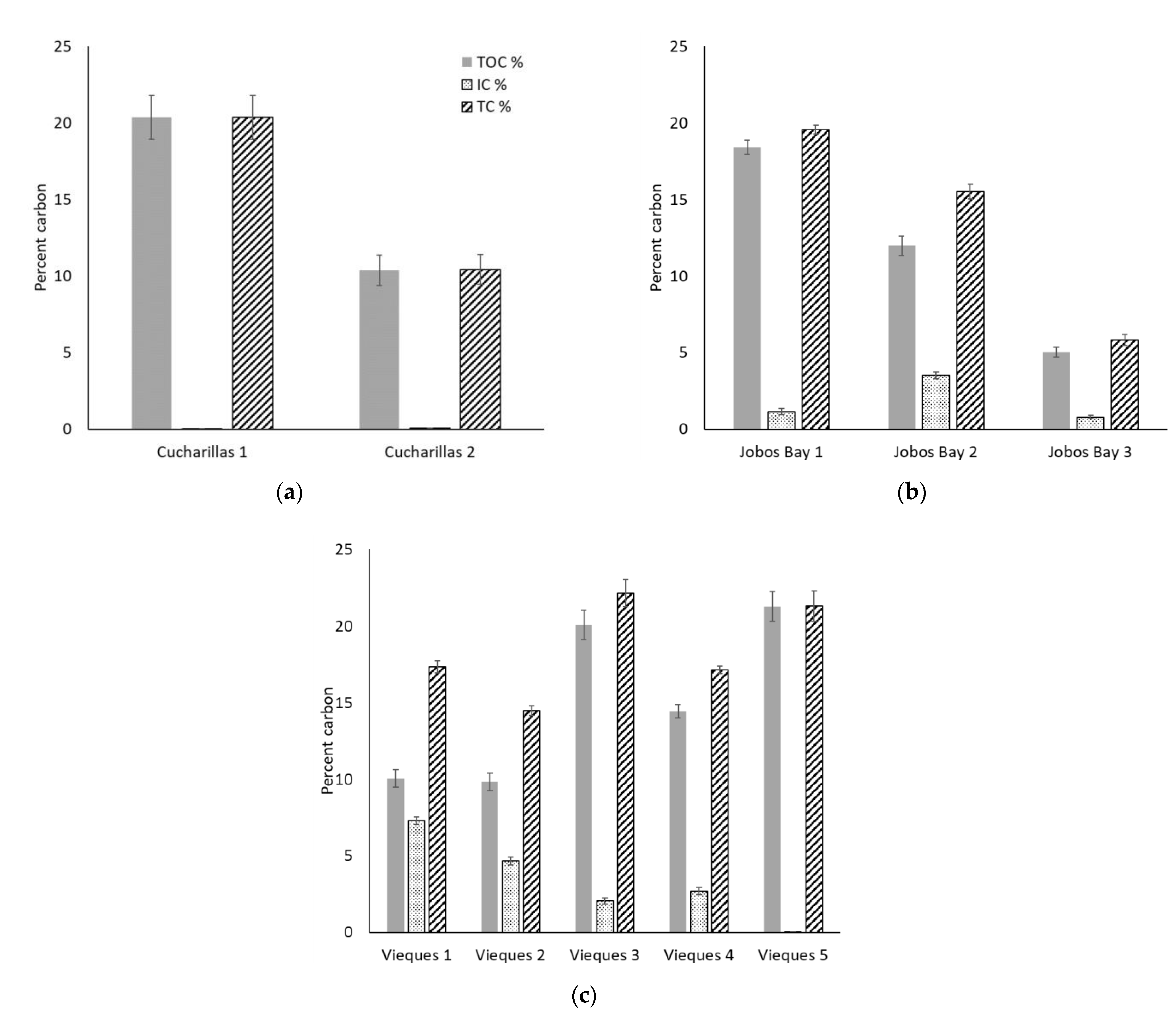
2.3. Soil Carbon Content
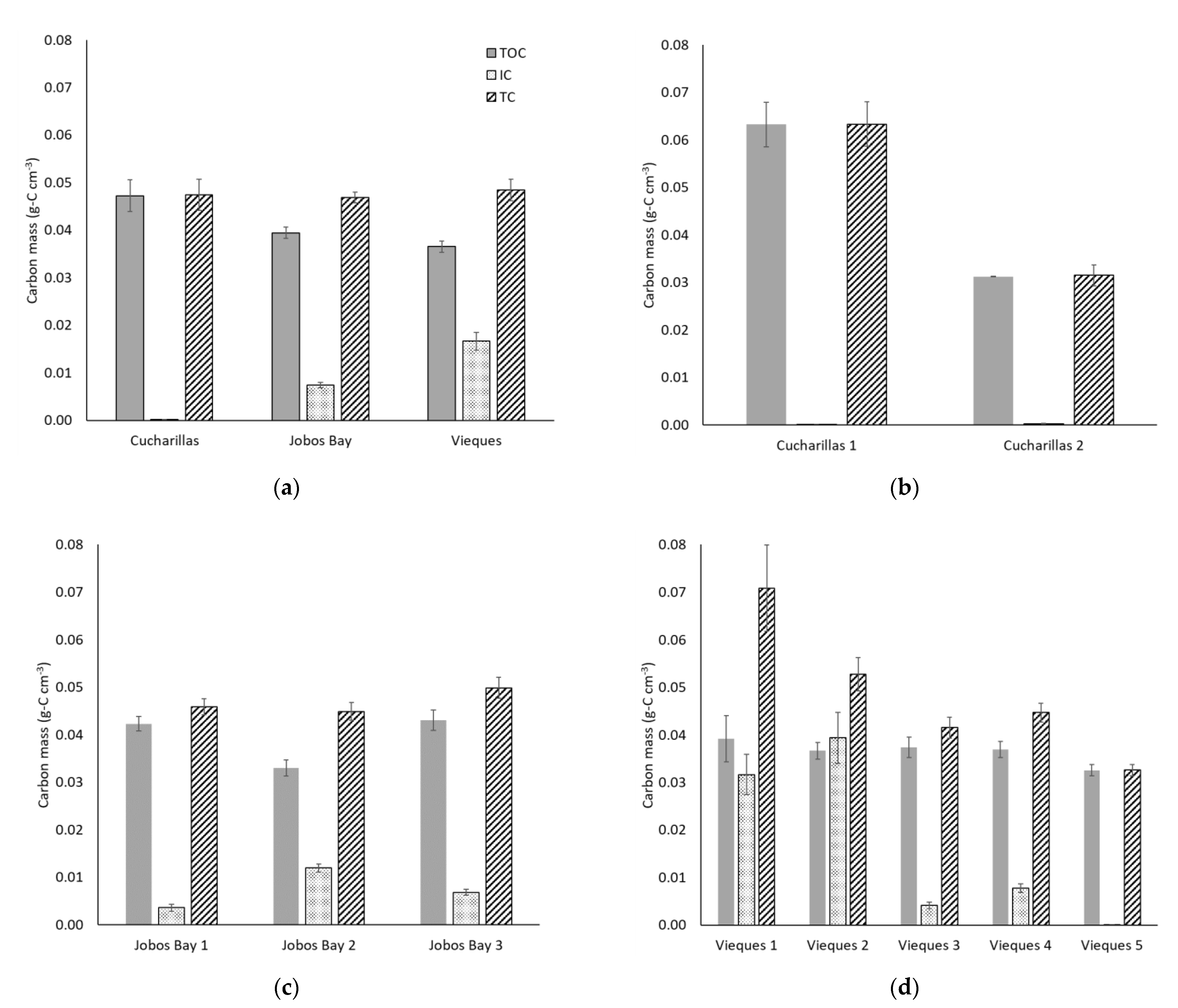
2.4. Carbon Mass Soil Profile
3. Discussion
3.1. Mangrove Forest Community Structure
3.2. Soil Carbon Storage
3.3. Total Carbon Stock
3.4. Limitations
4. Materials and Methods
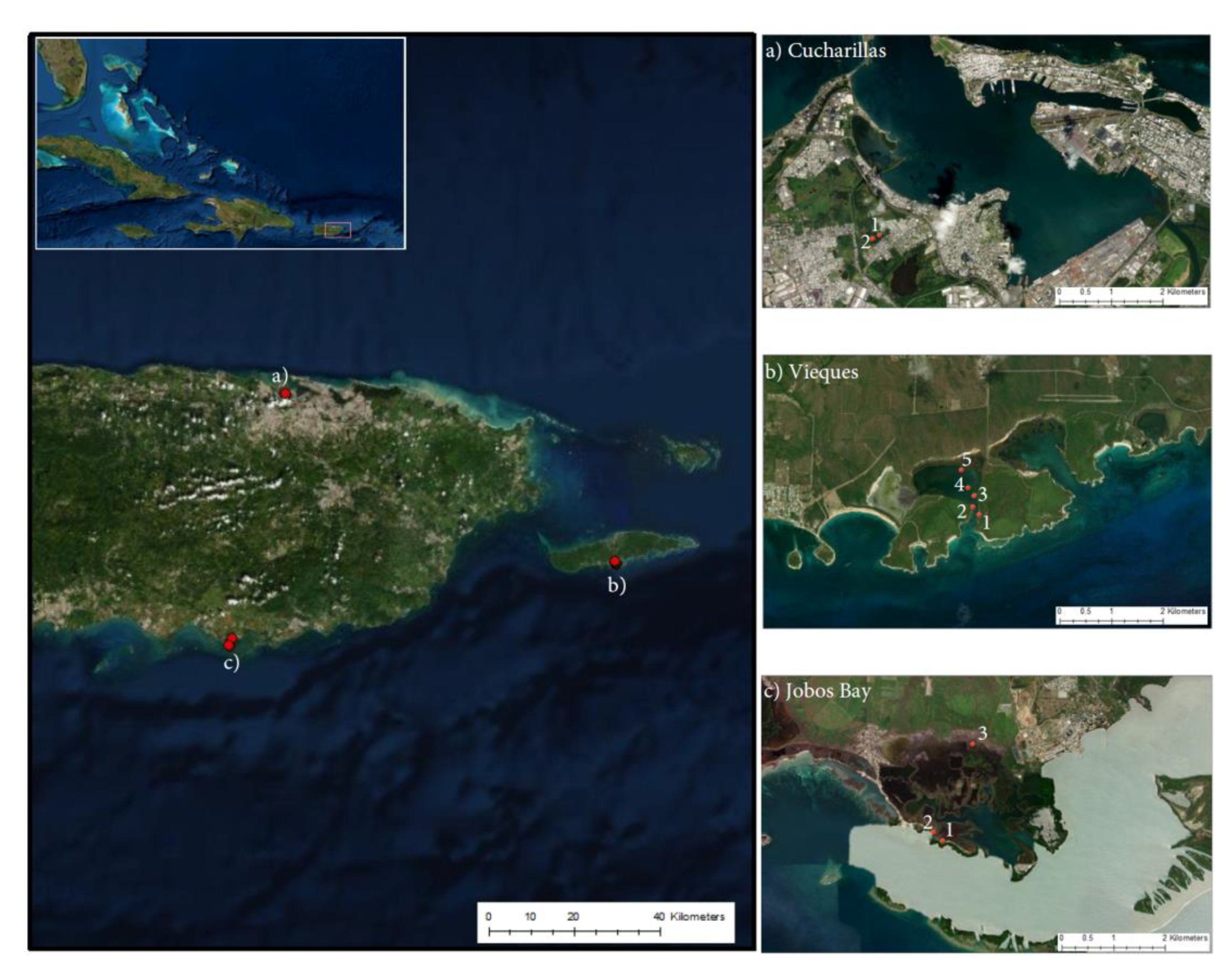
4.1. Study Sites
4.2. Sampling and Data Collection
4.3. Laboratory Procedure
4.4. Statistical Analysis
5. Conclusions
- Mangroves can maintain similar function of total carbon storage despite hydrologic disturbances.
- Inorganic carbon storage appears to be negatively affected by hydrologic disturbance.
- Rhizophora mangle dominated mangrove swamps may be less likely to recover after an intense hurricane than mangrove wetlands supporting other mangrove species.
- Mangrove forests have adapted to hydrologic and hurricane disturbance. By allocating carbon stocks to both above and belowground pools, mangroves are able to store carbon despite potential anthropogenic or natural disturbances that may affect one of these pools.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC Climate Change 2014: Synthesis Report. In Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014; p. 151.
- Hallegraeff, G.M. A Review of Harmful Algal Blooms and Their Apparent Global Increase. Phycologia 1993, 32, 79–99. [Google Scholar] [CrossRef] [Green Version]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human Domination of Earth’s Ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef] [Green Version]
- Webster, P.J.; Holland, G.J.; Curry, J.A.; Chang, H.R. Changes in Tropical Cyclone Number, Duration, and Intensity in a Warming Environment. Science 2005, 309, 1844–1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aumann, H.H.; Behrangi, A.; Wang, Y. Increased Frequency of Extreme Tropical Deep Convection: AIRS Observations and Climate Model Predictions. Geophys. Res. Lett. 2018, 45, 13530–13537. [Google Scholar] [CrossRef] [Green Version]
- Mitsch, W.J.; Bernal, B.; Nahlik, A.M.; Mander, Ü.; Zhang, L.; Anderson, C.J.; Jørgensen, S.E.; Brix, H. Wetlands, Carbon, and Climate Change. Landsc. Ecol. 2013, 28, 583–597. [Google Scholar] [CrossRef]
- The Puerto Rico Climate Change Council (PRCCC). Puerto Rico’s State of the Climate 2010–2013: Assessing Puerto Rico’s Social-Ecological Vulnerabilities in a Changing Climate; NOAA Office of Ocean and Coastal Resource Management: San Juan, Puerto Rico, 2013. [Google Scholar]
- Barbier, E.B. The Protective Service of Mangrove Ecosystems: A Review of Valuation Methods. Mar. Pollut. Bull. 2016, 109, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Barbier, E.B.; Hacker, S.D.; Kennedy, C.; Koch, E.W.; Stier, A.C.; Silliman, B.R. The Value of Estuarine and Coastal Ecosystem Services. Ecol. Monogr. 2011, 81, 169–193. [Google Scholar] [CrossRef]
- Mcleod, E.; Chmura, G.L.; Bouillon, S.; Salm, R.; Björk, M.; Duarte, C.M.; Lovelock, C.E.; Schlesinger, W.H.; Silliman, B.R. A Blueprint for Blue Carbon: Toward an Improved Understanding of the Role of Vegetated Coastal Habitats in Sequestering CO2. Front. Ecol. Environ. 2011, 9, 552–560. [Google Scholar] [CrossRef] [Green Version]
- Mitsch, W.J.; Gosselink, J.G. Wetlands, 5th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015. [Google Scholar]
- Davidson, N.C. How Much Wetland Has the World Lost? Long-Term and Recent Trends in Global Wetland Area. Mar. Freshw. Res. 2014, 65, 934. [Google Scholar] [CrossRef]
- Armentano, T.V.; Menges, E.S. Patterns of Change in the Carbon Balance of Organic Soil-Wetlands of the Temperate Zone. J. Ecol. 1986, 74, 774. [Google Scholar] [CrossRef]
- Alongi, D.M. Carbon Sequestration in Mangrove Forests. Carbon Manag. 2012, 3, 313–322. [Google Scholar] [CrossRef]
- Ezcurra, P.; Ezcurra, E.; Garcillán, P.P.; Costa, M.T.; Aburto-Oropeza, O. Coastal Landforms and Accumulation of Mangrove Peat Increase Carbon Sequestration and Storage. Proc. Natl. Acad. Sci. USA 2016, 113, 4404–4409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchio, D.A.; Savarese, M.; Bovard, B.; Mitsch, W.J. Carbon Sequestration and Sedimentation in Mangrove Swamps Influenced by Hydrogeomorphic Conditions and Urbanization in Southwest Florida. Forests 2016, 7, 116. [Google Scholar] [CrossRef] [Green Version]
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the Most Carbon-Rich Forests in the Tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- The Food and Agriculture Organization. Status and Trends in Mangrove Area Extent Worldwide; The Food and Agriculture Organization: Rome, Italy, 2003. [Google Scholar]
- Inoue, T. Carbon Sequestration in Mangroves. In Blue Carbon in Shallow Coastal Ecosystems; Springer: Singapore, 2019; pp. 73–99. [Google Scholar]
- McKee, K.L.; Cahoon, D.R.; Feller, I.C. Caribbean Mangroves Adjust to Rising Sea Level through Biotic Controls on Change in Soil Elevation. Glob. Ecol. Biogeogr. 2007, 16, 545–556. [Google Scholar] [CrossRef]
- Marois, D.E.; Mitsch, W.J. Coastal Protection from Tsunamis and Cyclones Provided by Mangrove Wetlands—A Review. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2015, 11, 71–83. [Google Scholar] [CrossRef]
- Danielsen, F.; Sørensen, M.K.; Olwig, M.F.; Selvam, V.; Parish, F.; Burgess, N.D.; Hiraishi, T.; Karunagaran, V.M.; Rasmussen, M.S.; Hansen, L.B.; et al. The Asian Tsunami: A Protective Role for Coastal Vegetation. Science 2005, 310, 643. [Google Scholar] [CrossRef] [Green Version]
- Sheng, Y.P.; Zou, R. Assessing the Role of Mangrove Forest in Reducing Coastal Inundation during Major Hurricanes. Hydrobiologia 2017, 803, 87–103. [Google Scholar] [CrossRef] [Green Version]
- Sanderman, J.; Hengl, T.; Fiske, G.; Solvik, K.; Adame, M.F.; Benson, L.; Bukoski, J.J.; Carnell, P.; Cifuentes-Jara, M.; Donato, D.; et al. A Global Map of Mangrove Forest Soil Carbon at 30 m Spatial Resolution. Environ. Res. Lett. 2018, 13, 055002. [Google Scholar] [CrossRef]
- Gilman, E.L.; Ellison, J.; Duke, N.C.; Field, C. Threats to Mangroves from Climate Change and Adaptation Options: A Review. Aquat. Bot. 2008, 89, 237–250. [Google Scholar] [CrossRef]
- Lewis, R.R.; Milbrandt, E.C.; Brown, B.; Krauss, K.W.; Rovai, A.S.; Beever, J.W.; Flynn, L.L. Stress in Mangrove Forests: Early Detection and Preemptive Rehabilitation Are Essential for Future Successful Worldwide Mangrove Forest Management. Mar. Pollut. Bull. 2016, 109, 764–771. [Google Scholar] [CrossRef]
- Villate Daza, D.A.; Moreno, H.S.; Portz, L.; Manzolli, R.P.; Bolívar-Anillo, H.J.; Anfuso, G. Mangrove Forests Evolution and Threats in the Caribbean Sea of Colombia. Water 2020, 12, 1113. [Google Scholar] [CrossRef] [Green Version]
- Kennish, M.J. Environmental Threats and Environmental Future of Estuaries. Environ. Conserv. 2002, 29, 78–107. [Google Scholar] [CrossRef]
- National Research Council. Urban Stormwater Management in the United States; National Academies Press: Washington, DC, USA, 2009. [Google Scholar]
- Mitchell, L.E. Developing a GIS of the Bioluminescent Bays on Vieques, Puerto Rico; National Geospatial Intelligence Agency: Bethesda, MD, USA, 2003. [Google Scholar]
- Schaffner, F.C.; Camacho-Fontánez, N.M.; Colón-Cruz, M.; Flusa, S.L.; Pérez-Cruz, I.J.; Rodríguez-Colón, I.; Santiago-Parrilla, R.; Torres-López, A. A First Assessment of the Effects of Hurricanes Irma and Maria on the Habitats and Terrestrial Bird Communities in the Coastal Dry Forest of Jobos; Universidad del Turabo: Gurabo, Puerto Rico, 2018. [Google Scholar]
- Baldwin, A.; Egnotovich, M.; Ford, M.; Platt, W. Regeneration in Fringe Mangrove Forests Damaged by Hurricane Andrew. Plant Ecol. 2001, 157, 149–162. [Google Scholar] [CrossRef]
- Smith, T.J.; Robblee, M.B.; Wanless, H.R.; Doyle, T.W. Mangroves, Hurricanes, and Lightning Strikes. BioScience 1994, 44, 256–262. [Google Scholar] [CrossRef]
- McKee, K.L.; Mendelssohn, I.A.; Hester, M.W. Reexamination of Pore Water Sulfide Concentrations and Redox Potentials near the Aerial Roots of Rhizophora Mangle and Avicennia Germinans. Am. J. Bot. 1988, 75, 1352–1359. [Google Scholar] [CrossRef]
- Ross, M.S.; Ruiz, P.L.; Telesnicki, G.J.; Meeder, J.F. Estimating Above-Ground Biomass and Production in Mangrove Communities of Biscayne National Park, Florida (U.S.A.). Wetl. Ecol. Manag. 2001, 9, 27–37. [Google Scholar] [CrossRef]
- Griffiths, L.N.; Mitsch, W.J. Nutrient Retention via Sedimentation in a Created Urban Stormwater Treatment Wetland. Sci. Total. Environ. 2020, 727, 138337. [Google Scholar] [CrossRef]
- Griffiths, L.N. Biogeochemical Cycling of Nutrients and Carbon in Subtropical Wetlands. Ph.D. Thesis, University of South Florida, Tampa, FL, USA, 2020. [Google Scholar]
- Komiyama, A.; Ong, J.E.; Poungparn, S. Allometry, Biomass, and Productivity of Mangrove Forests: A Review. Aquat. Bot. 2008, 89, 128–137. [Google Scholar] [CrossRef]
- Bouillon, S.; Borges, A.V.; Castañeda-Moya, E.; Diele, K.; Dittmar, T.; Duke, N.C.; Kristensen, E.; Lee, S.Y.; Marchand, C.; Middelburg, J.J.; et al. Mangrove Production and Carbon Sinks: A Revision of Global Budget Estimates. Glob. Biogeochem. Cycles 2008, 22. [Google Scholar] [CrossRef] [Green Version]
- Webb, R.M.T.; Gomez-Gomez, F. Synoptic Survey of Water Quality and Bottom Sediments, San Juan Bay Estuary System, Puerto Rico, December 1994-July 1995; US Geological Survey Water Resources Investigation; U.S. Geological Survey: Reston, VA, USA, 1998. [Google Scholar]
- Domínguez Cristóbal, C.M. La Politica Forestal Del Manglar En Puerot Rico Durante El Siglo XIX: El Caso Del Manglar de Jobos, Las Mareas, Caño Grande y Punta Caribe de Guayama. Acta Científica 2008, 22, 67–77. [Google Scholar]
- Torres-González, S.; Rodríguez, J.M. Hydrologic Conditions in the South Coast Aquifer, Puerto Rico, 2010–2015. U.S. Geological Survey: Reston, VA, USA, 2016. [Google Scholar]
- Whitall, D.R.; Costa, B.M.; Bauer, L.J.; Dieppa, A.; Hile, S.D. A Baseline Assessment of the Ecological Resources of Jobos Bay, Puerto Rico. NOAA Technichal Memorandum NOC NCCOS 133. National Oceanic and Atmospheric Administration: Silver Spring, MD, USA, 2011. [Google Scholar]
- Rovai, A.S.; Twilley, R.R.; Castañeda-Moya, E.; Riul, P.; Cifuentes-Jara, M.; Manrow-Villalobos, M.; Horta, P.A.; Simonassi, J.C.; Fonseca, A.L.; Pagliosa, P.R. Global Controls on Carbon Storage in Mangrove Soils. Nat. Clim. Chang. 2018, 8, 534–538. [Google Scholar] [CrossRef]
- Smoak, J.M.; Breithaupt, J.L.; Smith, T.J.; Sanders, C.J. Sediment Accretion and Organic Carbon Burial Relative to Sea-Level Rise and Storm Events in Two Mangrove Forests in Everglades National Park. Catena 2013, 104, 58–66. [Google Scholar] [CrossRef]
- Cahoon, D.R.; Hensel, P.; Rybczyk, J.; McKee, K.L.; Proffitt, C.E.; Perez, B.C. Mass Tree Mortality Leads to Mangrove Peat Collapse at Bay Islands, Honduras after Hurricane Mitch. J. Ecol. 2003, 91, 1093–1105. [Google Scholar] [CrossRef]
- Balco, G.; Finnegan, N.; Gendaszek, A.; Stone, J.O.; Thompson, N. Erosional Response to Northward-Propagating Crustal Thickening in the Coastal Ranges of the US Pacific Northwest. Am. J. Sci. 2013, 313, 790–806. [Google Scholar] [CrossRef]
- Alongi, D.M. Mangrove Forests: Resilience, Protection from Tsunamis, and Responses to Global Climate Change. Estuar. Coast. Shelf Sci. 2008, 76, 1–13. [Google Scholar] [CrossRef]
- Pasch, R.J.; Penny, A.B.; Berg, R. National Hurricane Center Tropical Cyclone Report: Hurricane Maria (AL152017); National Hurricane Center, National Oceanic and Atmospheric Administration: Miami, FL, USA, 2019. [Google Scholar]
- Mount, H.R.; Lynn, W.C. Soil Survey Laboratory Data and Soil Descriptions for Puerto Rico and the U.S. Virgin Islands: Soil Survey Investigations Report No. 49. United States Department of Agriculture, Natural Resources Conservation Service, National Soil Survey Center: Honolulu, HI, USA, 2004. [Google Scholar]
- D’Aluisio-Guerrieri, G.M.; Davis, R.A., Jr. Infilling of Coastal Lagoons by Terrigenous Siliciclastic and Marine Carbonate Sediment: Vieques, Puerto Rico. Dev. Sedimentol. 1988, 42, 207–230. [Google Scholar]
- Bush, D.M. Mixed Carbonate/Siliciclastic Sedimentation: Northern Insular Shelf of Puerto Rico. In Proceedings of the Mixed Carbonate-Siliciclastic Sequences; Lomardo, A.J., Harris, P.M., Eds.; SEPM Society for Sedimentary Geology: Tulsa, OK, USA, 1991; pp. 447–484. [Google Scholar]
- Lugo, A.E.; González, O.M.R.; Pedraza, C.R. The Río Piedras Watershed and Its Surrounding Environment. FS-980; U.S. Department of Agriculture, Forest Service, International Institute of Tropical Forestry: San Juan, Puerto Rico, 2011; pp. 1–46. [Google Scholar]
- Kennaway, T.; Helmer, E.H. The Forest Types and Ages Cleared for Land Development in Puerto Rico. GIScience Remote Sens. 2007, 44, 356–382. [Google Scholar] [CrossRef]
- Rivera De Jesus, W.O. Eficiencia En El Uso de Los Nutrientes: Nitrógeno (N), Fósforo (P) y Potasio (K) En Especies Arbóreas Utilizadas En Las Áreas Mitigadas de La Ciénaga Las Cucharillas. Ph.D. Thesis, Universidad Metropolitana, Atlántico, Colombia, 2011. [Google Scholar]
- O’Connell, S.; Ku, T.I.M.C.; Martini, A. The Hydrodynamics and Biogeochemistry of Bioluminescent Bays, Vieques, Puerto Rico, Proceedings of the 20th Annual Keck Research Symposium in Geology 2007; Sidor, S., Ed.; Keck Geology Consortium: Wooster, OH, USA, 2007; pp. 137–142. [Google Scholar]
- Viruet, Y.S. Land Use/Land Cover of Two Bioluminescent Bays in Puerto Rico. Bachelor’s Thesis, University of Puerto Rico, San Juan, Puerto Rico, 2002; pp. 1–26. [Google Scholar]
- Woodoruff, J.D.; Donnelly, J.P.; Mohrig, D.; Geyer, W.R. Reconstructing relative flooding intensities responsible for hurricane—induced deposits from laguna Playa Grande, Vieques, Puerto Rico. Geology 2008, 36, 391–394. [Google Scholar] [CrossRef]
- Martínex-Colón, M.; Hallock, P.; Green-Ruíz, C.R.; Smoak, J.M. Temporal variability in potentially toxic elements (PTE’s) and benthic Foraminifera in an estuarine environment in Puerto Rico. Micropaleontology 2017, 63, 357–382. [Google Scholar]
- Imbert, D.; Rollet, B. Phytmassaerienne et Production Primaire Dans La Mangrove Du Grand Cul-de-Sac Marine (Guadeloupe, Antilles Francaises). Bull. Ecol. 1989, 20, 27–39. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cucharillas | Jobos Bay | Vieques | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Coastal | Inland | Coastal | Inland | Coastal | Inland | ||||||||
 |  |  | |||||||||||
| 1 | 2 | 1 | 2 | 3 | 1 | 2 | 3 | 4 | 5 | ||||
| Tree density (stems m−2) | R. mangle | - | - | 0.10 | 0.10 | - | - | - | 0.32 | 0.16 | - | ||
| A. germinans | - | - | 0.05 | 0.18 | 0.03 | - | - | - | - | - | |||
| L. racemosa | 0.15 | 0.08 | 0.28 | 0.43 | - | - | - | - | - | - | |||
| Mean DBH (cm) ± SE | R. mangle | - | - | 2.1 ± 0.2 (4) | 2.1 ± 0.3 (4) | - | - | - | 15.4 ± 9.9 (2) | 13.0 (1) | - | ||
| A. germinans | - | - | 8.4 ± 0.9 (2) | 8.0 ± 2.5 (7) | 1.6 (1) | - | - | - | - | - | |||
| L. racemosa | 17.6 ± 4.9 (6) | 29.3 ± 7.3 (2) | 3.3 ± 0.3 (11) | 2.1 ± 0.2 (17) | - | - | - | - | - | - | |||
| Aboveground biomass (kg m−2) | R. mangle | - | - | 0.12 ± 0.01 (4) | 0.12 ± 0.01 (4) | - | - | - | 67.1 ± 28.1 (2) | 16.1 (1) | - | ||
| A. germinans | - | - | 1.72 ± 0.14 (2) | 7.49 ± 0.72 (7) | 0.01 (1) | - | - | - | - | - | |||
| L. racemosa | 30.0 ± 3.0 (6) | 21.8 ± 5.6 (2) | 0.93 ± 0.02 (11) | 0.59 ± 0.01 (17) | - | - | - | - | - | - | |||
| Total | 30.0 ± 3.0 (6) | 21.8 ± 5.6 (2) | 2.77 ± 0.04 (17) | 8.20 ± 0.17 (28) | 0.01 (1) | - | - | 67.1 ± 28.1 (2) | 16.1 (1) | - | |||
| Aboveground carbon stock (kg-C m−2) | R. mangle | - | - | 0.06 ± 0.01 (4) | 0.05 ± 0.01 (4) | - | - | - | 30.2 ± 12.6 (2) | 7.23 (1) | - | ||
| A. germinans | - | - | 0.77 ± 0.06 (2) | 3.37 ± 0.32 (7) | 0.003 (1) | - | - | - | - | - | |||
| L. racemosa | 13.5 ± 1.3 (6) | 9.82 ± 2.52 (2) | 0.42 ± 0.01 (11) | 0.26 ± 0.01 (17) | - | - | - | - | - | - | |||
| Total | 13.5 ± 1.3 (6) | 9.82 ± 2.52 (2) | 1.25 ± 0.02 (17) | 3.69 ± 0.08 (28) | 0.003 (1) | - | - | 30.2 ± 12.6 (2) | 7.23 (1) | - | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Griffiths, L.N.; Hernandez, E.; Cuevas, E.; Mitsch, W.J. Above- and Below-Ground Carbon Storage of Hydrologically Altered Mangrove Wetlands in Puerto Rico after a Hurricane. Plants 2021, 10, 1965. https://doi.org/10.3390/plants10091965
Griffiths LN, Hernandez E, Cuevas E, Mitsch WJ. Above- and Below-Ground Carbon Storage of Hydrologically Altered Mangrove Wetlands in Puerto Rico after a Hurricane. Plants. 2021; 10(9):1965. https://doi.org/10.3390/plants10091965
Chicago/Turabian StyleGriffiths, Lauren N., Elix Hernandez, Elvira Cuevas, and William J. Mitsch. 2021. "Above- and Below-Ground Carbon Storage of Hydrologically Altered Mangrove Wetlands in Puerto Rico after a Hurricane" Plants 10, no. 9: 1965. https://doi.org/10.3390/plants10091965
APA StyleGriffiths, L. N., Hernandez, E., Cuevas, E., & Mitsch, W. J. (2021). Above- and Below-Ground Carbon Storage of Hydrologically Altered Mangrove Wetlands in Puerto Rico after a Hurricane. Plants, 10(9), 1965. https://doi.org/10.3390/plants10091965