
Insight into Analysis of Essential Oil from Anisosciadium lanatum Boiss.—Chemical Composition, Molecular Docking, and Mitigation of Hepg2 Cancer Cells through Apoptotic Markers

 , ,
, ,  and
and
Abstract
1. Introduction
2. Results
2.1. Isolation and Identification of Chemical Components of EO
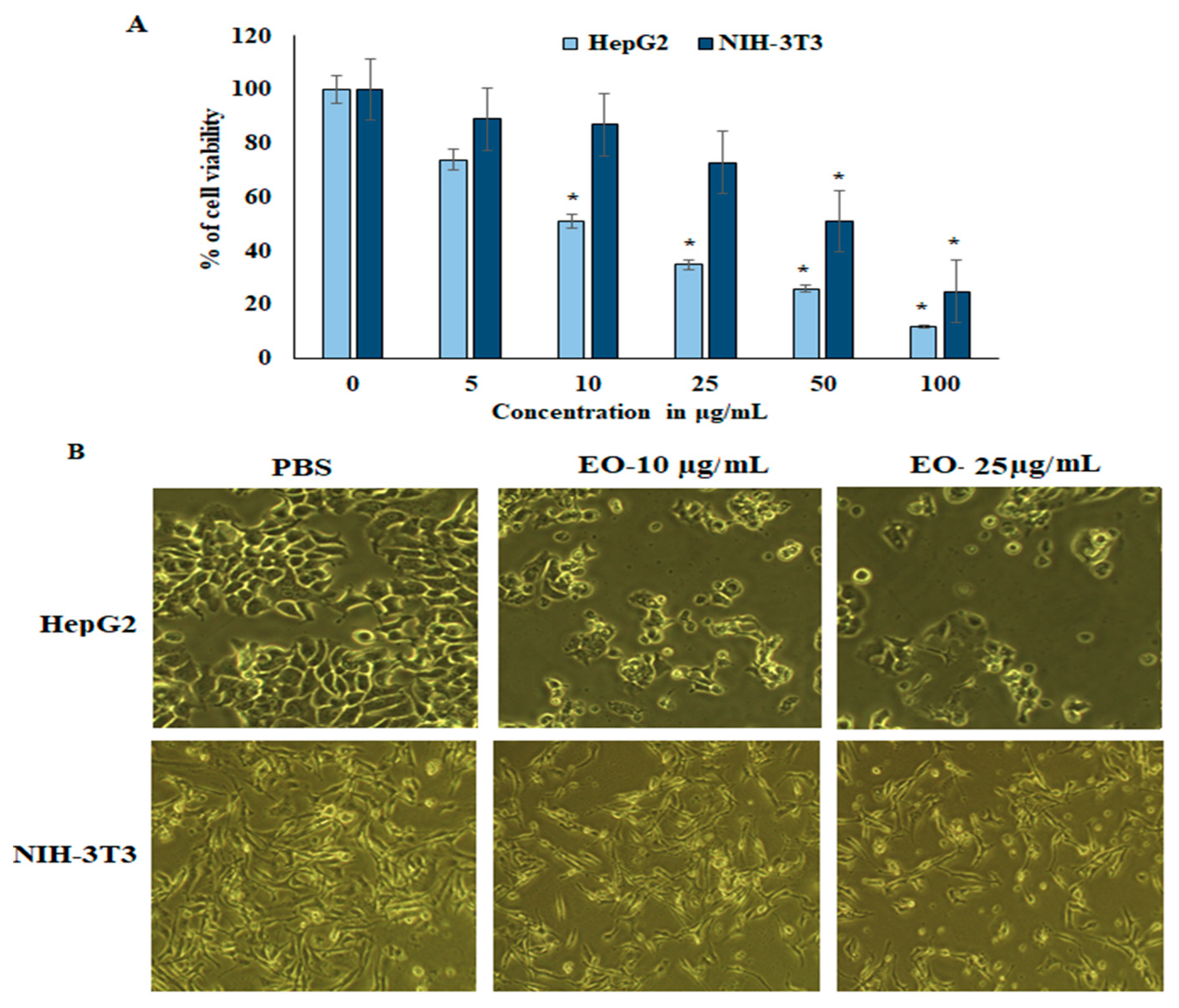
2.2. Proliferation and Cell Viability Assay
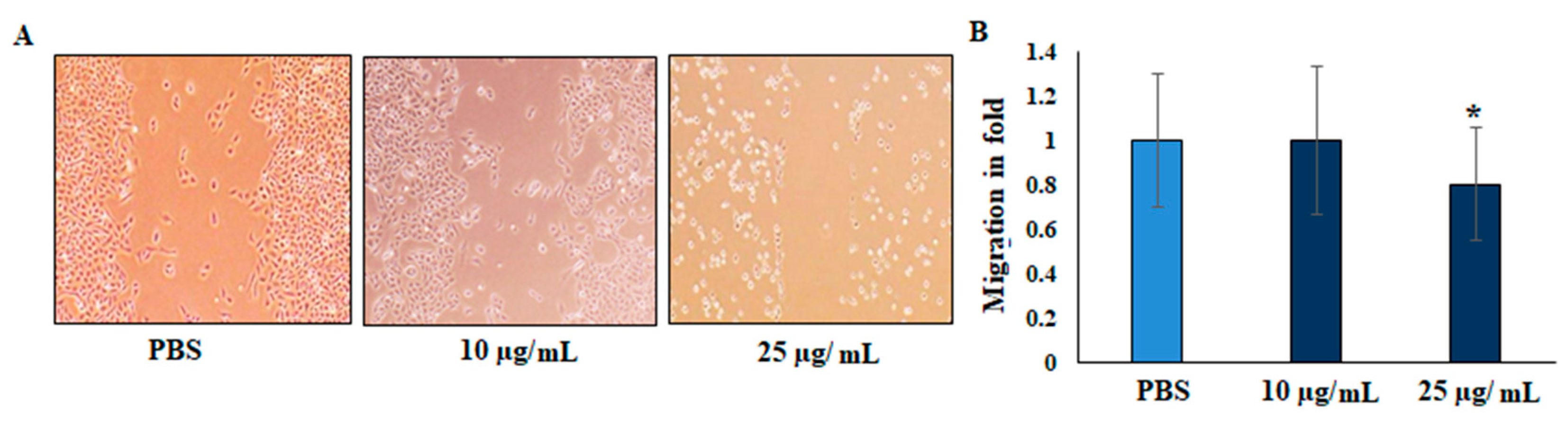
2.3. Migration (Scratch Wound Assay)
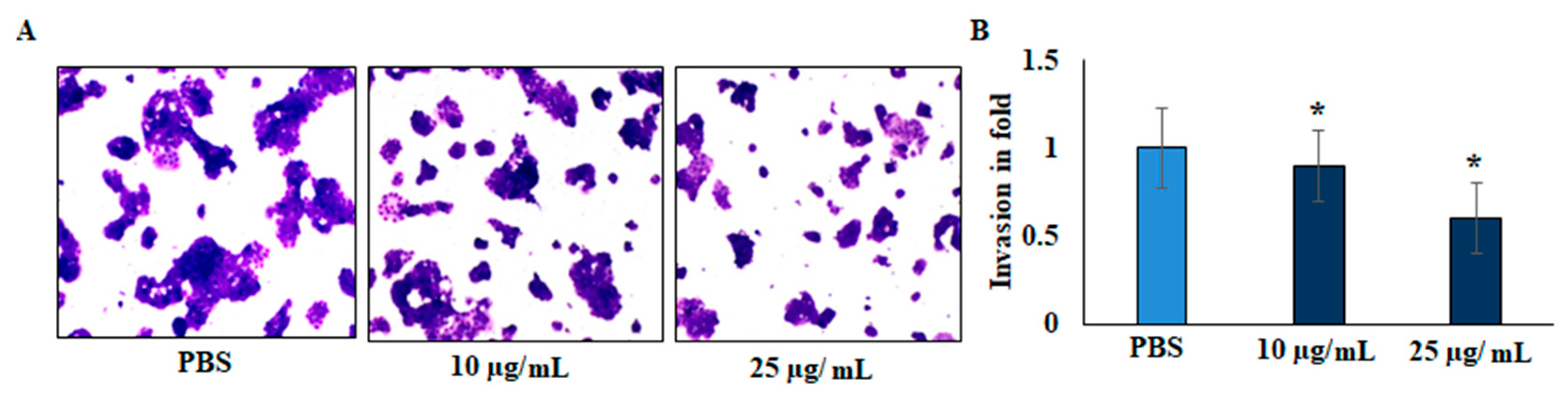
2.4. Invasion (Transwell Assay)
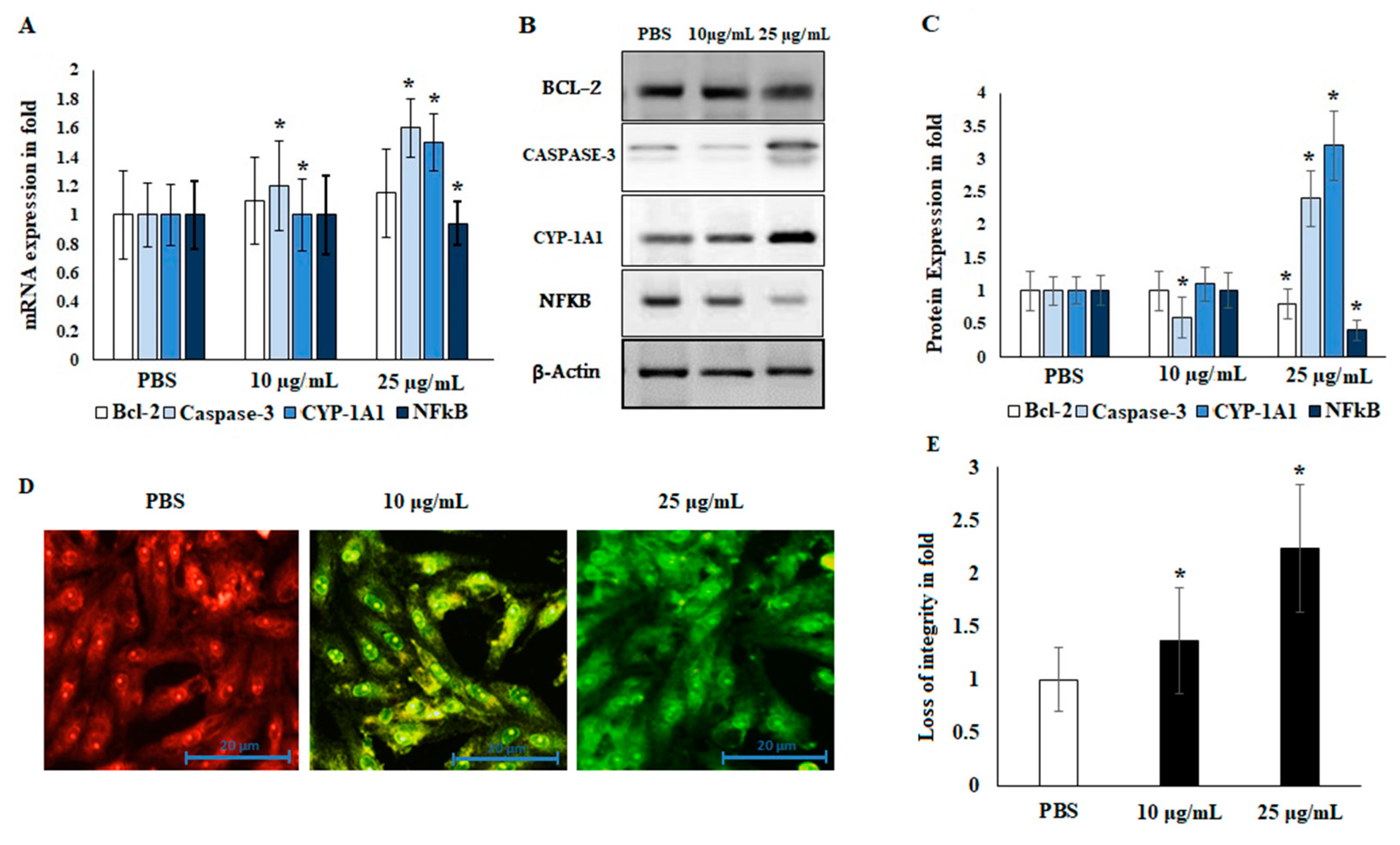
2.5. Immunoblotting and Localization of Cytochrome-c
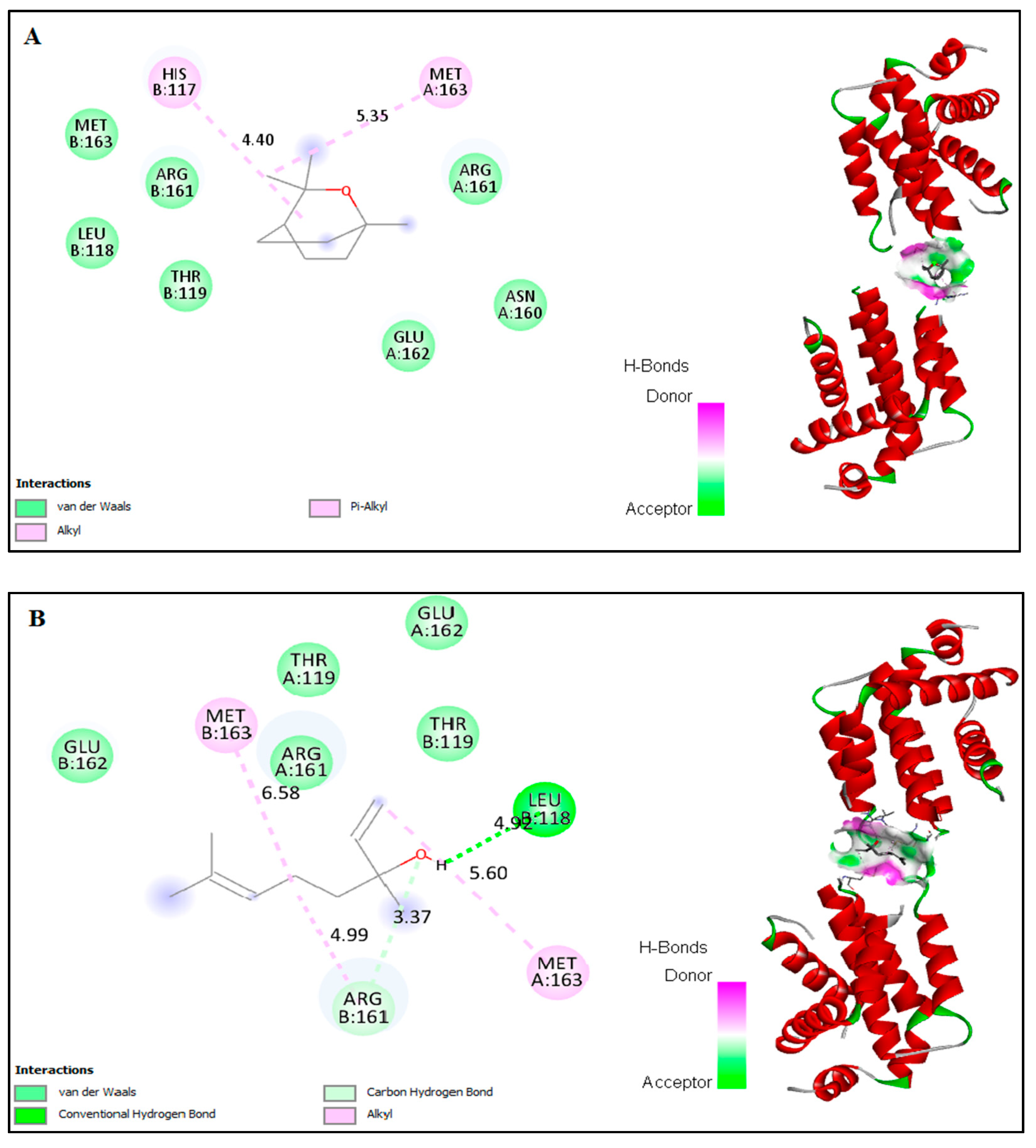
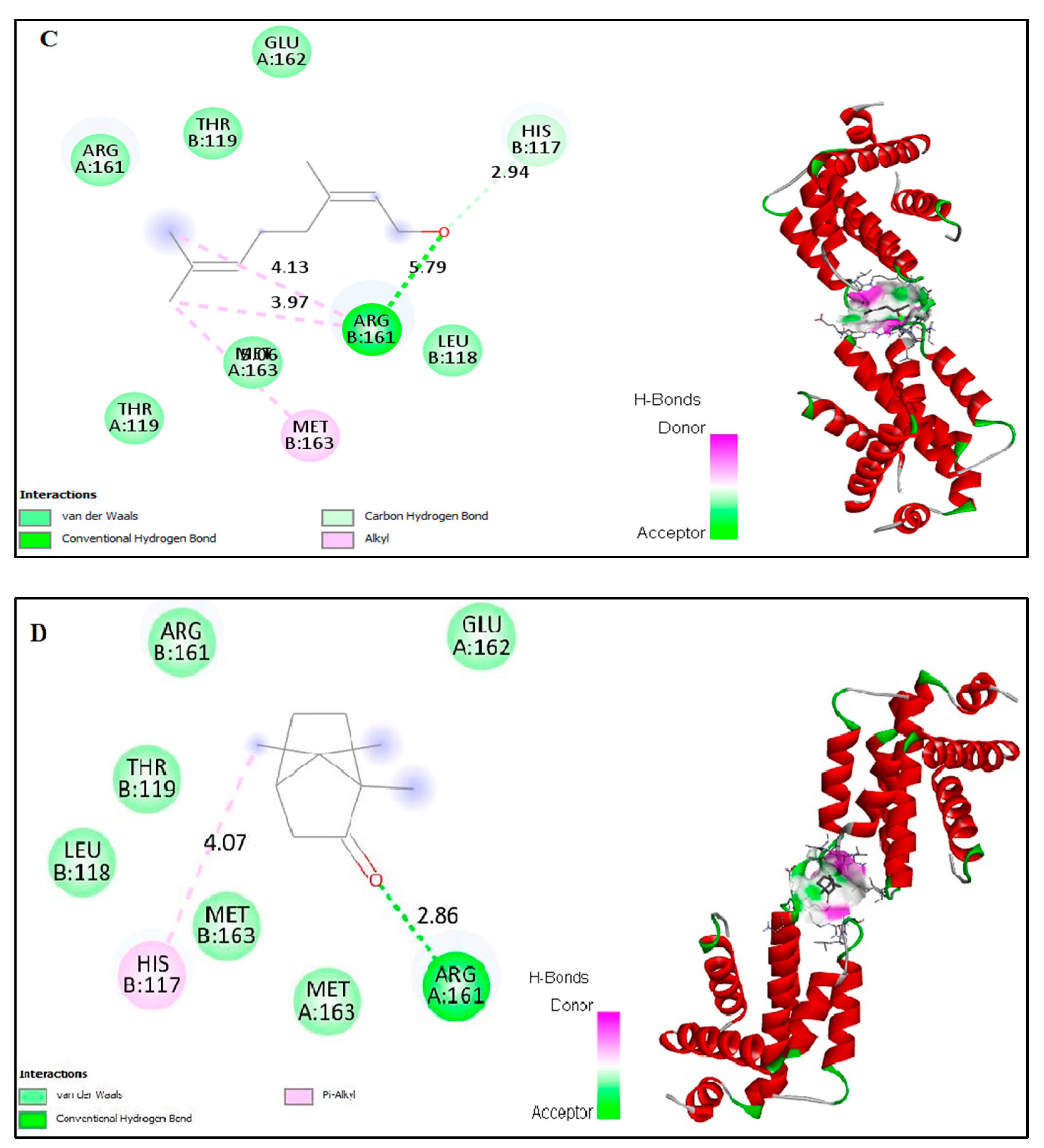
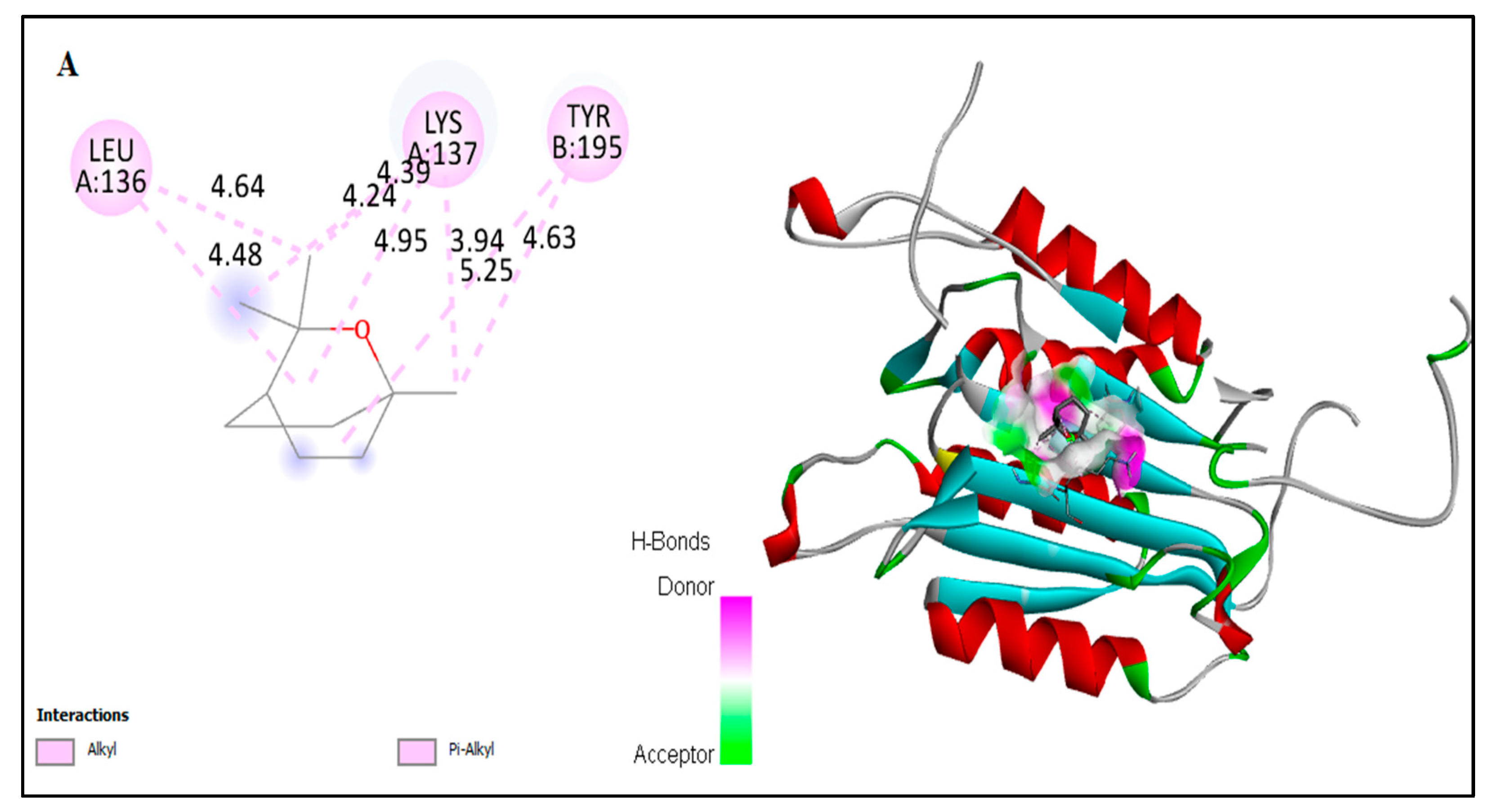
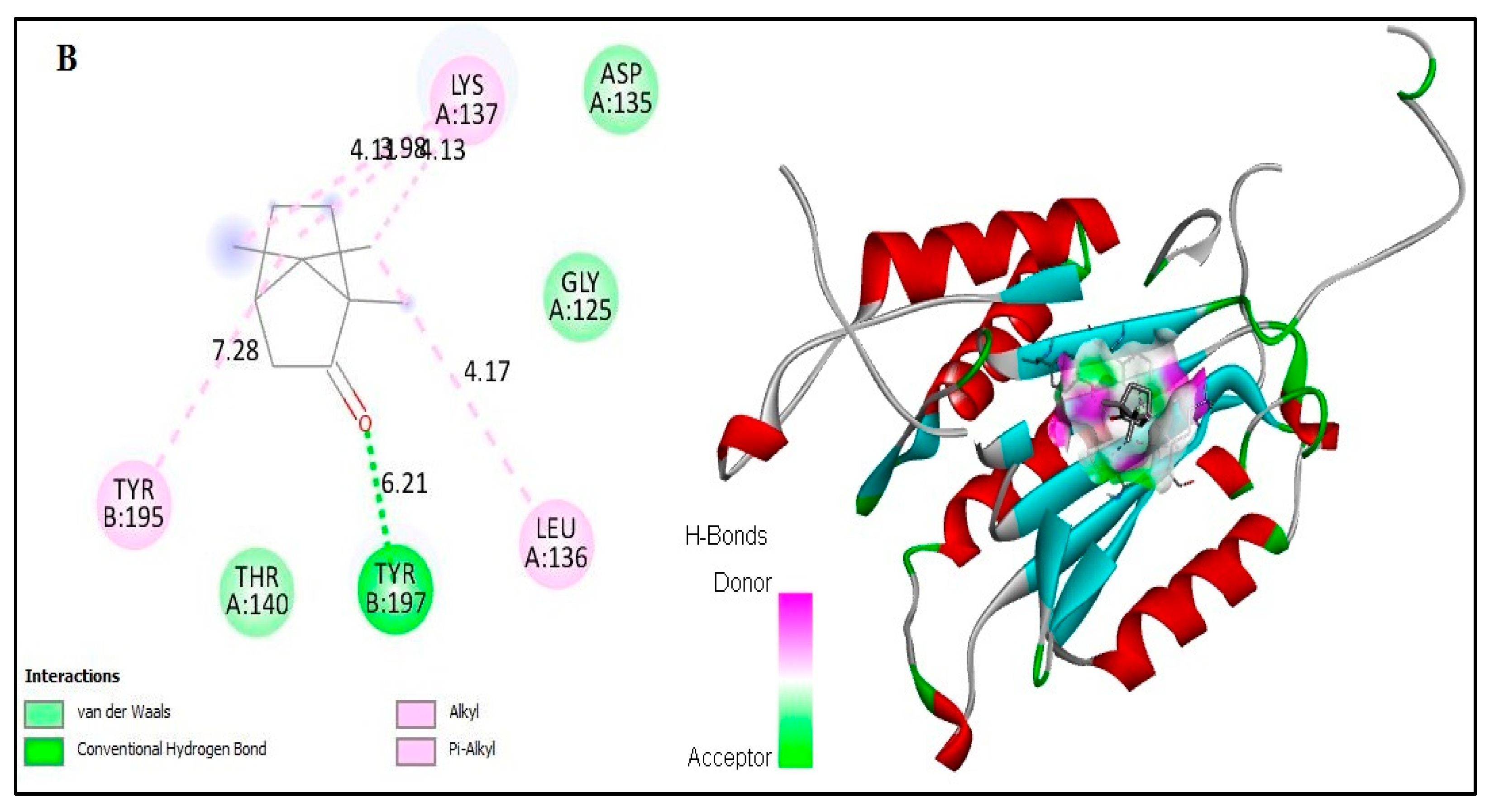
2.6. In Silico Docking of Major Constituents against BCL-2 and CASPASE-3
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Extraction of Essential Oil
4.3. Essential Oil Analysis
4.4. Cell Culture and MTT Assay
4.5. Migration (Scratch-Wound Assay)
4.6. Invasion (Transwell Assay)
4.7. Immunoblotting
4.8. Mitochondrial Membrane Potential (MMP) Assessment for Localization of Cytochrome-c
4.9. mRNA Expression
4.10. Computational Docking Analysis
4.10.1. Protein Preparation
4.10.2. Ligand Preparation
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Abd-ElGawad, A.M.; Elgamal, A.M.; Ei-Amier, Y.A.; Mohamed, T.A.; El Gendy, A.E.-N.G.; Elshamy, A.I. Chemical Composition, Allelopathic, Antioxidant, and Anti-Inflammatory Activities of Sesquiterpenes Rich Essential Oil of Cleome amblyocarpa Barratte & Murb. Plants 2021, 10, 1294. [Google Scholar] [PubMed]
- Camele, I.; Gruľová, D.; Elshafie, H.S. Chemical Composition and Antimicrobial Properties of Mentha× piperita cv.‘Kristinka’Essential Oil. Plants 2021, 10, 1567. [Google Scholar] [CrossRef]
- Abd-ElGawad, A.M.; Bonanomi, G.; Al-Rashed, S.A.; Elshamy, A.I. Persicaria lapathifolia Essential Oil: Chemical Constituents, Antioxidant Activity, and Allelopathic Effect on the Weed Echinochloa colona. Plants 2021, 10, 1798. [Google Scholar] [CrossRef]
- Mohamed, M.E.; Mohafez, O.M.; Khalil, H.E.; Alhaider, I.A. Essential Oil from Myrtle Leaves Growing in the Eastern Part of Saudi Arabia: Components, Anti-inflammatory and Cytotoxic Activities. J. Essent. Oil-Bear. Plants 2019, 22, 877–892. [Google Scholar] [CrossRef]
- Khalil, H.E.; Alqahtani, N.K.; Darrag, H.M.; Ibrahim, H.-I.M.; Emeka, P.M.; Badger-Emeka, L.I.; Matsunami, K.; Shehata, T.M.; Elsewedy, H.S. Date Palm Extract (Phoenix dactylifera) PEGylated Nanoemulsion: Development, Optimization and Cytotoxicity Evaluation. Plants 2021, 10, 735. [Google Scholar] [CrossRef]
- Khalil, H.E.; Mohamed, M.E.; Morsy, M.A.; Kandeel, M. Flavonoid and phenolic compounds from Carissa macrocarpa: Molecular docking and cytotoxicity studies. Pharmacogn. Mag. 2018, 14, 304. [Google Scholar] [CrossRef]
- Harvey, A.L. Natural products in drug discovery. Drug Discov. Today 2008, 13, 894–901. [Google Scholar] [CrossRef]
- Bamia, C. Dietary patterns in association to cancer incidence and survival: Concept, current evidence, and suggestions for future research. Eur. J. Clin. Nutr. 2018, 72, 818–825. [Google Scholar] [CrossRef] [PubMed]
- Clinton, S.K.; Giovannucci, E.L.; Hursting, S.D. The World Cancer Research Fund/American Institute for Cancer Research third expert report on diet, nutrition, physical activity, and cancer: Impact and future directions. J. Nutr. 2020, 150, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Karageorgou, D.; Magriplis, E.; Mitsopoulou, A.; Dimakopoulos, I.; Bakogianni, I.; Micha, R.; Michas, G.; Chourdakis, M.; Ntouroupi, T.; Tsaniklidou, S. Dietary patterns and lifestyle characteristics in adults: Results from the Hellenic National Nutrition and Health Survey (HNNHS). Public Health 2019, 171, 76–88. [Google Scholar] [CrossRef]
- Buriani, A.; Fortinguerra, S.; Sorrenti, V.; Dall’Acqua, S.; Innocenti, G.; Montopoli, M.; Gabbia, D.; Carrara, M. Human adenocarcinoma cell line sensitivity to essential oil phytocomplexes from Pistacia species: A multivariate approach. Molecules 2017, 22, 1336. [Google Scholar] [CrossRef]
- Amiri, M.S.; Joharchi, M.R. Ethnobotanical knowledge of Apiaceae family in Iran: A review. Avicenna J. Phytomed. 2016, 6, 621. [Google Scholar]
- Sayed-Ahmad, B.; Talou, T.; Saad, Z.; Hijazi, A.; Merah, O. The Apiaceae: Ethnomedicinal family as source for industrial uses. Ind. Crops. Prod. 2017, 109, 661–671. [Google Scholar] [CrossRef]
- Christensen, L.P.; Brandt, K. Bioactive polyacetylenes in food plants of the Apiaceae family: Occurrence, bioactivity and analysis. J. Pharm. Biomed. Anal. 2006, 41, 683–693. [Google Scholar] [CrossRef]
- Vieira, J.N.; Gonçalves, C.; Villarreal, J.; Gonçalves, V.; Lund, R.; Freitag, R.; Silva, A.; Nascente, P. Chemical composition of essential oils from the apiaceae family, cytotoxicity, and their antifungal activity in vitro against candida species from oral cavity. Braz. J. Biol. 2018, 79, 432–443. [Google Scholar] [CrossRef] [PubMed]
- Chizzola, R. Essential oil composition of wild growing Apiaceae from Europe and the Mediterranean. Nat. Prod. Commun. 2010, 5, 1934578X1000500925. [Google Scholar] [CrossRef]
- Masroorbabanari, M.; Miri, R.; Firuzi, O.; Jassbi, A.R. Antioxidant and Cytotoxic Activities of the Essential Oil and Extracts of Anisosciadium orientale. J. Chem. Soc. Pak. 2014, 36, 457–461. [Google Scholar]
- Rowshan, V.; Hatami, A.; Bahmanzadegan, A.; Yazdani, M. Chemical Constituents of the Essential Oil from Aerial Parts and Fruit of Anisosciadium Orientale. Nat. Prod. Commun. 2012, 7, 1934578X1200700127. [Google Scholar] [CrossRef]
- Sher, H.; Aldosari, A. Ethnobotanical survey on plants of veterinary importance around Al-Riyadh (Saudi Arabia). Afr. J. Pharm. Pharmacol. 2013, 7, 1404–1410. [Google Scholar] [CrossRef]
- Al-Mazroa, S. Chemical constituents of A. Lanatum, H. Tuberculatum, A. Garcini, S. Spinosa, H. Bacciferum, and A. Ludwigh grown in Saudi Arabia. J. Saudi Chem. 2003, 7, 255–258. [Google Scholar]
- Al-Yemeny, M.N.; Al-Yemeny, A. A check list of weeds in Al-kharj area of Saudi Arabia. Pak. J. Biol. Sci. 1999, 2, 7–13. [Google Scholar] [CrossRef]
- Joachim, W.; Kadereit, J.W.; Bittrich, V. The Families and Genera of Vascular Plants; Springer International Publishing AG: Steinhausen, Switzerland, 2018; p. 101. [Google Scholar]
- Mandaville, J.P. Bedouin Ethnobotany: Plant Concepts and Uses in a Desert Pastoral World; University of Arizona Press: Tucson, AZ, USA, 2019; p. 355. [Google Scholar]
- Middleditch, B.S. Kuwaiti Plants: Distribution, Traditional Medicine, Pytochemistry, Pharmacology and Economic Value; Elsevier: New York, NY, USA, 1991; p. 10. [Google Scholar]
- El-Sayed, W.M.; Hussin, W.A.; Mahmoud, A.A.; AlFredan, M.A. Antimutagenic Activities of Anisosciadium lanatum Extracts Could Predict the Anticancer Potential in Different Cell Lines. Int. J. Pharm. Pharm. Res. 2017, 9, 197–206. [Google Scholar] [CrossRef][Green Version]
- Mahmoud, A.A.; El-Sayed, W.M. The anti-proliferative activity of anisosciadone: A new guaiane sesquiterpene from Anisosciadium lanatum. Anti-Cancer Agents Med. Chem. 2019, 19, 1114–1119. [Google Scholar] [CrossRef] [PubMed]
- Mancianti, F.; Ebani, V.V. Biological activity of essential oils. Molecules 2020, 25, 678. [Google Scholar] [CrossRef]
- De Souza, E.L. The effects of sublethal doses of essential oils and their constituents on antimicrobial susceptibility and antibiotic resistance among food-related bacteria: A review. Trends Food Sci. Technol. 2016, 56, 1–12. [Google Scholar] [CrossRef]
- Spyridopoulou, K.; Fitsiou, E.; Bouloukosta, E.; Tiptiri-Kourpeti, A.; Vamvakias, M.; Oreopoulou, A.; Papavassilopoulou, E.; Pappa, A.; Chlichlia, K. Extraction, chemical composition, and anticancer potential of Origanum onites L. essential oil. Molecules 2019, 24, 2612. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Sureda, A.; Tenore, G.C.; Daglia, M.; Sharifi-Rad, M.; Valussi, M.; Tundis, R.; Sharifi-Rad, M.; Loizzo, M.R.; Ademiluyi, A.O. Biological activities of essential oils: From plant chemoecology to traditional healing systems. Molecules 2017, 22, 70. [Google Scholar] [CrossRef]
- Pan, W.; Zhang, G. Linalool monoterpene exerts potent antitumor effects in OECM 1 human oral cancer cells by inducing sub-G1 cell cycle arrest, loss of mitochondrial membrane potential and inhibition of PI3K/AKT biochemical pathway. J. Buon 2019, 24, 323–328. [Google Scholar]
- Bayala, B.; Bassole, I.H.; Scifo, R.; Gnoula, C.; Morel, L.; Lobaccaro, J.-M.A.; Simpore, J. Anticancer activity of essential oils and their chemical components-a review. Am. J. Cancer Res. 2014, 4, 591. [Google Scholar] [PubMed]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Candrasari, D.S.; Mubarika, S.; Wahyuningsih, M.S.H. The effect of a-terpineol on cell cycle, apoptosis and Bcl-2 family protein expression of breast cancer cell line MCF-7. J. Med. Sci. 2015, 47, 59–67. [Google Scholar]
- Kim, D.Y.; Kang, M.-K.; Lee, E.-J.; Kim, Y.-H.; Oh, H.; Kim, S.-I.; Oh, S.Y.; Na, W.; Kang, Y.-H. Eucalyptol Inhibits Amyloid-β-Induced Barrier Dysfunction in Glucose-Exposed Retinal Pigment Epithelial Cells and Diabetic Eyes. Antioxidants 2020, 9, 1000. [Google Scholar] [CrossRef]
- Gunaseelan, S.; Balupillai, A.; Govindasamy, K.; Ramasamy, K.; Muthusamy, G.; Shanmugam, M.; Thangaiyan, R.; Robert, B.M.; Prasad Nagarajan, R.; Ponniresan, V.K. Linalool prevents oxidative stress activated protein kinases in single UVB-exposed human skin cells. PLoS ONE 2017, 12, e0176699. [Google Scholar] [CrossRef]
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular principles of metastasis: A hallmark of cancer revisited. Signal Transduct. Target. Ther. 2020, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.-I.M.; Ismail, M.B.; Ammar, R.B.; Ahmed, E.A. Thidiazuron suppresses breast cancer via targeting miR-132 and dysregulation of the PI3K–Akt signaling pathway mediated by the miR-202-5p–PTEN axis. Biochem. Cell Biol. 2021, 99, 1–11. [Google Scholar] [CrossRef]
- Zhou, M.; Zhang, Q.; Zhao, J.; Liao, M.; Wen, S.; Yang, M. Phosphorylation of Bcl-2 plays an important role in glycochenodeoxycholate-induced survival and chemoresistance in HCC. Oncol. Rep. 2017, 38, 1742–1750. [Google Scholar] [CrossRef]
- Tsuchiya, S.; Tsuji, M.; Morio, Y.; Oguchi, K. Involvement of endoplasmic reticulum in glycochenodeoxycholic acid-induced apoptosis in rat hepatocytes. Toxicol. Lett. 2006, 166, 140–149. [Google Scholar] [CrossRef]
- Hata, A.N.; Engelman, J.A.; Faber, A.C. The BCL2 family: Key mediators of the apoptotic response to targeted anticancer therapeutics. Cancer Discov. 2015, 5, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Maji, S.; Panda, S.; Samal, S.K.; Shriwas, O.; Rath, R.; Pellecchia, M.; Emdad, L.; Das, S.K.; Fisher, P.B.; Dash, R. Bcl-2 antiapoptotic family proteins and chemoresistance in cancer. Adv. Cancer Res. 2018, 137, 37–75. [Google Scholar]
- Kumar, V.S.; Kumaresan, S.; Tamizh, M.M.; Islam, M.I.H.; Thirugnanasambantham, K. Anticancer potential of NF-κB targeting apoptotic molecule “flavipin” isolated from endophytic Chaetomium globosum. Phytomedicine 2019, 61, 152830. [Google Scholar] [CrossRef] [PubMed]
- Pan, R.; Hogdal, L.J.; Benito, J.M.; Bucci, D.; Han, L.; Borthakur, G.; Cortes, J.; DeAngelo, D.J.; Debose, L.; Mu, H. Selective BCL-2 inhibition by ABT-199 causes on-target cell death in acute myeloid leukemia. Cancer Discov. 2014, 4, 362–375. [Google Scholar] [CrossRef]
- Zhang, S.; Kim, K.; Jin, U.H.; Pfent, C.; Cao, H.; Amendt, B.; Liu, X.; Wilson-Robles, H.; Safe, S. Aryl hydrocarbon receptor agonists induce microRNA-335 expression and inhibit lung metastasis of estrogen receptor negative breast cancer cells. Mol. Cancer Ther. 2012, 11, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Hanieh, H.; Mohafez, O.; Hairul-Islam, V.I.; Alzahrani, A.; Bani Ismail, M.; Thirugnanasambantham, K. Novel aryl hydrocarbon receptor agonist suppresses migration and invasion of breast cancer cells. PLoS ONE 2016, 11, e0167650. [Google Scholar] [CrossRef] [PubMed]
- Gautam, N.; Mantha, A.K.; Mittal, S. Essential oils and their constituents as anticancer agents: A mechanistic view. Biomed. Res. Int. 2014, 2014, 154106. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, J.; Tan, J.; Wang, M.; Yang, J.; Zhang, Z.-M.; Li, C.; Basnakian, A.G.; Tang, H.-W.; Perrimon, N. Endonuclease G promotes autophagy by suppressing mTOR signaling and activating the DNA damage response. Nat. Commun. 2021, 12, 476. [Google Scholar] [CrossRef]
- Zhang, X.; Fan, C.; Zhang, H.; Zhao, Q.; Liu, Y.; Xu, C.; Xie, Q.; Wu, X.; Yu, X.; Zhang, J. MLKL and FADD are critical for suppressing progressive lymphoproliferative disease and activating the NLRP3 inflammasome. Cell Rep. 2016, 16, 3247–3259. [Google Scholar] [CrossRef] [PubMed]
- Kroemer, G.; Galluzzi, L.; Brenner, C. Mitochondrial membrane permeabilization in cell death. Physiol. Rev. 2007, 87, 99–163. [Google Scholar] [CrossRef]
- Noor, Z.I.; Nithyanan, A.; Nur, N.M.Z.; Hasni, A. Molecular docking of selected compounds from Clinacanthus Nutans with Bcl-2, P53, Caspase-3 and Caspase-8 proteins in the apoptosis pathway. J. Biol. Sci. Opin. 2020, 8, 4–11. [Google Scholar]
- Darrag, H.M.; Alhajhoj, M.R.; Khalil, H.E. Bio-Insecticide of Thymus vulgaris and Ocimum basilicum Extract from Cell Suspensions and Their Inhibitory Effect against Serine, Cysteine, and Metalloproteinases of the Red Palm Weevil (Rhynchophorus ferrugineus). Insects 2021, 12, 405. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; Volume 456. [Google Scholar]
- Ibrahim, H.-I.M.; Darrag, H.M.; Alhajhoj, M.R.; Khalil, H.E. Biomolecule from Trigonella stellata from Saudi Flora to Suppress Osteoporosis via Osteostromal Regulations. Plants 2020, 9, 1610. [Google Scholar] [CrossRef]
- Marvibaigi, M.; Amini, N.; Supriyanto, E.; Abdul Majid, F.A.; Kumar Jaganathan, S.; Jamil, S.; Hamzehalipour Almaki, J.; Nasiri, R. Antioxidant activity and ROS-dependent apoptotic effect of Scurrula ferruginea (Jack) danser methanol extract in human breast cancer cell MDA-MB-231. PLoS ONE 2016, 11, e0158942. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, E.A.; Ibrahim, H.-I.M.; Khalil, H.E. Pinocembrin Reduces Arthritic Symptoms in Mouse Model via Targeting Sox4 Signaling Molecules. J. Med. Food. 2021, 24, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Sivamani, P.; Singaravelu, G.; Thiagarajan, V.; Jayalakshmi, T.; Kumar, G.R. Comparative molecular docking analysis of essential oil constituents as elastase inhibitors. Bioinformation 2012, 8, 457–460. [Google Scholar] [CrossRef] [PubMed]
- Kandeel, M.; Al-Taher, A. Metabolic drug targets of the cytosine metabolism pathways in the dromedary camel (Camelus dromedarius) and blood parasite Trypanosoma evansi. Trop. Anim. Health Prod. 2020, 52, 3337–3358. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Constituents | Rt(min) | RI(Exp) | RI(Lit) | RA% | MS |
|---|---|---|---|---|---|---|
| 1 | α-pinene | 9.03 | 937 | 932 | 10.14 ± 0.6 | 136.2340 |
| 2 | camphene | 9.29 | 952 | 946 | 4.4 ± 0.1 | 136.2340 |
| 3 | β-pinene | 9.76 | 977 | 977 | 6.2 ± 0.09 | 136.2340 |
| 4 | β-myrcene | 9.99 | 991 | 988 | 8.45 ± 0.2 | 136.2340 |
| 5 | α-phellandrene | 10.28 | 1005 | 1002 | 0.2 ± 0.02 | 136.2340 |
| 6 | car-4-ene | 13.41 | 1009 | 1004 | 5.8 ± 0.2 | 136.2340 |
| 7 | α-terpinene | 13.53 | 1017 | 1014 | 3.62 ± 0.07 | 136.2340 |
| 8 | limonene | 13.74 | 1030 | 1224 | 6.7 ± 0.3 | 136.2340 |
| 9 | p-cymene | 13.79 | 1026 | 1023 | 6.58 ± 0.1 | 134.2182 |
| 10 | terpinolene | 15.09 | 1088 | 1086 | 2.07 ± 0.07 | 136.2340 |
| 11 | eucalyptol | 15.36 | 1031 | 1031 | 6.35 ± 0.2 | 154.2493 |
| 12 | linalool | 16.37 | 1099 | 1095 | 4.34 ± 0.1 | 154.2493 |
| 13 | camphor | 16.57 | 1145 | 1141 | 4.3 ± 0.03 | 152.2334 |
| 14 | isoborneol | 17.03 | 1167 | 1165 | 0.1 ± 0.04 | 154.2493 |
| 15 | terpinen-4-ol | 17.29 | 1177 | 1174 | 2.4 ± 0.2 | 154.2493 |
| 16 | α-terpineol | 17.76 | 1189 | 1186 | 1.5 ± 0.05 | 154.2493 |
| 17 | fenchyl acetate | 17.81 | 1214 | 1214 | 0.1 ± 0.01 | 196.2860 |
| 18 | nerol | 17.92 | 1228 | 1227 | 3.1 ± 0.1 | 154.2493 |
| 19 | bornyl acetate | 18.57 | 1285 | 1284 | 0.6 ± 0.04 | 196.2860 |
| 20 | methyl geranate | 18.61 | 1321 | 1319 | 0.2 ± 0.02 | 182.2594 |
| 21 | neryl acetate | 18.74 | 1364 | 1359 | 0.1 ± 0.02 | 196.2860 |
| 22 | α-copaene | 19.56 | 1376 | 1374 | 0.9 ± 0.1 | 204.3511 |
| 23 | β-cubebene | 19.89 | 1389 | 1387 | 1.3 ± 0.2 | 204.3511 |
| 24 | β-caryophyllene | 20.45 | 1424 | 1424 | 7.25 ± 0.4 | 204.3511 |
| 25 | trans-α-bergamotene | 21.63 | 1435 | 1432 | 0.1 ± 0.04 | 204.3511 |
| 26 | α-guaiene | 21.21 | 1439 | 1437 | 2.02 ± 0.2 | 204.3511 |
| 27 | α-humulene | 22.69 | 1455 | 1452 | 6.45 ± 0.5 | 204.3511 |
| 28 | β-farnesene | 22.78 | 1457 | 1454 | 8.25 ± 0.4 | 204.3511 |
| 29 | germacrene D | 30.98 | 1481 | 1484 | 2.31 ± 0.07 | 204.3511 |
| 30 | β-selinene | 31.04 | 1486 | 1489 | 0.6 ± 0.03 | 204.3511 |
| 31 | β-bisabolene | 32.12 | 1509 | 1512 | 0.2 ± 0.03 | 204.3511 |
| 32 | γ-cadinene | 32.78 | 1513 | 1513 | 0.2 ± 0.04 | 204.3511 |
| 33 | caryophyllene oxide | 33.86 | 1640 | 1638 | 5.68 ± 0.02 | 220.3505 |
| 34 | α-eudesmol | 33.94 | 1653 | 1652 | 0.8 ± 0.06 | 222.3663 |
| 35 | chavicol | 17.73 | 1256 | 1247 | 0.07 ± 0.01 | 134.1751 |
| 36 | eugenol | 17.92 | 1357 | 1356 | 1.5 ± 0.02 | 164.2011 |
| 37 | methyl eugenol | 19.94 | 1406 | 1402 | 0.2 ± 0.02 | 178.2277 |
| 38 | ethyl isovalerate | 8.01 | 853 | 856 | 0.2 ± 0.02 | 130.1849 |
| Classes of Constituents | RA% (No of Constituents) | |||||
| Total monoterpene hydrocarbons | 47.86 (10) | |||||
| Total oxygenated monoterpenes | 16.44 (11) | |||||
| Total sesquiterpene hydrocarbons | 22.13 (11) | |||||
| Total oxygenated sesquiterpenes | 6.48 (2) | |||||
| Total phenylpropanoids | 1.77 (3) | |||||
| Total non-terpene derivatives | 0.20 (1) | |||||
| Total identified constituents | 94.68 (38) | |||||
| Ligand | Eucalyptol | Camphor | Linalool | Nerol |
|---|---|---|---|---|
| PubChem ID | CID:2758 | CID:2537 | CID:6549 | CID:643820 |
| Binding energy | −3.76 | −4.29 | −3.37 | −5.5 |
| Ligand efficiency | −0.34 | −0.39 | −0.31 | −0.32 |
| Intermol energy | −3.76 | −4.29 | −4.58 | −6.1 |
| Ligand atoms (ring) | Alkyl hydrophobic bond:C9 Pi-alkyl hydrophobic bond:C7 | Hydrogen bonds:C2-O Pi-alkyl hydrophobic bond:C8 | Alkyl hydrophobic bond:C8, C8 , C1 ,C1 ,C3’-O ,C3’-O Pi-alkyl hydrophobic bond:C5, C3’-O Carbon–hydrogen bond interaction: C3-OH | C-1 C-1-OH C-1 C-1-OH C-1 |
| Docked amino acid residue (bond length) | Chain A: MET`163 (4.75 Å) Chain B: HIS`117 (5.21Å) | Chain A: ARG`161/ HE (2.86 Å) Chain B: HIS`117 (4.07Å) | Chain A: LEU`136 (4.62Å) Chain A: LYS`137 (4.16Å) Chain A: LEU`136 (4.48Å) Chain A: LYS`137 (4.23Å) Chain A: LYS`137/CE Chain B: VAL`266 (3.50 Å) Chain B: TYR`266 (3.50 Å) Chain B: TYR`195 (3.65Å) Chain B: TYR`195 (4.93Å) (3.18Å) | Chain A : MET Chain B : ARG Chain B : HIS |
| Ligand | Camphor | Eucalyptol |
|---|---|---|
| PubChem ID | CID_2537 | CID_2758 |
| Binding energy | −4.29 | −3.81 |
| Ligand efficiency | −0.39 | −0.35 |
| Intermol energy | −4.29 | −3.81 |
| Ligand atoms (ring) | Hydrogen bonds:C2-O Alkyl hydrophobic bond:C9, C9, O, C8 Pi-alkyl hydrophobic bond: O | Alkyl hydrophobic bond:C7, O, C9, C10, C10, O Pi-alkyl hydrophobic bond:O |
| Docked amino acid residue (bond length) | Chain A: LEU`136/CG (4.56Å) Chain A: LYS`137/CG (3.88Å) Chain A: LYS`137 (4.38Å) Chain A: LYS`137 (3.95Å) Chain B: TYR`197/ HH (2.04 Å) Chain B: TYR`195 (5.01Å) | Chain A: LYS`137 (4.27Å) Chain A: LYS`137 (4.35Å) Chain A: LEU`136 (4.64Å) Chain A: LEU`136 (4.48Å) Chain B: LYS`137 (3.94 Å) Chain B: LYS`137 (4.95 Å) Chain B: TYR`195 (5.01Å) |
| Primer Name | Forward | Reverse | Product Size |
|---|---|---|---|
| BCL-2 | TGTGGATGACTGACTACCTGAACC | CAGCCAGGAGAAATCAAACAGAGG | 186 |
| CASPASE-3 | GTGGAACTGACGATGATATGGC | CGCAAAGTGACTGGATGAACC | 212 |
| CYP-1A1 | GGCCACTTTGACCCTTACAA | CAGGTAACGGAGGACAGGAA | 236 |
| NFκB | TGAAGAGAAGACACTGACCATGGAAA | TGGATAGAGGCTAAGTGTAGACACG | 254 |
| β-Actin | AAGATCCTGACCGAGCGTGG | CAGCACTGTGTTGGCATAGAGG | 225 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khalil, H.E.; Ibrahim, H.-I.M.; Darrag, H.M.; Matsunami, K. Insight into Analysis of Essential Oil from Anisosciadium lanatum Boiss.—Chemical Composition, Molecular Docking, and Mitigation of Hepg2 Cancer Cells through Apoptotic Markers. Plants 2022, 11, 66. https://doi.org/10.3390/plants11010066
Khalil HE, Ibrahim H-IM, Darrag HM, Matsunami K. Insight into Analysis of Essential Oil from Anisosciadium lanatum Boiss.—Chemical Composition, Molecular Docking, and Mitigation of Hepg2 Cancer Cells through Apoptotic Markers. Plants. 2022; 11(1):66. https://doi.org/10.3390/plants11010066
Chicago/Turabian StyleKhalil, Hany Ezzat, Hairul-Islam Mohamed Ibrahim, Hossam M. Darrag, and Katsuyoshi Matsunami. 2022. "Insight into Analysis of Essential Oil from Anisosciadium lanatum Boiss.—Chemical Composition, Molecular Docking, and Mitigation of Hepg2 Cancer Cells through Apoptotic Markers" Plants 11, no. 1: 66. https://doi.org/10.3390/plants11010066
APA StyleKhalil, H. E., Ibrahim, H.-I. M., Darrag, H. M., & Matsunami, K. (2022). Insight into Analysis of Essential Oil from Anisosciadium lanatum Boiss.—Chemical Composition, Molecular Docking, and Mitigation of Hepg2 Cancer Cells through Apoptotic Markers. Plants, 11(1), 66. https://doi.org/10.3390/plants11010066