
Angiopteris cochinchinensis de Vriese Ameliorates LPS-Induced Acute Lung Injury via Src Inhibition

 , , and
, , and 
Abstract
:1. Introduction
2. Results
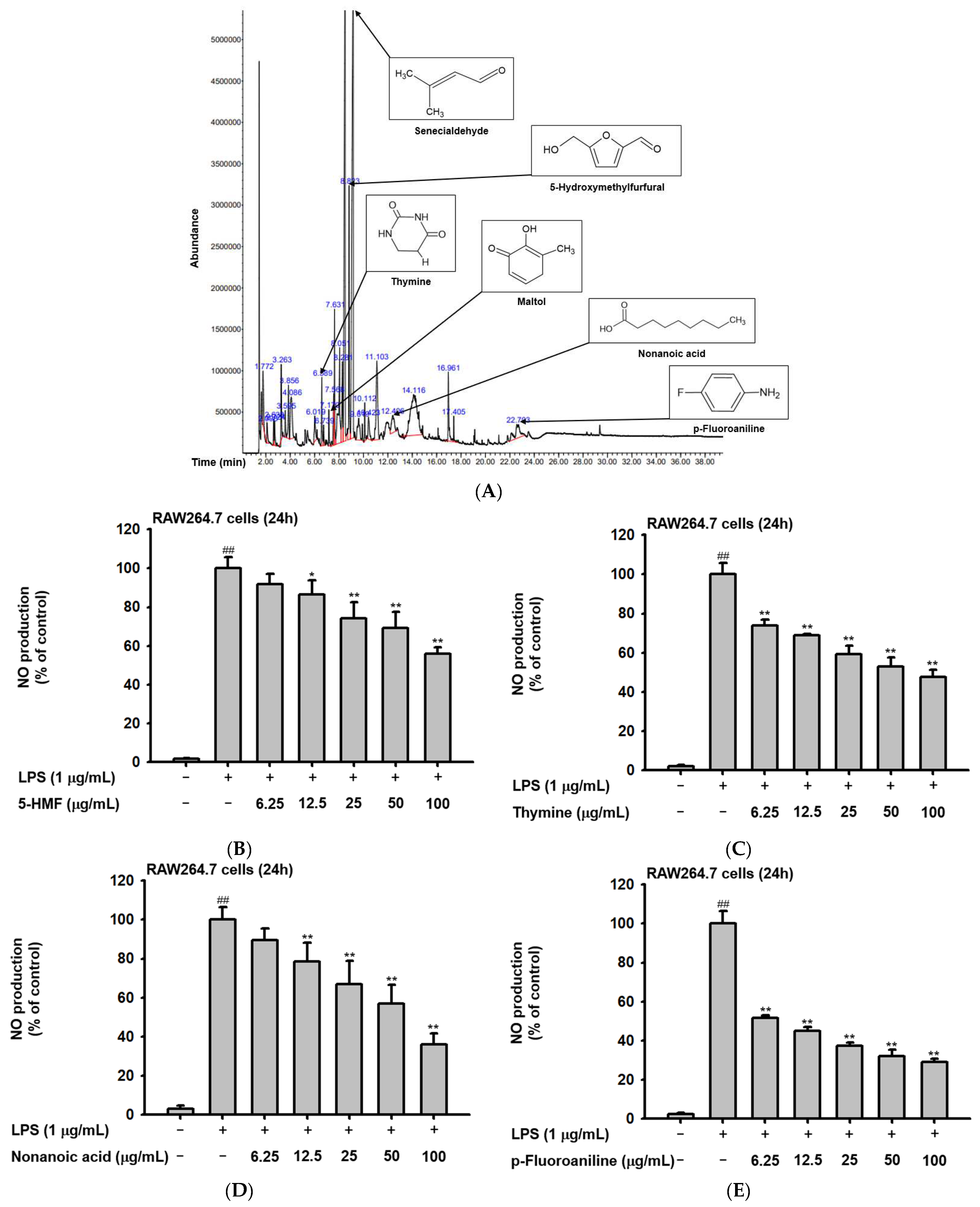
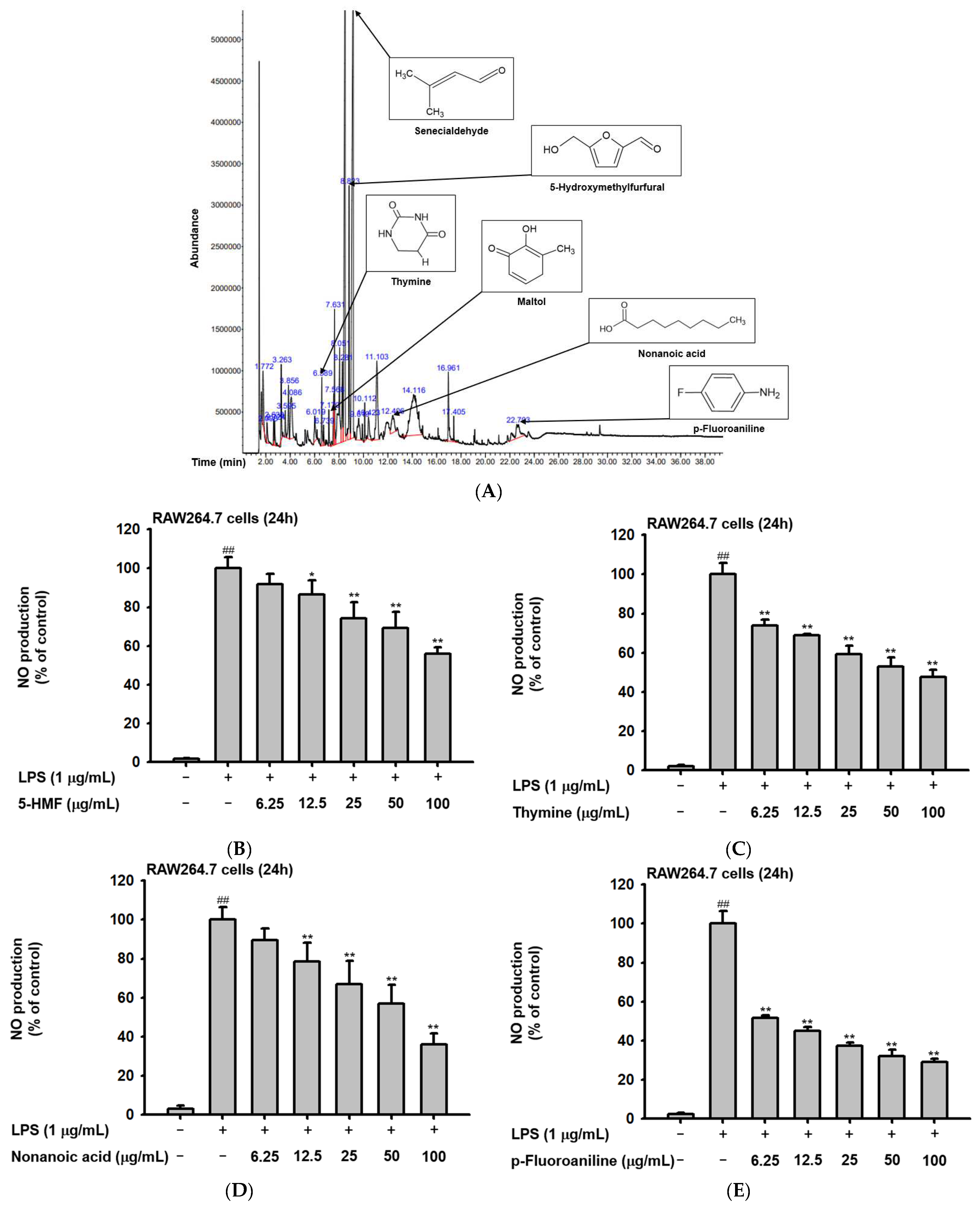
2.1. Phytochemical Components of Ac-EE
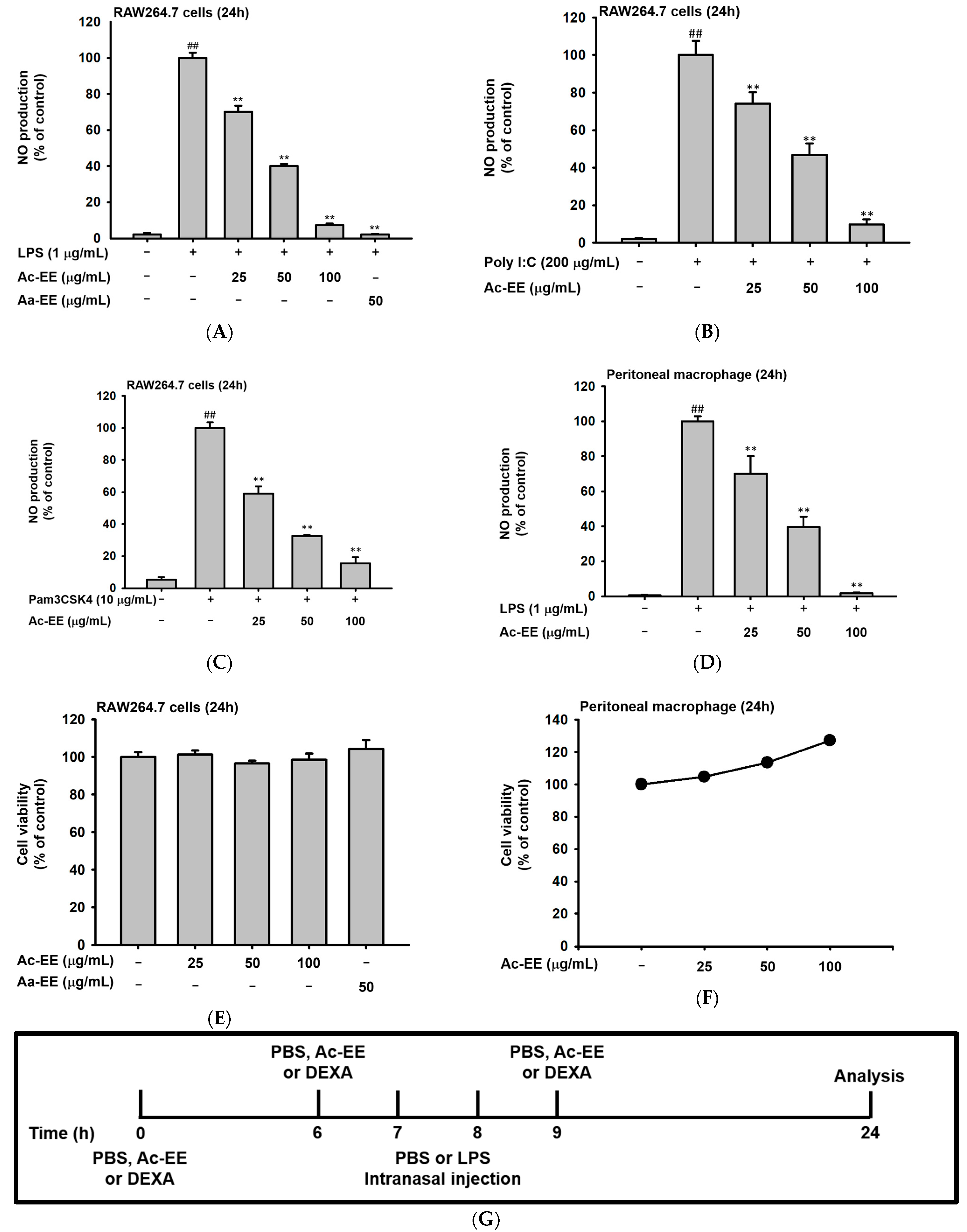
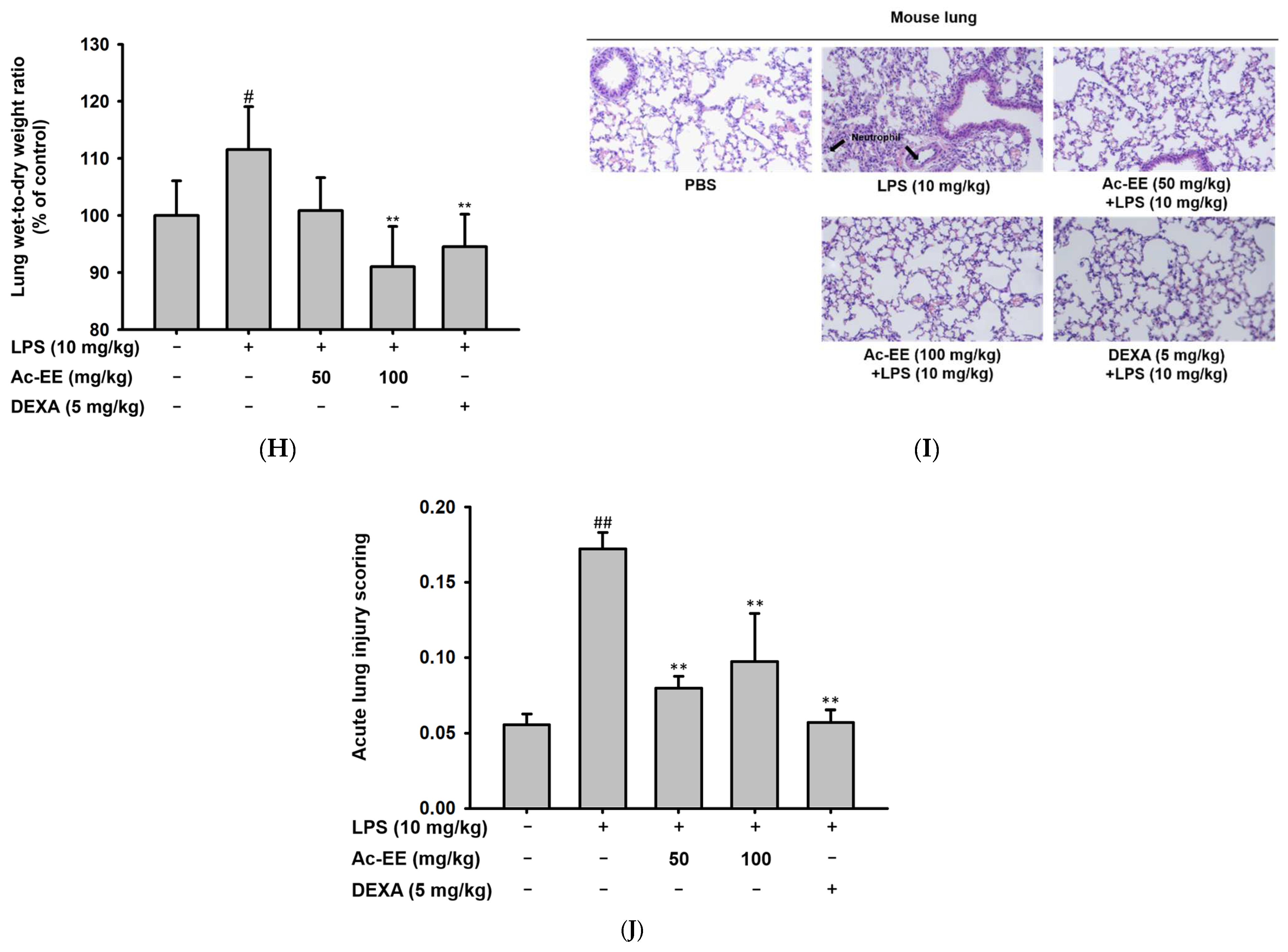
2.2. Ac-EE Alleviates Inflammation in Both Macropahge and Acute Lung Injury Mouse Model
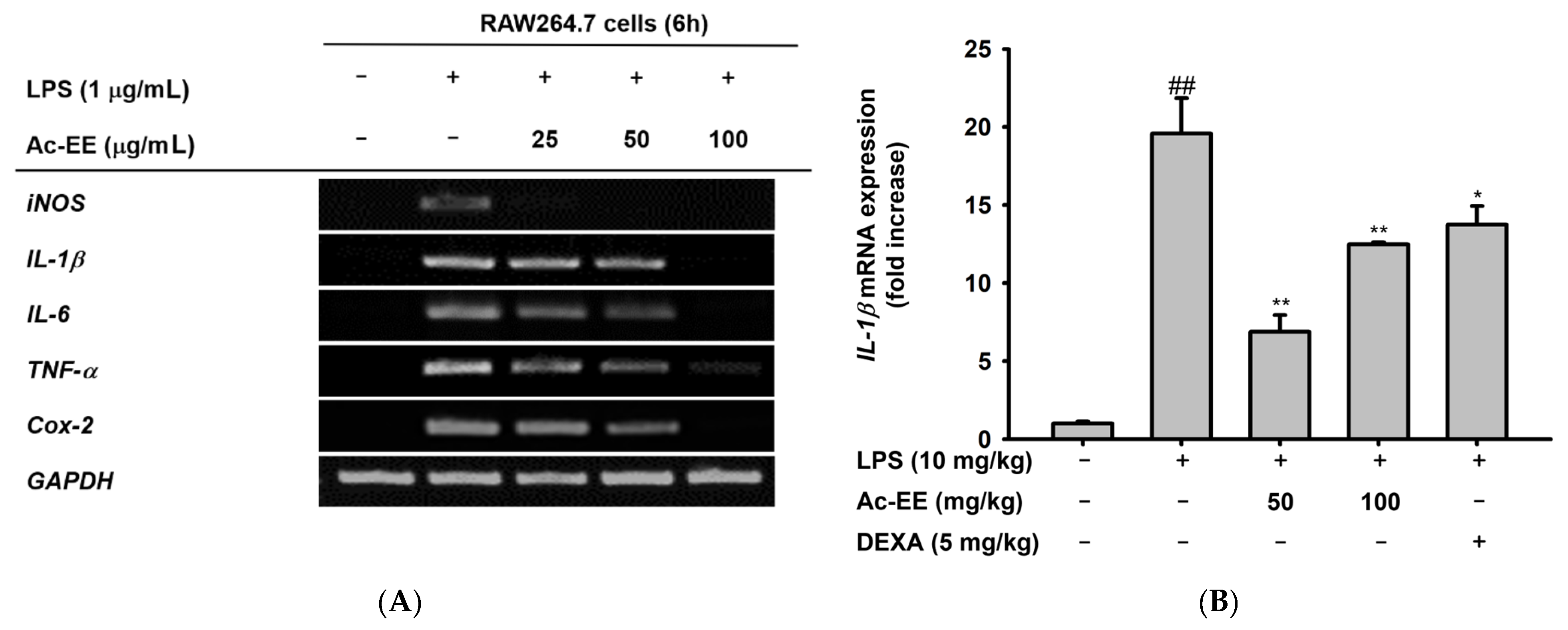
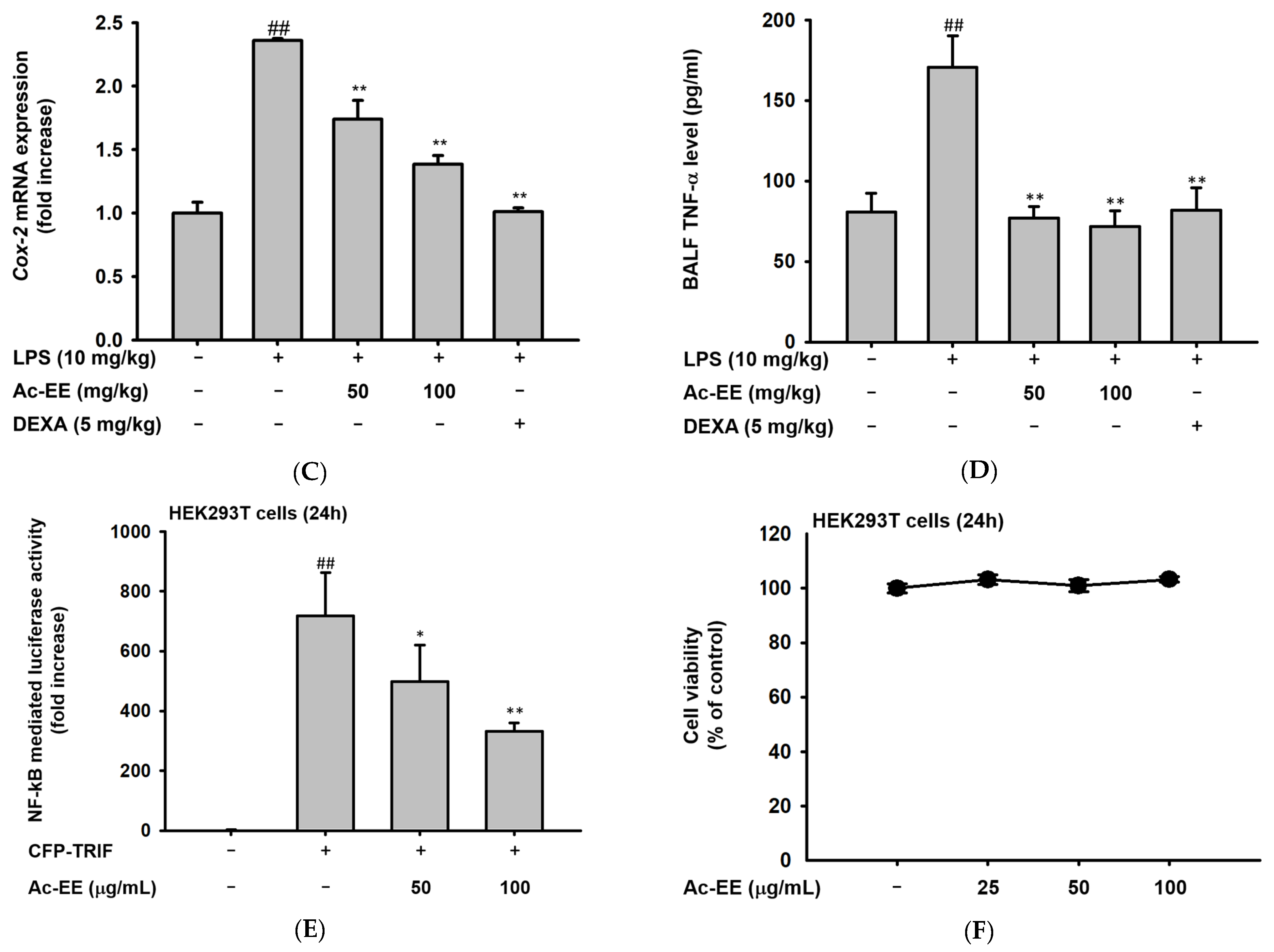
2.3. Ac-EE Reduces LPS-Induced Expression and Secretion of Proinflammatory Mediators
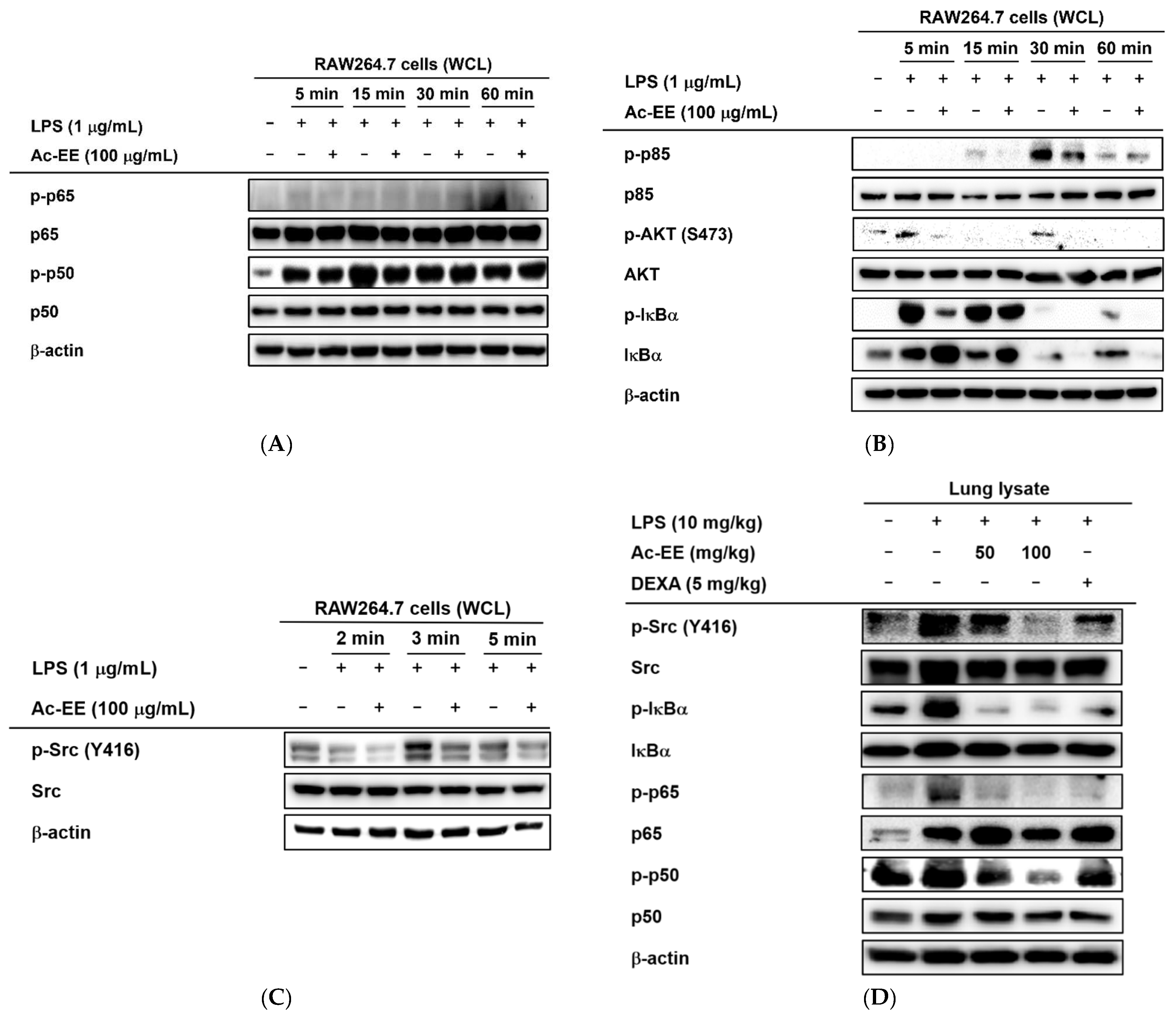
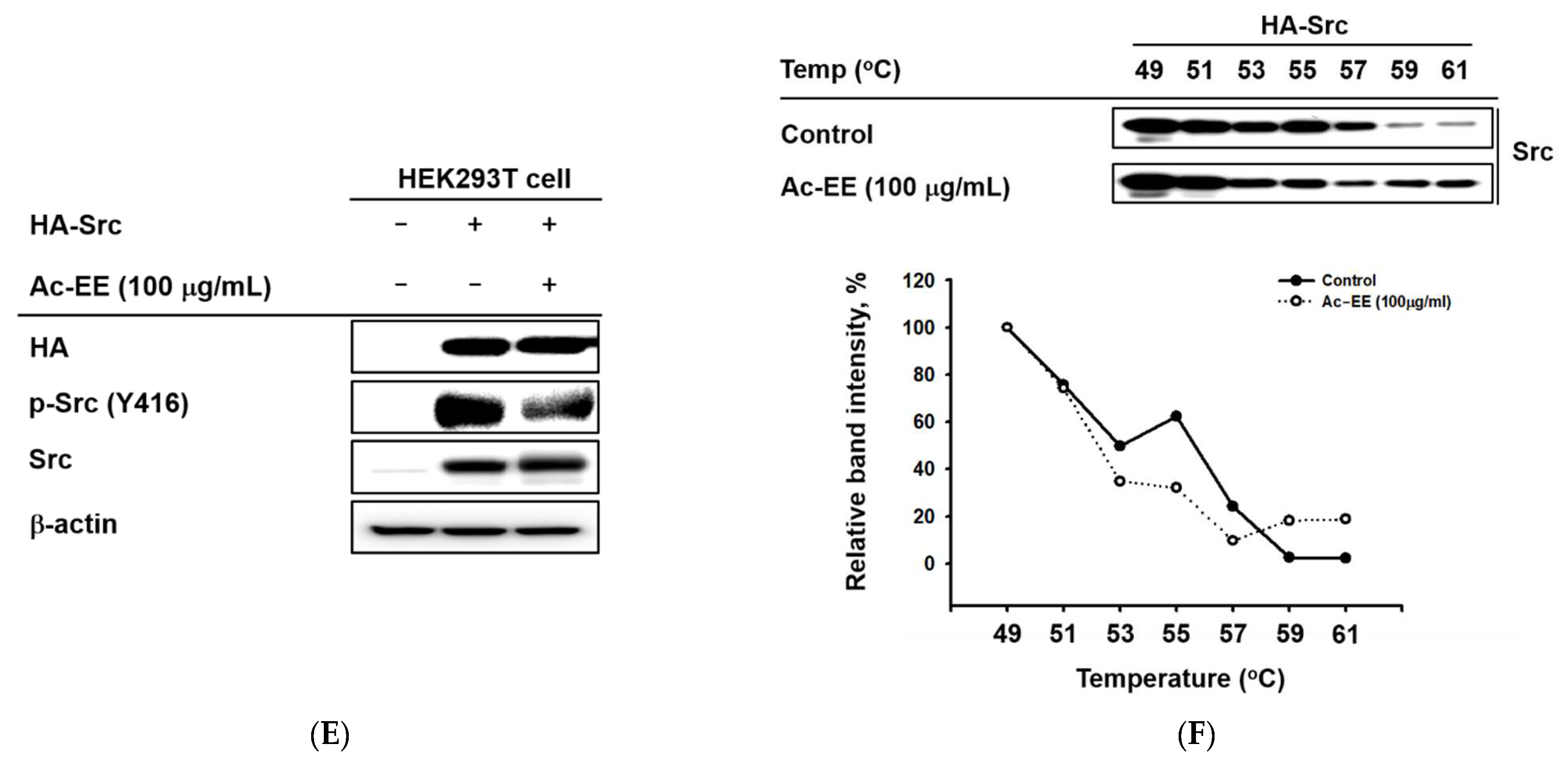
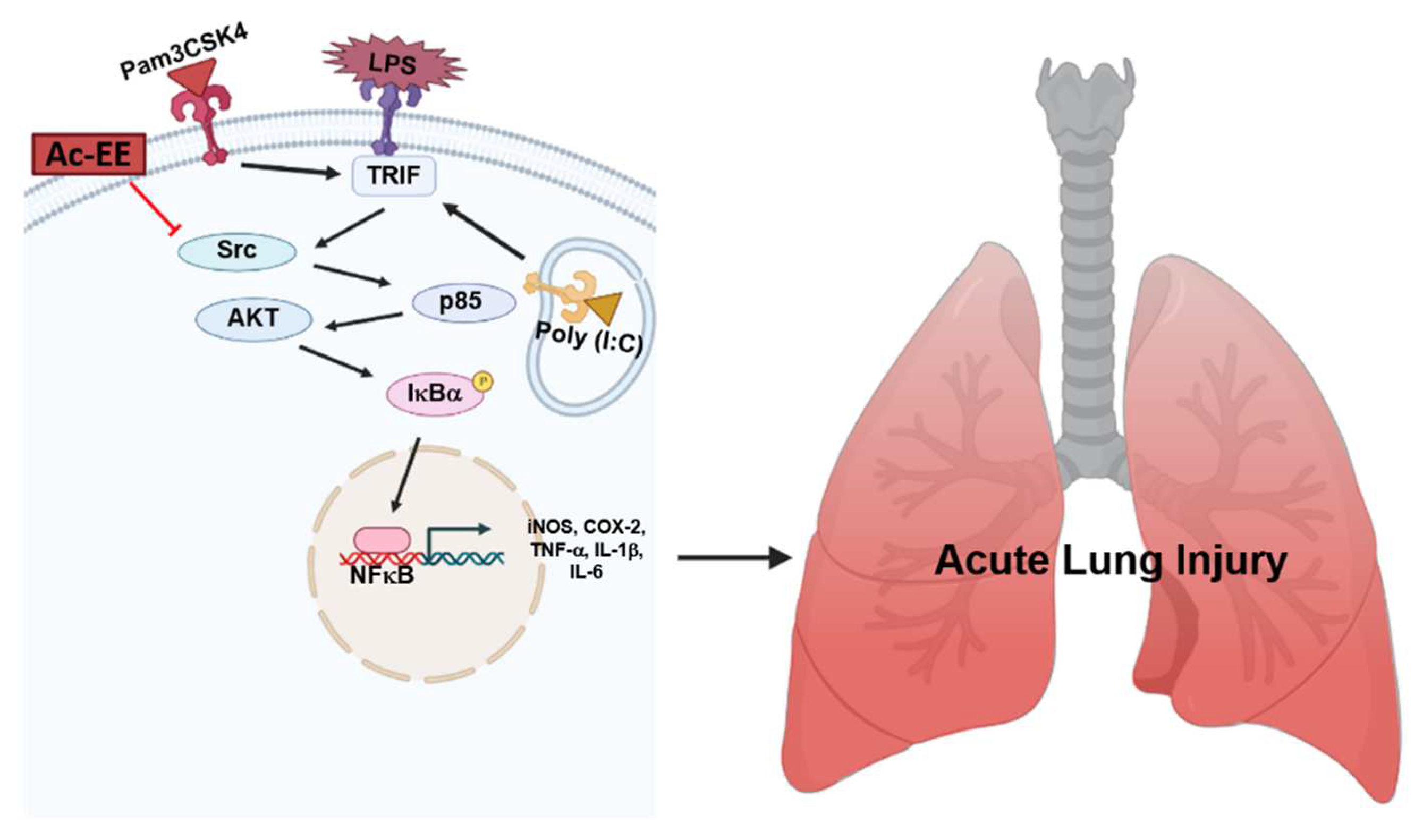
2.4. Ac-EE Inhibits NF-κB Signaling Pathway via Targeting Src
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Ac-EE Preparation and Gas Chromatography–Mass Spectrometry
4.3. Cell Culture
4.4. Isolation of Peritoneal Macrophages
4.5. In Vivo LPS-Induced Acute Lung Injury Mouse Model
4.6. Nitric Oxide Production Assay
4.7. Cell Viability Assay
4.8. Lung Wet-to-Dry Weight Ratio Measurement
4.9. Histological Analysis of Lung Tissue
4.10. mRNA Expression Level Measurement Using Semiquantitative RT-PCR and Quantitative Real-Time PCR
4.11. ELISA in Bronchoalveolar Lavage Fluid
4.12. Luciferase Reporter Assay
4.13. Whole Cell Lysate Preparation and Western Blotting Analysis
4.14. Cellular Thermal Shift Assay
4.15. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ALI | Acute lung injury |
| Ac-EE | Ethanol extract of Angiopteris cochinchinensis |
| PRR | Pattern recognition receptors |
| PAMP | Pattern-associated molecular patterns |
| LPS | Lipopolysaccharide |
| TLR | Toll-like receptors |
| NF-κB | Nuclear factor kappa B |
| iNOS | Induced nitric oxide synthase |
| COX-2 | Cyclooxygenase-2 |
| IL-1β | Interleukin-1 beta |
| IL-6 | Interleukin-6 |
| TNF-α | Tumor necrosis factor alpha |
| PI3K | Phosphoinositide 3-kinase |
| GC-MS | Gas chromatography–mass spectrometry |
| CETSA | Cellular thermal shift assay |
References
- Butt, Y.; Kurdowska, A.; Allen, T.C. Acute lung injury: A clinical and molecular review. Arch. Pathol. Lab. Med. 2016, 140, 345–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, E.; Matthay, M.A.; Dinarello, C.A.; Vincent, J.L.; Cohen, J.; Opal, S.M.; Glauser, M.; Parsons, P.; Fisher, C.J., Jr.; Repine, J.E. Consensus conference definitions for sepsis, septic shock, acute lung injury, and acute respiratory distress syndrome: Time for a reevaluation. Crit. Care Med. 2000, 28, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Tirunavalli, S.K.; Gourishetti, K.; Kotipalli, R.S.S.; Kuncha, M.; Kathirvel, M.; Kaur, R.; Jerald, M.K.; Sistla, R.; Andugulapati, S.B. Dehydrozingerone ameliorates lipopolysaccharide induced acute respiratory distress syndrome by inhibiting cytokine storm, oxidative stress via modulating the MAPK/NF-κB pathway. Phytomedicine 2021, 92, 153729. [Google Scholar] [CrossRef]
- Cole, P. The damaging role of bacteria in chronic lung infection. J. Antimicrob. Chemother. 1997, 40 (Suppl. A), 5–10. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, A.; Gonçalves, J.S.; Dourado, Á.W.A.; de Sousa, E.M.; Brito, N.M.; Silva, L.K.; Batista, M.C.A.; de Sá, J.C.; Monteiro, C.; Fernandes, E.S.; et al. Punica granatum L. leaf extract attenuates lung inflammation in mice with acute lung injury. J. Immunol. Res. 2018, 2018, 6879183. [Google Scholar] [CrossRef] [Green Version]
- Cochi, S.E.; Kempker, J.A.; Annangi, S.; Kramer, M.R.; Martin, G.S. Mortality trends of acute respiratory distress syndrome in the United States from 1999 to 2013. Ann. Am. Thorac. Soc. 2016, 13, 1742–1751. [Google Scholar] [CrossRef] [Green Version]
- Standiford, T.J.; Ward, P.A. Therapeutic targeting of acute lung injury and acute respiratory distress syndrome. Transl. Res. 2016, 167, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Mogensen, T.H. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin. Microbiol. Rev. 2009, 22, 240–273. [Google Scholar] [CrossRef] [Green Version]
- Davicino, R.C.; Eliçabe, R.J.; di Genaro, M.S.; Rabinovich, G.A. Coupling pathogen recognition to innate immunity through glycan-dependent mechanisms. Int. Immunopharmacol. 2011, 11, 1457–1463. [Google Scholar] [CrossRef]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef]
- Funderburg, N.T.; Jadlowsky, J.K.; Lederman, M.M.; Feng, Z.; Weinberg, A.; Sieg, S.F. The toll-like receptor 1/2 agonists pam(3) CSK(4) and human β-defensin-3 differentially induce interleukin-10 and nuclear factor-κB signalling patterns in human monocytes. Immunology 2011, 134, 151–160. [Google Scholar] [CrossRef]
- Field, R.; Campion, S.; Warren, C.; Murray, C.; Cunningham, C. Systemic challenge with the TLR3 agonist poly I:C induces amplified IFNα/β and IL-1β responses in the diseased brain and exacerbates chronic neurodegeneration. Brain Behav. Immun. 2010, 24, 996–1007. [Google Scholar] [CrossRef] [Green Version]
- Fujitani, S.; Sun, H.Y.; Yu, V.L.; Weingarten, J.A. Pneumonia due to Pseudomonas aeruginosa: Part I: Epidemiology, clinical diagnosis, and source. Chest 2011, 139, 909–919. [Google Scholar] [CrossRef]
- Pier, G.B. Pseudomonas aeruginosa lipopolysaccharide: A major virulence factor, initiator of inflammation and target for effective immunity. Int. J. Med. Microbiol. IJMM 2007, 297, 277–295. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, Y.; Murakami, H.; Akbar, S.M.; Matsui, H.; Onji, M. A novel and effective approach of developing aggressive experimental autoimmune gastritis in neonatal thymectomized BALB/c mouse by polyinosinic:polycytidylic acid. Clin. Exp. Immunol. 2004, 136, 423–431. [Google Scholar] [CrossRef]
- Kim, H.S.; Go, H.; Akira, S.; Chung, D.H. TLR2-mediated production of IL-27 and chemokines by respiratory epithelial cells promotes bleomycin-induced pulmonary fibrosis in mice. J. Immun. Balt. 2011, 187, 4007–4017. [Google Scholar] [CrossRef] [Green Version]
- Hebert-Chatelain, E. Src kinases are important regulators of mitochondrial functions. Int. J. Biochem. Cell Biol. 2013, 45, 90–98. [Google Scholar] [CrossRef]
- Avizienyte, E.; Frame, M.C. Src and FAK signalling controls adhesion fate and the epithelial-to-mesenchymal transition. Curr. Opin. Cell Biol. 2005, 17, 542–547. [Google Scholar] [CrossRef]
- Sánchez-Bailón, M.P.; Calcabrini, A.; Gómez-Domínguez, D.; Morte, B.; Martín-Forero, E.; Gómez-López, G.; Molinari, A.; Wagner, K.U.; Martín-Pérez, J. Src kinases catalytic activity regulates proliferation, migration and invasiveness of MDA-MB-231 breast cancer cells. Cell. Signal. 2012, 24, 1276–1286. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.Y.; Kim, H.G.; Park, S.H.; Jang, S.G.; Park, K.J.; Kim, D.S.; Kim, J.H.; Cho, J.Y. Anti-inflammatory functions of alverine via targeting Src in the NF-κB pathway. Biomolecules 2020, 10, 611. [Google Scholar] [CrossRef] [Green Version]
- Shin, K.K.; Park, J.G.; Hong, Y.H.; Aziz, N.; Park, S.H.; Kim, S.; Kim, E.; Cho, J.Y. Anti-inflammatory effects of Licania macrocarpa Cuatrec methanol extract target Src- and TAK1-mediated pathways. Evid. Based Complement. Altern. Med. 2019, 2019, 4873870. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Park, J.G.; Hong, Y.H.; Shin, K.K.; Kim, J.K.; Kim, Y.D.; Yoon, K.D.; Kim, K.H.; Yoo, B.C.; Sung, G.H.; et al. Sauropus brevipes ethanol extract negatively regulates inflammatory responses in vivo and in vitro by targeting Src, Syk and IRAK1. Pharm. Biol. 2021, 59, 74–86. [Google Scholar] [CrossRef]
- Viatour, P.; Merville, M.P.; Bours, V.; Chariot, A. Phosphorylation of NF-κB and IκB proteins: Implications in cancer and inflammation. Trends Biochem. Sci. 2005, 30, 43–52. [Google Scholar] [CrossRef]
- Won, J.H.; Im, H.T.; Kim, Y.H.; Yun, K.J.; Park, H.J.; Choi, J.W.; Lee, K.T. Anti-inflammatory effect of buddlejasaponin IV through the inhibition of iNOS and COX-2 expression in RAW 264.7 macrophages via the NF-κB inactivation. Br. J. Pharmacol. 2006, 148, 216–225. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.S.; Yang, E.J.; Shin, D.H.; Son, K.H.; Park, H.Y.; Lee, J.S. Effect of arazyme on the lipopolysaccharide-induced inflammatory response in human endothelial cells. Mol. Med. Rep. 2014, 10, 1025–1029. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.R.; Omoloso, A.D. Antibacterial and antifungal activities of Angiopteris evecta. Fitoterapia 2008, 79, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, R.; Pandeya, P.R.; Lee, K.H.; Kim, S.G.; Devkota, H.P.; Jung, H.J. Anti-adipogenic and anti-inflammatory activities of (-)-epi-Osmundalactone and angiopteroside from Angiopteris helferiana C. Presl. Molecules 2020, 25, 1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamitakahara, H.; Okayama, T.; Praptiwi, A.A.; Tobimatsu, Y.; Takano, T. Two-dimensional NMR analysis of Angiopteris evecta rhizome and improved extraction method for angiopteroside. Phytochem. Anal. PCA 2019, 30, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Kong, F.; Lee, B.H.; Wei, K. 5-hydroxymethylfurfural mitigates lipopolysaccharide-stimulated inflammation via suppression of MAPK, NF-κB and mTOR activation in RAW 264.7 cells. Molecules 2019, 24, 275. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Hao, W.; Hu, J.; Mi, X.; Han, Y.; Ren, S.; Jiang, S.; Wang, Y.; Li, X.; Li, W. Maltol improves APAP-induced hepatotoxicity by inhibiting oxidative stress and inflammation response via NF-κB and PI3K/akt signal pathways. Antioxidants 2019, 8, 395. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Jiang, Z.; Shen, C.; Zou, H.; Zhang, Z.; Wang, K.; Bai, R.; Kang, Y.; Ye, X.Y.; Xie, T. 5-hydroxymethylfurfural alleviates inflammatory lung injury by inhibiting endoplasmic reticulum stress and NLRP3 inflammasome activation. Front. Cell Dev. Biol. 2021, 9, 782427. [Google Scholar] [CrossRef]
- Jeong, D.; Yi, Y.S.; Sung, G.H.; Yang, W.S.; Park, J.G.; Yoon, K.; Yoon, D.H.; Song, C.; Lee, Y.; Rhee, M.H.; et al. Anti-inflammatory activities and mechanisms of Artemisia asiatica ethanol extract. J. Ethnopharmacol. 2014, 152, 487–496. [Google Scholar] [CrossRef]
- Al-Harbi, N.O.; Imam, F.; Al-Harbi, M.M.; Ansari, M.A.; Zoheir, K.M.; Korashy, H.M.; Sayed-Ahmed, M.M.; Attia, S.M.; Shabanah, O.A.; Ahmad, S.F. Dexamethasone attenuates LPS-induced acute lung injury through inhibition of NF-κB, COX-2, and pro-inflammatory mediators. Immunol. Investig. 2016, 45, 349–369. [Google Scholar] [CrossRef]
- Martinez Molina, D.; Nordlund, P. The cellular thermal shift assay: A novel biophysical assay for in situ drug target engagement and mechanistic biomarker studies. Annu. Rev. Pharmacol. Toxicol. 2016, 56, 141–161. [Google Scholar] [CrossRef]
- Wang, C.; Wang, X.; Long, X.; Xia, D.; Ben, D.; Wang, Y. Publication trends of research on acute lung injury and acute respiration distress syndrome during 2009–2019: A 10-year bibliometric analysis. Am. J. Transl. Res. 2020, 12, 6366–6380. [Google Scholar]
- Rubenfeld, G.D.; Caldwell, E.; Peabody, E.; Weaver, J.; Martin, D.P.; Neff, M.; Stern, E.J.; Hudson, L.D. Incidence and outcomes of acute lung injury. N. Engl. J. Med. 2005, 353, 1685–1693. [Google Scholar] [CrossRef] [Green Version]
- Mokrá, D. Acute lung injury—From pathophysiology to treatment. Physiol. Res. 2020, 69, S353–S366. [Google Scholar] [CrossRef]
- Marini, J.J. Limitations of clinical trials in acute lung injury and acute respiratory distress syndrome. Curr. Opin. Crit. Care 2006, 12, 25–31. [Google Scholar] [CrossRef]
- Menezes, S.L.; Bozza, P.T.; Neto, H.C.; Laranjeira, A.P.; Negri, E.M.; Capelozzi, V.L.; Zin, W.A.; Rocco, P.R. Pulmonary and extrapulmonary acute lung injury: Inflammatory and ultrastructural analyses. J. Appl. Physiol. 2005, 98, 1777–1783. [Google Scholar] [CrossRef] [Green Version]
- Phumthum, M.; Balslev, H. Anti-infectious plants of the Thai Karen: A meta-analysis. Antibiotics 2020, 9, 298. [Google Scholar] [CrossRef]
- Zhu, D.C.; Wang, Y.H.; Lin, J.H.; Miao, Z.M.; Xu, J.J.; Wu, Y.S. Maltol inhibits the progression of osteoarthritis via the nuclear factor-erythroid 2-related factor-2/heme oxygenase-1 signal pathway in vitro and in vivo. Food Funct. 2021, 12, 1327–1337. [Google Scholar] [CrossRef] [PubMed]
- Hendrickson, C.M.; Matthay, M.A. Viral pathogens and acute lung injury: Investigations inspired by the SARS epidemic and the 2009 H1N1 influenza pandemic. Semin. Respir. Crit. Care Med. 2013, 34, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yan, F.; Zhou, H.; Lin, X.; Wu, Y.; Chen, C.; Zhou, N.; Chen, Z.; Li, J.D.; Shen, H. P. aeruginosa lipopolysaccharide-induced MUC5AC and CLCA3 expression is partly through Duox1 in vitro and in vivo. PLoS ONE 2013, 8, e63945. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, S.; Zawieja, S.D.; Wang, W.; Lee, Y.; Wang, Y.J.; von der Weid, P.Y.; Zawieja, D.C.; Muthuchamy, M. Lipopolysaccharide modulates neutrophil recruitment and macrophage polarization on lymphatic vessels and impairs lymphatic function in rat mesentery. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H2042–H2057. [Google Scholar] [CrossRef] [Green Version]
- Jiang, K.; Zhang, T.; Yin, N.; Ma, X.; Zhao, G.; Wu, H.; Qiu, C.; Deng, G. Geraniol alleviates LPS-induced acute lung injury in mice via inhibiting inflammation and apoptosis. Oncotarget 2017, 8, 71038–71053. [Google Scholar] [CrossRef] [Green Version]
- Cheung, O.-Y.; Graziano, P.; Leslie, K.O. 5—Acute lung injury. In Practical Pulmonary Pathology: A Diagnostic Approach, 2nd ed.; Leslie, K.O., Wick, M.R., Eds.; W.B. Saunders: Philadelphia, PA, USA, 2011; pp. 117–136. [Google Scholar]
- Lin, W.C.; Chen, C.W.; Huang, Y.W.; Chao, L.; Chao, J.; Lin, Y.S.; Lin, C.F. Kallistatin protects against sepsis-related acute lung injury via inhibiting inflammation and apoptosis. Sci. Rep. 2015, 5, 12463. [Google Scholar] [CrossRef] [Green Version]
- Bordag, N.; Klie, S.; Jürchott, K.; Vierheller, J.; Schiewe, H.; Albrecht, V.; Tonn, J.C.; Schwartz, C.; Schichor, C.; Selbig, J. Glucocorticoid (dexamethasone)-induced metabolome changes in healthy males suggest prediction of response and side effects. Sci. Rep. 2015, 5, 15954. [Google Scholar] [CrossRef]
- Cea, L.A.; Balboa, E.; Puebla, C.; Vargas, A.A.; Cisterna, B.A.; Escamilla, R.; Regueira, T.; Sáez, J.C. Dexamethasone-induced muscular atrophy is mediated by functional expression of connexin-based hemichannels. Biochim. Biophys. Acta 2016, 1862, 1891–1899. [Google Scholar] [CrossRef]
- Bindu, S.; Mazumder, S.; Bandyopadhyay, U. Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: A current perspective. Biochem. Pharmacol. 2020, 180, 114147. [Google Scholar] [CrossRef]
- Bhandari, K.; Venables, B. Ibuprofen bioconcentration and prostaglandin E2 levels in the bluntnose minnow Pimephales notatus. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2011, 153, 251–257. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, M.; Xiong, R.P.; Chen, X.Y.; Li, P.; Ning, Y.L.; Yang, N.; Peng, Y.; Zhou, Y.G. Somatostatin reduces the acute lung injury of mice via increasing the affinity of glucocorticoid receptor. Cell. Physiol. Biochem. 2016, 38, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Van Hoecke, L.; Job, E.R.; Saelens, X.; Roose, K. Bronchoalveolar lavage of murine lungs to analyze inflammatory cell infiltration. J. Vis. Exp. 2017, 123, e55398. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.H.; Kim, J.H.; Cho, J.Y. Ranunculus bulumei methanol extract exerts anti-inflammatory activity by targeting Src/Syk in NF-κB signaling. Biomolecules 2020, 10, 546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Shin, K.K.; Kim, H.G.; Jo, M.; Kim, J.K.; Lee, J.S.; Choung, E.S.; Li, W.Y.; Lee, S.W.; Kim, K.H.; et al. Src/NF-κB-targeted anti-inflammatory effects of Potentilla glabra var. Mandshurica (Maxim.) hand.-mazz. ethanol extract. Biomolecules 2020, 10, 648. [Google Scholar] [CrossRef]
- Song, C.; Kim, M.Y.; Cho, J.Y. Olea europaea suppresses inflammation by targeting TAK1-mediated MAP kinase activation. Molecules 2021, 26, 1540. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | RT | Name of the Compound | Corrected Area | % of Total |
|---|---|---|---|---|
| 1 | 1.772 | Acetic acid | 29216024 | 1.64 |
| 2 | 2.098 | 2-Propanone, 1-hydoxy- | 12843838 | 0.72 |
| 3 | 2.638 | 2-Propenoyl chloride | 8113271 | 0.46 |
| 4 | 2.724 | 1,2,3-Propanetriol, 1-acetate | 11290321 | 0.63 |
| 5 | 3.263 | Glyceraldehyde | 34415997 | 1.93 |
| 6 | 3.595 | 2-Furanmethanol | 10248331 | 0.57 |
| 7 | 3.856 | Dihydro-2(3H)-thiophenone | 28723699 | 1.61 |
| 8 | 4.086 | Dihydroxyacetone | 52160462 | 2.93 |
| 9 | 6.019 | 1,4-Cyclohex-2-enedione | 18462433 | 1.04 |
| 10 | 6.589 | Thymine | 25694457 | 1.44 |
| 11 | 6.739 | 3-Furanacarboxylic acid, methyl ester | 8426843 | 0.47 |
| 12 | 7.170 | Maltol | 11986818 | 0.67 |
| 13 | 7.568 | Isothiazole, 3-methyl- | 26854159 | 1.51 |
| 14 | 7.631 | 4H-Pyran-4one, 2,3-dihydro-3,5-dihydroxy-6-methyl- | 44906782 | 2.52 |
| 15 | 8.051 | Cyclopentanone ethylene ketal | 92735782 | 5.20 |
| 16 | 8.281 | Pyrrolidin-1-acetic acid | 44123254 | 2.47 |
| 17 | 8.492 | Divinyl sulfide | 330210094 | 18.52 |
| 18 | 8.823 | 5-Hydroxymethylfurfural | 85088443 | 4.77 |
| 19 | 9.156 | 2-Butenal, 3-methyl- | 435954466 | 24.45 |
| 20 | 9.619 | 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl- | 32698466 | 1.83 |
| 21 | 10.112 | Isosorbide | 13278795 | 0.74 |
| 22 | 10.423 | Heptane, 2,3-epoxy- | 19897179 | 1.12 |
| 23 | 11.103 | Formic acid, hex-2-yl ester | 86990006 | 4.88 |
| 24 | 12.406 | Nonanoic acid | 33776334 | 1.89 |
| 25 | 14.116 | N-Methoxymethyl-N-methylacetamide | 190707640 | 10.70 |
| 26 | 16.961 | 2-Amino-actadecane-1,3,4 triol 1,3:2,4-bis-methaneboronate | 39099092 | 2.19 |
| 27 | 17.405 | n-Hexadecanoic acid | 7408037 | 0.42 |
| 28 | 22.703 | p-Fluoroaniline | 47763799 | 2.68 |
| Measurement Criteria | Score | ||
|---|---|---|---|
| 0 | 1 | 2 | |
| A. Neutrophil infiltration to the interstitial space | Not found | 1 to 5 | More than 5 |
| B. Neutrophils infiltration to the alveolar space | Not found | 1 to 5 | More than 5 |
| C. Numbers of hyaline membrane | Not found | 3 | More than 3 |
| D. Septal thickening of alveolar wall | More than 2× | 2 to 4× | More than 4× |
| Score = [(20 × A) + (14 × B) + (7 × C) + (2 × D)]/(field number × 100) | |||
| Gene Name | Sequence (5′–3′) | |
|---|---|---|
| iNOS | Forward | TGCCAGGGTCACAACTTTACA |
| Reverse | ACCCCAAGCAAGACTTGGAC | |
| COX-2 | Forward | TGAGTACCGCAACGCTTCT |
| Reverse | TGGGAGGCACTTGCATTGAT | |
| TNF-α | Forward | TTGACCTCAGCGCTGAGTTG |
| Reverse | CCTGTAGCCCACGTCGTAGC | |
| IL-1β | Forward | CAGGATGAGGACATGAGCACC |
| Reverse | CTCTGCAGACTCAAACTCCAC | |
| IL-6 | Forward | GGAAATCGTGGAAATGAG |
| Reverse | GCTTAGGCATAACGCACT | |
| GAPDH | Forward | GAAGGTCGGTGTGAACGGAT |
| Reverse | AGTGATGGCATGGACTGTGG | |
| Gene Name | Sequence (5′–3′) | |
|---|---|---|
| COX-2 | Forward | TTGGAGGCGAAGTGGGTTTT |
| Reverse | TGGCTGTTTTGGTAGGCTGT | |
| IL-1β | Forward | GTGAAATGCCACCTTTTACAGTG |
| Reverse | CCTGCCTGAAGCTCTTGTTG | |
| GAPDH | Forward | GGAGAGTGTTTCCTCGTCCC |
| Reverse | ATGAAGGGGTCGTTGATGGC | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, W.Y.; Lee, H.P.; Kim, S.A.; Huang, L.; Yoon, J.H.; Shin, C.Y.; Mitra, A.; Kim, H.G.; Cho, J.Y. Angiopteris cochinchinensis de Vriese Ameliorates LPS-Induced Acute Lung Injury via Src Inhibition. Plants 2022, 11, 1306. https://doi.org/10.3390/plants11101306
Jang WY, Lee HP, Kim SA, Huang L, Yoon JH, Shin CY, Mitra A, Kim HG, Cho JY. Angiopteris cochinchinensis de Vriese Ameliorates LPS-Induced Acute Lung Injury via Src Inhibition. Plants. 2022; 11(10):1306. https://doi.org/10.3390/plants11101306
Chicago/Turabian StyleJang, Won Young, Hwa Pyoung Lee, Seung A Kim, Lei Huang, Ji Hye Yoon, Chae Yun Shin, Ankita Mitra, Han Gyung Kim, and Jae Youl Cho. 2022. "Angiopteris cochinchinensis de Vriese Ameliorates LPS-Induced Acute Lung Injury via Src Inhibition" Plants 11, no. 10: 1306. https://doi.org/10.3390/plants11101306
APA StyleJang, W. Y., Lee, H. P., Kim, S. A., Huang, L., Yoon, J. H., Shin, C. Y., Mitra, A., Kim, H. G., & Cho, J. Y. (2022). Angiopteris cochinchinensis de Vriese Ameliorates LPS-Induced Acute Lung Injury via Src Inhibition. Plants, 11(10), 1306. https://doi.org/10.3390/plants11101306