
Virus and Virus-like Pathogens in the Grapevine Virus Collection of Croatian Autochthonous Grapevine Cultivars

 ,
,  ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Virus Screening by ELISA and Collection Establishment
2.2. Grapevine Rupestris Stem Pitting-Associated Virus (GRSPaV) Screening by Western Blot and Other Molecular Methods, including Phylogenetic Analysis
2.3. High-Throughput Sequencing (HTS)
2.4. Loop-Mediated Isothermal Amplification (LAMP) Screening for “Flavescence Dorée”
3. Discussion
4. Materials and Methods
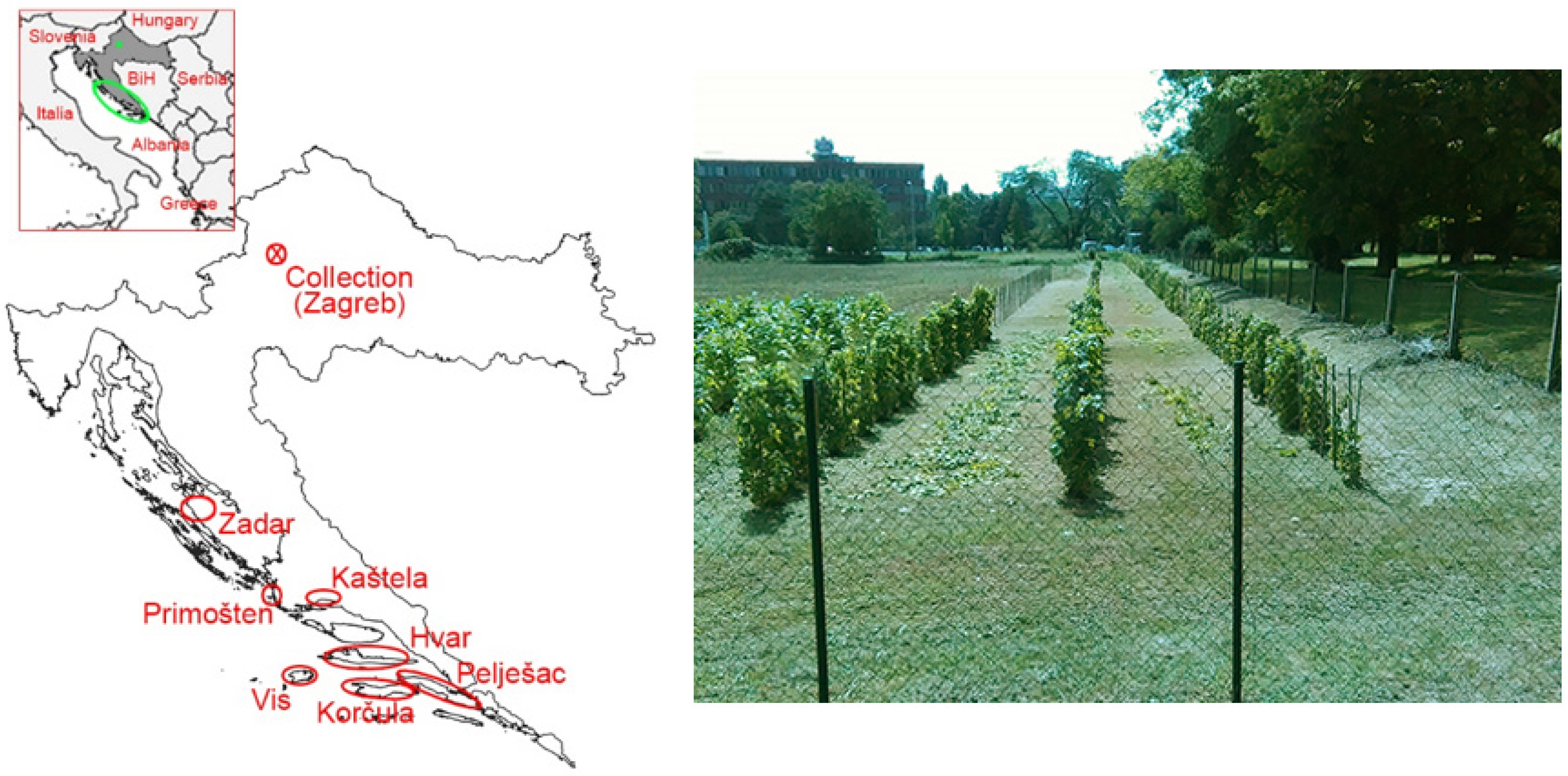
4.1. ELISA Screening and Plant Material Used for the Grapevine Virus Collection
4.2. Grapevine Rupestris Stem Pitting-Associated Virus (GRSPaV) Screening by Western Blot and Other Molecular Methods, including Phylogenetic Analysis
4.3. High-Throughput Sequencing (HTS)
4.4. Loop-Mediated Isothermal Amplification (LAMP) Screening for “Flavescence Dorée”
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fuchs, M. Grapevine viruses: A multitude of diverse species with simple but overall poorly adopted management solutions in the vineyard. J. Plant. Pathol. 2020, 102, 643–653. [Google Scholar] [CrossRef]
- Golino, D.A. The Davis grapevine virus collection. Am. J. Enol. Vitic. 1992, 43, 200–205. [Google Scholar]
- Vončina, D.; Šimon, S.; Đermić, E.; Cvjetković, B.; Pejić, I.; Maletić, E.; Karoglan Kontić, J. Distribution and partial molecular characterization of Grapevine leafroll-associated virus 2 (GLRaV-2) found in Croatian autochthonous grapevine (Vitis vinifera L.) germplasm. J. Plant Dis. Prot. 2010, 5, 194–200. [Google Scholar] [CrossRef]
- Vončina, D.; Šimon, S.; Đermić, E.; Cvjetković, B.; Pejić, I.; Maletić, E.; Karoglan Kontić, J. Differential properties of Grapevine virus B isolates from Croatian autochthonous grapevine cultivars. J. Plant Path. 2011, 93, 283–289. [Google Scholar]
- Vončina, D.; Al Rwahnih, M.; Rowhani, A.; Gouran, M.; Almeida, R.P.P. Viral Diversity in Autochthonous Croatian Grapevine Cultivars. Plant Dis. 2017, 7, 1230–1235. [Google Scholar] [CrossRef] [Green Version]
- Vončina, D.; Almeida, R.P.P. Screening of some Croatian autochthonous grapevine varieties reveals a multitude of viruses, including novel ones. Arch. Virol. 2018, 163, 2239–2243. [Google Scholar] [CrossRef] [Green Version]
- Debat, H.; Zavallo, D.; Brisbane, R.S.; Vončina, D.; Almeida, R.P.P.; Blouin, A.G.; Al Rwahnih, M.; Gomez-Talquenca, S.; Asurmendi, S. Grapevine virus L: A novel vitivirus in grapevine. Eur. J. Plant Pathol. 2019, 1, 319–328. [Google Scholar] [CrossRef]
- Meng, B.; Rowhani, A. Grapevine rupestris stem pitting-associated virus. In Grapevine Viruses: Molecular Biology, Diagnostics and Management, 1st ed.; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 257–288. [Google Scholar]
- Meng, B.; Rebelo, A.R.; Fisher, H. Genetic diversity analysis of Grapevine rupestris stem pitting-associated virus: Revelation of distinct population structures in scion versus rootstock varieties. J. Gen. Virol. 2006, 87, 1725–1733. [Google Scholar] [CrossRef]
- Nolasco, G.; Mansinho, A.; Santos, M.T.; Soares, C.; Sequeira, Z.; Sequeira, C.; Correia, P.K.; Sequeira, O.A. Large scale evaluation of primers for diagnosis of rupestris stem pitting associated virus-1. Eur. J. Plant Pathol. 2000, 106, 311–318. [Google Scholar] [CrossRef]
- Terlizzi, F.; Li, C.; Ratti, C.; Qu, W.; Credi, R.; Meng, B. Detection of multiple sequence variants of Grapevine rupestris stem pitting-associated virus using primers targeting the polymerase domain and partial genome sequencing of a novel variant. Ann. Appl. Biol. 2011, 159, 478–490. [Google Scholar] [CrossRef]
- Alabi, O.J.; Martin, R.R.; Naidu, R.A. Sequence diversity, population genetics and potential recombination events in Grapevine rupestris stem pitting-associated virus in Pacific North-West vineyards. J. Gen. Virol. 2010, 91, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Meng, B.; Credi, R.; Petrovic, N.; Tomazic, I.; Gonsalves, D. Antiserum to recombinant virus coat protein detects Rupestris stem pitting associated virus in grapevines. Plant Dis. 2003, 87, 515–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minafra, A.; Casati, P.; Elicio, V.; Rowhani, A.; Saldarelli, P.; Savino, V.; Martelli, G.P. Serological detection of grapevine rupestris stem pitting-associated virus (GRSPaV) by a polyclonal antiserum to recombinant virus coat protein. Vitis 2000, 39, 115–118. [Google Scholar]
- Al Rwahnih, M.; Daubert, S.; Golino, D.A.; Rowhani, A. Deep sequencing analysis of RNAs from a grapevine showing Syrah decline symptoms reveals a multiple virus infection that included a novel virus. Virology 2009, 387, 395–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Čarija, M.; Radić, T.; Černi, S.; Mucalo, A.; Zdunić, G.; Vončina, D.; Jagunić, M.; Hančević, K. Prevalence of Virus Infections and GLRaV-3 Genetic Diversity in Selected Clones of Croatian Indigenous Grapevine Cultivar Plavac Mali. Pathogens 2022, 11, 176. [Google Scholar] [CrossRef] [PubMed]
- Boudon-Padieu, E. Grapevine yellows diseases. In Compendium of Grape Diseases, Disorders, and Pests, 2nd ed.; Wilcox, W.F., Gubler, W.D., Uyemoto, J.K., Eds.; APS Press: St. Paul, MN, USA, 2015; pp. 103–114. [Google Scholar]
- Meng, B.; Johnson, R.; Peressini, S.; Forsline, P.L.; Gonsalves, D. Rupestris stem pitting associated virus-1 is consistently detected in grapevines that are infected with rupestris stem pitting. Eur. J. Plant Pathol. 1999, 105, 191–199. [Google Scholar] [CrossRef]
- Lima, M.F.; Alkowni, R.; Uyemoto, J.K.; Golino, D.; Osman, F.; Rowhani, A. Molecular analysis of a California strain of Rupestris stem pitting-associated virus isolated from declining Syrah grapevines. Arch. Virol. 2006, 151, 1889–1894. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acid. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Higgins, D.; Thompson, J.; Gibson, T.; Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucl. Acid. Res. 1994, 22, 4673–4680. [Google Scholar]
- Terlizzi, F.; Ratti, C.; Filippini, G.; Pisi, A.; Credi, R. Detection and molecular characterization of Italian isolates. Plant Pathol. 2010, 59, 48–58. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Al Rwahnih, M.; Diaz-Lara, A.; Arnold, K.; Cooper, M.L.; Smith, R.J.; Zhuang, G.; Battany, M.C.; Bettiga, L.J.; Rowhani, A.; Golino, D. Incidence and Genetic Diversity of Grapevine Pinot gris Virus in California. Am. J. Enol. Vitic. 2021, 72, 164–169. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comp. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kogovšek, P.; Hodgetts, J.; Hall, J.; Prezelj, N.; Nikolić, P.; Mehle, N.; Lenarčič, R.; Rotter, A.; Dickinson, M.; Boonham, N.; et al. LAMP assay and rapid sample preparation method for on-site detection of flavescence dorée phytoplasma in grapevine. Plant Pathol. 2015, 64, 286–296. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| No. on the Gels | Cultivar/Grapevine Accession | Western Blot | RT-PCR | Cloning/ Sequencing | ||
|---|---|---|---|---|---|---|
| As7-276 | As2003 | RSP13/RSP14 | 48V/49C | |||
| 1. | Babic 015 | + | + | + | + | + |
| 2. | Babic 056 | + | + | + | + | N/A |
| 3. | Babic 113 | +/− | − | + | + | N/A |
| 4. | Babic 076 | + | + | + | + | N/A |
| 5. | Marastina 034 | − | − | + | + | N/A |
| 6. | Marastina 058 | + | + | + | + | + |
| 7. | Marastina 101 | + | + | + | + | N/A |
| 8. | Posip 076 | + | + | + | + | + |
| 9. | Vugava 060 | + | + | + | + | N/A |
| 10. | Vugava 115 | + | + | + | + | + |
| 11. | Plavac mali 013 | + | + | + | + | N/A |
| 12. | Plavac mali 085 | + | + | + | + | + |
| 13. | Plavac mali 234 | + | + | + | − | + |
| 14. | Plavac mali 286 | + | + | + | + | N/A |
| 15. | Plavac mali 111 | + | + | + | + | N/A |
| 16. | Plavac mali 128 | − | − | + | + | + |
| 17. | Plavac mali 159 | + | + | + | + | N/A |
| 18. | Plavac mali 181 | + | + | + | + | N/A |
| 19. | Plavac mali 219 | + | + | + | + | N/A |
| 20. | Babica 106 | + | + | + | + | N/A |
| 21. | Dobricic 101 | − | − | + | + | N/A |
| 22. | Glavinusa 110 | + | + | + | + | + |
| 23. | Ljutun 167 | + | + | + | + | N/A |
| 24. | Mladenka 154 | + | + | + | + | N/A |
| 25. | Nincusa 108 | + | + | + | + | + |
| 26. | Vlaska 137 | +/− | +/− | + | + | N/A |
| Helicase (RSP13/RSP14) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Babic 015 | 100% | 96.96% | 97.97% | 95.95% | 97.97% | 97.97% | 97.97% | 98.98% | 97.97% |
| Glavinusa 108 | 92.64% | 100% | 96.96% | 94.94% | 98.98% | 98.98% | 98.98% | 97.97% | 98.98% |
| Marastina 058 | 92.64% | 96.65% | 100% | 95.95% | 97.97% | 97.97% | 97.97% | 98.98% | 97.97% |
| Nincusa 108 | 88.29% | 90.63% | 92.64% | 100% | 95.95% | 95.95% | 95.95% | 96.96% | 95.95% |
| Plavac mali 085 | 88.29% | 90.63% | 92.3% | 87.62% | 100% | 100% | 100% | 98.98% | 97.97% |
| Plavac mali 128 | 88.29% | 90.63% | 92.3% | 87.62% | 100% | 100% | 100% | 98.98% | 97.97% |
| Plavac mali 234 | 88.29% | 90.63% | 92.3% | 87.62% | 100% | 100% | 100% | 98.98% | 97.97% |
| Posip 076 | 93.31% | 96.98% | 98.32% | 91.63% | 92.97% | 92.97% | 92.97% | 100% | 98.98% |
| Vugava 115 | 92.3% | 98.99% | 96.32% | 90.3% | 90.96% | 90.96% | 90.96% | 96.65% | 100% |
| Babic 015 | Glavinusa 108 | Marastina 058 | Nincusa 108 | Plavac mali 085 | Plavac mali 128 | Plavac mali 234 | Posip 076 | Vugava 115 | |
| Coat protein (48V/49C) | |||||||||
| Babic 015 | 100% | 98.95% | 100% | 100% | 98.95% | 98.95% | N/A | 100% | 96.87% |
| Glavinusa 108 | 97.57% | 100% | 98.95% | 98.95% | 97.91% | 97.91% | N/A | 98.95% | 97.91% |
| Marastina 058 | 96.88% | 97.23% | 100% | 100% | 98.95% | 98.95% | N/A | 100% | 96.87% |
| Nincusa 108 | 96.88% | 97.23% | 100% | 100% | 98.95% | 98.95% | N/A | 100% | 96.87% |
| Plavac mali 085 | 95.15% | 94.8% | 94.8% | 94.8% | 100% | 100% | N/A | 98.95% | 95.83% |
| Plavac mali 128 | 95.15% | 94.8% | 94.8% | 94.8% | 100% | 100% | N/A | 98.95% | 95.83% |
| Plavac mali 234 * | N/A | N/A | N/A | N/A | N/A | N/A | 100% | 98.95% | 95.83% |
| Posip 076 | 95.15% | 94.8% | 94.8% | 94.8% | 99.3% | 99.3% | 99.3% | 100% | 96.87% |
| Vugava 115 | 96.88% | 97.92% | 96.53% | 96.53% | 94.11% | 94.11% | 94.11% | 94.11% | 100% |
| Babic 015 | Glavinusa 108 | Marastina 058 | Nincusa 108 | Plavac mali 085 | Plavac mali 128 | Plavac mali 234 | Posip 076 | Vugava 115 | |
| Croatian Isolate | Helicase | Coat Protein | ||||||
|---|---|---|---|---|---|---|---|---|
| GenBank Isolate | Country | % Identity | Accession No. | GenBank Isolate | Country | % Identity | Accession No. | |
| Babic 015 | BS | Canada | 98.66 | AY881627 | AV95 VVSY2 | Tunisia Brazil | 100 | LT855242 KT008377 |
| Glavinusa 108 | NV_60U OV_104I OV_104G OV_104A | South Africa South Africa South Africa South Africa | 99.33 | MG050684 MG050602 MG050601 MG050600 | NV_48E NV_48D NV_25D NV_25C LN-YEM-1 | South Africa South Africa South Africa South Africa China | 98.96 | MG050300 MG050299 MG050262 MG050261 KF731972 |
| Marastina 058 | Pagadebit 2–5 | South Africa | 97.99 | MG050684 | NV_09E | South Africa | 99.65 | MG050227 |
| Nincusa 108 | SK704-C | Slovakia | 98.32 | KX274276 | NV_09E | South Africa | 99.65 | MG050227 |
| Plavac mali 085 | GRSPaV-1 | - | 99.67 | AF057136 | NV_05C E78-RSP LN-XY1–3 AFH-2 PG | South Africa Spain China China Italy | 99.65 | MG050219 KJ466314 KJ634636 MK867356 HE591388 * |
| Plavac mali 128 | Pagadebit 2–5 | Italy | 97.99 | DQ278627 | NV_09E | South Africa | 98.62 | MG050227 |
| Plavac mali 234 | 34 clone 2 | France | 100 | MG938304 | N/A ** | N/A ** | N/A ** | N/A ** |
| Posip 076 | Pagadebit 2–5 | Italy | 98.32 | DQ278627 | NUB1_BR AMCF clone 2 NV_38U NV_38B MB_93B | Brazil France South Africa South Africa South Africa | 100 | MK804766 MG938310 MG050435 MG050288 MG050192 * |
| Vugava 115 | NV_60U OV_104I OV_104G OV_104A | South Africa South Africa South Africa South Africa | 99% | MG050684 MG050602 MG050601 MG050600 | NV_60B NV_25D NV_25C NV_06B LN-YEM-1 | South Africa South Africa South Africa South Africa China | 98.96 | MG050326 MG050262 MG050261 MG050220 KF731972 * |
| Grapevine Accession ID | ArMV | GFLV | GFkV | GLRaV-1 | GLRaV-2 | GLRaV-3 | GVA | GVB | GVE | GVF | GVG | GVT | GLRaV-4 | GLRaV-4 strain 5 | GLRaV-4 strain 6 | GLRaV-4 strain 9 | GRSPaV | GDefV | GRLDaV | GBV-1 | GRVFV | GSyV-1 | GYSVd-1 | GYSVd-2 | HSVd |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Babica 122 | + | + + | + + | + + | + | + | + | + | + | + | |||||||||||||||
| Babic 055 | + | + + | + + | + + | + | + | + | + | |||||||||||||||||
| Babic 063 | + + | + + | + + | + + | + | + | + | + | + | ||||||||||||||||
| Dobricic 021 | + + | + + | + | + | + | + | + | ||||||||||||||||||
| Glavinusa 102 | + + | + + | + + | + | + | + | + | + | + | + | |||||||||||||||
| Marastina 016 | + | + + | + | + | + | + | + | ||||||||||||||||||
| Marastina 037 | + | + + | + + | + + | + | + | + | + | |||||||||||||||||
| Mladenka 160 | + + | + + | + + | + | + | + | + | ||||||||||||||||||
| Plavac mali 010 | + + | + + | + + | + | + | + | + | + | + | ||||||||||||||||
| Plavac mali 084 | + + | + | + + | + | + | + | + | + | |||||||||||||||||
| Plavac mali 113 | + + | + + | + + | + | + | ||||||||||||||||||||
| Plavac mali 219 | + + | + + | + + | + + | + + | + | + | + | + | + | + | ||||||||||||||
| Plavac mali 235 | + | + + | + + | + + | + | + | + | + | + | + | |||||||||||||||
| Plavac mali 275 | + + | + + | + | + | + | + | + | ||||||||||||||||||
| Plavac mali 282 | + + | + + | + + | + + | + | + | + | + | |||||||||||||||||
| Plavac mali 313 | + + | + + | + | + | + | ||||||||||||||||||||
| Posip 056 | + + | + | + | + | + | ||||||||||||||||||||
| Posip 084 | + | + + | + | + | + | ||||||||||||||||||||
| Posip 123 | + | + + | + | + | + | ||||||||||||||||||||
| Vlaska 150 | + | + + | + | + | + | + | + | ||||||||||||||||||
| Vugava 036 | + | + + | + + | + | + | ||||||||||||||||||||
| Vugava 112 | + + | + + | + | + | + | + | + | + | + |
| Grapevine Accession | FD-LAMP | |
|---|---|---|
| 2020 | 2021 | |
| Grk 003 | − | N/A |
| Mladenka 148 | − | N/A |
| Plavac mali 010 | N/A | − |
| Plavac mali 011 | N/A | − |
| Plavac mali 013 | N/A | − |
| Plavac mali 105 | N/A | − |
| Plavac mali 272 | + | L |
| Plavac mali 313 | + | + |
| Plavac mali 401 | N/A | + |
| Posip 064 | N/A | − |
| Vlaska 122 | N/A | + |
| Vlaška 123 | N/A | − |
| Vlaska 131 | + | L |
| Vlaska 143 | − | − |
| Vlaska 150 | + | L |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vončina, D.; Diaz-Lara, A.; Preiner, D.; Al Rwahnih, M.; Stevens, K.; Jurić, S.; Malenica, N.; Šimon, S.; Meng, B.; Maletić, E.; et al. Virus and Virus-like Pathogens in the Grapevine Virus Collection of Croatian Autochthonous Grapevine Cultivars. Plants 2022, 11, 1485. https://doi.org/10.3390/plants11111485
Vončina D, Diaz-Lara A, Preiner D, Al Rwahnih M, Stevens K, Jurić S, Malenica N, Šimon S, Meng B, Maletić E, et al. Virus and Virus-like Pathogens in the Grapevine Virus Collection of Croatian Autochthonous Grapevine Cultivars. Plants. 2022; 11(11):1485. https://doi.org/10.3390/plants11111485
Chicago/Turabian StyleVončina, Darko, Alfredo Diaz-Lara, Darko Preiner, Maher Al Rwahnih, Kristian Stevens, Snježana Jurić, Nenad Malenica, Silvio Šimon, Baozhong Meng, Edi Maletić, and et al. 2022. "Virus and Virus-like Pathogens in the Grapevine Virus Collection of Croatian Autochthonous Grapevine Cultivars" Plants 11, no. 11: 1485. https://doi.org/10.3390/plants11111485
APA StyleVončina, D., Diaz-Lara, A., Preiner, D., Al Rwahnih, M., Stevens, K., Jurić, S., Malenica, N., Šimon, S., Meng, B., Maletić, E., Fulgosi, H., & Cvjetković, B. (2022). Virus and Virus-like Pathogens in the Grapevine Virus Collection of Croatian Autochthonous Grapevine Cultivars. Plants, 11(11), 1485. https://doi.org/10.3390/plants11111485