
Potential Distribution of and Sensitivity Analysis for Urochloa panicoides Weed Using Modeling: An Implication of Invasion Risk Analysis for China and Europe

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Material and Methods
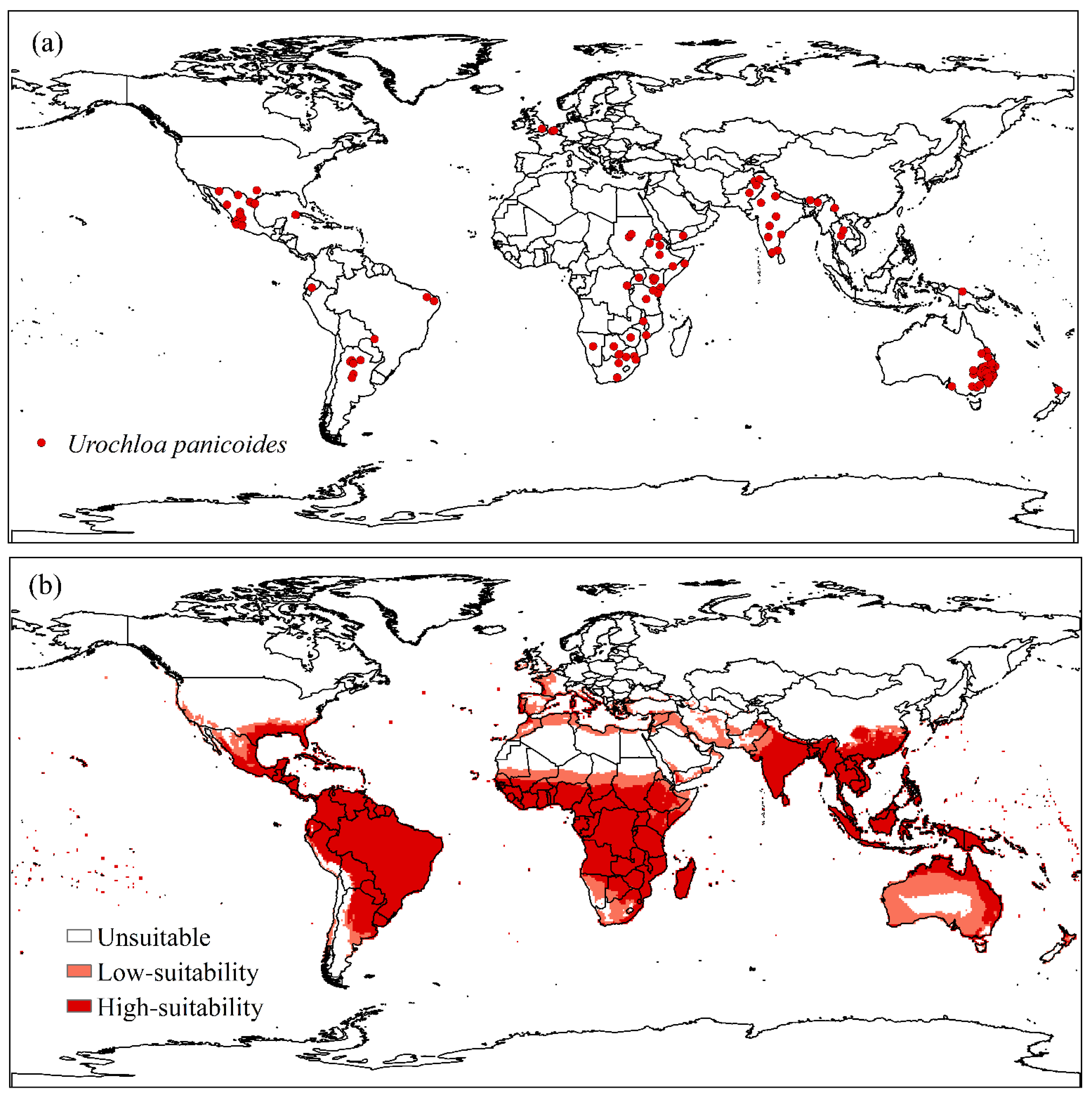
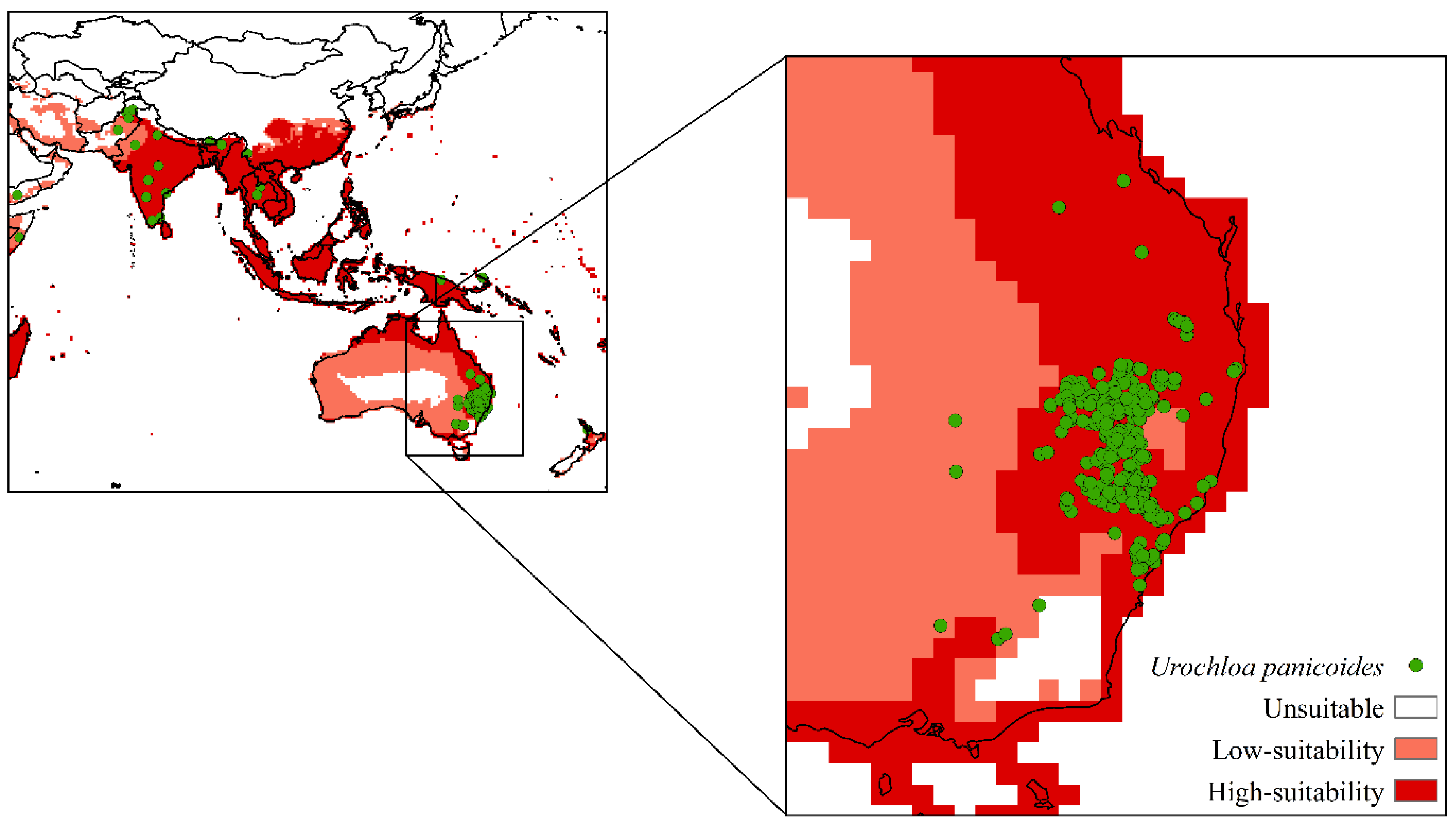
2.1. Global Distribution of Urochloa panicoides
2.2. CLIMEX
2.3. Parameter Adjustments and Model Validation in CLIMEX Software
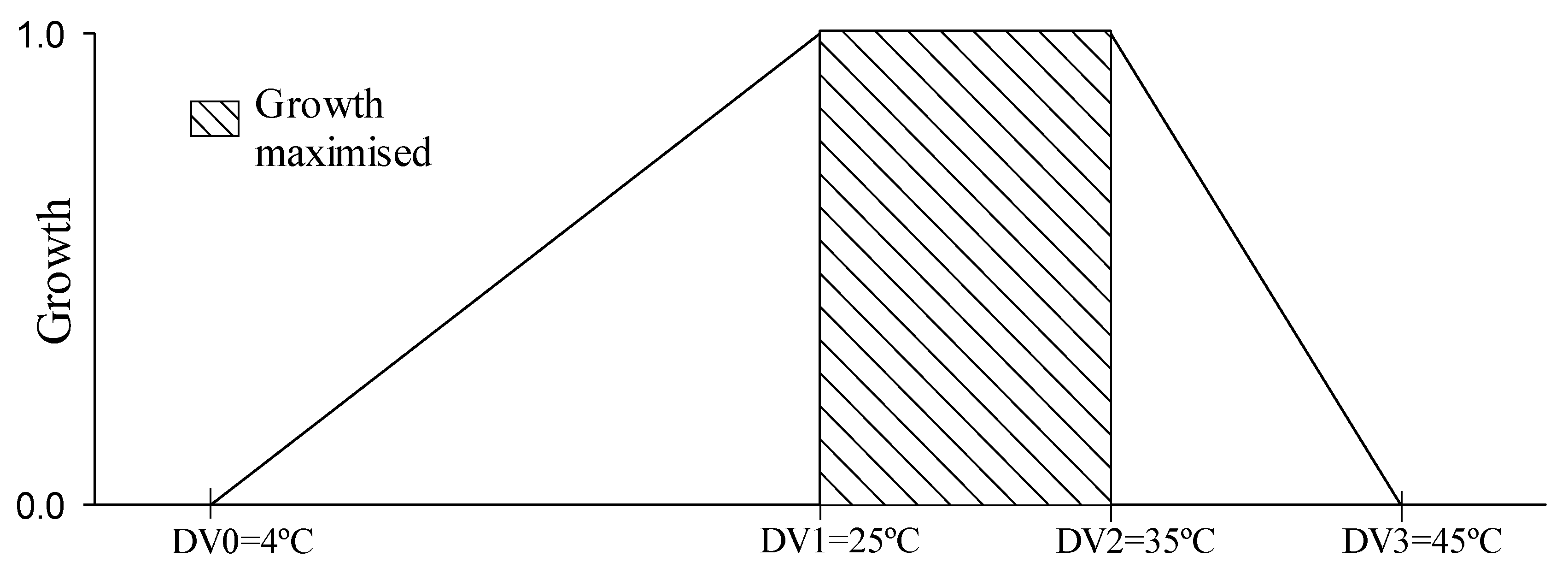
2.3.1. Growth Indices
2.3.2. Stress Parameters
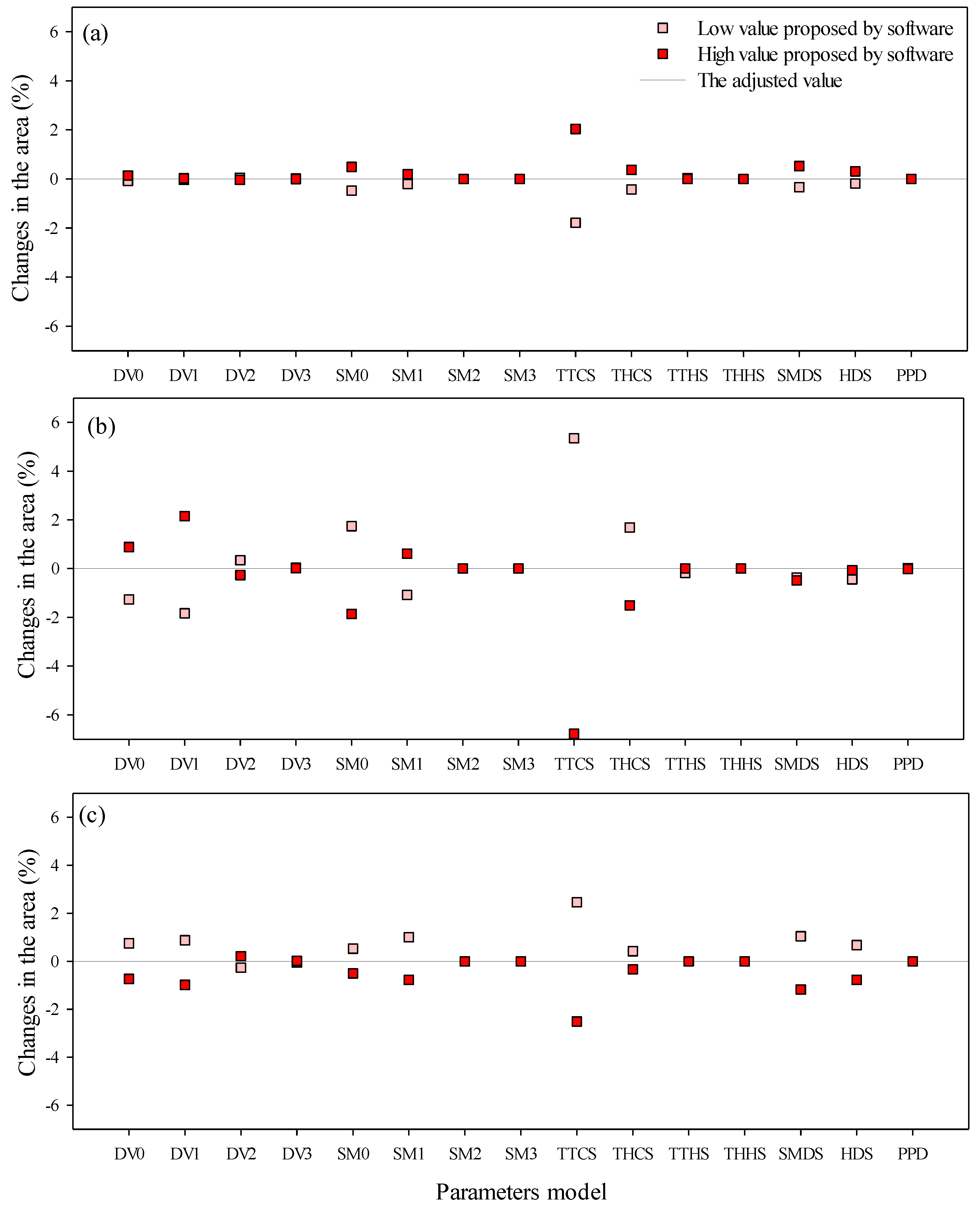
2.4. Sensitivity Analysis Using CLIMEX
2.5. Climate Data, Models, and Scenarios
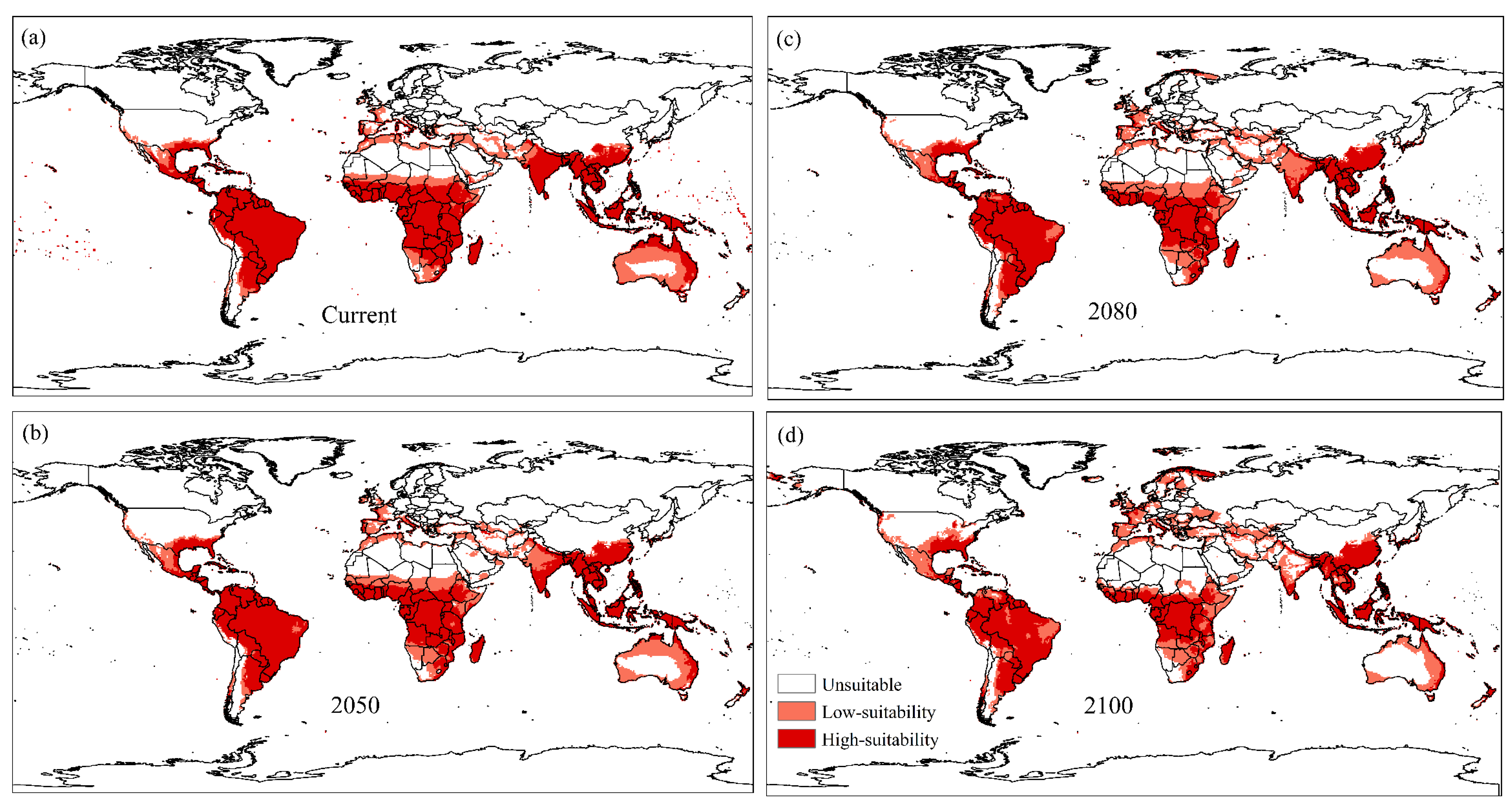
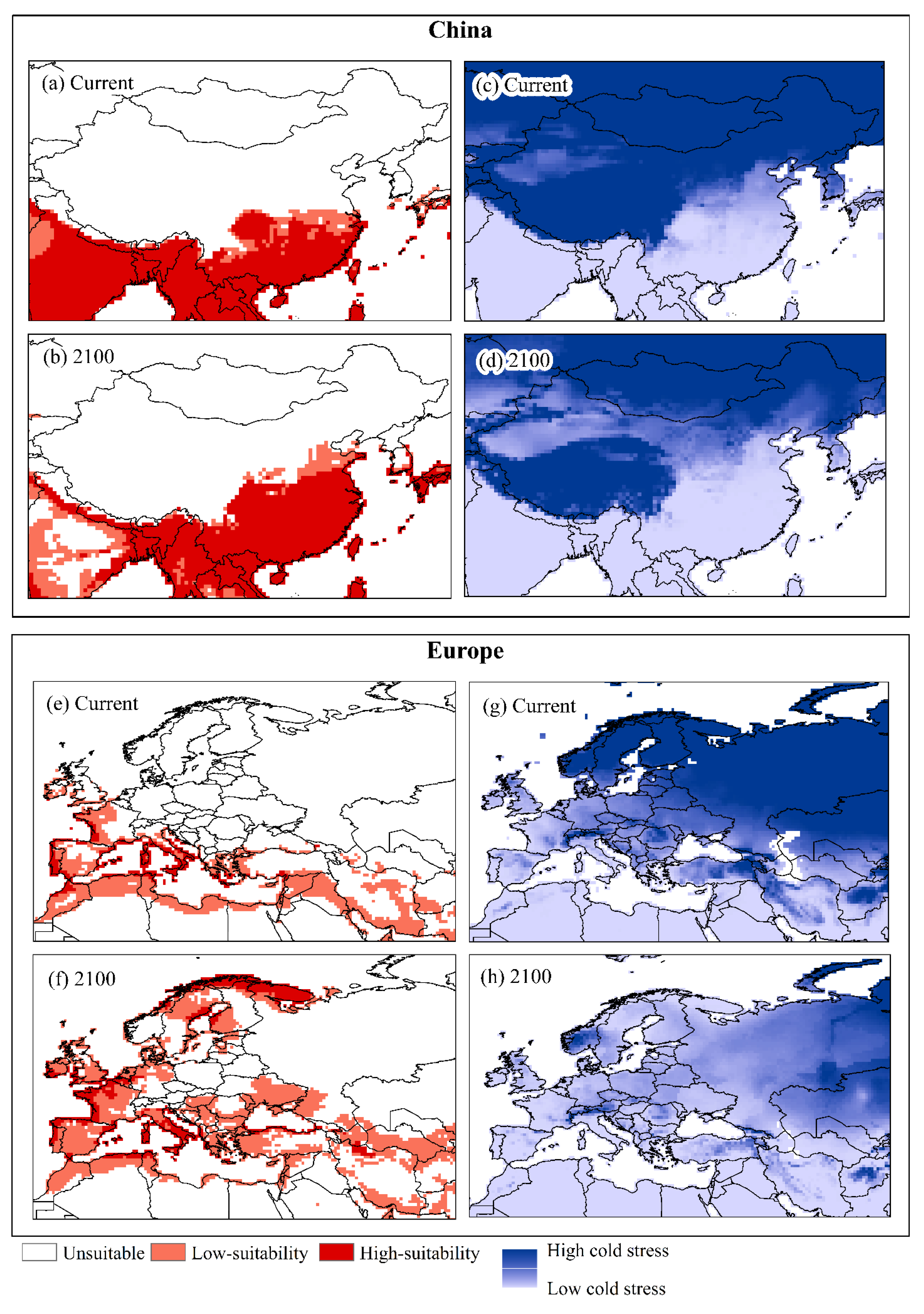
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonnamour, A.; Gippet, J.M.; Bertelsmeier, C. Insect and plant invasions follow two waves of globalisation. Ecol. Lett. 2021, 24, 2418–2426. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Richardson, D.M. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef] [PubMed]
- Diagne, C.; Leroy, B.; Gozlan, R.E.; Vaissière, A.C.; Assailly, C.; Nuninger, L.; Roiz, D.; Jourdain, F.; Jaric, I.; Courchamp, F. InvaCost, a public database of the economic costs of biological invasions worldwide. Sci. Data 2020, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Paini, D.R.; Sheppard, A.W.; Cook, D.C.; De Barro, P.J.; Worner, S.P.; Thomas, M.B. Global threat to agriculture from invasive species. Proc. Natl. Acad. Sci. USA 2016, 113, 7575–7579. [Google Scholar] [CrossRef] [PubMed]
- Carboni, M.; Livingstone, S.W.; Isaac, M.E.; Cadotte, M.W. Invasion drives plant diversity loss through competition and ecosystem modification. J. Ecol. 2021, 109, 3587–3601. [Google Scholar] [CrossRef]
- Zhang, Z.; Suseela, V. Nitrogen availability modulates the impacts of plant invasion on the chemical composition of soil organic matter. Soil Biol. Biochem. 2021, 156, 108195. [Google Scholar] [CrossRef]
- Ni, M.; Deane, D.C.; Li, S.; Wu, Y.; Sui, X.; Xu, H.; Chu, C.; He, F.; Fang, S. Invasion success and impacts depend on different characteristics in non-native plants. Divers Distrib. 2021, 27, 1194–1207. [Google Scholar] [CrossRef]
- Van Kleunen, M.; Pyšek, P.; Dawson, W.; Essl, F.; Kreft, H.; Pergl, J.; Winter, M. The global naturalized alien Flora (Glo NAF) database. Ecology 2018, 100, e02542. [Google Scholar] [CrossRef] [PubMed]
- Roura-Pascual, N.; Leung, B.; Rabitsch, W.; Rutting, L.; Vervoort, J.; Bacher, S.; Essl, F. Alternative futures for global biological invasions. Sustain. Sci. 2021, 16, 1637–1650. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Essl, F.; Evans, T.; Hulme, P.E.; Jeschke, J.M.; Kühn, I.; Bacher, S. A unified classification of alien species based on the magnitude of their environmental impacts. PLoS Biol. 2014, 12, e1001850. [Google Scholar] [CrossRef] [PubMed]
- Foxcroft, L.C.; Richardson, D.M.; Rejmánek, M.; Pyšek, P. Alien plant invasions in tropical and sub-tropical savannas: Patterns, processes and prospects. Biol. Invasions 2010, 12, 3913–3933. [Google Scholar] [CrossRef]
- Anderson, N.O.; Galatowitsch, S.M.; Gomez, N. Selection strategies to reduce invasive potential in introduced plants. Euphytica 2006, 148, 203–216. [Google Scholar] [CrossRef]
- Ustarroz, D.; Kruk, B.C.; Satorre, E.H.; Ghersa, C.M. Dormancy, germination and emergence of Urochloa panicoides regulated by temperature. Weed Res. 2016, 56, 59–68. [Google Scholar] [CrossRef]
- Malan, L. Biology and Germination Characteristics of Urochloa mosambicensis and Urochloa panicoides. Ph.D. Thesis, North-West University, Vanderbijlpark, South Africa, 2018. [Google Scholar]
- U.S. Department of Agriculture. Federal Noxious Weed List. Available online: https://www.aphis.usda.gov/plant_health/plant_pest_info/weeds/downloads/weedlist.pdf (accessed on 30 June 2022).
- Johnson, W.G.; Davis, V.M.; Kruger, G.R.; Weller, S.C. Influence of glyphosate-resistant cropping systems on weed species shifts and glyphosate-resistant weed populations. Eur J Agron 2009, 31, 162–172. [Google Scholar] [CrossRef]
- Riaz, T.; Javaid, A. Invasion of exotic weed Parthenium hysterophorus in district Sheikhupura, Pakistan. Int. J. Biol. Biotech. 2007, 4, 163–166. [Google Scholar]
- Villaseñor, J.L.J.; Espinosa-Garcia, F. The alien flowering plants of Mexico. Divers Distrib. 2004, 10, 113–123. [Google Scholar] [CrossRef]
- Souza, E.B.; Junior, G.A.D.; Pott, A. Soil seed bank in Pantanal riparian forest: Persistence, abundance, functional diversity and composition. Oecologia Aust. 2019, 23, 891–903. [Google Scholar] [CrossRef]
- CABI. Invasive Species Compendium. Urochloa panicoides (Liverseed Grass). Available online: https://www.cabi.org/isc/datasheet/55773 (accessed on 5 May 2022).
- GBIF.org. Global Biodiversity Information Facility. Urochloa panicoides P. Beauv. Available online: https://www.gbif.org/pt/species/2705869 (accessed on 5 May 2022).
- INDEX MUNDI. Available online: https://www.indexmundi.com/agriculture/?commodity=cotton&graph=production&display=map (accessed on 4 March 2022).
- Bogale, G.A.; Tolossa, T.T. Climate change intensification impacts and challenges of invasive species and adaptation measures in Eastern Ethiopia. Sustain. Environ. Res. 2021, 7, 1875555. [Google Scholar] [CrossRef]
- Van Kleunen, M.; Weber, E.; Fischer, M. A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol. Lett. 2010, 13, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Dukes, J.S.; Mooney, H.A. Does global change increase the success of biological invaders? Trends Ecol. Evol. 1999, 14, 135–139. [Google Scholar] [CrossRef]
- Westphal, M.I.; Browne, M.; MacKinnon, K.; Noble, I. The link between international trade and the global distribution of invasive alien species. Biol. Invasions 2008, 10, 391–398. [Google Scholar] [CrossRef]
- MacKenzie, D. Sick to death. New Sci. 2000, 167, 32–35. [Google Scholar]
- Sutherst, R.W. The vulnerability of animal and human health to parasites under global change. Int. J. Parasitol. 2001, 31, 933–948. [Google Scholar] [CrossRef]
- Sharma, G.P.; Esler, K.J.; Blignaut, J.N. Determining the relationship between invasive alien species density and a country’s socio-economic status. S. Afr. J. Sci. 2010, 106, 1–6. [Google Scholar] [CrossRef][Green Version]
- Pyšek, P.; Richardson, D.M. Traits associated with invasiveness in alien plants: Where do we stand? Biol. Invasions 2008, 193, 97–125. [Google Scholar]
- Allen, J.M.; Bradley, B.A. Out of the weeds? Reduced plant invasion risk with climate change in the continental United States. Biol. Conserv. 2016, 203, 306–312. [Google Scholar] [CrossRef]
- Renteria, J.L.; Darin, G.M.S.; Grosholz, E.D. Assessing the risk of plant species invasion under different climate change scenarios in California. Invasive Plant Sci. Manag. 2021, 14, 172–182. [Google Scholar] [CrossRef]
- Pratt, C.J.; Denley, D.; Metaxas, A. Selection of predictor variables for species distribution models: A case study with an invasive marine bryozoan. Oecologia 2022, 198, 319–336. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models: With Applications in R; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- Da Silva, R.S.; Kumar, L.; Shabani, F.; Da Silva, E.M.; Da Silva Galdino, T.V.; Picanço, M.C. Spatio-temporal dynamic climate model for Neoleucinodes elegantalis using CLIMEX. Int. J. Biometeorol. 2017, 61, 785–795. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Maywald, G.F.; Yonow, T.; Zurcher, E.J.; Herrmann, N.I.; Sutherst, R. Exploring the effects of climate on plants, animals and diseases. CLIMEX Version 2015, 4, 184. [Google Scholar]
- Jae-Min, J.; Sung-Hoon, J.; Byeon, D.H.; Wang-Hee, L. Invasion risk of the yellow crazy ant (Anoplolepis gracilipes) under the Representative Concentration Pathways 8.5 climate change scenario in South Korea. J Asia Pac Biodivers 2016, 10, 548–554. [Google Scholar]
- Ramos, R.S.; Kumar, L.; Shabani, F.; Da Silva, R.S.; De Araújo, T.A.; Picanço, M.C. Climate model for seasonal variation in Bemisia tabaci using CLIMEX in tomato crops. Int. J. Biometeorol. 2019, 63, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Luna, I.M. Caracterización de la dinámica de emergencia de cuatro especies poáceas en el departamento Moreno (Santiago del Estero). Ph.D. Thesis, Universidad Católica de Córdoba, Córdoba, Argentina, 2018. [Google Scholar]
- Kriticos, D.J.; Webber, B.L.; Leriche, A.; Ota, N.; Macadam, I.; Bathols, J.; Scott, J.K. CliMond: Global high-resolution historical and future scenario climate surfaces for bioclimatic modelling. Methods Ecol. Evol. 2012, 3, 53–64. [Google Scholar] [CrossRef]
- Gordon, H.B.; Rotstayn, L.D.; McGregor, J.L.; Dix, M.R.; Kowalczyk, E.A.; O’Farrell, S.P.; Waterman, L.J.; Hirst, A.C.; Wilson, S.G.; Collier, M.A.; et al. The CSIRO Mk3 Climate System Model; CSIRO Atmospheric Research: Aspendale, VIC, Australia, 2002. [Google Scholar]
- Chiew, F.; Kirono, D.; Kent, D.; Vaze, J. Assessment of rainfall simulations from global climate models and implications for climate change impact on runoff studies. In Proceedings of the 18th World Imacs Congress and Modsim09 International Congress on Modelling and Simulation: Interfacing Modelling and Simulation with Mathematical and Computational Sciences, Cairns, Australia, 13–17 July 2009; pp. 3907–3913. [Google Scholar]
- Van Vuuren, D.P.; Carter, T.R. Climate and socio-economic scenarios for climate change research and assessment: Reconciling the new with the old. Clim. Change 2004, 122, 415–429. [Google Scholar] [CrossRef]
- Suppiah, R.; Hennessy, K.J.; Whetton, P.H.; McInnes, K.; Macadam, I.; Bathols, J.; Page, C.M. Australian climate change projections derived from simulations performed for the IPCC 4th Assessment Report. Aust. Meteorol. Mag. 2007, 56, 131–152. [Google Scholar]
- Shoko, C.; Mutanga, O.; Dube, T. Optimal season for discriminating C3 and C4 grass functional types using multi-date Sentinel 2 data. GISci Remote Sens. 2020, 57, 127–139. [Google Scholar] [CrossRef]
- Barroso, G.M.; Da Silva, R.S.; Mucida, D.P.; Borges, C.E.; Ferreira, S.R.; Dos Santos, J.C.B.; Dos Santos, J.B. Spatio-Temporal Distribution of Digitaria insularis: Risk Analysis of Areas with Potential for Selection of Glyphosate-Resistant Biotypes in Eucalyptus Crops in Brazil. Sustain. Sci. 2021, 13, 10405. [Google Scholar] [CrossRef]
- Jung, J.M.; Jung, S.; Byeon, D.H.; Lee, W.H. Model-based prediction of potential distribution of the invasive insect pest, spotted lanternfly Lycorma delicatula (Hemiptera: Fulgoridae), by using CLIMEX. J. Asia-Pac. Biodivers. 2017, 10, 532–538. [Google Scholar] [CrossRef]
- Pattison, R.R.; Mack, R.N. Potential distribution of the invasive tree Triadica sebifera (Euphorbiaceae) in the United States: Evaluating CLIMEX predictions with field trials. Global Change Biol. 2008, 14, 813–826. [Google Scholar] [CrossRef]
- Shabani, F.; Kumar, L.; Esmaeili, A. Future distributions of Fusarium oxysporum f. spp. in European, Middle Eastern and North African agricultural regions under climate change. Agric. Ecosyst. Environ. 2014, 197, 96–105. [Google Scholar] [CrossRef]
- Walker, S.; Wu, H.; Bell, K. Emergence and Seed Persistence of Echinochloa colona, Urochloa panicoides and Hibiscus trionum in the Sub-tropical Environment of North-Eastern Australia. Plant Prot. Q. 2010, 25, 127–132. [Google Scholar]
- Anwar, M.; Islam, A.K.M.; Yeasmin, S.; Rashid, M.; Juraimi, A.S.; Ahmed, S.; Shrestha, A. Weeds and Their Responses to Management Efforts in A Changing Climate. Agron J. 2021, 11, 1921. [Google Scholar] [CrossRef]
- Amare, T. Review on impact of climate change on weed and their management. AJBES. 2016, 2, 21–27. [Google Scholar] [CrossRef]
- Da Silva, R.S. Impactos do aquecimento global na distribuição espaço-temporal do tomateiro e do inseto praga Neoleucinodes elegantalis. Ph.D. Thesis, Universidade Federal de Viçosa, Viçosa, Brazil, 2016. [Google Scholar]
- Walker, S.R.; Taylor, I.N.; Milne, G.; Osten, V.A.; Hoque, Z.; Farquharson, R.J. A survey of management and economic impact of weeds in dryland cotton cropping systems of subtropical Australia. Aust J. Exp. Agric. 2005, 45, 79–91. [Google Scholar] [CrossRef]
- Chandrasena, N. How will weed management change under climate change? Some perspectives. J. Crop. Weed 2009, 5, 95–105. [Google Scholar]
- Singh, M.C.; Dubey, S.C.; Yaduraju, N.T. Climate change and its possible impacts on weeds. Int. J. Environ. Sci. Technol. 2016, 5, 1530–1539. [Google Scholar]
- Yin, X.; Struik, P.C. Applying modelling experiences from the past to shape crop systems biology: The need to converge crop physiology and functional genomics. New Phytol. 2008, 179, 629–642. [Google Scholar] [CrossRef]
- Adkins, S.W.; Wills, D.; Boersma, M.; Walker, S.R.; Robinson, G.; McLeod, R.J.; Einam, J.P. Weeds resistant to chlorsulfuron and atrazine from the north east grain region of Australia. Weed Res. 1997, 37, 343–349. [Google Scholar] [CrossRef]
- Peters, K.; Breitsameter, L.; Gerowitt, B. Impact of climate change on weeds in agriculture: A review. Agron. Sustain. Dev. 2014, 34, 707–721. [Google Scholar] [CrossRef]
- Flora do Brasil. Urochloa panicoides P.Beauv. Available online: http://reflora.jbrj.gov.br/reflora/listaBrasil/FichaPublicaTaxonUC/FichaPublicaTaxonUC.do?id=FB135135 (accessed on 1 November 2021).
- Varela, A.E.; Cabrera, D.C.; De La Veja, M.; De La Fuente, E.B. Dinámica de emergencia de plántulas y control de Urochloa panicoides P. Beauv resistente a glifosato en Tucumán (Argentina). Rev. Agron. Noroeste Argent. 2011, 41, 27–38. [Google Scholar]
- Dellafiore, C.M.; Brignone, E.; Scilingo, V. Rol Ecológico Del Zorro Pampeano (Lycalopex gymnocercus) Como Dispersor De Semillas En Ambientes De Agroecosistema Y Bosques Serranos De Argentina. Eur. J. Sci. 2020, 16, 223–233. [Google Scholar] [CrossRef]
- Olea, I.; Sabaté, S.; Vinciguerra, F.; Devani, L. Pautas para el manejo de malezas en cultivos extensivos en el Noroeste Argentino. Avanço Agroind. 2014, 35, 25–34. [Google Scholar]
- Edgar, E.; Shand, J.E. Checklist of Panicoid grasses naturalised in New Zealand; with a key to native and naturalised genera and species. N. Z. J. Bot. 1987, 25, 343–353. [Google Scholar] [CrossRef]
- Naples, N.E.R.; Oviedo, I.B.; Cruz, R.E. Diversidad florística de la Cordillera de Guaniguanico, Cuba/Floristic diversity of the Cordillera de Guaniguanico, Cuba. Acta Botánica Cubana 2018, 217, 1–32. [Google Scholar]
- Wipff, J.K.; Lonard, R.I.; Jones, S.D.; Hatch, S.L. The genus Urochloa (poaceae: Paniceae) in Texas, including one previously unreported species for the state. SIDA Contrib. Bot. 1993, 15, 405–413. [Google Scholar]
- Dávila, P.; Mejia-Saulés, M.; Soriano-Martínez, A.M.; Herrera-Arrieta, Y. Conocimiento taxonómico de la familia Poaceae en México. Bot. Sci. 2018, 96, 462–514. [Google Scholar] [CrossRef]
- Morrone, O.; Zuloaga, F.O. Sinopsis del género Urochloa (Poaceae: Panicoideae: Paniceae) para México y América Central. Darwiniana 1993, 32, 59–75. [Google Scholar]
- Rivera, L.M.M.; Ortiz-Arrona, C.I. Investigación y Gestión de los Recursos Naturales de la Cuenca del Rio Ayuquila-Armería, Jalisco; Universidad de Guadalajara: Guadalajara, México, 2020. [Google Scholar]
- Sánchez-Escalante, J.J.; Van Devender, T.R.; Reina-Guerrero, A.L. Preliminary Flora of the Sierra La Púrica, Sonora, Mexico. Available online: https://www.researchgate.net/profile/Jose-Sanchez-Escalante/publication/329761058_Preliminary_Flora_of_the_Sierra_La_Purica_Sonora_Mexico/links/5c197064a6fdccfc70586e81/Preliminary-Flora-of-the-Sierra-La-Purica-Sonora-Mexico.pdf (accessed on 30 June 2022).
- Arrieta, Y.H.; Ortiz, A.C. Diversidad y distribución de las gramíneas (poaceae) en el estado de Zacatecas. J. Bot. Res. Inst. Tex. 2009, 3, 775–792. [Google Scholar]
- Steinmann, V.W. Flora y Vegetación de la Reserva de la Biosfera Zicuirán-Infiernillo, Michoacán, México. Bot. Sci. 2021, 99, 661–707. [Google Scholar] [CrossRef]
- Nigatu, L.; Hassen, A.; Sharma, J.; Adkins, S.W. Impact of Parthenium hysterophorus on grazing land communities in north-eastern Ethiopia. Weed Biol. Manag. 2010, 10, 143–152. [Google Scholar] [CrossRef]
- Ahmad, Z.; Khan, S.M.; Ali, S.; Rahman, I.U.; Ara, H.; Noreen, I.; Khan, A. Indicator species analyses of weed communities of maize crop in district Mardan, Pakistan. Pak. J. Weed Sci. Res. 2016, 22, 227–238. [Google Scholar]
- Ernst, W.H.O.; Veenendaal, E.M.; Kebakile, M.M. Possibilities for dispersal in annual and perennial grasses in a savanna in Botswana. Vegetatio 1992, 102, 1–11. [Google Scholar] [CrossRef]
- Sintayehu, A. Weed flora survey in field crops of Northwestern Ethiopia. Afri. J. Agric. Res. 2019, 14, 749–758. [Google Scholar] [CrossRef]
- Sadasivaiah, B.; Priyadarshini, P.; Rao, B.R.P. Three rare grasses (Eragrostis ciliaris var. clarkei, E. tenella var. insularis and Urochloa panicoides var. velutina) from Andhra Pradesh, India. J. Sci. Trans. Eviron. Technov. 2013, 6, 166–167. [Google Scholar]
- Kiran, M.; Rahees, N.; Vishal, V.; Vidyasagaran, K. Floristic diversity and structural dynamics of mangroves in the north west coast of kerala, India. J. Plant Dev. 2015, 7, 549–553. [Google Scholar]
- Kabir, M.E.; Webb, E.L. Community in Conservation: A Case from a Deciduous Forest under Community Management in Northeast Thailand. In Proceedings of the FORTROP II: Tropical Forestry Change in a Changing World, Bangkok, Thailand, 17–20 November 2008. [Google Scholar]
- Rahman, I.U.; Afzal, A.; Iqbal, Z.; Ijaz, F.; Ali, N.; Asif, M.; Alam, J.; Majid, A.; Hart, R.; Bussmann, R.W. First insights into the floristic diversity, biological spectra and phenology of Manoor Valley, Pakistan. Pak. J. Bot. 2018, 50, 1113–1124. [Google Scholar]
- Labrada, R. Revisión de la lista de malezas cuarentenarias de Ecuador. Technique 2015, 15, 58–68. [Google Scholar] [CrossRef]
- Hemp, A. Ecology of the pteridophytes on the southern slopes of Mt. Kilimanjaro. Part II: Habitat selection. Plant Biol. 2001, 3, 493–523. [Google Scholar] [CrossRef]
- Dangerfield, J.M.; Veenendaal, E.M.; Riddoch, B.J.; Black, H. Termites and land use in south-east Botswana: Variety and abundance of termite surface features. Botsw. Notes Rec. 1992, 24, 165–179. [Google Scholar]
- Kumar, K.K.; Babu, P.K. Diversity of Weed Flora in Citrus at ICAR-Regional Research Centre for Citrus, Biswanath Chariali, Assam, India. Int. J. Curr. Microbiol. App. Sci. 2019, 8, 2100–2107. [Google Scholar] [CrossRef]
- Velayudham, L.; Jeyaprakash, K.; Balachandran, N.; Rajantheran, M. Lindernia micrantha D. Don (Linderniaceae): A red listed plant species new discovery to Tamil Nadu, India. Plant Arch. 2020, 20, 1404–1408. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Code | Unit | Low Values | Adjusted Parameter Values | High Values | References |
|---|---|---|---|---|---|---|
| Limiting low temperature | DV0 | °C | 3 | 4 | 5 | Ustarroz, 2011; Ustarroz et al., 2015 |
| Lower optimal temperature | DV1 | °C | 24 | 25 | 26 | |
| Upper optimal temperature | DV2 | °C | 34 | 35 | 36 | |
| Limiting high temperature | DV3 | °C | 44 | 45 | 46 | |
| Limiting low moisture | SM0 | -- | 0.09 | 0.1 | 0.11 | ---- |
| Lower optimal moisture | SM1 | -- | 0.18 | 0.2 | 0.22 | |
| Upper optimal moisture | SM2 | -- | 7.2 | 8 | 8.8 | |
| Limiting high moisture | SM3 | -- | 9 | 10 | 11 | |
| Cold stress temperature threshold | TTCS | °C | 3 | 4 | 5 | Ustarroz, 2011; Ustarroz et al., 2015 |
| Cold stress temperature rate | THCS | week−1 | −0.0018 | −0.002 | −0.0022 | |
| Heat stress temperature threshold | TTHS | °C | 44 | 45 | 46 | |
| Heat stress temperature rate | THHS | week−1 | 0.018 | 0.02 | 0.022 | |
| Dry stress threshold | SMDS | -- | 0.09 | 0.1 | 0.11 | ---- |
| Dry stress rate | HDS | week−1 | −0.009 | −0.01 | −0.011 | |
| Degree-days | PPD | °C days | 1497 | 1517 | 1537 | Luna, 2018 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duque, T.S.; da Silva, R.S.; Maciel, J.C.; Silva, D.V.; Fernandes, B.C.C.; Júnior, A.P.B.; Santos, J.B.d. Potential Distribution of and Sensitivity Analysis for Urochloa panicoides Weed Using Modeling: An Implication of Invasion Risk Analysis for China and Europe. Plants 2022, 11, 1761. https://doi.org/10.3390/plants11131761
Duque TS, da Silva RS, Maciel JC, Silva DV, Fernandes BCC, Júnior APB, Santos JBd. Potential Distribution of and Sensitivity Analysis for Urochloa panicoides Weed Using Modeling: An Implication of Invasion Risk Analysis for China and Europe. Plants. 2022; 11(13):1761. https://doi.org/10.3390/plants11131761
Chicago/Turabian StyleDuque, Tayna Sousa, Ricardo Siqueira da Silva, Josiane Costa Maciel, Daniel Valadão Silva, Bruno Caio Chaves Fernandes, Aurélio Paes Barros Júnior, and José Barbosa dos Santos. 2022. "Potential Distribution of and Sensitivity Analysis for Urochloa panicoides Weed Using Modeling: An Implication of Invasion Risk Analysis for China and Europe" Plants 11, no. 13: 1761. https://doi.org/10.3390/plants11131761
APA StyleDuque, T. S., da Silva, R. S., Maciel, J. C., Silva, D. V., Fernandes, B. C. C., Júnior, A. P. B., & Santos, J. B. d. (2022). Potential Distribution of and Sensitivity Analysis for Urochloa panicoides Weed Using Modeling: An Implication of Invasion Risk Analysis for China and Europe. Plants, 11(13), 1761. https://doi.org/10.3390/plants11131761