
Phenoplasticity of Essential Oils from Two Species of Piper (Piperaceae): Comparing Wild Specimens and Bi-Generational Monoclonal Cultivars


 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Chemical Variation in P. gaudichaudianum
2.2. Chemical Variation in P. mollicomum
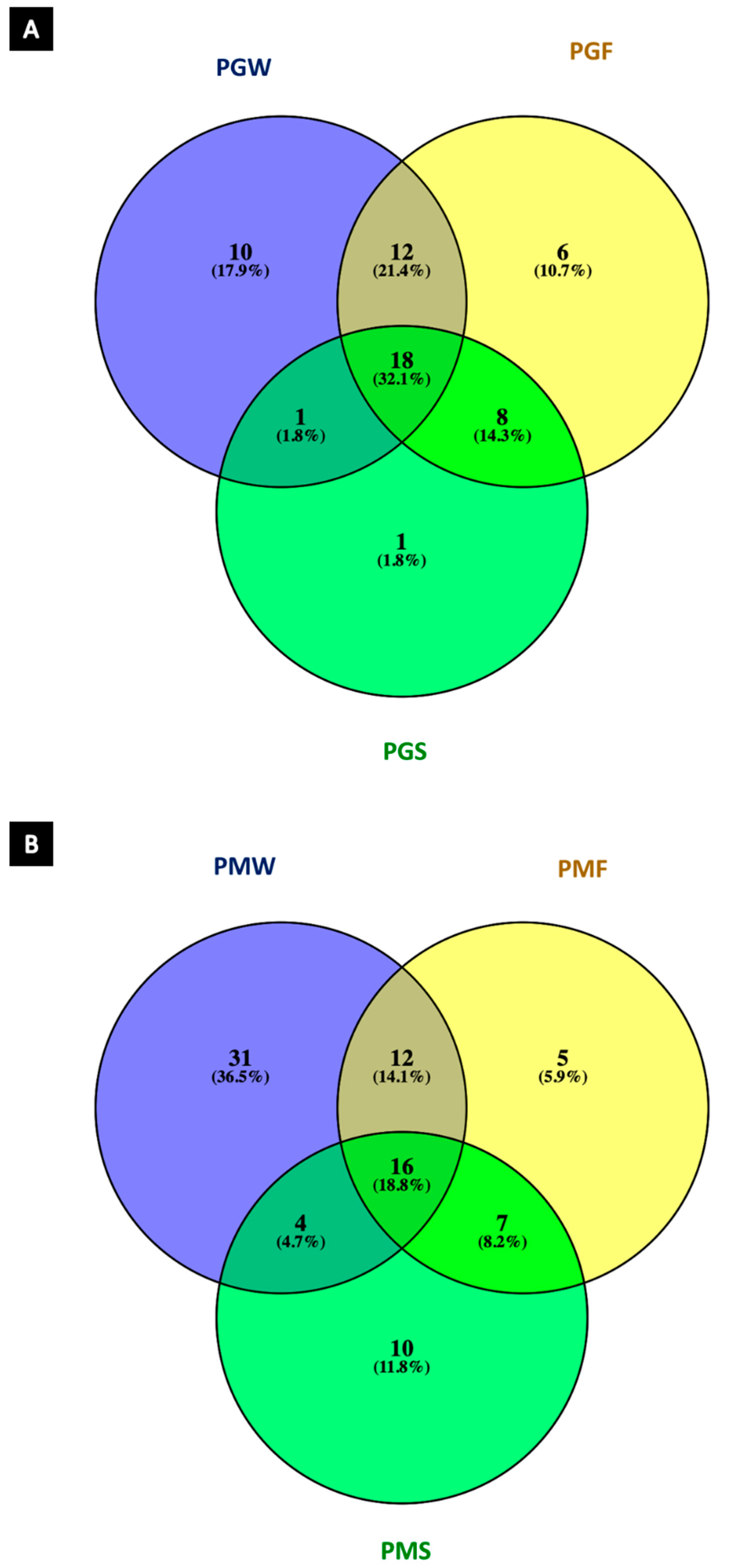
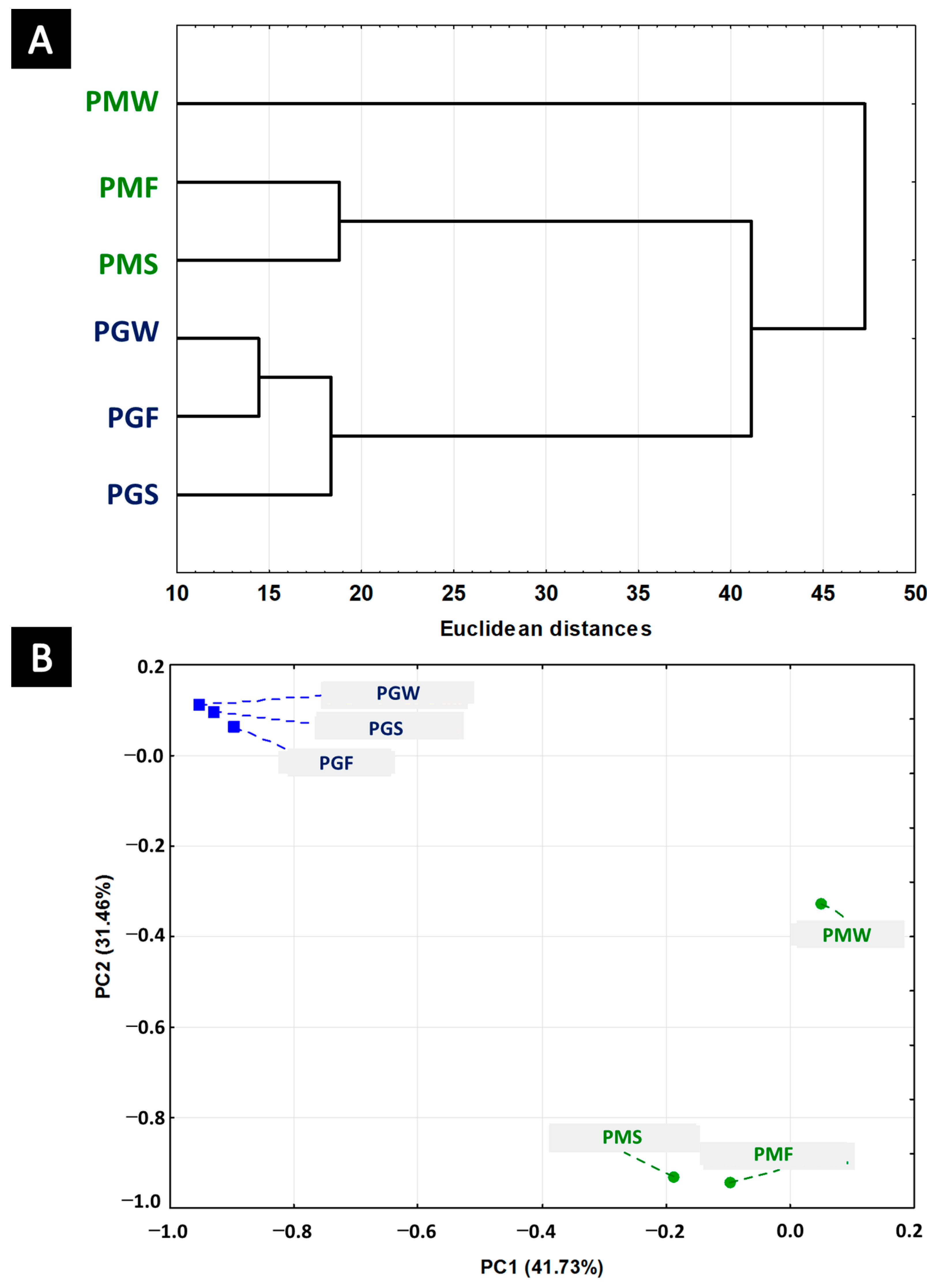
2.3. Chemometric Analysis
2.4. Soil Characteristics
2.5. Chemodiversity and Micromolecular Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Species Propagation Protocol
4.3. Essential Oil Production
4.4. Essential Oil Analysis
4.5. Evaluation of Chemodiversity and Micromolecular Parameters
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Takeara, R.; Gonçalves, R.; Ayres, V.; Guimarães, A.C. Biological properties of essential oils from the Piper species of Brazil: A review. Aromat. Med. Plants-Back Nat. 2017, 4, 81–93. [Google Scholar]
- Gogosz, A.M.; Boeger, M.R.T.; Negrelle, R.R.B.; Bergo, C. Anatomia foliar comparativa de nove espécies do gênero Piper (Piperaceae). Rodriguésia 2012, 63, 405–417. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L.A.; Lima, R.B.; Guimarães, E.F.; Almeida, M.Z.; Monteiro, E.D.C.Q.; Sobrinho, F.D.A.P. Plantas medicinais de uma comunidade quilombola na Amazônia Oriental: Aspectos utilitários de espécies das famílias Piperaceae e Solanaceae. Rev. Bras. De Agroecol. 2007, 2, 1385–1388. (In Portuguese) [Google Scholar]
- Silva, R.J.F.; de Aguiar-Dias, A.C.A.; Faial, K.D.C.F.; de Mendonca, M.S. Caracterização farmacognóstica de Piper arboreum var. arboreum e P. tuberculatum (Piperaceae). Acta Amaz. 2016, 46, 195–208. (In Portuguese) [Google Scholar]
- Guedes, R.R.; Profice, S.R.; Costa, E.d.L.; Baumgratz, J.F.A.; de Lima, C.H. Plantas utilizadas em rituais afro-brasileiros no Estado do Rio de Janeiro—um ensaio Etnobotânico. Rodriguésia 1985, 37, 3–9. (In Portuguese) [Google Scholar] [CrossRef] [Green Version]
- Verger, P. Ewé: O Uso das Plantas na Sociedade Iorubá; Companhia das letras: São Paulo, Brazil, 1995. (In Portuguese) [Google Scholar]
- Peckolt, T.; Peckolt, G. Historia das Plantas Medicinaes e Uteis do Brazil: I; Editora Lemmert: Rio de Janeiro, Brazil, 1888. (In Portuguese) [Google Scholar]
- Brito, M.R.d.; Senna-Valle, L.d. Plantas medicinais utilizadas na comunidade caiçara da Praia do Sono, Paraty, Rio de Janeiro, Brasil. Acta Bot. Bras. 2011, 25, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Corrêa, M.P. Dicionário das Plantas Úteis do Brasil e das Exóticas Cultivadas; Ministério da Agricultura: Rio de Janeiro, Brazil, 1984; p. 687. (In Portuguese) [Google Scholar]
- Duarte, M.C.T.; Leme, E.E.; Delarmelina, C.; Soares, A.A.; Figueira, G.M.; Sartoratto, A. Activity of essential oils from Brazilian medicinal plants on Escherichia coli. J. Ethnopharmacol. 2007, 111, 197–201. [Google Scholar] [CrossRef]
- Messias, M.C.T.B.; Menegatto, M.; Prado, A.; Santos, B.; Guimarães, M. Uso popular de plantas medicinais e perfil socioeconômico dos usuários: Um estudo em área urbana em Ouro Preto, MG, Brasil. Rev. Bras. De Plantas Med. 2015, 17, 76–104. [Google Scholar] [CrossRef]
- Mors, W.B.; Rizzini, C.T.; Pereira, N.A. Medicinal Plants of Brazil; Reference Publications, Inc.: Algonac, MI, USA, 2000. [Google Scholar]
- Lago, J.H.G.; Young, M.C.M.; Reigada, J.B.; Soares, M.G.; Roesler, B.P.; Kato, M.J. Antifungal derivatives from Piper mollicomum and P. ihotzkyanum (Piperaceae). Quim. Nova 2007, 30, 1222–1224. [Google Scholar] [CrossRef]
- Cordova, S.d.; Benfatti, C.S.; Magina, M.D.A.; Guedes, A.; Cordova, C.M.M.D. Análise da atividade antimicrobiana de extratos isolados de plantas nativas da flora brasileira frente a Mycoplasma arginini, M. hominis e Ureaplasma urealyticum. RBAC 2010, 42, 241–244. [Google Scholar]
- Valverde, S.S.; Costa, N.F.; Calheiros, A.S.; Lima, K.S.; Frutuoso, V.S.; Lima, A.; LS, N. Chemical composition and antinociceptive activity of the essential oil of Piper mollicomum and Piper rivinoides. J. Med. Plants Res. 2014, 8, 788–793. [Google Scholar]
- Guimarães, E.F.; Giordano, L.C.D.S. Piperaceae do Nordeste brasileiro I: Estado do Ceará. Rodriguésia 2004, 55, 21–46. [Google Scholar] [CrossRef] [Green Version]
- Salehi, B.; Zakaria, Z.A.; Gyawali, R.; Ibrahim, S.A.; Rajkovic, J.; Shinwari, Z.K.; Khan, T.; Sharifi-Rad, J.; Ozleyen, A.; Turkdonmez, E.; et al. Piper Species: A Comprehensive Review on Their Phytochemistry, Biological Activities and Applications. Molecules 2019, 24, 1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Stasi, L.C.; Hiruma-Lima, C.A. Plantas Medicinais na Amazônia e na Mata Atlântica; Editora Unesp: São Paulo, Brazil, 2002. [Google Scholar]
- Lago, J.H.G.; Ramos, C.S.; Casanova, D.C.C.; Morandim, A.D.A.; Bergamo, D.C.B.; Cavalheiro, A.J.; Bolzani, V.D.S.; Furlan, M.; Guimarães, E.F.; Young, M.C.M.; et al. Benzoic Acid Derivatives from Piper Species and Their Fungitoxic Activity against Cladosporium cladosporioides and C. sphaerospermum. J. Nat. Prod. 2004, 67, 1783–1788. [Google Scholar] [CrossRef] [PubMed]
- de Morais, S.M.; Facundo, V.A.; Bertini, L.M.; Cavalcanti, E.S.B.; Anjos Júnior, J.F.D.; Ferreira, S.A.; de Brito, E.S.; de Souza Neto, M.A. Chemical composition and larvicidal activity of essential oils from Piper species. Biochem. Syst. Ecol. 2007, 35, 670–675. [Google Scholar] [CrossRef]
- Ramos, Y.J.; da Costa-Oliveira, C.; Candido-Fonseca, I.; de Queiroz, G.A.; Guimarães, E.F.; Defaveri, A.C.; Sadgrove, N.J.; Moreira, D.D.L. Advanced Chemophenetic Analysis of Essential Oil from Leaves of Piper gaudichaudianum Kunth (Piperaceae) Using a New Reduction-Oxidation Index to Explore Seasonal and Circadian Rhythms. Plants 2021, 10, 2116. [Google Scholar] [CrossRef]
- Chaaban, A.; Santos, V.M.C.S.; Gomes, E.N.; Martins, C.E.N.; Amaral, W.D.; Deschamps, C.; Molento, M.B. Chemical composition of Piper gaudichaudianum essential oil and its bioactivity against Lucilia cuprina (Diptera: Calliphoridae). J. Essent. Oil Res. 2018, 30, 159–166. [Google Scholar] [CrossRef]
- Marques, A.M.; Peixoto, A.C.C.; Provance, D.W.; Kaplan, M.A.C. Separation of Volatile Metabolites from the Leaf-Derived Essential Oil of Piper mollicomum Kunth (Piperaceae) by High-Speed Countercurrent Chromatography. Molecules 2018, 23, 3064. [Google Scholar] [CrossRef] [Green Version]
- Ramos, Y.J.; Machado, D.D.B.; Queiroz, G.A.D.; Guimarães, E.F.; Defaveri, A.C.A.E.; Moreira, D.D.L. Chemical composition of the essential oils of circadian rhythm and of different vegetative parts from Piper mollicomum Kunth—A medicinal plant from Brazil. Biochem. Syst. Ecol. 2020, 92, 104116. [Google Scholar] [CrossRef]
- Sperotto, A.R.M.; Moura, D.J.; Péres, V.F.; Damasceno, F.C.; Caramão, E.B.; Henriques, J.A.P.; Saffi, J. Cytotoxic mechanism of Piper gaudichaudianum Kunth essential oil and its major compound nerolidol. Food Chem. Toxicol. 2013, 57, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, G.L.; Moreira, D.D.L.; Mendes, A.D.R.; Guimarães, E.F.; Figueiredo, L.S.; Kaplan, M.A.C.; Martins, E.R. Growth study and essential oil analysis of Piper aduncum from two sites of Cerrado biome of Minas Gerais State, Brazil. Rev. Bras. De Farmacogn. 2013, 23, 743–753. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, R.L.C. Etnobotânica e Plantas Medicinais: Estratégias de Conservação. Rev. De Biol. E Ciências Da Terra 2010, 10, 76–82. [Google Scholar]
- Dousseau, S.; de Alvarenga, A.A.; Alves, E.; Chaves, I.d.S.; Souza, E.D.S.; Alves, J.D.S. Physiological, morphological and biochemical characteristics of the sexual propagation of Piper aduncum (Piperaceae). Braz. J. Bot. 2011, 34, 297–305. [Google Scholar] [CrossRef] [Green Version]
- Ferriani, A.P.; Krinski, D. Propagation of pariparoba (Piperaceae) by different types of stem cuttings and substrates. Acta Biológica Catarin. 2019, 6, 75–80. [Google Scholar] [CrossRef]
- Gomes, E.N.; Krinski, D. Enraizamento de estacas caulinares de Piper crassinervium Kunth sob differentes concentracoes de acido indolbutirico. Rev. De Agric. Neotrop. 2019, 6, 92–97. [Google Scholar] [CrossRef]
- de Andrade, J.H.C.; Rodrigues, J.; Benites, A.; Benites, C.; Acosta, A.; Benites, M.; Benites, C.; Gomes, I.; da Silva, J.V.; Antunes, E.; et al. Notes on current Mbyá-Guarani medicinal plant exchanges in southern Brazil. J. Ethnobiol. Ethnomed. 2021, 17, 38. [Google Scholar] [CrossRef]
- Tng, D.Y.; Apgaua, D.M.G.; Lisboa, M.M.; El-Hani, C.N. Gender differences in plant use knowledge within a traditional fishing community in northeastern Brazil. Ethnobot. Res. Appl. 2021, 21, 1–36. [Google Scholar] [CrossRef]
- Jannuzzi, H.; Mattos, J.K.A.; Vieira, R.F.; Silva, D.B.; Bizzo, H.R. Avaliação agronômica e identificação de quimiotipos de erva cidreira no Distrito Federal. Hortic. Bras. 2010, 28, 412–417. [Google Scholar] [CrossRef] [Green Version]
- Nantongo, J.S.; Potts, B.M.; Davies, N.W.; Fitzgerald, H.; Rodemann, T.; O’Reilly-Wapstra, J.M. Additive genetic variation in Pinus radiata bark chemistry and the chemical traits associated with variation in mammalian bark stripping. Heredity 2021, 127, 498–509. [Google Scholar] [CrossRef]
- Scheiner, S.M. Genetics and Evolution of Phenotypic Plasticity. Annu. Rev. Ecol. Syst. 1993, 24, 35–68. [Google Scholar] [CrossRef]
- Kitavi, M.; Cashell, R.; Ferguson, M.; Lorenzen, J.; Nyine, M.; McKeown, P.C.; Spillane, C. Heritable epigenetic diversity for conservation and utilization of epigenetic germplasm resources of clonal East African Highland banana (EAHB) accessions. Theor. Appl. Genet. 2020, 133, 2605–2625. [Google Scholar] [CrossRef] [PubMed]
- Vallecillos, A.; María-Dolores, E.; Villa, J.; Rueda, F.M.; Carrillo, J.; Ramis, G.; Soula, M.; Afonso, J.M.; Armero, E. Phenotypic and Genetic Components for Growth, Morphology, and Flesh-Quality Traits of Meagre (Argyrosomus regius) Reared in Tank and Sea Cage. Animals 2021, 11, 3285. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, P.R.D.; Moreira, D.D.L.; Guimarães, E.F.; Kaplan, M.A.C. Essential oil analysis of 10 Piperaceae species from the Brazilian Atlantic Forest. Phytochemistry 2001, 58, 547–551. [Google Scholar] [CrossRef]
- Péres, V.F.; Moura, D.J.; Sperotto, A.R.M.; Damasceno, F.C.; Caramão, E.B.; Zini, C.A.; Saffi, J. Chemical composition and cytotoxic, mutagenic and genotoxic activities of the essential oil from Piper gaudichaudianum Kunth leaves. Food Chem. Toxicol. 2009, 47, 2389–2395. [Google Scholar] [CrossRef]
- Schindler, B.; Heinzmann, B.M. Piper gaudichaudianum Kunth: Seasonal Characterization of the Essential Oil Chemical Composition of Leaves and Reproductive Organs. Braz. Arch. Biol. Technol. 2017, 60. [Google Scholar] [CrossRef] [Green Version]
- Krinski, D.; Foerster, L.A. Toxicity of essential oils from leaves of Piperaceae species in rice stalk stink bug eggs, Tibraca limbativentris (Hemiptera: Pentatomidae). Ciência E Agrotecnol. 2016, 40, 676–687. [Google Scholar] [CrossRef] [Green Version]
- Bernuci, K.Z.; Iwanaga, C.C.; Fernandez-Andrade, C.M.M.; Lorenzetti, F.B.; Torres-Santos, E.C.; Faiões, V.D.S.; Gonçalves, J.E.; Do Amaral, W.; Deschamps, C.; Scodro, R.B.D.L.; et al. Evaluation of Chemical Composition and Antileishmanial and Antituberculosis Activities of Essential Oils of Piper Species. Molecules 2016, 21, 1698. [Google Scholar] [CrossRef] [Green Version]
- Krinski, D.; Foerster, L.A.; Deschamps, C. Ovicidal effect of the essential oils from 18 Brazilian Piper species: Controlling Anticarsia gemmatalis (Lepidoptera, Erebidae) at the initial stage of development. Acta Scientiarum. Agron. 2018, 40. [Google Scholar] [CrossRef] [Green Version]
- Del Quiqui, E.M.; Deschamps, C.; do Amaral, W.; Sipriano, R.R.; Machado, M.P. Yield and chemical composition of essential oil of Piperaceae in one segment of the semi deciduous forest of Paraná state, Brazil, in seasonal samplings. Int. J. Adv. Eng. Res. Sci. 2019, 6, 355–367. [Google Scholar] [CrossRef]
- da Silva, A.C.A.; Matias, E.F.F.; Rocha, J.E.; de Araújo, A.C.J.; de Freitas, T.S.; Campina, F.F.; Costa, M.d.S.; Silva, L.E.; do Amaral, W.; Maia, B.H.L.N.S.; et al. Gas chromatography coupled to mass spectrometry (GC-MS) characterization and evaluation of antibacterial bioactivities of the essential oils from Piper arboreum Aubl., Piper aduncum L. e Piper gaudichaudianum Kunth. Z. Für Nat. C 2021, 76, 35–42. [Google Scholar] [CrossRef]
- Silva, A.C.A.; Diodato, J.S.; Castro, J.W.; Matias, E.F.F.; Silva, L.E.; do Amaral, W.; Maia, B.H.L.N.S.; Ferriani, A.P.; Souza, A.K.; Quintans-Júnior, L.J.; et al. Effect of the essential oils from Piper sp. and blue led lights in the enhancement of the antibiotic activity of drugs against MDR bacterial strains. J. Photochem. Photobiol. B Biol. 2019, 199, 111604. [Google Scholar] [CrossRef] [PubMed]
- Morandim-Giannetti, A.d.A.; Pin, A.R.; Pietro, N.A.S.; Oliveira, H.C.d.; Mendes-Giannini, M.J.S.; Alecio, A.C.; Kato, M.J.; Oliveira, J.E.d.; Furlan, M. Composition and antifungal activity against Candida albicans, C. parapsilopisis, Candida krusei and Cryptococcus neoformans of essential oils from leaves of Piper and Peperomia species. J. Med. Plant Res. 2010, 4, 1810–1814. [Google Scholar]
- Santana, A.I.; Vila, R.; Cañigueral, S.; Gupta, M.P. Chemical Composition and Biological Activity of Essential Oils from Different Species of Piper from Panama. Planta Med. 2016, 82, 986–991. [Google Scholar] [CrossRef]
- Hammer, K.A.; Carson, C.F.; Riley, T.V. Antifungal activity of the components of Melaleuca alternifolia (tea tree) oil. J. Appl. Microbiol. 2003, 95, 853–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman, A.; Tambor, K.; Herman, A. Linalool Affects the Antimicrobial Efficacy of Essential Oils. Curr. Microbiol. 2016, 72, 165–172. [Google Scholar] [CrossRef]
- Park, S.-N.; Lim, Y.K.; Freire, M.O.; Cho, E.; Jin, D.; Kook, J.-K. Antimicrobial effect of linalool and α-terpineol against periodontopathic and cariogenic bacteria. Anaerobe 2012, 18, 369–372. [Google Scholar] [CrossRef]
- Villamizar, L.H.; Cardoso, M.D.G.; Andrade, J.D.; Teixeira, M.L.; Soares, M.J. Linalool, a Piper aduncum essential oil component, has selective activity against Trypanosoma cruzi trypomastigote forms at 4 C. Memórias do Inst. Oswaldo Cruz 2017, 112, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Gil, M.; Bottini, R.; Berli, F.; Pontin, M.; Silva, M.F.; Piccoli, P. Volatile organic compounds characterized from grapevine (Vitis vinifera L. cv. Malbec) berries increase at pre-harvest and in response to UV-B radiation. Phytochemistry 2013, 96, 148–157. [Google Scholar] [CrossRef]
- de Barros, F.M.C.; Zambarda, E.D.O.; Heinzmann, B.M.; Mallmann, C.A. Variabilidade sazonal e biossíntese de terpenóides presentes no óleo essencial de Lippia alba (Mill.) N. E. Brown (Verbenaceae). Química Nova 2009, 32, 861–867. [Google Scholar] [CrossRef]
- Blande, J.D.; Glinwood, R. Deciphering Chemical Language of Plant Communication; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Hieu, L.D.; Thang, T.D.; Hoi, T.M.; Ogunwande, I.A. Chemical Composition of Essential Oils from Four Vietnamese Species of Piper (Piperaceae). J. Oleo Sci. 2014, 63, 211–217. [Google Scholar] [CrossRef] [Green Version]
- Muturi, E.J.; Selling, G.W.; Doll, K.M.; Hay, W.T.; Ramirez, J.L. Leptospermum scoparium essential oil is a promising source of mosquito larvicide and its toxicity is enhanced by a biobased emulsifier. PLoS ONE 2020, 15, e0229076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.-K.; Yusoff, K.; Thomas, W.; Akseer, R.; Alhosani, M.S.; Abushelaibi, A.; Lim, S.-H.-E.; Lai, K.-S. Lavender essential oil induces oxidative stress which modifies the bacterial membrane permeability of carbapenemase producing Klebsiella Pneumoniae. Sci. Rep. 2020, 10, 819. [Google Scholar] [CrossRef] [PubMed]
- Zago, J.A.; Ushimaru, P.I.; Barbosa, L.N.; Fernandes Junior, A. Sinergismo entre óleos essenciais e drogas antimicrobianas sobre linhagens de Staphylococcus aureus e Escherichia coli isoladas de casos clínicos humanos. Rev. Bras. de Farmacogn. 2009, 19, 828–833. [Google Scholar] [CrossRef] [Green Version]
- Sadgrove, N.J.; Padilla-González, G.F.; Phumthum, M. Fundamental Chemistry of Essential Oils and Volatile Organic Compounds, Methods of Analysis and Authentication. Plants 2022, 11, 789. [Google Scholar] [CrossRef] [PubMed]
- Sadgrove, N.J. Purely Australian Essential Oils Past and Present: Chemical Diversity, Authenticity, Bioactivity, and Commercial Value. Diversity 2022, 14, 124. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Padilla-Gonzalez, G.F.; Leuner, O.; Melnikovova, I.; Fernandez-Cusimamani, E. Pharmacology of Natural Volatiles and Essential Oils in Food, Therapy, and Disease Prophylaxis. Front. Pharmacol. 2021, 12, 740302. [Google Scholar] [CrossRef]
- Sadgrove, N.J. Southern Africa as a ‘cradle of incense’ in wider African aromatherapy. Sci. Afr. 2020, 9, e00502. [Google Scholar] [CrossRef]
- Ceole, L.F.; Cardoso, M.D.A.S.G.; Soares, M.J. Nerolidol, the main constituent of Piper aduncum essential oil, has anti-Leishmania braziliensis activity. Parasitology 2017, 144, 1179–1190. [Google Scholar] [CrossRef]
- Marques, A.M.; Barreto, A.L.S.; Batista, E.M.; Curvelo, J.A.D.R.; Velozo, L.S.M.; Moreira, D.D.L.; Guimarães, E.F.; Soares, R.M.A.; Kaplan, M.A.C. Chemistry and Biological Activity of Essential Oils from Piper Claussenianum (Piperaceae). Nat. Prod. Commun. 2010, 5, 1837–1840. [Google Scholar] [CrossRef] [Green Version]
- Chan, W.-K.; Tan, L.T.-H.; Chan, K.-G.; Lee, L.-H.; Goh, B.-H. Nerolidol: A Sesquiterpene Alcohol with Multi-Faceted Pharmacological and Biological Activities. Molecules 2016, 21, 529. [Google Scholar] [CrossRef] [Green Version]
- Curvelo, J.A.R.; Marques, A.M.; Barreto, A.L.S.; Romanos, M.T.V.; Portela, M.B.; Kaplan, M.A.C.; Soares, R.M.A. A novel nerolidol-rich essential oil from Piper claussenianum modulates Candida Albicans biofilm. J. Med. Microbiol. 2014, 63, 697–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, A.M.; Barreto, A.L.S.; Curvelo, J.A.D.R.; Romanos, M.T.V.; Soares, R.M.D.A.; Kaplan, M.A.C. Antileishmanial activity of nerolidol-rich essential oil from Piper claussenianum. Rev. Bras. De Farmacogn. 2011, 21, 908–914. [Google Scholar] [CrossRef]
- Simas, N.K.; Lima, E.d.C.; Conceição, S.D.R.; Kuster, R.M.; Filho, A.M.D.O.; Lage, C.L.S. Produtos naturais para o controle da transmissão da dengue: Atividade larvicida de Myroxylon balsamum (óleo vermelho) e de terpenóides e fenilpropanóides. Química Nova 2004, 27, 46–49. [Google Scholar] [CrossRef] [Green Version]
- Sangwan, N.S.; Farooqi, A.H.A.; Shabih, F.; Sangwan, R.S. Regulation of essential oil production in plants. Plant Growth Regul. 2001, 34, 3–21. [Google Scholar] [CrossRef]
- Baser, K.H.C.; Özek, T.; Kirimer, N.; Tümen, G. A Comparative Study of the Essential Oils of Wild and Cultivated Satureja hor tensis L. J. Essent. Oil Res. 2004, 16, 422–424. [Google Scholar] [CrossRef]
- Gervasi, D.D.L.; Schiestl, F.P. Real-time divergent evolution in plants driven by pollinators. Nat. Commun. 2017, 8, 14691. [Google Scholar] [CrossRef] [Green Version]
- Maeda, H.A.; Fernie, A.R. Evolutionary history of plant metabolism. Annu. Rev. Plant Biol. 2021, 72, 185–216. [Google Scholar] [CrossRef]
- Kessler, A.; Kalske, A. Plant Secondary Metabolite Diversity and Species Interactions. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 115–138. [Google Scholar] [CrossRef]
- Agrawal, A.A.; Hastings, A.P.; Johnson, M.T.J.; Maron, J.L.; Salminen, J.-P. Insect Herbivores Drive Real-Time Ecological and Evolutionary Change in Plant Populations. Science 2012, 338, 113–116. [Google Scholar] [CrossRef]
- Muniz, A.W.; de Sá, E.L.; Dalagnol, G.L.; Filho, J.A. Rooting and acclimatization of micropropagated marubakaido apple rootstock using Adesmia latifolia rhizobia. SpringerPlus 2013, 2, 437. [Google Scholar] [CrossRef] [Green Version]
- Wamser, G.H.; Coimbra, J.L.M.; Guidolin, A.F.; Lannes, S.D.; Dalagnol, G.L. Caracterização de genótipos de cebola com a utilização de marcadores moleculares RAPD. Rev. Ciência Agronômica 2014, 45, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Henderson, I.R.; Jacobsen, S.E. Epigenetic inheritance in plants. Nature 2007, 447, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Miryeganeh, M.; Saze, H. Epigenetic inheritance and plant evolution. Popul. Ecol. 2020, 62, 17–27. [Google Scholar] [CrossRef]
- Bemowska-Kałabun, O.; Panufnik-Mędrzycka, D.; Wierzbicka, M. Evolution caught ‘red-handed’–the transformation of plants in industrial areas (microevolution). In Buckler Mustard (Biscutella laevigata L.) an Extraordinary Plant on Ordinary Mine Heaps near Olkusz; Łukaszewska, R.G.S., Ed.; ZGH Boleslaw: Kraków, Poland, 2020; pp. 117–146. [Google Scholar]
- Preite, V.; Oplaat, C.; Biere, A.; Kirschner, J.; van der Putten, W.H.; Verhoeven, K.J.F. Increased transgenerational epigenetic variation, but not predictable epigenetic variants, after environmental exposure in two apomictic dandelion lineages. Ecol. Evol. 2018, 8, 3047–3059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.-Z.; Li, H.-L.; Li, J.-M.; Yu, F.-H. Correlations between genetic, epigenetic and phenotypic variation of an introduced clonal herb. Heredity 2020, 124, 146–155. [Google Scholar] [CrossRef]
- Zhang, Y.-Y.; Fischer, M.; Colot, V.; Bossdorf, O. Epigenetic variation creates potential for evolution of plant phenotypic plasticity. New Phytol. 2013, 197, 314–322. [Google Scholar] [CrossRef]
- de Arruda, M.R.; Moreira, A.; Pereira, J.C.R. Amostragem e Cuidados na Coleta de Solo para fins de Fertilidade. Embrapa Amazônia Ocidental-Documentos (INFOTECA-E). 2014. Available online: https://www.embrapa.br/busca-de-publicacoes/-/publicacao/1007420/amostragem-e-cuidados-na-coleta-de-solo-para-fins-de-fertilidade (accessed on 26 June 2022).
- Teixeira, P.C.; Donagemma, G.K.; Fontana, A.; Teixeira, W.G. Manual de Métodos de Análise de Solo; Editora Embrapa Solos: Rio de Janeiro, Brazil, 2011. [Google Scholar]
- Wasicky, R. Uma modificação do aparelho de Clevenger para extração de óleos essenciais. Rev. Fac. De Farmácia E Bioquímica 1963, 1, 77–81. [Google Scholar]
- Dool, H.v.D.; Kratz, P.D. A generalization of the retention index system including linear temperature programmed gas—liquid partition chromatography. J. Chromatogr. A 1963, 11, 463–471. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; ISBN 978-1-932633-21-4. [Google Scholar]
- Feng, X.; Zhang, W.; Wu, W.; Bai, R.; Kuang, S.; Shi, B.; Li, D. Chemical composition and diversity of the essential oils of Juniperus rigida along the elevations in Helan and Changbai Mountains and correlation with the soil characteristics. Ind. Crops Prod. 2021, 159, 113032. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| No. a | Compounds b | RIlit | RIcalc | Relative Concentration (%) ± Standard Deviation c | |||||
|---|---|---|---|---|---|---|---|---|---|
| PGW | PGF | PGS | PMW | PMF | PMS | ||||
| 1 | (3H)-Hexanol | 844 | 844 | 0.03 ± 0.01 | |||||
| 2 | α-Pinene # | 932 | 928 | 0.02 ± 0.02 | 15.20 ± 0.03 | 1.07 ± 0.02 | |||
| 3 | Camphene | 946 | 954 | 0.04 ± 0.00 | |||||
| 4 | β-Pinene # | 974 | 979 | 0.31 ± 0.02 | 12.10 ± 1.03 | 0.68 ± 0.08 | |||
| 5 | α-Phellendrene | 1002 | 1000 | 1.19 ± 0.09 | 0.62 ± 0.04 | ||||
| 6 | 1,8-Cineole # | 1026 | 1024 | 34.1 ± 1.54 | 0.83 ± 0.05 | ||||
| 7 | Limonene | 1024 | 1026 | 0.12 ± 0.03 | 0.03 ± 0.02 | 2.14 ± 0.12 | |||
| 8 | Z-β-Ocimene | 1032 | 1035 | 0.19 ± 0.02 | |||||
| 9 | E-β-Ocimene | 1044 | 1048 | 0.09 ± 0.03 | |||||
| 10 | Z-Linalool oxide | 1067 | 1069 | 0.16 ± 0.02 | 18.95 ± 0.74 | 0.34 ± 0.02 | |||
| 11 | E-Linalool oxide | 1084 | 1083 | 1.92 ± 0.08 | |||||
| 12 | α-Terpinolene | 1086 | 1089 | 0.30 ± 0.03 | |||||
| 13 | Linalool # | 1095 | 1094 | 0.02 ± 0.01 | 7.26 ± 0.46 | 37.88 ± 1.01 | 36.99 ± 1.32 | ||
| 14 | E-Pinocarveol | 1135 | 1138 | 0.11 ± 0.03 | |||||
| 15 | Camphor | 1141 | 1143 | 1.23 ± 0.02 | 0.31 ± 0.03 | 0.10 ± 0.01 | |||
| 16 | Camphene hydrate | 1145 | 1152 | 0.16 ± 0.02 | |||||
| 17 | Pinocarvone | 1160 | 1162 | 0.79 ± 0.03 | |||||
| 18 | δ-Terpineol | 1162 | 1164 | 4.69 ± 0.02 | |||||
| 19 | Borneol | 1165 | 1170 | 0.12 ± 0.03 | |||||
| 20 | Terpinen-4-ol | 1174 | 1174 | 0.87 ± 0.03 | |||||
| 21 | α-Terpineol # | 1186 | 1190 | 0.23 ± 0.02 | 4.87 ± 0.02 | 5.62 ± 0.02 | 0.07 ± 0.02 | ||
| 22 | 1-Tridecene | 1290 | 1291 | 0.14 ± 0.03 | |||||
| 23 | 2-Undecanone | 1293 | 1294 | 0.06 ± 0.02 | 0.31 ± 0.03 | 0.32 ± 0.02 | |||
| 24 | δ-Elemene | 1335 | 1337 | 2.31 ± 0.03 | 1.34 ± 0.04 | 1.87 ± 0.33 | 1.07 ± 0.01 | 0.99 ± 0.06 | |
| 25 | Benzyl butanoate | 1343 | 1345 | 1.08 ± 0.03 | |||||
| 26 | α-Cubebene | 1345 | 1352 | 0.32 ± 0.02 | 0.98 ± 0.06 | 1.89 ± 0.02 | 0.09 ± 0.01 | ||
| 27 | α-Ylangene | 1373 | 1374 | 0.78 ± 0.05 | 0.10 ± 0.02 | ||||
| 28 | α-Copaene | 1374 | 1376 | 1.23 ± 0.05 | 1.45 ± 0.03 | 1.67 ± 0.07 | 0.17 ± 0.01 | ||
| 29 | β-Bourbonene | 1387 | 1383 | 0.32 ± 0.02 | 0.89 ± 0.08 | 1.80 ± 0.34 | |||
| 30 | β-Elemene | 1389 | 1388 | 1.23 ± 0.04 | 1.37 ± 0.03 | 1.67 ± 0.04 | 2.13 ± 0.03 | 1.61 ± 0.05 | 1.15 ± 0.13 |
| 31 | α-Gurjunene | 1409 | 1409 | 1.45 ± 0.06 | 0.32 ± 0.07 | ||||
| 32 | E-Caryophyllene # | 1417 | 1418 | 2.34 ± 0.04 | 5.43 ± 0.06 | 8.43 ± 0.07 | 2.44 ± 0.22 | 2.49 ± 0.55 | |
| 33 | β-Gurjunene | 1431 | 1435 | 0.45 ± 0.02 | 0.23 ± 0.02 | 0.21 ± 0.03 | 1.47 ± 0.22 | ||
| 34 | γ-Elemene | 1434 | 1438 | 0.78 ± 0.06 | 0.03 ± 0.01 | 0.08 ± 0.03 | |||
| 35 | α-Guaiene | 1437 | 1439 | 0.40 ± 0.06 | |||||
| 36 | Aromadendrene | 1439 | 1441 | 1.23 ± 0.07 | 1.04 ± 0.03 | 1.65 ± 0.76 | 0.17 ± 0.03 | 0.79 ± 0.02 | |
| 37 | Z-β-Farnesene | 1440 | 1442 | 0.57 ± 0.02 | 3.43 ± 0.06 | ||||
| 38 | Z-Muurola-3,5-diene | 1448 | 1450 | 0.24 ± 0.03 | 0.89 ± 0.06 | ||||
| 39 | α-Humulene | 1452 | 1453 | 1.21 ± 0.06 | 0.32 ± 0.02 | 1.22 ± 0.03 | 2.22 ± 0.12 | 2.47 ± 0.04 | |
| 40 | E-β-Farnesene | 1454 | 1454 | 1.73 ± 0.08 | |||||
| 41 | E-Muurola-3,5-diene | 1454 | 1455 | 0.21 ± 0.05 | |||||
| 42 | β-Santalene | 1457 | 1459 | 0.34 ± 0.01 | |||||
| 43 | Allo-Aromadendrene | 1458 | 1461 | 2.34 ± 0.04 | 0.07 ± 0.01 | 0.50 ± 0.04 | 0.54 ± 0.02 | ||
| 44 | 9-epi-E-Caryophyllene | 1464 | 1468 | 1.23 ± 0.03 | 0.43 ± 0.02 | 0.33 ± 0.02 | |||
| 45 | γ-Muurolene | 1478 | 1477 | 0.23 ± 0.00 | 1.21 ± 0.00 | ||||
| 46 | Amorpha-4,7(11)-diene | 1479 | 1478 | 0.23 ± 0.02 | |||||
| 47 | Ar-Curcumene | 1479 | 1480 | 0.08 ± 0.02 | 0.14 ± 0.01 | 0.18 ± 0.02 | 0.37 ± 0.03 | ||
| 48 | α-Amorphene | 1483 | 1483 | 5.21 ± 0.04 | 1.43 ± 0.34 | 0.76 ± 0.04 | 0.20 ± 0.01 | ||
| 49 | Germacrene D | 1484 | 1484 | 0.05 ± 0.01 | 0.32 ± 0.03 | 0.57 ± 0.04 | 0.60 ± 0.09 | ||
| 50 | Z-Eudesma-6,11-diene | 1489 | 1489 | 4.32 ± 0.35 | 2.31 ± 0.12 | 3.34 ± 0.31 | |||
| 51 | β-Selinene | 1489 | 1490 | 3.45 ± 0.06 | 1.23 ± 0.03 | 1.87 ± 0.02 | 0.14 ± 0.03 | 0.72 ± 0.04 | 0.54 ± 0.01 |
| 52 | δ-Selinene | 1492 | 1492 | 0.14 ± 0.04 | 0.52 ± 0.05 | ||||
| 53 | E-Muurola-4(14),5-diene | 1493 | 1493 | ||||||
| 54 | γ-Amorphene | 1495 | 1496 | 4.21 ± 0.00 | |||||
| 55 | α-Selinene | 1498 | 1497 | 4.87 ± 0.00 | 3.25 ± 0.00 | 1.01 ± 0.00 | |||
| 56 | 2-Tridecanone | 1495 | 1497 | 0.29 ± 0.02 | |||||
| 57 | Bicyclogermacrene # | 1500 | 1499 | 14.23 ± 0.0 | 16.12 ± 0.00 | 28.16 ± 0.00 | 0.81 ± 0.06 | 1.34 ± 0.05 | 1.30 ± 0.03 |
| 58 | α-Muurolene | 1500 | 1501 | 0.76 ± 0.00 | 1.23 ± 0.00 | 0.06 ± 0.01 | |||
| 59 | E-β-Guaiene | 1502 | 1503 | 0.20 ± 0.02 | |||||
| 60 | E,E-α-Farnesene | 1505 | 1505 | 0.49 ± 0.06 | 0.43 ± 0.02 | ||||
| 61 | Cubebol | 1515 | 1514 | 0.18 ± 0.06 | 0.38 ± 0.02 | ||||
| 62 | γ-Cadinene | 1513 | 1515 | 1.23 ± 0.11 | 2.32 ± 0.27 | 0.52 ± 0.02 | 0.23 ± 0.01 | 0.39 ± 0.01 | |
| 63 | 7-epi-α-Selinene | 1520 | 1521 | 1.23 ± 0.05 | 1.23 ± 0.04 | ||||
| 64 | δ-Cadinene | 1522 | 1522 | 5.67 ± 0.08 | 3.56 ± 0.05 | 0.18 ± 0.03 | 0.81 ± 0.08 | 1.36 ± 0.07 | |
| 65 | Z-Calamenene | 1528 | 1527 | 0.21 ± 0.03 | |||||
| 66 | Zonarene | 1528 | 1530 | 0.24 ± 0.04 | |||||
| 67 | E-γ-Bisabolene | 1529 | 1532 | 0.03 ± 0.02 | |||||
| 68 | E-Cadina-1,4-diene | 1533 | 1534 | 1.87 ± 0.08 | 0.67 ± 0.02 | 0.31 ± 0.04 | |||
| 69 | α-Cadinene | 1537 | 1537 | 1.98 ± 0.06 | 0.89 ± 0.04 | ||||
| 70 | Selina-3,7(11)-diene | 1545 | 1546 | 1.45 ± 0.08 | 1.94 ± 0.09 | 2.31 ± 0.06 | 0.18 ± 0.02 | ||
| 71 | Elemol | 1548 | 1551 | 0.08 ± 0.02 | 0.38 ± 0.10 | ||||
| 72 | Germacrene B | 1559 | 1558 | 1.23 ± 0.04 | 2.23 ± 0.05 | 0.12 ± 0.02 | |||
| 73 | E-Nerolidol | 1561 | 1563 | 12.11 ± 1.0 | 6.32 ± 0.21 | 8.03 ± 0.09 | 2.12 ± 0.06 | 2.14 ± 0.09 | 11.39 ± 1.04 |
| 74 | β-Calacorene | 1564 | 1566 | 0.09 ± 0.02 | |||||
| 75 | Palustrol | 1567 | 1568 | 0.09 ± 0.02 | 0.22 ± 0.03 | 0.31 ± 0.02 | |||
| 76 | Spathulenol | 1577 | 1573 | 5.32 ± 0.02 | 3.32 ± 0.02 | 0.82 ± 0.04 | 1.13 ± 0.04 | ||
| 77 | Caryophyllene oxide | 1582 | 1581 | 1.23 ± 0.02 | 1.14 ± 0.12 | 2.07 ± 0.02 | |||
| 78 | Globulol | 1590 | 1585 | 0.50 ± 0.02 | 1.94 ± 0.01 | ||||
| 79 | Gleenol | 1586 | 1586 | 0.36 ± 0.02 | |||||
| 80 | Viridiflorol | 1592 | 1594 | 5.43 ± 0.05 | 7.89 ± 0.56 | 5.21 ± 0.40 | 0.10 ± 0.01 | 0.57 ± 0.05 | 1.17 ± 0.07 |
| 81 | Guaiol | 1600 | 1602 | 0.35 ± 0.04 | |||||
| 82 | Ledol | 1602 | 1604 | 5.08 ± 0.02 | 5.54 ± 0.07 | ||||
| 83 | 5-epi-7-epi-α-Eudesmol | 1607 | 1607 | 0.32 ± 0.01 | |||||
| 84 | Humulene epoxide II | 1608 | 1607 | 2.08 ± 0.03 | |||||
| 85 | 2,(7Z) -Bisaboladien-4-ol | 1618 | 1618 | 0.21 ± 0.03 | |||||
| 86 | 10-epi-γ-Eudesmol | 1622 | 1622 | 0.39 ± 0.02 | |||||
| 87 | E-Isolongifolanone | 1625 | 1625 | 0.18 ± 0.03 | |||||
| 88 | 1-epi-Cubenol | 1627 | 1629 | 0.06 ± 0.02 | 0.48 ± 0.02 | 1.20 ± 0.01 | |||
| 89 | E-Sesquilavandulol | 1631 | 1630 | 1.11 ± 0.03 | |||||
| 90 | γ-Eudesmol | 1630 | 1631 | 0.28 ± 0.02 | 1.68 ± 0.16 | ||||
| 91 | Eremoligenol | 1629 | 1632 | 0.04 ± 0.03 | |||||
| 92 | epi-α-Cadinol | 1638 | 1636 | 3.65 ± 0.19 | |||||
| 93 | Z-Cadin-4-en-7-ol | 1635 | 1637 | 0.62 ± 0.08 | |||||
| 94 | Caryophylla-4(12),8(13)-dien-5α-ol | 1639 | 1637 | 0.32 ± 0.02 | 0.50 ± 0.08 | ||||
| 95 | epi-α-Muurolol | 1640 | 1641 | 1.99 ± 0.09 | 1.03 ± 0.09 | ||||
| 96 | α -Muurolol | 1644 | 1645 | 3.42 ± 0.04 | 0.98 ± 0.00 | 0.28 ± 0.03 | 0.60 ± 0.19 | 3.66 ± 0.02 | |
| 97 | α-Eudesmol | 1652 | 1652 | 1.21 ± 0.05 | 0.56 ± 0.04 | ||||
| 98 | α-Cadinol | 1652 | 1653 | 3.21 ± 0.12 | 9.32 ± 1.12 | 7.32 ± 0.68 | 0.20 ± 0.01 | 2.22 ± 0.07 | |
| 99 | neo-Intermedeol | 1658 | 1658 | 0.08 ± 0.01 | |||||
| 100 | Selin-11-en-4-α-ol | 1658 | 1660 | 0.26 ± 0.03 | |||||
| 101 | 7-epi-α-Eudesmol | 1662 | 1663 | 0.32 ± 0.02 | 0.09 ± 0.01 | 0.34 ± 0.04 | |||
| 102 | Intermedeol | 1665 | 1667 | 1.34 ± 0.01 | 2.32 ± 0.03 | 0.22 ± 0.03 | |||
| 103 | 14-hydroxy-9-epi-E-Caryophyllene | 1668 | 1672 | 0.63 ± 0.01 | 1.14 ± 0.08 | ||||
| 104 | β-Bisabolol | 1674 | 1677 | 0.24 ± 0.03 | |||||
| 105 | α-Bisabolol | 1685 | 1684 | 0.07 ± 0.01 | |||||
| 106 | Eudesm-7(11)-en-4-ol | 1700 | 1699 | 0.76 ± 0.06 | |||||
| 107 | Benzyl benzoate # | 1759 | 1762 | 0.11 ± 0.01 | 1.05 ± 0.09 | 5.69 ± 0.09 | |||
| Monoterpene hydrocarbons | 0.06 | 0.43 | 0.03 | 31.10 | 2.39 | 0.00 | |||
| Oxygenated monoterpenes | 0.37 | 6.10 | 5.93 | 48.30 | 59.55 | 37.33 | |||
| Sesquiterpene hydrocarbons | 65.73 | 53.46 | 65.07 | 13.00 | 13.17 | 12.37 | |||
| Oxygenated sesquiterpenes | 31.81 | 37.91 | 26.20 | 5.57 | 15.45 | 32.88 | |||
| Other compounds | 0.03 | 0.00 | 0.00 | 0.31 | 1.36 | 7.38 | |||
| Identified Compounds in Numbers | 43 | 44 | 28 | 63 | 40 | 37 | |||
| Total compounds (%) | 98.01 | 97.90 | 97.23 | 98.34 | 92.32 | 89.96 | |||
| Oil yielding (%) | 0.17 ± 0.03 | 0.13 ± 0.02 | 0.18 ± 0.02 | 0.86 ± 0.08 | 0.12 ± 0.03 | 0.17 ± 0.06 | |||
| Shannon index | 3.81 | 3.78 | 3.30 | 4.14 | 3.86 | 3.67 | |||
| ‘Ramos and Moreira Index (GMRO) | −3.39 | −3.46 | −6.94 | −2.55 | −3.49 | −3.73 | |||
| Soil Attributes | PM | PG | THT |
|---|---|---|---|
| pH in Water | 4.90 ± 0.95 | 5.40 ± 0.09 | 5.80 ± 0.18 |
| Total acidity (cmolc/dm3) | 11.88 ± 2,63 | 12.15 ± 0.89 | 8.91 ± 0.24 |
| Al (cmolc/dm3) | 0.10 ± 0.05 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Ca (cmolc/dm3) | 2.30 ± 0.09 | 2.00 ± 0.18 | 14.40 ± 5.89 |
| Mg (cmolc/dm3) | 1.30 ± 0.09 | 1.60 ± 0.08 | 6.90 ± 0.08 |
| Na (mg/dm3) | 18.40 ± 0.12 | 11.50 ± 1.70 | 27.60 ± 3.75 |
| K (mg/dm3) | 276.90 ± 32.45 | 202.90 ± 25.12 | 557.70 ± 41.04 |
| P (mg/dm3) | 7.54 ± 1.09 | 6.31 ± 1.07 | 25.33 ± 3.05 |
| C (g/kg) | 44.00 ± 6.31 | 67.00 ± 9.78 | 107.50 ± 13.43 |
| N (g/kg) | 4.30 ± 0.19 | 3.80 ± 0.13 | 4.20 ± 0.29 |
| Cu (mg/dm3) | 2.04 ± 0.04 | 2.77 ± 0.06 | 0.26 ± 0.08 |
| Fe (mg/dm3) | 29.70 ± 9.21 | 49.50 ± 8.45 | 23.60 ± 3.23 |
| Mn (mg/dm3) | 83.50 ± 4.32 | 82.40 ± 3.07 | 13.70 ± 1.45 |
| Zn (mg/dm3) | 4.43 ± 0.03 | 3.76 ± 0.04 | 3.46 ± 0.05 |
| Value S (cmolc/dm3) | 8.05 ± 0.032 | 7.15 ± 0.06 | 4.15 ± 0.32 |
| Value T (cmolc/dm3) | 19.93 ± 0.98 | 16.30 ± 0.21 | 6.30 ± 0.78 |
| Value V (%) | 40.39 ± 6.03 | 46.31 ± 4.20 | 65.93 ± 2.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos, Y.J.; Felisberto, J.S.; Gouvêa-Silva, J.G.; de Souza, U.C.; da Costa-Oliveira, C.; de Queiroz, G.A.; Guimarães, E.F.; Sadgrove, N.J.; de Lima Moreira, D. Phenoplasticity of Essential Oils from Two Species of Piper (Piperaceae): Comparing Wild Specimens and Bi-Generational Monoclonal Cultivars. Plants 2022, 11, 1771. https://doi.org/10.3390/plants11131771
Ramos YJ, Felisberto JS, Gouvêa-Silva JG, de Souza UC, da Costa-Oliveira C, de Queiroz GA, Guimarães EF, Sadgrove NJ, de Lima Moreira D. Phenoplasticity of Essential Oils from Two Species of Piper (Piperaceae): Comparing Wild Specimens and Bi-Generational Monoclonal Cultivars. Plants. 2022; 11(13):1771. https://doi.org/10.3390/plants11131771
Chicago/Turabian StyleRamos, Ygor Jessé, Jéssica Sales Felisberto, João Gabriel Gouvêa-Silva, Ulisses Carvalho de Souza, Claudete da Costa-Oliveira, George Azevedo de Queiroz, Elsie Franklin Guimarães, Nicholas John Sadgrove, and Davyson de Lima Moreira. 2022. "Phenoplasticity of Essential Oils from Two Species of Piper (Piperaceae): Comparing Wild Specimens and Bi-Generational Monoclonal Cultivars" Plants 11, no. 13: 1771. https://doi.org/10.3390/plants11131771
APA StyleRamos, Y. J., Felisberto, J. S., Gouvêa-Silva, J. G., de Souza, U. C., da Costa-Oliveira, C., de Queiroz, G. A., Guimarães, E. F., Sadgrove, N. J., & de Lima Moreira, D. (2022). Phenoplasticity of Essential Oils from Two Species of Piper (Piperaceae): Comparing Wild Specimens and Bi-Generational Monoclonal Cultivars. Plants, 11(13), 1771. https://doi.org/10.3390/plants11131771