Abstract
Soil salinity is a problem that affects soil fertility and threatens agri-food crop production worldwide. Biotechnology, through plant micropropagation and the use of biofertilizers such as arbuscular mycorrhizal fungi (AMF), is an alternative to increase productivity and induce tolerance to salinity stress in different crops. This study aimed to evaluate the effect of different doses of the fungus Glomus intraradices on the ex vitro development of taro (Colocasia esculenta L. Schott cv. Criolla) plantlets under salinity stress during the acclimatization stage. In vitro-obtained C. esculenta plantlets were inoculated at different doses (0, 100, and 200 spores per plantlet) of G. intraradices during acclimatization. At 60 d of acclimatization in the greenhouse, plantlets were exposed to 100 mM NaCl salinity stress for 10 d. After the stress period, plantlet development, colonization percentage, and biomass were evaluated. In addition, the content of chlorophyll, carotenoids, proteins, proline, glycine-betaine, soluble phenols, and antioxidant capacity were quantified. The results showed differences in the developmental, physiological, and biochemical variables evaluated; however, no changes in total protein content were observed. Spore colonization showed that the symbiotic association has positive effects on the development of plantlets with or without salinity stress. This symbiotic interaction contributes to salinity stress tolerance in C. esculenta plantlets. The early application of AMF in in vitro-obtained taro plantlets is an alternative to increase or maintain the productivity of this crop in saline soils.
1. Introduction
Taro (Colocasia esculenta L. Schott), a member of the plant family Araceae, is cultivated due to its importance in the food industry as a result of its high content of starch, proteins, vitamins, polysaccharides, and various trace elements [1,2]. It is also sought by the pharmaceutical industry for its secondary metabolites with antitumor, antimetastatic, antioxidant, and anti-inflammatory properties [3], and by the biorefinery industry for use in bioethanol production [4]. The uses and benefits of C. esculenta make it a genetic resource with high commercial value. However, taro cultivation faces great challenges in terms of productivity due to little renewal of certified plant material and the abiotic stress caused by an excess accumulation of salts in the soil. Salinity affects soil fertility with effects on low crop yields [5]. Soil salinity, both of natural origin (primary salinization) and as a consequence of anthropogenic activities (secondary salinity), is a threat to agriculture and a constraint on agri-food crop production. The main factors contributing to the increase in salinity are low precipitation, high surface evaporation, use of wastewater for irrigation, groundwater depletion, and poor agricultural practices caused by the excessive use of chemical fertilizers such as carbonates, sulfates, sodium nitrates and chlorides, potassium, and magnesium, among others [6,7], coupled with a lack of leaching practices.
Plant and microbial biotechnology offer alternatives to increase crop productivity and mitigate the effects of abiotic stress on plants. One option is micropropagation using plant tissue culture (PTC) techniques to obtain certified plants with high genetic and phytosanitary quality [8], while another is the use of microorganisms such as mycorrhizae that can establish symbiotic associations with plants to stimulate greater development and increase tolerance to different types of abiotic stress [9]. Acclimatization is the final stage of micropropagation and consists of the gradual transfer of in vitro-grown plantlets to ex vitro conditions under a greenhouse [10]. In addition, plantlet acclimatization is a process that allows early inoculation of microorganisms prior to transplanting in the field for seedbed establishment. Mycorrhizae play an important role in the development and survival of their constituent species (plants and fungi) [11,12]. Arbuscular mycorrhizal fungi (AMF) are obligate biotrophs that establish endosymbiotic associations with the roots of vascular plants [13,14]. AMF enhance mineral nutrient uptake in the host plant [13,15], have positive effects on photosynthetic activity [16], decrease disease invasion [17], and confer tolerance to abiotic stresses caused by abrupt temperature changes [18], heavy metals [19], and drought and salinity stress. The positive effects of AMF on plants are the production of antioxidants and polyamines, osmotic adjustment, improved water transport in plants exposed to salinity, and maintenance of an ionic balance in the Na+/K+ ratio through the external mycelium to enhance plant development and survival under salt stress [20,21,22,23]. The use of mycorrhizae to improve salinity tolerance in plants has been evaluated in Rhizophagus irregularis in swamp she-oak (Casuarina glauca) [24], Funneliformis mosseae in Chinese honey locust (Gleditsia sinensis) [25], Glomus mosseae in wheat (Triticum aestivum L.) [26], and Rhizophagus irregularis in maize (Zea mays L.) [27]. This study aimed to evaluate the effect of different doses of Glomus intraradices spores during salinity stress induced during ex vitro development of C. esculenta plantlets at the acclimatization stage.
2. Results and Discussion
2.1. Colonization Percentage, Plantlet Development, and Biomass
The different AMF application doses and induced salinity stress had an effect on the colonization percentage and developmental and biomass variables evaluated (Table 1). Regarding colonization, the highest colonization percentages were observed in plantlets inoculated with mycorrhizae without salinity stress conditions, followed by plantlets with mycorrhizae without salinity stress conditions, followed by plantlets with mycorrhizae + NaCl, whereas the lowest colonization percentage was observed in plantlets without mycorrhizae. The largest plantlets were observed at doses of 100 and 200 spores per plant without salinity stress application, whereas the smallest plantlets were obtained under salinity stress without mycorrhizae. For the number of leaves per plantlet, the highest number of leaves was observed in the mycorrhizal treatments, whereas the lowest number of leaves per plantlet was observed in the treatments without mycorrhizae. The highest percentage of senescent leaves was found in plantlets under salinity stress and without mycorrhizae, whereas the lowest percentage of senescent leaves was observed in the treatments without salinity stress, with or without mycorrhizae. For the variable number of roots, the highest number of roots was found at doses of 100 and 200 spores per plantlet without salinity stress, while the lowest number of roots was observed in the treatment without mycorrhizae and without salinity stress, and in the treatment with salinity stress without mycorrhizae. On the other hand, the greatest root length was observed at doses of 100 and 200 spores per plantlet, whereas the smallest roots were observed under salinity stress and without mycorrhizae (Figure 1). Regarding the biomass variables, the highest fresh weight was observed at doses of 100 and 200 spores with salinity stress, whereas the lowest fresh weight was observed in the treatment without mycorrhizae and without salinity stress, and in the treatment with salinity stress without mycorrhizae. The highest dry weight was observed in all mycorrhizal treatments, with or without salinity stress, whereas the lowest dry weight content was observed in the treatment without mycorrhizae and without salinity stress. For the dry matter (DM) percentage, the highest percentage was observed at doses of 100 and 200 spores per plantlet under salinity stress, whereas the lowest percentage was observed in the treatment without mycorrhizae and without salinity stress.

Table 1.
Effect of sodium chloride (NaCl) and arbuscular mycorrhizal fungi (Glomus intraradices) on development of taro (Colocasia esculenta L. Schott) during acclimatization stage.

Figure 1.
Effect of sodium chloride (NaCl) and arbuscular mycorrhizal fungi (Glomus intraradices) on ex vitro development of taro (Colocasia esculenta L. Schott) during acclimatization stage. (a) Non-mycorrhizal plantlets, (b) 100 spores per plantlet, (c) 200 spores per plantlet, (d) non-mycorrhizal plantlets + 100 mM NaCl, (e) 100 spores per plantlet + 100 mM NaCl, and (f) 200 spores per plantlet + 100 mM NaCl. White bars = 5 cm.
The results obtained in this study demonstrate the effect of AMF with or without salinity stress on colonization percentage and plantlet development and biomass variables. The AMF doses evaluated had a positive effect on plantlet development and tolerance to salinity stress. This effect could be due to the fact that mycorrhizal colonization percentages allow an adequate symbiotic interaction that could favor some physiological processes for the plant such as photosynthetic efficiency and water and nutrient uptake, keeping a high K+/Na+ ratio and higher osmoprotectants and enzyme activities [28].
Increased development and biomass in plants inoculated with mycorrhizae have been reported in melon (Cucumis melo) [29], watermelon (Citrullus lanatus L. cv. Qilin) [17] and sugarcane (Saccharum spp. Hybrids) [30]. Meddich et al. [29] observed in C. melo that when using a mycorrhizal consortium with Glomus sp. at a dose of 44 spores per plant, shoot and root biomass increased significantly. Wu et al. [17] observed in C. lanatus that colonization with mycorrhizae at a dose of 300 spores per plant promoted greater biomass accumulation. Sales et al. [30] in Saccharum spp. obtained an increase in yield when inoculating with native AMF at a dose of 260 spores per plant. In this study, mycorrhizal treatments with or without salinity stress increased plantlet length, number of leaves, number of roots, root length, fresh weight, and dry weight compared to treatments without mycorrhizae. On the other hand, plantlets that were not inoculated with mycorrhizae had a lower rate of development; however, none of the plantlets died at a dose of 100 mM NaCl. The zero-mortality rate in plantlets was probably due to Na+ accumulation in microcorm tissues; however, leaf senescence was observed in NaCl-treated plantlets. In our study, senescence in NaCl treatments could be due to older leaves showing the effects of salinity stress toxicity first, probably because they have transpired longer and therefore accumulated more Na+ in their tissues during the stress period causing leaf death. Lloyd et al. [31] found that C. esculenta was able to continue to grow and exclude sodium from transpiring leaves with 100 and 200 mM NaCl. In addition, an increase in senescent leaves and marginal leaf chlorosis were observed in salt treatments.
In crops such as licorice (Glycyrrhiza glabra) [32], wheat (Triticum aestivum) [33], pistachio (Pistacia vera L. cv. Ohadi) [34], Saudi pearl millet (Pennisetum spicatum) [35], and maize (Zea mays L.) [27], an increase in the developmental and biomass variables of plants inoculated with mycorrhizae under salinity stress has been observed. The effects of mycorrhizae on development depend on the type and dose of AMF in relation to the host plant species. Amanifar et al. [32] in G. glabra observed that by using Funneliformis mosseae at a dose of 840 spores per plant, root length increased significantly under salinity stress for 40 days. Abbaspour et al. [34] in P. vera cv. Ohadi obtained greater plant length and an increase in stem diameter when inoculating with 1000 spores per plant with the fungus Rhizophagus irregularis Becker and Gerdemann (Gec) at concentrations of 0 and 250 mM NaCl for seven days. Chen et al. [27] observed that colonization with Rhizophagus irregularis at a dose of 1800 spores per plant and a concentration of 100 mM NaCl promoted greater biomass accumulation in Z. mays plantlets.
In this study, the increase in dry matter in the AMF treatments could be due to the fact that G. intraradices is an endomycorrhizal fungus. This type of fungus inhabits the cells and tissues of the plant and could cause a relative increase in root dry matter compared to the control treatment, without the presence of colonies in the roots. According to the results obtained, the increase in developmental variables in mycorrhized taro plantlets could be due to better water absorption through the external mycelium and a reduction in the accumulation of Na+ and Cl− in the aerial parts of the plants. In addition, the increased growth of plantlets with AMF + NaCl is probably due not only to ion storage in the microcosms, but also to the fungus.
2.2. Total Chlorophyll and Carotenoid Content
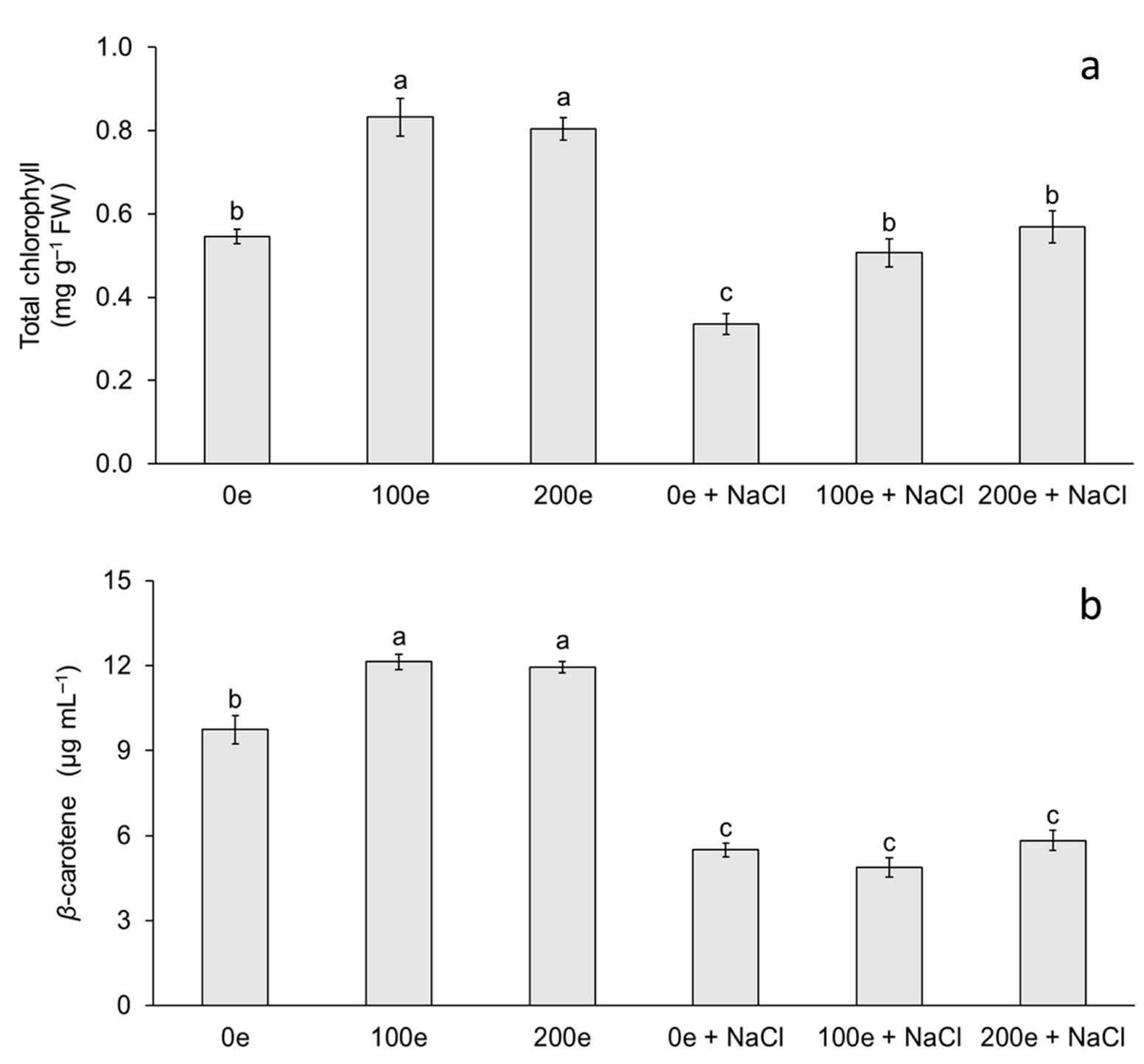
The different AMF doses and induced salinity stress had an effect on total chlorophyll and carotenoid content (Figure 2). The highest chlorophyll content was observed in the treatments with 100 and 200 spores per plantlet without salinity stress, whereas the lowest chlorophyll contents were observed when neither mycorrhizae nor salinity stress were applied, and at doses of 100 and 200 spores per plantlet under salinity stress (Figure 2a). For carotenoid content, the highest level was observed in the treatments with 100 and 200 spores per plantlet without salinity stress, whereas the lowest level was observed when mycorrhizae were applied with or without induced salinity stress (Figure 2b).
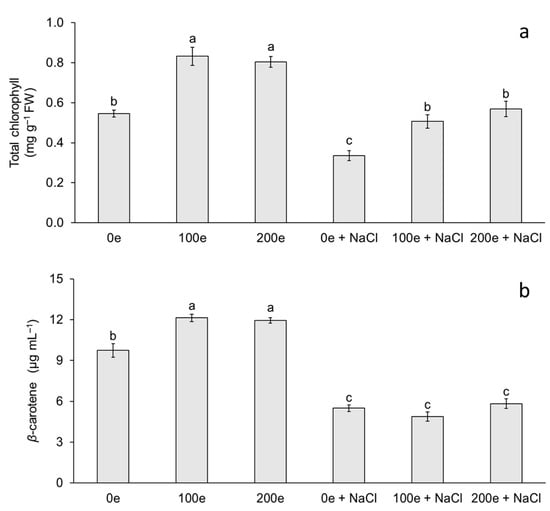
Figure 2.
Effect of sodium chloride (NaCl) and arbuscular mycorrhizal fungi (Glomus intraradices) on total chlorophyll and β-carotene content in taro plantlets (Colocasia esculenta L. Schott) during acclimatization stage. (a) Total chlorophyll and (b) β-carotene. Bars represent mean ± standard error. Means with a different letter are significantly different (Tukey, p < 0.05). e = spores per plantlet and NaCl is expressed in 100 mM.
The results obtained in this study demonstrate the effect of different treatments with mycorrhizae and NaCl on chlorophyll and carotenoid content in taro plantlets. Chlorophyll and carotenoid accumulation in plants inoculated with mycorrhizae has been reported in several species such as tomato (Lycopersicon esculentum) [14], sugarcane (Saccharum spp.) [36], rice (Oryza sativa) [37], and melon (Cucumis melo) [29]. Di Martino et al. [14] in L. esculentum found that mycorrhizal colonization with Glomus mosseae increases the concentration of photosynthetic pigments. Meddich et al. [29] in C. melo observed that when using a consortium with Glomus sp. at a dose of 44 spores per plant, photosynthetic pigments increased significantly. In this study, total chlorophyll content increased when plantlets were inoculated with mycorrhizae (with or without NaCl); however, plants that were treated with mycorrhizae + NaCl showed an increase in chlorophyll content compared to NaCl treatment without mycorrhizae. Chlorophyll and carotenoid accumulation in plants inoculated with mycorrhizae under salinity stress has been reported in licorice (Glycyrrhiza glabra) [32], wheat (Triticum aestivum) [33], Russian olive (Elaeagnus angustifolia) [38], Saudi pearl millet (Pennisetum spicatum) [35], and eucalyptus (Eucalyptus camaldulensis) [22]. Amanifar et al. [32] in G. glabra, using Funneliformis mosseae at 840 spores per plant, found that the relative leaf chlorophyll index increased significantly in salinity stress at a concentration of 40 mM NaCl for 40 days. Liang et al. [38] observed in E. angustifolia that total chlorophyll content increased when using Rhizophagus irregularis at a dose of 1500 spores per plant under salinity stress with 300 mM NaCl for 21 days. Klinsukon et al. [22] in E. camaldulensis observed that total chlorophyll content increased when using a consortium of mycorrhizae (Glomus sp., Gigaspora albida and Gigaspora decipiens) at a dose of 960 spores per plant under salinity stress at a concentration of 100 mM NaCl for 90 days. Variation in chlorophyll content is an important factor that indirectly determines the photosynthetic capacity of plants [39]. A low photosynthetic capacity can disrupt carbon stabilization and eventually decrease the development of plants under stress [39,40]. Increased chlorophyll content in plantlets under salinity stress and with mycorrhizae could be a mechanism to maintain photosynthetic metabolism associated with salinity tolerance. Hashem et al. [41] and Bouskout et al. [42] note that mycorrhizae generate greater availability of N and cofactors involved in chlorophyll synthesis such as Mn, Mg, and Fe. The latter are essential components of the electron carriers in the photosynthetic apparatus, improve stomatal conductance, and consequently increase CO2 assimilation.
In relation to carotenoids, taro plantlets showed a decrease at different doses of mycorrhizae with NaCl (with or without mycorrhizae), with carotenoid content being higher when plantlets were treated with mycorrhizae without NaCl. Ye et al. [28] observed in C. lanatus that carotenoid content increased when using Funneliformis mosseae at a dose of 700 spores per plantlet and a period of salinity stress with 60 mM NaCl for 6 days. Ndiate et al. [26] observed in T. aestivum that carotenoid content increased when using Glomus mosseae at a dose of 160 spores per plant in combination with a biochar amendment during a period of salinity stress with 50–150 mM NaCl for 90 days. Therefore, the increase in these antioxidant compounds could decrease the photodegradation and photoinhibition of these photosynthetic pigments by acting as a salinity stress tolerance mechanism [42]. According to Sun et al. [43], carotenoids function as light scavengers for photosynthesis, absorbing light energy and transferring it to chlorophylls in an absorption range of 450–550 nm. Carotenoid levels could be explained by several factors: changes in enzymatic activities, damaged photosynthetic apparatus, inhibition of the electron transport chain, and changes in gene expression involved in the biosynthetic pathway [44]. In this study, the reduction in carotenoid content was probably due to a decrease in their biosynthesis and/or a degradation caused by NaCl-induced salinity stress.
2.3. Protein, Proline, and Glycine-Betaine Content
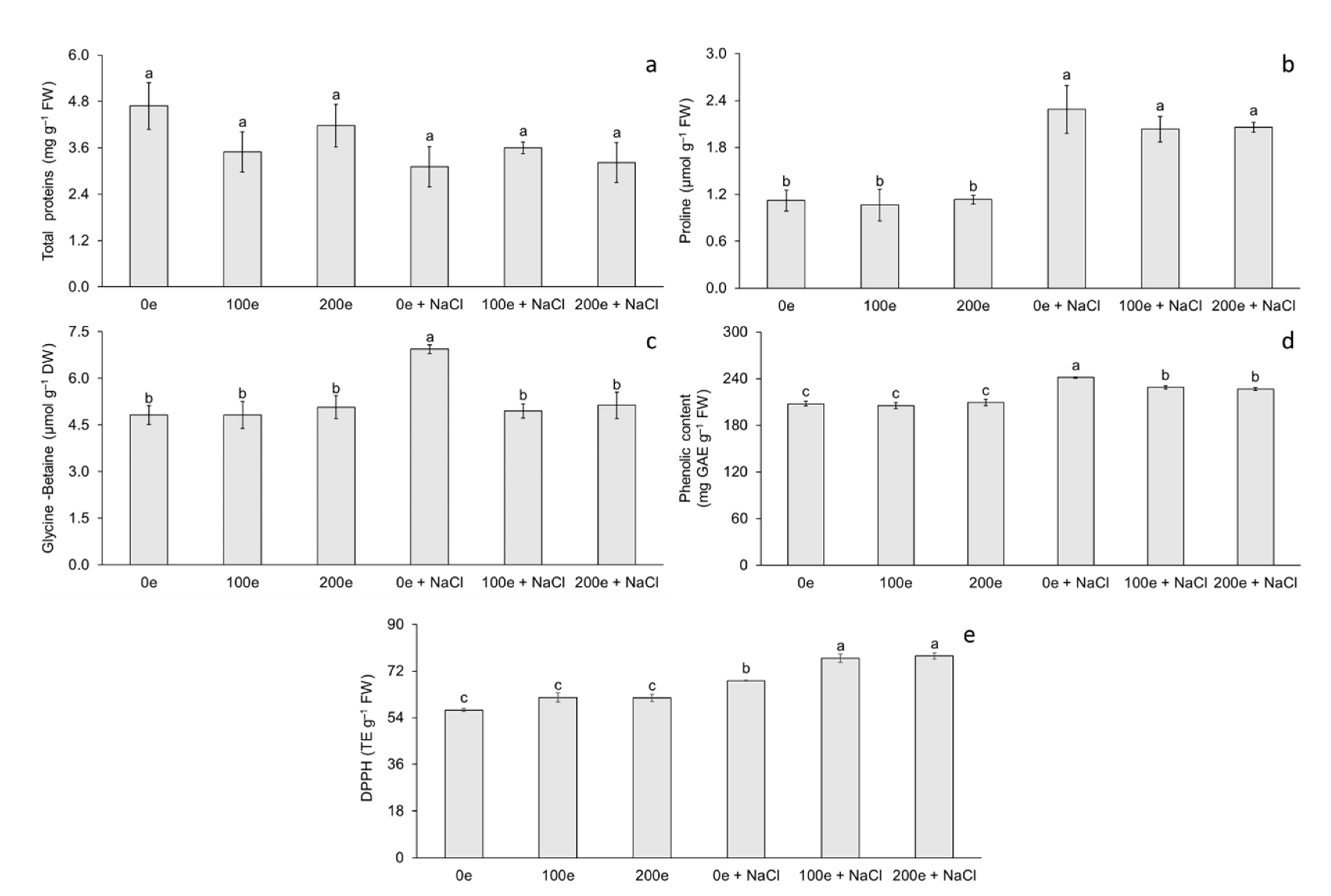
The different AMF doses and induced salinity stress had no effect on total protein (TP) content; however, differences were observed for proline (Pr) and glycine-betaine (GB) content (Figure 3). The highest proline content was observed when mycorrhizae were applied with or without induced salinity stress, whereas the lowest proline content was observed when salinity stress was not applied, with or without mycorrhizae (Figure 3b). Regarding glycine-betaine, the highest content was observed in plantlets under salinity stress without mycorrhizae, whereas the lowest content was observed in the rest of the treatments (Figure 3c).
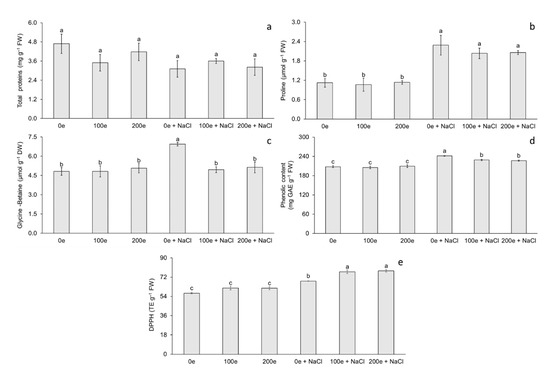
Figure 3.
Effect of sodium chloride (NaCl) and arbuscular mycorrhizal fungi (Glomus intraradices) on total proteins, proline, and glycine-betaine content in taro plantlets (Colocasia esculenta L. Schott) during acclimatization stage. (a) Total proteins, (b) proline, (c) glycine-betaine, (d) phenolic content expressed in GAE (milligrams of gallic acid equivalents per g of fresh weight) and (e) antioxidant capacity expressed in DPPH (trolox equivalents per g of fresh weight). Bars represent mean ± standard error. Means with a different letter are significantly different (Tukey, p < 0.05). e = spores per plantlet and NaCl is expressed in 100 mM.
The results obtained in this study demonstrate the effect of different doses of mycorrhizae with or without NaCl on Pr and GB content in taro plantlets. Accumulation of TP and/or Pr in plants inoculated with mycorrhizae has been reported in Ashwagandha (Withania somnifera) [45], Thymus species (Thymus daenensis Celak and Thymus vulgaris L.) [46], melon (Cucumis melo) [29], and basil (Ocimum basilicum) [47]. In W. somnifera, Parihar and Bora [45] found that, when using Glomus mosseae at 100 spores per plant, the protein content increased significantly. Meddich et al. [29] observed in C. melo that when using mycorrhizae with Glomus sp. at 44 spores per plant for eight weeks, protein and proline content increased significantly. In this study, TP content did not show significant differences as the plantlets were inoculated with mycorrhizae (with or without NaCl). This was probably due to the specific expression of new proteins under salinity stress in taro; these proteins are not quantifiable by the Bradford [48] method, but rather a qualitative method is required to identify the proteins expressed under salinity stress. However, in other studies the accumulation of TP in plants inoculated with mycorrhizae under salinity stress has been reported in watermelon (Citrullus lanatus L.) [28], stevia (Stevia rebaudiana B.) [49], eucalyptus (Eucalyptus camaldulensis) [22], Saudi pearl millet (Pennisetum spicatum) [35], and pistachio (Pistacia vera L. cv. Ohadi) [34]. Janah et al. [49] in S. rebaudiana, when using a mycorrhizal consortium at 125 spores per plant and at a NaCl concentration of 80 mM for 2 months, found that the protein content increased significantly.
In this study, taro plantlets showed an increase in Pr and GB content when plantlets were treated with NaCl with or without AMF. This effect confirms that Pr and GB are biochemical indicators contributing to salinity stress tolerance mechanisms in taro. Ye et al. [28] observed in C. lanatus that Pr content increased when using Funneliformis mosseae at a dose of 700 spores per plantlet and a period of salinity stress with 60 mM NaCl for 6 days. Furthermore, [35] found that, in P. spicatum, when using Glomus mosseae at 2400 spores per plant and at a NaCl concentration of 60 mM for 2 months, the Pr content increased significantly. The Pr is used as a biomarker of osmotic stress; this compatible osmolyte maintains cell turgor, acts as a chaperone protein, and protects cells against free radical damage [50,51].
To date, no GB accumulation has been reported in taro plantlets under salinity stress conditions during the acclimatization stage and inoculated with AMF. However, GB accumulation has been reported in other species during salinity stress. Abbaspour et al. [34] found that, in P. vera, when inoculating with 1000 spores per plant with Rhizophagus irregularis Becker and Gerdemann (Gec) and at an NaCl concentration of 250 mM for seven days, the GB content increased significantly. GB acts as a compatible osmolyte and promotes antioxidant activity [52]. In addition, it can protect the enzymatic activity of Rubisco and the photosystem II complex during photosynthesis [53].
The symbiosis between AMF and plants under salinity stress could have also induced the synthesis of other compatible osmolytes such as polyamines, sugars, and polyols. These have an important role in osmotic adjustment and protect cells from ROS [54].
2.4. Soluble Phenols and Antioxidant Capacity
The different AMF doses and induced salinity stress had no effect on phenol content and antioxidant capacity (DPPH) (Figure 3). The highest phenol content was observed in plantlets under salinity stress without mycorrhizae, whereas the lowest phenol content was observed when no salinity stress was applied, with or without mycorrhizae (Figure 3d). Regarding antioxidant capacity, the highest DPPH content was observed at doses of 100 and 200 spores per plantlet under salinity stress, whereas the lowest DPPH content was observed when salinity stress was not applied, with or without mycorrhizae (Figure 3e).
The results obtained in this study demonstrate the effect of different treatments with mycorrhizae and NaCl on the content of soluble phenols and antioxidant capacity in taro plantlets. The accumulation of soluble phenols and antioxidant capacity in plants inoculated with mycorrhizae has been reported in French tamarisk (Tamarix gallica) [55], globe artichoke (Cynara cardunculus L. cv. scolymus Fiori) [56], prickly pear cactus (Opuntia ficus-indica) [57], and lettuce (Lactuca sativa L.) [58]. Bencherif et al. [55] in T. gallica found that when using a consortium (Funnneliformis mosseae, Septoglomus constrictum, Gigaspora gigantea, Glomus sp1., and Glomus sp2.) at a dose of 165 spores per plant, total phenolic content increased significantly more in roots than in leaves. Lahbouki et al. [57] observed in O. ficus-indica that when using a consortium with seven genera (Glomus (32%), Acaulospora (23%), Rhizophagus (14%), Scutellospora (9%), Diversispora (9%), Claroideoglomus (9%), and Gigaspora (4%)) distributed into 22 species at a dose of 344 spores per plant, total phenolic content increased significantly and antioxidant capacity decreased.
In this study, soluble phenol content increased as plantlets were inoculated with mycorrhizae (with or without NaCl); however, plantlets that were treated with mycorrhizae alone did not show a significant increase in soluble phenol content. The increase in soluble phenols in mycorrhizal-inoculated plantlets under salinity stress has been reported in lettuce (Lactuca sativa L.) [59], valerian (Valeriana officinalis L.) [60], forage grass (Lasiurus scindicus Henrard) [50], and Casuarinaceae species (Casuarina equisetifolia and Casuarina obesa) [61]. Santander et al. [59] in L. sativa observed that when using a native consortium with Funneliformis mosseae (MN264635), Claroideoglomus lamellosum (MN263071), and Diversispora celata (MN264508) at a dose of 2000 spores per plant and a concentration of 40 mM NaCl for 60 days, total phenolic content increased significantly. Malik et al. [50] in L. scindicus found that when using a native consortium with Claroideoglomus etunicatum, Funneliformis mosseae, Gigaspora margarita, and Scutellospora calospora at a dose of 282 spores per plant and a concentration of 100 mM NaCl, total phenolic content increased significantly.
Therefore, the increase in soluble phenol content could be a biochemical indicator of salinity stress tolerance. In addition, under salinity stress, plants synthesize antioxidant enzymes and non-enzymatic antioxidants as tolerance mechanisms [54]. Phenolic compounds are non-enzymatic antioxidants that scavenge free radicals [62]. Phenolic compounds can scavenge reactive oxygen intermediates while preventing the initiation of subsequent oxidative processes [50]. In addition, phenolic compounds are characterized by the availability of phenolic hydrogens as scavengers of hydrogen-donating radicals and consequently an increase in antioxidant capacity for scavenging activity [57].
Similarly, to soluble phenol content, taro plantlets showed an increase in the content of antioxidant capacity at different doses of mycorrhizae with NaCl; however, the highest antioxidant capacity content was obtained when plantlets were treated with mycorrhizae. This effect confirms that soluble phenols and antioxidant capacity are biochemical indicators that contribute to salinity stress tolerance mechanisms. Djighaly et al. [61] in C. equisetifolia and C. obesa observed that when using Rhizophagus fasciculatus (Thaxt.) C. Walker and A. Schüßler strain DAOM227130 at a dose of 648 spores per plant and a concentration of 200 mM NaCl for 15 days, total antioxidant capacity increased significantly. The accumulation of phenolic compounds and antioxidant capacity are a mechanism against oxidative stress. In this study, taro plantlets tolerance to NaCl could be related to the increase in soluble phenols and antioxidant capacity under salinity conditions.
In this study, the application of Glomus intraradices under salinity stress had an effect on plantlet development, photosynthetic pigments, Pr and GB, phenolic compounds, and antioxidant capacity in taro plantlets; no effects on PT accumulation were observed with the evaluated technique. However, the fungus–plant symbiosis specifically induces gene expression, activation of aquaporins, and the activity of enzymatic and non-enzymatic antioxidant compounds related to a salinity stress tolerance response [54,63]. In addition, AMF protect host plants against salinity stress through different mechanisms, where fungal structures (hyphae and mycelia) function as extensions of roots to maintain direct uptake of water and nutrients from the extraradical mycelium, improve soil structure, compartmentalization and ion exchange in vacuoles [64], and induce expression of genes coding for Na and K transporters, as well as H+ pumps that regulate ion transport in fungal cells [65]. Taro plantlets exposed to salinity stress showed senescent leaves compared to plantlets inoculated with AMF and without NaCl. In general, AMF-inoculated plantlets had better tolerance to salinity stress through the induction of different physiological and biochemical processes to maintain adequate plantlet development.
3. Materials and Methods
3.1. Plant Material and Micropropagation
For in vitro establishment of taro (Colocasia esculenta L. Schott cv. Criolla), 10 cm apices were collected. The apices were washed with water and Axion complete® commercial soap (Mission Hills, S.A. de C.V., San Jose of Iturbide, GT, Mexico), transferred to the laboratory where they were immersed in a solution containing 1 g L−1 fungicide (Cupravit, Bayer AG, Leverkusen, NW, DE) and 1 g L−1 bactericide (Agrimycin, Pfizer, New York, NY, USA) for 15 min, and then rinsed with tap water. In a laminar flow hood, the apices were reduced to 2 cm and then rinsed for five min in a 15% (v/v) solution of Cloralex™ commercial chlorine bleach (Industrias Alen, NL, Mexico) (5% a.i.) and with three drops of Tween 20® (Sigma-Aldrich® Chemical Company, Saint Louis, MO, USA) per 100 mL of water for 20 min. Subsequently, they were immersed in 70% ethanol for 1 min and rinsed three times with sterile distilled water. Finally, 1 cm apical meristems were excised with a scalpel and cultured individually in test tubes containing 10 mL of Murashige and Skoog (MS) [66] medium supplemented with 30 g L−1 sucrose and 1 mg L−1 BAP (6-Bencilaminopurine, Sigma-Aldrich®). The culture medium was adjusted to pH 5.8 with 0.1 N NaOH, after which 2.5 g L−1 Phytagel™ (Sigma-Aldrich®) were added as a gelling agent before being sterilized in an autoclave at 120 °C and 115 kPa for 15 min. The explants were incubated at 24 ± 2 °C, under irradiance of 40 ± 5 μmol m−2 s−1 and a photoperiod of 16 h light. After one week of culture, the apices were transferred for the multiplication phase to MS medium supplemented with 3 mg L−1 BAP (Sigma-Aldrich®). After three subcultures (30 d each), 2 cm shoots were individualized and transferred to MS rooting medium without growth regulators. After 15 days of culture, 5 cm long plantlets were rinsed with tap water and taken to the greenhouse for acclimatization.
3.2. Mycorrhizal Fungi Inoculation and Culture Conditions
Inoculation with mycorrhizae was performed under ex vitro greenhouse conditions using ex vitro plantlets with a length of 5 cm and mycorrhizae of the species Glomus intraradices (Biofertilizante INIFAP®, Chiapas, Mexico). The plant–fungus inoculation was carried out in 32-cavity polypropylene trays with a substrate made up of compost, peat moss, and agrolite (2:1:1 v/v). The substrate was sterilized in the autoclave for 30 min at 120 °C and 115 kPa. Different doses (0, 100, and 200 spores per plantlet) of G. intraradices were evaluated. Taro plantlets were covered with a translucent polyvinyl chloride dome to control humidity conditions. The inoculated plantlets were kept under greenhouse conditions with 60% shade at 30 ± 2 °C, relative humidity of 60 ± 10%, and natural light at an irradiance of 80 ± 10 μmol m−2 s−1 for 30 days. In a second phase, the dome was removed from the plantlets which were then kept at a temperature of 35 ± 2 °C, relative humidity of 30%, and natural light at an irradiance of 150 ± 10 μmol m−2 s−1 for 30 d. Throughout the experiment the plantlets were placed above a dome containing 3 L of osmosis water for two months. After 60 d of acclimatization, the plantlets were exposed to salinity stress with 3 L of 100 mM NaCl solution (Fermont, NL, Mexico) for 10 d. The salt concentrations were determined based on a previous study by [31] in C. esculenta, which found a maximum survivable concentration of 100 mM NaCl. After the salinity stress time, the colonization percentage, plantlet length, number of leaves, percentage of senescent leaves, roots per plantlet, root length, fresh weight, dry weight, and dry matter percentage were evaluated. In addition, the content of chlorophyll, carotenoids, proteins, proline, glycine-betaine, soluble phenols, and antioxidant capacity were determined.
3.3. Mycorrhizal Colonization
To visualize the effect of the different AMF doses on mycorrhizal colonization, segments of the roots were obtained and fixed in 4% paraformaldehyde, and then incubated for 48 h at room temperature. Root segments were washed three times with distilled water and then incubated in 10% KOH for 15 min at 120 °C. An alkaline hydrogen peroxide solution was added and incubated for 20 min at room temperature, after which 0.05% methylene blue (Sigma-Aldrich®) was added and incubated for 24 h at room temperature. The samples were observed under a microscope (M5LCD Velab, Co., Pharr, TX, USA) using a 40× objective. The mycorrhizal colonization percentages were determined using the following formula: Percentage of root colonization (%) = No. of infected segments/No. of examined root segments × 100.
3.4. Total Chlorophyll and Carotenoid Content
Total chlorophyll content was determined using the methodology proposed by [67]. A total of 250 mg of fresh leaf tissue were taken and macerated in a mortar with 80% acetone and allowed to stand at −4 °C for 24 h in 2.5 mL of 80% acetone. Subsequently, the mixture was filtered using No. 41 filter paper and adjusted to a final volume of 6.25 mL with 80% acetone. Finally, it was read at an absorbance of 663 and 645 nm for chlorophyll a and b, respectively. Readings were made using a spectrophotometer (Thermo Scientific Genesys 10S; Madison, WI, USA). Carotenoid (beta-carotene) content was determined according to the method described by [68] and quantification was done using the following formula:
where:
C = A450 × M × 1000/ε × δ
A = absorption determined at 450 nm
M = β-carotene molecular mass (537 g−1 mol)
ε = molar extinction coefficient of β-carotene in acetone (140,663 L−1 mol cm).
δ = optical path (cm).
3.5. Total Protein (TP) Estimation
The TP estimation was carried out by the method proposed by [48]. Ten milligrams of dry plant material were taken and macerated in a mortar in cold acetone. The macerated tissue was vacuum filtered until obtaining acetone powder. Next, 1.25 mL of 0.1 M tris-HCl pH 7.1 buffer was added. Subsequently, the solution was centrifuged at 3100× g for 20 min at 4 °C. Finally, 5 mL of Bradford solution were added to a 0.1 mL sample of the supernatant and read at an absorbance of 595 nm in a spectrophotometer (Thermo Scientific Genesys 10S, Madison, WI, USA). The values were interpolated in the calibration curve made with bovine albumin (Sigma-Aldrich®).
3.6. Proline (Pr) Determination
Pr was estimated according to the colorimetric method described by [69]. First, 250 mg of fresh leaf tissue were homogenized with 5 mL of 3% sulfosalicylic acid. The resulting paste was sieved through Whatman’s No. 2 filter paper and a 1 mL aliquot was taken, to which 1 mL of glacial acetic acid and 1 mL of ninhydrin were added. This mixture was left to incubate in a thermoregulated bath for 1 h at 100 °C. The tubes were removed and rapidly chilled on ice. Two mL of toluene were added and mixed for 30 s. The chromophore containing the toluene was separated to measure absorbance. The absorbance was read at 520 nm in a spectrophotometer (Thermo Scientific Genesys 10S, Madison, WI, USA) and the values were interpolated in the calibration curve made with L-proline standard (Sigma-Aldrich®).
3.7. Glycine-Betaine (GB) Determination
GB was determined using the colorimetric method proposed by [70]. First, 250 mg of dry macerated plant tissue were taken, to which 10 mL of deionized water were added and left to incubate for 24 h. An aliquot of 0.5 mL diluted at a 1:1 ratio with 2 N H2SO4 was taken, after which 0.1 mL of KI-I2 was added. The samples were mixed and left under refrigeration at 0–4 °C for 16 h. They were then centrifuged at 3100× g for 15 min at 0 °C and placed on ice for 1 h. Finally, the supernatant was separated and 4.5 mL of 1,2-Dichloroethane were added and left at room temperature for 2 h. The chromophore was separated to measure absorbance at 365 nm in the spectrophotometer (Thermo Scientific Genesys 10S, Madison, WI, USA). The values obtained were interpolated in the calibration curve made with glycine-betaine standard (Sigma-Aldrich®).
3.8. Determination of Soluble Phenols and Antioxidant Capacity
Phenolic content was determined according to [71]. First, 250 mg fresh weight of plant tissue were taken and macerated in a mortar; the extraction was performed with 10 mL of methanol: water (80:20). Subsequently, the solution was centrifuged at 3100× g for 10 min at 10 °C. Next, 150 µL of the supernatant were taken and 750 µL of 10% Folin-Ciocalteu’s reagent (E. Merck, Darmstadt, Germany) were added; it was homogenized gently, 600 µL of 20% calcium carbonate (Sigma-Aldrich®) were added as well, and then it was incubated for 2 h at 26 °C. Finally, the absorbance was measured at 765 nm using distilled water as a blank. Phenolic content was calculated from a gallic acid calibration curve (0–10,000 µg mL−1) and expressed as milligrams of gallic acid equivalents (GAE) per g of fresh weight (g FW) of taro plantlets.
The antioxidant capacity was expressed in DPPH (2, 2-Diphenyl-1-picrylhydrazyl). The DPPH was performed by the methodology proposed by [72]. An aliquot of 2900 µL of DPPH and 100 µL of methanolic extract obtained in the phenolic content determination was taken. The mixture was incubated at 26 °C for 1 h and the absorbance was measured at 515 nm. A calibration curve with Trolox (Sigma-Aldrich®) was used at different concentrations. Values obtained were interpolated in the calibration curve and expressed as trolox equivalents (TE) per g of fresh weight (g FW) of taro plantlets.
3.9. Experimental Design and Statistical Analysis
All experiments were performed in a completely randomized design and were run in triplicate. An analysis of variance was performed followed by Tukey’s range test (p < 0.05) using IBM SPSS® statistical software (version 22 for Windows). The percentage data were transformed with the formula Y = arcsine (√ (×/100)), where × is the value of the percentage.
4. Conclusions
In this study, since no significant differences were found for the developmental and biomass variables evaluated at doses of 100 and 200 spores per plantlet, it is suggested to apply 100 spores per plantlet to reduce spore handling, product transport, and arbuscular mycorrhizal fungi cost. Biochemical studies of proline, glycine-betaine, soluble phenols, and antioxidant capacity indicate the degree of plant stress and can be used as early selection markers for salinity-tolerant taro cultivars in breeding programs. It is suggested that future studies use a qualitative technique to determine the specific expression of proteins under salinity stress. In addition, reactive oxygen species production and antioxidant enzyme activities could be evaluated in a future study for a better understanding of salinity stress tolerance. Finally, the arbuscular mycorrhizal fungi evaluated in this study induce salinity tolerance in C. esculenta plantlets during the acclimatization stage. These plants can be transplanted to the field for further development under saline soil conditions in order to evaluate their tolerance to salinity stress.
Author Contributions
Conceptualization, writing—original draft preparation, and methodology, O.B.-B.; writing—review and editing, J.L.S.-C.; visualization, E.M.-Á. and J.J.B.-B. designed the experiments and head lab. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data in this study can be found in the manuscript.
Conflicts of Interest
The authors declare no conflict of interests.
References
- Gupta, K.; Kumar, A.; Tomer, V.; Kumar, V.; Saini, M. Potential of Colocasia leaves in human nutrition: Review on nutritional and phytochemical properties. J. Food Biochem. 2019, 43, e12878. [Google Scholar] [CrossRef] [PubMed]
- Chand, N.; Suthar, S.; Kumar, K.; Tyagi, V.K. Enhanced removal of nutrients and coliforms from domestic wastewater in cattle dung biochar-packed Colocasia esculenta-based vertical subsurface flow constructed wetland. J. Water Process Eng. 2021, 41, 101994. [Google Scholar] [CrossRef]
- Mitharwal, S.; Kumar, A.; Chauhan, K.; Taneja, N.K. Nutritional, phytochemical composition and potential health benefits of taro (Colocasia esculenta L.) leaves: A review. Food Chem. 2022, 383, 132406. [Google Scholar] [CrossRef] [PubMed]
- Farias, F.D.A.C.; de Souza Moretti, M.M.; Costa, M.S.; BordignonJunior, S.E.; Cavalcante, K.B.; Boscolo, M.; da Silva, R. Structural and physicochemical characteristics of taioba starch in comparison with cassava starch and its potential for ethanol production. Ind. Crops. Prod. 2020, 157, 112825. [Google Scholar] [CrossRef]
- Valenzuela, F.J.; Reineke, D.; Leventini, D.; Chen, C.C.L.; Barrett-Lennard, E.G.; Colmer, T.D.; Bazihizina, N. Plant responses to heterogeneous salinity: Agronomic relevance and research priorities. Ann. Bot. 2022, 129, 499–518. [Google Scholar] [CrossRef]
- Schwantes, A.M.; Parolari, A.J.; Swenson, J.J.; Johnson, D.M.; Domec, J.C.; Jackson, R.B.; Pelak, N.; Porporato, A. Accounting for landscape heterogeneity improves spatial predictions of tree vulnerability to drought. New Phytol. 2018, 220, 132–146. [Google Scholar] [CrossRef] [Green Version]
- Chourasia, K.N.; More, S.J.; Kumar, A.; Kumar, D.; Singh, B.; Bhardwaj, V.; Kumar, A.; Das, S.K.; Singh, R.K.; Zinta, G.; et al. Salinity responses and tolerance mechanisms in underground vegetable crops: An integrative review. Planta 2022, 255, 1–25. [Google Scholar] [CrossRef]
- Martínez-Santos, E.; Cruz-Cruz, C.A.; Spinoso-Castillo, J.L.; Bello-Bello, J.J. In vitro response of vanilla (Vanilla planifolia Jacks. ex Andrews) to PEG-induced osmotic stress. Sci. Rep. 2021, 11, 22611. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, B.; Raigond, P.; Sahu, C.; Mishra, U.N.; Sharma, S.; Lal, M.K. Phytic acid: Blessing in disguise, a prime compound required for both plant and human nutrition. Food Res. Int. 2021, 142, 110193. [Google Scholar] [CrossRef]
- Gómez-Kosky, R.; Jaramillo, D.N.; Esquiro, C.R.; Villegas, A.B.; Calimano, M.B.; Armas, P.M.; Ferreiro, J.A.; Pineda, E.; Kukurtcu, B.; Daniels, D.D. Effect of VIUSID Agro® and FitoMas-E® on the Ex-Vitro Acclimatization of Sugarcane Plants (Saccharum spp.) Cultivar C90-469. Sugar Tech 2020, 22, 42–51. [Google Scholar] [CrossRef]
- de Souza Ferrari, M.P.; da Cruz, R.M.S.; dos Santos Queiroz, M.; de Andrade, M.M.; Alberton, O.; Magalhães, H.M. Efficient ex vitro rooting, acclimatization, and cultivation of Curcuma longa L. from mycorrhizal fungi. J. Crop Sci. Biotechnol. 2020, 23, 469–482. [Google Scholar] [CrossRef]
- Wang, L.; Yang, D.; Ma, F.; Wang, G.; You, Y. Recent advances in responses of arbuscular mycorrhizal fungi-Plant symbiosis to engineered nanoparticles. Chemosphere 2022, 286, 131644. [Google Scholar] [CrossRef]
- Choi, J.; Summers, W.; Paszkowski, U. Mechanisms underlying establishment of arbuscular mycorrhizal symbioses. Annu. Rev. Phytopathol. 2018, 56, 135–160. [Google Scholar] [CrossRef]
- Di Martino, C.; Fioretto, A.; Palmieri, D.; Torino, V.; Palumbo, G. Influence of tomato plant mycorrhization on nitrogen metabolism, growth and fructification on P-limited soil. J. Plant Growth Regul. 2019, 38, 1183–1195. [Google Scholar] [CrossRef]
- Ortas, I. Under filed conditions, mycorrhizal inoculum effectiveness depends on plant species and phosphorus nutrition. J. Plant Nutr. 2019, 42, 2349–2362. [Google Scholar] [CrossRef]
- Al-Karaki, G.N.; Williams, M. Mycorrhizal mixtures affect the growth, nutrition, and physiological responses of soybean to water deficit. Acta Physiol. Plant. 2021, 43, 1–9. [Google Scholar] [CrossRef]
- Wu, M.; Yan, Y.; Wang, Y.; Mao, Q.; Fu, Y.; Peng, X.; Yang, Z.; Ren, J.; Liu, A.; Chen, S.; et al. Arbuscular mycorrhizal fungi for vegetable (VT) enhance resistance to Rhizoctonia solani in watermelon by alleviating oxidative stress. Biol. Control 2021, 152, 104433. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Boopathi, T.; Manivannan, P. Comprehensive assessment of ameliorative effects of AMF in alleviating abiotic stress in tomato plants. J. Fungi 2021, 7, 303. [Google Scholar] [CrossRef]
- Gupta, S.; Thokchom, S.D.; Kapoor, R. Arbuscular mycorrhiza improves photosynthesis and restores alteration in sugar metabolism in Triticum aestivum L. grown in arsenic contaminated soil. Front. Plant Sci. 2021, 12, 334. [Google Scholar] [CrossRef]
- Salwan, R.; Sharma, A.; Sharma, V. Microbes mediated plant stress tolerance in saline agricultural ecosystem. Plant Soil 2019, 442, 1–22. [Google Scholar] [CrossRef]
- He, J.D.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Mycorrhizas enhance drought tolerance of trifoliate orange by enhancing activities and gene expression of antioxidant enzymes. Sci. Hortic. 2020, 262, 108745. [Google Scholar] [CrossRef]
- Klinsukon, C.; Lumyong, S.; Kuyper, T.W.; Boonlue, S. Colonization by arbuscular mycorrhizal fungi improves salinity tolerance of eucalyptus (Eucalyptus camaldulensis) seedlings. Sci. Rep. 2021, 11, 4362. [Google Scholar] [CrossRef]
- Thangavel, P.; Anjum, N.A.; Muthukumar, T.; Sridevi, G.; Vasudhevan, P.; Maruthupandian, A. Arbuscular mycorrhizae: Natural modulators of plant–nutrient relation and growth in stressful environments. Arch. Microbiol. 2022, 204, 1–22. [Google Scholar] [CrossRef]
- Wang, Y.; Dong, F.; Tang, M. Transcriptome Analysis of Arbuscular Mycorrhizal Casuarina glauca in Damage Mitigation of Roots on NaCl Stress. Microorganisms 2021, 10, 15. [Google Scholar] [CrossRef]
- Wang, J.; Yuan, J.; Ren, Q.; Zhang, B.; Zhang, J.; Huang, R.; Wang, G.G. Arbuscular mycorrhizal fungi enhanced salt tolerance of Gleditsia sinensis by modulating antioxidant activity, ion balance and P/N ratio. Plant Growth Regul. 2022, 97, 33–49. [Google Scholar] [CrossRef]
- Ndiate, N.I.; Zaman, Q.U.; Francis, I.N.; Dada, O.A.; Rehman, A.; Asif, M.; Goffner, D.; Kane, A.; Liqun, C.; Haider, F.U. Soil Amendment with Arbuscular Mycorrhizal Fungi and Biochar Improves Salinity Tolerance, Growth, and Lipid Metabolism of Common Wheat (Triticum aestivum L.). Sustainability 2022, 14, 3210. [Google Scholar] [CrossRef]
- Chen, Q.; Deng, X.; Elzenga, J.T.M.; van Elsas, J.D. Effect of soil bacteriomes on mycorrhizal colonization by Rhizophagus irregularis—interactive effects on maize (Zea mays L.) growth under salt stress. Biol. Fertil. Soils 2022, 58, 515–525. [Google Scholar] [CrossRef]
- Ye, L.; Zhao, X.; Bao, E.; Cao, K.; Zou, Z. Effects of arbuscular mycorrhizal fungi on watermelon growth, elemental uptake, antioxidant, and photosystem II activities and stress-response gene expressions under salinity-alkalinity stresses. Front. Plant Sci. 2019, 10, 863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meddich, A.; Ait Rahou, Y.; Boutasknit, A.; Ait-El-Mokhtar, M.; Fakhech, A.; Lahbouki, S.; Wahbi, S.; Wahbi, S. Role of mycorrhizal fungi in improving the tolerance of melon (Cucumus melo) under two water deficit partial root drying and regulated deficit irrigation. Plant Biosyst.- Int. J. Deal. All Asp. Plant Biol. 2021, 156, 469–479. [Google Scholar] [CrossRef]
- Sales, F.R.; Silva, A.O.; Sales, L.R.; Rodrigues, T.L.; de Souza Moreira, F.M.; Carneiro, M.A.C. Native Arbuscular Mycorrhizal Fungi Exhibit Biotechnological Potential in Improvement of Soil Biochemical Quality and in Increasing Yield in Sugarcane Cultivars. Sugar Tech 2021, 23, 1235–1246. [Google Scholar] [CrossRef]
- Lloyd, G.R.; Uesugi, A.; Gleadow, R.M. Effects of Salinity on the Growth and Nutrition of Taro (Colocasia esculenta): Implications for Food Security. Plants 2021, 10, 2319. [Google Scholar] [CrossRef]
- Amanifar, S.; Khodabandeloo, M.; Fard, E.M.; Askari, M.S.; Ashrafi, M. Alleviation of salt stress and changes in glycyrrhizin accumulation by arbuscular mycorrhiza in liquorice (Glycyrrhiza glabra) grown under salinity stress. Environ. Exp. Bot. 2019, 160, 25–34. [Google Scholar] [CrossRef]
- Eroğlu, Ç.G.; Cabral, C.; Ravnskov, S.; Bak Topbjerg, H.; Wollenweber, B. Arbuscular mycorrhiza influences carbon-use efficiency and grain yield of wheat grown under pre-and post-anthesis salinity stress. Plant Biol. 2020, 22, 863–871. [Google Scholar] [CrossRef]
- Abbaspour, H.; Pour, F.S.; Abdel-Wahhab, M.A. Arbuscular mycorrhizal symbiosis regulates the physiological responses, ion distribution and relevant gene expression to trigger salt stress tolerance in pistachio. Physiol. Mol. Biol. Plants 2021, 27, 1765–1778. [Google Scholar] [CrossRef]
- Alshoaibi, A. Improved Tolerance of Three Saudi Pearl Millet Cultivars (Pennisetum spicatum) to Salt Stress by Mycorrhiza. Phyton 2021, 90, 731–745. [Google Scholar] [CrossRef]
- Fors, R.O.; Saggin Júnior, O.J.; Carneiro, M.A.C.; Barbara, R.L.L. Berbara, R.L.L. Selection of arbuscular mycorrhizal fungi for sugarcane in four soils with the presence of dark septate endophytes. Acta Sci.-Agron. 2020, 42, e42477. [Google Scholar] [CrossRef] [Green Version]
- Campo, S.; Martín-Cardoso, H.; Olivé, M.; Pla, E.; Catala-Forner, M.; Martínez-Eixarch, M.; San Segundo, B. Effect of root colonization by arbuscular mycorrhizal fungi on growth, productivity and blast resistance in rice. Rice 2020, 13, 1–14. [Google Scholar] [CrossRef]
- Liang, B.B.; Wang, W.J.; Fan, X.X.; Kurakov, A.V.; Liu, Y.F.; Song, F.Q.; Chang, W. Arbuscular mycorrhizal fungi can ameliorate salt stress in Elaeagnus angustifolia by improving leaf photosynthetic function and ultrastructure. Plant Biol. J. 2021, 23, 232–241. [Google Scholar] [CrossRef]
- Liang, D.; Ni, Z.; Xia, H.; Xie, Y.; Lv, X.; Wang, J.; Lin, L.; Deng, Q.; Luo, X. Exogenous melatonin promotes biomass accumulation and photosynthesis of kiwifruit seedlings under drought stress. Sci. Hortic. 2019, 246, 34–43. [Google Scholar] [CrossRef]
- Aalipour, H.; Nikbakht, A.; Ghasemi, M.; Amiri, R. Morpho-physiological and biochemical responses of two turfgrass species to arbuscular mycorrhizal fungi and humic acid under water stress condition. J. Soil Sci. Plant Nutr. 2020, 20, 566–576. [Google Scholar] [CrossRef]
- Hashem, A.; Kumar, A.; Al-Dbass, A.M.; Alqarawi, A.A.; Al-Arjani, A.B.F.; Singh, G.; Farooq, M.; Abd Allah, E.F. Arbuscular mycorrhizal fungi and biochar improves drought tolerance in chickpea. Saudi J. Biol. Sci. 2019, 26, 614–624. [Google Scholar] [CrossRef]
- Bouskout, M.; Bourhia, M.; Al Feddy, M.N.; Dounas, H.; Salamatullah, A.M.; Soufan, W.; Ouahmane, L. Mycorrhizal Fungi Inoculation Improves Capparis spinosa’s Yield, Nutrient Uptake and Photosynthetic Efficiency under Water Deficit. Agronomy 2022, 12, 149. [Google Scholar] [CrossRef]
- Sun, T.; Rao, S.; Zhou, X.; Li, L. Plant carotenoids: Recent advances and future perspectives. Mol. Hortic. 2022, 2, 3. [Google Scholar] [CrossRef]
- Zahra, N.; Al Hinai, M.S.; Hafeez, M.B.; Rehman, A.; Wahid, A.; Siddique, K.H.; Farooq, M. Regulation of photosynthesis under salt stress and associated tolerance mechanisms. Plant Physiol. Biochem. 2022, 178, 55–69. [Google Scholar] [CrossRef]
- Parihar, P.; Bora, M. Effect of mycorrhiza (Glomus mosseae) on morphological and biochemical properties of Ashwagandha (Withania somnifera) (L.) Dunal. J. Appl. Nat. Sci. 2018, 10, 1115–1123. [Google Scholar] [CrossRef]
- Arpanahi, A.A.; Feizian, M.; Mehdipourian, G.; Khojasteh, D.N. Arbuscular mycorrhizal fungi inoculation improve essential oil and physiological parameters and nutritional values of Thymus daenensis Celak and Thymus vulgaris L. under normal and drought stress conditions. Eur. J. Soil Biol. 2020, 100, 103217. [Google Scholar] [CrossRef]
- Abd-Elghany, S.E.; Moustafa, A.A.; Gomaa, N.H.; Hamed, B.-E.A. Mycorrhizal impact on Ocimum basilicum grown under drought stress. Beni-Suef Univ. J. Basic Appl. Sci. 2021, 10, 72. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Janah, I.; Meddich, A.; Elhasnaoui, A.; Khayat, S.; Anli, M.; Boutasknit, A.; Aissam, S.; Loutfi, K. Arbuscular Mycorrhizal Fungi Mitigates Salt Stress Toxicity in Stevia rebaudiana Bertoni Through the Modulation of Physiological and Biochemical Responses. J. Soil Sci. Plant Nutr. 2021, 1–11. [Google Scholar] [CrossRef]
- Malik, J.A.; AlQarawi, A.A.; Dar, B.A.; Hashem, A.; Alshahrani, T.S.; AlZain, M.N.; Habib, M.M.; Javed, M.M.; Abd_Allah, E.F. Arbuscular Mycorrhizal Fungi Isolated from Highly Saline “Sabkha Habitat” Soil Alleviated the NaCl-Induced Stress and Improved Lasiurus scindicus Henr. Growth. Agriculture 2022, 12, 337. [Google Scholar] [CrossRef]
- Bahadur, A.; Batool, A.; Nasir, F.; Jiang, S.; Mingsen, Q.; Zhang, Q.; Pan, J.; Liu, Y.; Feng, H. Mechanistic insights into arbuscular mycorrhizal fungi-mediated drought stress tolerance in plants. Int. J. Mol. Sci. 2019, 20, 4199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Pérez, C.A.; Gómez-Merino, F.C.; Spinoso-Castillo, J.L.; Bello-Bello, J.J. In vitro screening of sugarcane cultivars (Saccharum spp. hybrids) for tolerance to polyethylene glycol-induced water stress. Agronomy 2021, 11, 598. [Google Scholar] [CrossRef]
- Masood, A.; Per, T.S.; Asgher, M.; Fatma, M.; Khan, M.I.R.; Rasheed, F.; Hussain, S.J.; Khan, N.A. Glycine betaine: Role in shifting plants toward adaptation under extreme environments. In Osmolytes and Plants Acclimation to Changing Environment: Emerging Omics Technologies; Springer: New Delhi, India, 2016; pp. 69–82. [Google Scholar] [CrossRef]
- Tomar, R.S.; Kataria, S.; Jajoo, A. Behind the scene: Critical role of reactive oxygen species and reactive nitrogen species in salt stress tolerance. J. Agron. Crop Sci. 2021, 207, 577–588. [Google Scholar] [CrossRef]
- Bencherif, K.; Djaballah, Z.; Brahimi, F.; Boutekrabt, A.; Dalpè, Y.; Sahraoui, A.L.H. Arbuscular mycorrhizal fungi affect total phenolic content and antimicrobial activity of Tamarix gallica in natural semi-arid Algerian areas. S. Afr. J. Bot. 2019, 125, 39–45. [Google Scholar] [CrossRef]
- Avio, L.; Maggini, R.; Ujvári, G.; Incrocci, L.; Giovannetti, M.; Turrini, A. Phenolics content and antioxidant activity in the leaves of two artichoke cultivars are differentially affected by six mycorrhizal symbionts. Sci. Hortic. 2020, 264, 109153. [Google Scholar] [CrossRef]
- Lahbouki, S.; Anli, M.; El Gabardi, S.; Ait-El-Mokhtar, M.; Ben-Laouane, R.; Boutasknit, A.; Ait-Rahou, Y.; Outzourhit, A.; Wahbi, S.; Douira, A.; et al. Evaluation of arbuscular mycorrhizal fungi and vermicompost supplementation on growth, phenolic content and antioxidant activity of prickly pear cactus (Opuntia ficus-indica). Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2021, 155, 1–11. [Google Scholar] [CrossRef]
- Rasouli, F.; Amini, T.; Asadi, M.; Hassanpouraghdam, M.B.; Aazami, M.A.; Ercisli, S.; Skrovankova, S.; Mlcek, J. Growth and Antioxidant Responses of Lettuce (Lactuca sativa L.) to Arbuscular Mycorrhiza Inoculation and Seaweed Extract Foliar Application. Agronomy 2022, 12, 401. [Google Scholar] [CrossRef]
- Santander, C.; Ruiz, A.; García, S.; Aroca, R.; Cumming, J.; Cornejo, P. Efficiency of two arbuscular mycorrhizal fungal inocula to improve saline stress tolerance in lettuce plants by changes of antioxidant defense mechanisms. J. Sci. Food Agric. 2020, 100, 1577–1587. [Google Scholar] [CrossRef]
- Amanifar, S.; Toghranegar, Z. The efficiency of arbuscular mycorrhiza for improving tolerance of Valeriana officinalis L. and enhancing valerenic acid accumulation under salinity stress. Ind. Crops Prod. 2020, 147, 112234. [Google Scholar] [CrossRef]
- Djighaly, P.I.; Diagne, N.; Ngom, D.; Cooper, K.; Pignoly, S.; Hocher, V.; Farrant, J.M.; Svistoonoff, S. Effect of symbiotic associations with Frankia and arbuscular mycorrhizal fungi on antioxidant activity and cell ultrastructure in C. equisetifolia and C. obesa under salt stress. J. For. Res. 2022, 27, 117–127. [Google Scholar] [CrossRef]
- Bors, W.; Michel, C.; Saran, M. Flavonoid antioxidants: Rate constants for reactions with oxygen radicals. Methods Enzymol. 1994, 234, 420–429. [Google Scholar] [CrossRef]
- Kataria, S.; Verma, S.K. Salinity stress responses and adaptive mechanisms in major glycophytic crops: The story so far. In Salinity Responses and Tolerance in Plants; Volume 1: Targeting Sensory, Transport and Signaling Mechanisms; Kumar, V., Wani, S.H., Suprasanna, P., Tran, L.-S., Eds.; Springer: Cham, Switzerland, 2018; Chapter 1; pp. 1–39. [Google Scholar]
- Zou, Y.N.; Wu, Q.S.; Kuča, K. Unravelling the role of arbuscular mycorrhizal fungi in mitigating the oxidative burst of plants under drought stress. Plant Biol. J. 2021, 23, 50–57. [Google Scholar] [CrossRef]
- Corratgé, C.; Zimmermann, S.; Lambilliotte, R.; Plassard, C.; Marmeisse, R.; Thibaud, J.B.; Lacombe, B.; Sentenac, H. Molecular and functional characterization of a Na+-K+ transporter from the Trk family in the ectomycorrhizal fungus Hebeloma cylindrosporum. J. Biol. Chem. 2007, 282, 26057–26066. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Harborne, J.B. Nitrogen Compounds. In Phytochemical Methods; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 1973; pp. 166–211. [Google Scholar]
- Biehler, E.; Mayer, F.; Hoffmann, L.; Krause, E.; Bohn, T. Comparison of 3 spectrophotometric methods for carotenoid determination in frequently consumed fruits and vegetables. J. Food Sci. 2010, 75, C55–C61. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I. D Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R. Rapid assay for determination of water-soluble quaternary ammonium compounds. Plant Soil 1983, 70, 303–307. [Google Scholar] [CrossRef]
- Payet, B.; Shum Cheong Sing, A.; Smadja, J. Comparison of the concentrations of phenolic constituents in cane sugar manufacturing products with their antioxidant activities. J. Agric. Food Chem. 2006, 54, 7270–7276. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-throughput assay of oxygen radical absorbance capacity (ORAC) using a multichannel liquid handling system coupled with a microplate fluorescence reader in 96-well format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).