
Multi-Omics Approaches to Study Molecular Mechanisms in Cannabis sativa


Abstract
:1. Introduction
2. Studying the Metabolomic Profile of Cannabis
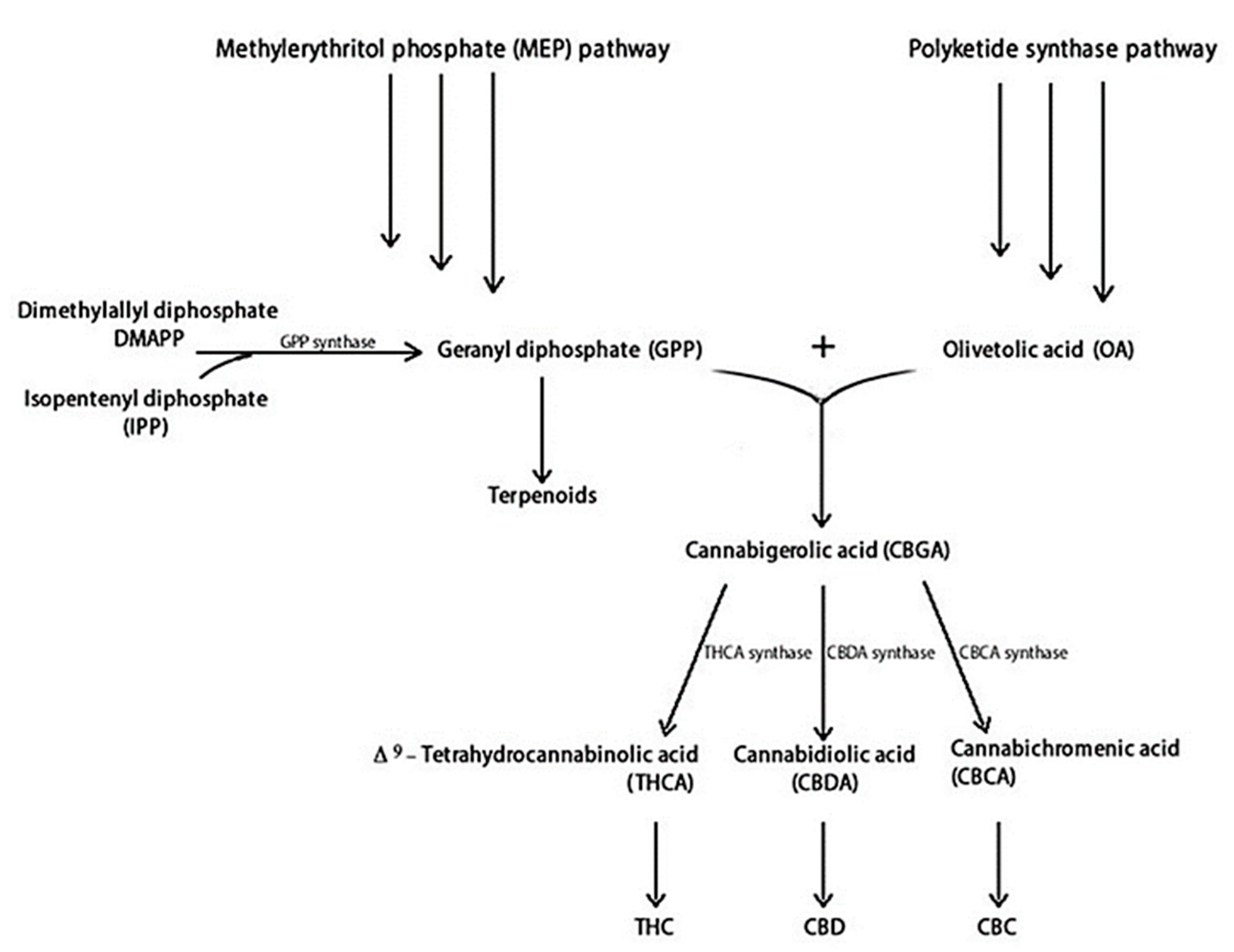
2.1. Key Metabolites: An Overview
2.2. Cannabis Metabolite Profiling Techniques
3. Genomic and Transcriptomic Cannabis Profiles
4. Multi-Omics Approaches in Cannabis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Clarke, R.C.; Merlin, M.D. Cannabis domestication, breeding history, present day genetic diversity, and future prospects. Crit. Rev. Plant Sci. 2016, 35, 293–327. [Google Scholar] [CrossRef]
- Andre, C.M.; Hausman, J.F.; Guerriero, G. Cannabis sativa: The Plant of the Thousand and One Molecules. Front. Plant Sci. 2016, 7, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avico, U.; Pacifici, R.; Zuccaro, P. Variations of tetrahydrocannabinol content in cannabis plants to distinguish the fibre-type from drug-type plants. Bull. Narc. 1985, 37, 61–65. [Google Scholar]
- Morales, P.; Hurst, D.P.; Reggio, P.H. Molecular targets of the phytocannabinoids: A complex picture. Prog. Chem. Org. Nat. Prod. 2017, 103, 103–131. [Google Scholar]
- Kovalchuk, I.; Pellino, M.; Rigault, P.; van Velzen, R.; Ebersbach, J.; Ashnest, J.R.; Mau, M.; Schranz, M.; Alcorn, J.; Laprairie, R. The genomics of cannabis and its close relatives. Annu. Rev. Plant Biol. 2020, 71, 713–739. [Google Scholar] [CrossRef] [Green Version]
- Piluzza, G.; Delogu, G.; Cabras, A.; Marceddu, S.; Bullitta, S. Differentiation between fiber and drugs of hemp (Cannabis sativa L.) from a collection of wild and domesticated accessions. Genet. Resour. Crop Evol. 2013, 60, 2331–2342. [Google Scholar] [CrossRef]
- Small, E.; Cronquist, A. A practical and natural taxonomy for Cannabis. Taxon 1976, 4, 405–435. [Google Scholar] [CrossRef]
- Henry, P.; Khatodia, S.; Kapoor, K.; Gonzales, B.; Middleton, A.; Hong, K.; Hilyard, A.; Johnson, S.; Allen, D.; Chester, Z.A.; et al. Single Nucleotide Polymorphism assay sheds light on the extent and distribution of genetic diversity, population structure and functional basis of key traits in cultivated North American cannabis. J. Cannabis Res. 2020, 2, 26. [Google Scholar] [CrossRef]
- Vergara, D.; Baker, H.; Clancy, K.; Keepers, K.G.; Mendieta, J.P.; Pauli, C.S.; Tittes, S.B.; White, K.H.; Kane, N.C. Genetic and genomic tools for Cannabis sativa. Crit. Rev. Plant Sci. 2016, 35, 364–377. [Google Scholar] [CrossRef]
- Vergara, D.; Feathers, C.; Huscher, E.L.; Holmes, B.; Haas, J.A.; Kane, N.C. Widely assumed phenotypic associations in Cannabis sativa lack a shared genetic basis. PeerJ 2021, 9, e10672. [Google Scholar] [CrossRef]
- McPartland, J.M.; Russo, E.B. Cannabis and cannabis extracts: Greater than the sum of their parts? J. Cannabis Ther. 2001, 1, 103–132. [Google Scholar] [CrossRef]
- Russo, E.B.; Jiang, H.E.; Li, X.; Sutton, A.; Carboni, A.; Del Bianco, F.; Mandolino, G.; Potter, D.J.; Zhao, Y.X.; Bera, S.; et al. Phytochemical and genetic analyses of ancient cannabis from Central Asia. J. Exp. Bot. 2008, 59, 4171–4182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Fuente, A.; Zamberlan, F.; Ferrán, A.S.; Carrillo, F.; Tagliazucchi, E.; Pallavicini, C. Relationship among subjective responses, flavor, and chemical composition across more than 800 commercial cannabis varieties. J. Cannabis Res. 2020, 2, 21. [Google Scholar] [CrossRef]
- Reimann-Philipp, U.; Speck, M.; Orser, C.; Johnson, S.; Hilyard, A.; Turner, H.; Stokes, A.J.; Small-Howard, A.L. Cannabis chemovar nomenclature misrepresents chemical and genetic diversity; survey of variations in chemical profiles and genetic markers in Nevada Medical Cannabis samples. Cannabis Cannabinoid Res. 2019, 5, 215–230. [Google Scholar] [CrossRef] [Green Version]
- Lynch, R.C.; Vergara, D.; Tittes, S.; White, K.; Schwartz, C.J.; Gibbs, M.J.; Ruthenburg, T.C.; deCesare, K.; Land, D.P.; Kane, N.C. Genomic and Chemical Diversity in Cannabis. Crit. Rev. Plant Sci. 2016, 35, 349–363. [Google Scholar] [CrossRef] [Green Version]
- Mandolino, G.; Carboni, A. Potential of Marker-Assisted Selection in Hemp Genetic Improvement. Euphytica 2004, 140, 107–120. [Google Scholar] [CrossRef]
- Green, G. (Ed.) The Cannabis Grow Bible: The Definitive Guide to Growing Marijuana for Recreational and Medicinal Use; Green Candy Press: San Francisco, CA, USA, 2017. [Google Scholar]
- Cervantes, J. (Ed.) Marijuana Horticulture: The Indoor/Outdoor Medical Grower’s Bible; Van Patten Publishing: Vancouver, WA, USA, 2007. [Google Scholar]
- Hurgobin, B.; Tamiru-Oli, M.; Welling, M.T.; Doblin, M.S.; Bacic, A.; Whelan, J.A.; Lewsey, M.G. Recent advances in Cannabis sativa genomics research. New Phytol. 2020, 230, 73–89. [Google Scholar] [CrossRef]
- Aliferis, K.A.; Bernard-Perron, D. Cannabinomics: Application of Metabolomics in Cannabis (Cannabis sativa L.) Research and Development. Front. Plant. Sci. 2020, 11, 554. [Google Scholar] [CrossRef]
- Pacula, R.L.; Smart, R. Medical Marijuana and Marijuana Legalization. Ann. Rev. Clin. Psychol. 2017, 13, 397–419. [Google Scholar] [CrossRef]
- Cox, C. The Canadian Cannabis Act legalizes and regulates recreational Cannabis use in 2018. Health Policy 2018, 122, 205–209. [Google Scholar] [CrossRef]
- Van Bakel, H.; Stout, J.; Cote, A.; Tallon, C.; Sharpe, A.; Hughes, T.; Page, J. The draft genome and transcriptome of Cannabis sativa. Genome Biol. 2011, 12, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braich, S.; Baillie, R.C.; Jewell, L.S.; Spangenberg, G.C.; Cogan, N.O.I. Generation of a comprehensive transcriptome atlas and transcriptome dynamics in medicinal cannabis. Sci. Rep. 2019, 9, 16583. [Google Scholar] [CrossRef]
- McKernan, K.J.; Helbert, Y.; Kane, L.T.; Ebling, H.; Zhang, L.; Liu, B.; Eaton, Z.; McLaughlin, S.; Kingan, S.; Baybayan, P. Sequence and annotation of 42 cannabis genomes reveals extensive copy number variation in cannabinoid synthesis and pathogen resistance genes. bioRxiv 2020. [Google Scholar] [CrossRef]
- Pisupati, R.; Vergara, D.; Kane, N.C. Diversity and evolution of the repetitive genomic content in Cannabis sativa. BMC Genom. 2018, 19, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.; Wang, B.; Xie, S.; Xu, X.; Zhang, J.; Pei, L.; Yu, Y.; Yang, W.; Zhang, Y. A high quality reference genome of wild Cannabis sativa. Hortic. Res. 2020, 7, 73. [Google Scholar] [CrossRef] [PubMed]
- Lubell, J.D.; Brand, M.H. Foliar sprays of silver thiosulfate produce male flowers on female hemp plants. Horttechnology 2018, 28, 743–747. [Google Scholar] [CrossRef] [Green Version]
- Crandall, S.G.; Gold, K.M.; Jiménez-Gasco, M.M.; Filgueiras, C.C.; Willett, D.S. A multi-omics approach to solving problems in plant disease ecology. PLoS ONE 2020, 15, e0237975. [Google Scholar] [CrossRef]
- Yang, Y.; Saand, M.A.; Huang, L.; Abdelaal, W.B.; Zhang, J.; Wu, Y.; Li, J.; Sirohi, M.H.; Wang, F. Applications of Multi-Omics Technologies for Crop Improvement. Front. Plant Sci. 2021, 12, 563953. [Google Scholar] [CrossRef]
- Amer, B.; Baidoo, E.E.K. Omics-Driven Biotechnology for Industrial Applications. Front. Bioeng. Biotechnol. 2021, 9, 613307. [Google Scholar] [CrossRef]
- Chandra, S.; Lata, H.; Khan, I.A.; Elsohly, M.A. Cannabis sativa L.: Botany and horticulture. In Cannabis sativa L.—Botany and Biotechnology; Chandra, S., Lata, H., Elsohly, M.A., Eds.; Springer: Cham, Switzerland, 2017; pp. 79–100. [Google Scholar]
- Livingston, S.; Quilichini, T.; Booth, J.; Wong, D.; Rensing, K.; Laflamme-Yonkman, J.; Castellarin, S.; Bohlmann, J.; Page, J.; Samuels, A.L. Cannabis glandular trichomes alter morphology and metabolite content during flower maturation. Plant J. 2019, 101, 37–56. [Google Scholar] [CrossRef]
- Tahir, M.N.; Shahbazi, F.; Rondeau-Gagné, S.; Trant, J.F. The biosynthesis of the cannabinoids. J. Cannabis Res. 2021, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, N.; Gorelick, J.; Zerahia, R.; Koch, S. Impact of N, P, K, and humic acid supplementation on the chemical profile of medical cannabis (Cannabis sativa L. ). Front. Plant Sci. 2019, 10, 736. [Google Scholar] [CrossRef] [Green Version]
- Eichhorn Bilodeau, S.; Wu, B.S.; Rufyikiri, A.S.; MacPherson, S.; Lefsrud, M. An update on plant photobiology and implications for Cannabis production. Front. Plant Sci. 2019, 10, 296. [Google Scholar] [CrossRef] [PubMed]
- Mostafaei Dehnavi, M.; Ebadi, A.; Peirovi, A.; Taylor, G.; Salami, S.A. THC and CBD Fingerprinting of an Elite Cannabis Collection from Iran: Quantifying Diversity to Underpin Future Cannabis Breeding. Plants 2022, 11, 129. [Google Scholar] [CrossRef]
- Weiblen, G.D.; Wenger, J.P.; Craft, K.J.; ElSohly, M.A.; Mehmedic, Z.; Treiber, E.L.; Marks, M.D. Gene duplication and divergence affecting drug content in Cannabis sativa. New Phytol. 2015, 208, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Gülck, T.; Møller, B.L. Phytocannabinoids: Origins and Biosynthesis. Trends Plant Sci. 2020, 25, 985–1004. [Google Scholar] [CrossRef]
- Walsh, K.B.; McKinney, A.E.; Holmes, A.E. Minor Cannabinoids: Biosynthesis, Molecular Pharmacology and Potential Therapeutic Uses. Front. Pharmacol. 2021, 12, 777804. [Google Scholar] [CrossRef]
- Grof, C.P. Cannabis, from plant to pill. Br. J. Clin. Pharmacol. 2018, 84, 2463–2467. [Google Scholar] [CrossRef]
- Gallily, R.; Yekhtin, Z.; Hanuš, L.O. The anti-inflammatory properties of terpenoids from Cannabis. Cannabis Cannabinoid Res. 2018, 3, 282–290. [Google Scholar] [CrossRef] [Green Version]
- Zager, J.J.; Lange, I.; Srividya, N.; Smith, A.; Lange, B.M. Gene networks underlying cannabinoid and terpenoid accumulation in Cannabis. Plant Physiol. 2019, 180, 1877–1897. [Google Scholar] [CrossRef] [Green Version]
- Leghissa, A.; Hildenbrand, Z.L.; Schug, K.A. A review of methods for the chemical characterization of Cannabis natural products. J. Sep. Sci. 2018, 41, 398–415. [Google Scholar] [CrossRef] [PubMed]
- Fuhrer, T.; Zamboni, N. High-throughput discovery metabolomics. Curr. Opin. Biotechnol. 2015, 31, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Magagnini, G.; Grassi, G.; Kotiranta, S. The Effect of Light Spectrum on the Morphology and Cannabinoid Content of Cannabis sativa L. Med. Cannabis Cannabinoids 2018, 1, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Basas-Jaumandreu, J.; De las Heras, F.X.C. GC-MS metabolite profile and identification of unusual homologous cannabinoids in high potency Cannabis sativa. Planta Med. 2020, 86, 338–347. [Google Scholar] [CrossRef]
- Fulvio, F.; Paris, R.; Montanari, M.; Citti, C.; Cilento, V.; Bassolino, L.; Moschella, A.; Alberti, I.; Pecchioni, N.; Cannazza, G.; et al. Analysis of Sequence Variability and Transcriptional Profile of Cannabinoid synthase Genes in Cannabis sativa L. Chemotypes with a Focus on Cannabichromenic acid synthase. Plants 2021, 10, 1857. [Google Scholar] [CrossRef]
- Burgel, L.; Hartung, J.; Pflugfelder, A.; Graeff-Hönninger, S. Impact of Growth Stage and Biomass Fractions on Cannabinoid Content and Yield of Different Hemp (Cannabis sativa L.) Genotypes. Agronomy 2020, 10, 372. [Google Scholar] [CrossRef] [Green Version]
- Fischedick, J.T. Identification of terpenoid chemotypes among High (−)-trans-Δ9-Tetrahydrocannabinol-Producing Cannabis sativa L. Cultivars. Cannabis Cannabinoid Res. 2017, 2, 34–47. [Google Scholar] [CrossRef] [Green Version]
- Sapkale, G.N.; Patil, S.M.; Surwase, U.S.; Bhatbhage, P.K. Supercritical Fluid Extraction: A Review. Int. J. Chem. Sci. 2010, 8, 729–743. [Google Scholar]
- Gallo-Molina, A.C.; Castro-Vargas, H.I.; Garzón-Méndez, W.F.; Ramírez, J.A.M.; Monroy, Z.J.R.; King, J.W.; Parada-Alfonso, F. Extraction, isolation and purification of tetrahydrocannabinol from the Cannabis sativa L. plant using supercritical fluid extraction and solid phase extraction. J. Supercrit. Fluids 2019, 146, 208–216. [Google Scholar]
- Ramirez, C.L.; Fanovich, M.A.; Churio, M.S. Cannabinoids: Extraction methods, analysis, and physicochemical characterization. Stud. Nat. Prod. Chem. 2019, 61, 143–173. [Google Scholar]
- Leghissa, A.; Hildenbrand, Z.L.; Foss, F.W.; Schug, K.A. Determination of cannabinoids from a surrogate hops matrix using multiple reaction monitoring gas chromatography with triple quadrupole mass spectrometry. J. Sep. Sci. 2018, 41, 459–468. [Google Scholar] [CrossRef]
- Markley, J.L.; Brüschweiler, R.; Edison, A.S.; Eghbalnia, H.R.; Powers, R.; Raftery, D.; Wishart, D.S. The future of NMR-based metabolomics. Curr. Opin. Biotechnol. 2017, 43, 34–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjerrum, J.T. Metabonomics; Springer: New York, NY, USA, 2015. [Google Scholar]
- Choi, Y.H.; Kim, H.K.; Hazekamp, A.; Erkelens, C.; Lefeber, A.W.M.; Verpoorte, R. Metabolomic differentiation of Cannabis sativa cultivars using 1H NMR spectroscopy and principal component analysis. J. Nat. Prod. 2004, 67, 953–957. [Google Scholar] [CrossRef] [PubMed]
- Peschel, W.; Politi, M. 1H NMR and HPLC/DAD for Cannabis sativa L. chemotype distinction, extract profiling and specification. Talanta 2015, 140, 150–165. [Google Scholar] [CrossRef] [PubMed]
- Happyana, N.; Agnolet, S.; Muntendam, R.; Van Dam, A.; Schneider, B.; Kayser, O. Analysis of cannabinoids in laser-microdissected trichomes of medicinal Cannabis sativa using LCMS and cryogenic NMR. Phytochemistry 2013, 87, 51–59. [Google Scholar] [CrossRef]
- Wang, X.; Harrington, P.D.B.; Baugh, S.F. Effect of preprocessing high-resolution mass spectra on the pattern recognition of Cannabis, hemp, and liquor. Talanta 2018, 180, 229–238. [Google Scholar] [CrossRef]
- Vásquez-Ocmín, P.G.; Marti, G.; Bonhomme, M.; Mathis, F.; Fournier, S.; Bertani, S.; Maciuk, A. Cannabinoids vs. whole metabolome: Relevance of cannabinomics in analyzing Cannabis varieties. Anal. Chim. Acta 2021, 1184, 339020. [Google Scholar] [CrossRef]
- Laverty, K.U.; Stout, J.M.; Sullivan, M.J.; Shah, H.; Gill, N.; Holbrook, L.; Deikus, G.; Sebra, R.; Hughes, T.R.; Page, J.E. A physical and genetic map of Cannabis sativa identifies extensive rearrangements at the THC/CBD acid synthase loci. Genome Res. 2019, 29, 146–156. [Google Scholar] [CrossRef] [Green Version]
- Grassa, C.J.; Weiblen, G.D.; Wenger, J.P.; Dabney, C.; Poplawski, S.G.; Motley, S.T.; Michael, T.P.; Schwartz, C.J. A new Cannabis genome assembly associates elevated cannabidiol (CBD) with hemp introgressed into marijuana. New Phytol. 2021, 230, 1665–1679. [Google Scholar] [CrossRef]
- Ren, G.; Zhang, X.; Li, Y.; Ridout, K.; Serrano-Serrano, M.L.; Yang, Y.; Liu, A.; Ravikanth, G.; Nawaz, M.A.; Mumtaz, A.S.; et al. Large-scale whole-genome resequencing unravels the domestication history of Cannabis sativa. Sci. Adv. 2021, 7, 29. [Google Scholar] [CrossRef]
- McGarvey, P.; Huang, J.; McCoy, M.; Orvis, J.; Katsir, Y.; Lotringer, N.; Nesher, I.; Kavarana, M.; Sun, M.; Peet, R.; et al. De novo assembly and annotation of transcriptomes from two cultivars of Cannabis sativa with different cannabinoid profiles. Gene 2020, 762, 145026. [Google Scholar] [CrossRef]
- Van Velzen, R.; Schranz, M.E. Origin and Evolution of the Cannabinoid Oxidocyclase Gene Family. Genome Biol. Evol. 2021, 13, evab130. [Google Scholar] [CrossRef] [PubMed]
- Prentout, D.; Razumova, O.; Rhoné, B.; Badouin, H.; Henri, H.; Feng, C.; Käfer, J.; Karlov, G.; Marais, G.A. An efficient RNA-seq-based segregation analysis identifies the sex chromosomes of Cannabis sativa. Genome Res. 2020, 30, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Welling, M.T.; Liu, L.; Kretzschmar, T.; Mauleon, R.; Ansari, O.; King, G.J. An extreme-phenotype genome-wide association study identifies candidate cannabinoid pathway genes in Cannabis. Sci. Rep. 2020, 10, 18643. [Google Scholar] [CrossRef]
- Petit, J.; Salentijn, E.M.J.; Paulo, M.J.; Denneboom, C.; Trindade, L.M. Genetic Architecture of Flowering Time and Sex Determination in Hemp (Cannabis sativa L.): A Genome-Wide Association Study. Front. Plant Sci. 2020, 11, 569958. [Google Scholar] [CrossRef]
- Wang, S.; Wei, J.; Li, R.; Qu, H.; Chater, J.M.; Ma, R.; Li, Y.; Xie, W.; Jia, Z. Identification of optimal prediction models using multi-omic data for selecting hybrid rice. Heredity 2019, 123, 395–406. [Google Scholar] [CrossRef]
- Knoch, D.; Werner, C.R.; Meyer, R.C.; Riewe, D.; Abbadi, A.; Lücke, S.; Altmann, T. Multi-omics-based prediction of hybrid performance in canola. Theor. Appl. Genet. 2021, 134, 1147–1165. [Google Scholar] [CrossRef]
- Allen, K.D.; McKernan, K.; Pauli, C.; Roe, J.; Torres, A.; Gaudino, R. Genomic characterization of the complete terpene synthase gene family from Cannabis sativa. PLoS ONE 2019, 14, e0222363. [Google Scholar] [CrossRef]
- Guerriero, G.; Mangeot-Peter, L.; Legay, S.; Behr, M.; Lutts, S.; Siddiqui, K.S.; Hausman, J.F. Identification of fasciclin-like arabinogalactan proteins in textile hemp (Cannabis sativa L.): In silico analyses and gene expression patterns in different tissues. BMC Genom. 2017, 18, 741. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Tian, F.; Yang, D.C.; Meng, Y.Q.; Kong, L.; Luo, J.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res. 2017, 45, D1040–D1045. [Google Scholar] [CrossRef] [Green Version]
- Matoušek, J.; Kocábek, T.; Patzak, J.; Bříza, J.; Siglová, K.; Mishra, A.K.; Duraisamy, G.S.; Týcová, A.; Ono, E.; Krofta, K. The “putative” role of transcription factors from HlWRKY family in the regulation of the fnal steps of prenylfavonid and bitter acids biosynthesis in hop (Humulus lupulus L.). Plant. Mol. Biol. 2016, 92, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhu, P.; Cai, S.; Haughn, G.; Page, J.E. Three novel transcription factors involved in cannabinoid biosynthesis in Cannabis sativa L. Plant Mol. Biol. 2021, 106, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, G.; Behr, M.; Legay, S.; Mangeot-Peter, L.; Zorzan, S.; Ghoniem, M.; Hausman, J.F. Transcriptomic profiling of hemp bast fibres at different developmental stages. Sci. Rep. 2017, 7, 4961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rengasamy, P. Soil Salinization. In Oxford Research Encyclopedia of Environmental Science; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Cao, K.; Sun, Y.; Han, C.; Zhang, X.; Zhao, Y.; Jiang, Y.; Jiang, Y.; Sun, X.; Guo, Y.; Wang, X. The transcriptome of saline-alkaline resistant industrial hemp (Cannabis sativa L.) exposed to NaHCO3 stress. Ind. Crops Prod. 2021, 170, 113766. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, C.; Huang, S.; Chang, L.; Li, J.; Tang, H.; Dey, S.; Biswas, A.; Du, D.; Li, D.; et al. Key Cannabis Salt-Responsive Genes and Pathways Revealed by Comparative Transcriptome and Physiological Analyses of Contrasting Varieties. Agronomy 2021, 11, 2338. [Google Scholar] [CrossRef]
- Weng, Q.; Zhao, Y.; Yanan, Z.; Song, X.; Yuan, J.; Liu, Y. Identification of salt stress-responsive proteins in maize (Zea may) seedlings using itraq-based proteomic technique. Iran. J. Biotechnol. 2021, 19, e2512. [Google Scholar]
- Pandita, D.; Pandita, A.; Wani, S.H.; Abdelmohsen, S.A.M.; Alyousef, H.A.; Abdelbacki, A.M.M.; Al-Yafrasi, M.A.; Al-Mana, F.A.; Elansary, H.O. Crosstalk of Multi-Omics Platforms with Plants of Therapeutic Importance. Cells 2021, 10, 1296. [Google Scholar] [CrossRef]
- Onofri, C.; de Meijer, E.P.M.; Mandolino, G. Sequence heterogeneity of cannabidiolic- and tetrahydrocannabinolic acid-synthase in Cannabis sativa L. and its relationship with chemical phenotype. Phytochemistry 2015, 116, 57–68. [Google Scholar] [CrossRef]
- Rasmann, S.; Köllner, T.G.; Degenhardt, J.; Hiltpold, I.; Toepfer, S.; Kuhlmann, U.; Gershenzon, J.; Turlings, T.C.J. Recruitment of entomopathogenic nematodes by insect-damaged maize roots. Nature 2005, 434, 732–737. [Google Scholar] [CrossRef]
- Booth, J.K.; Yuen, M.M.; Jancsik, S.; Madilao, L.; Page, J.; Bohlmann, J. Terpene synthases and terpene variation in Cannabis sativa. Plant Physiol. 2020, 184, 130–147. [Google Scholar] [CrossRef]
- Vergara, D.; Huscher, E.L.; Keepers, K.G.; Givens, R.M.; Cizek, C.G.; Torres, A.; Gaudino, R.; Kane, N.C.J.B. Gene copy number is associated with phytochemistry in Cannabis sativa. AoB Plants 2019, 11, plz074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muyle, A.; Shearn, R.; Marais, G.A. The evolution of sex chromosomes and dosage compensation in plants. Genome Biol. Evol. 2017, 9, 627–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muyle, A.; Zemp, N.; Fruchard, C.; Cegan, R.; Vrana, J.; Deschamps, C.; Tavares, R.; Hobza, R.; Picard, F.; Widmer, A.; et al. Genomic imprinting mediates dosage compensation in a young plant XY system. Nat. Plants 2018, 4, 677–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woods, P.; Campbell, B.J.; Nicodemus, T.J.; Cahoon, E.B.; Mullen, J.L.; McKay, J.K. Quantitative trait loci controlling agronomic and biochemical traits in Cannabis sativa. Genetics 2021, 219, iyab099. [Google Scholar] [CrossRef]
- Gagne, S.J.; Stout, J.M.; Liu, E.; Boubakir, Z.; Clark, S.M.; Page, J.E. Identification of olivetolic acid cyclase from Cannabis sativa reveals a unique catalytic route to plant polyketides. Proc. Natl. Acad. Sci. USA 2012, 109, 12811–12816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, S.; Zhang, Z.; Huang, S.; Bai, X.; Huang, Z.; Zhang, Y.J.; Huang, L.; Tang, W.; Haughn, G.; You, S.; et al. CannabisGDB: A comprehensive genomic database for Cannabis sativa L. Plant Biotechnol. J. 2021, 19, 857–859. [Google Scholar] [CrossRef]
- Dolgin, E. The bioengineering of cannabis. Nature 2019, 572, S5–S7. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, G.; Cheng, C.; Lei, L.; Sun, J.; Xu, Y.; Deng, C.; Dai, Z.; Yang, Z.; Chen, X.; et al. Establishment of an Agrobacterium-mediated genetic transformation and CRISPR/Cas9-mediated targeted mutagenesis in Hemp (Cannabis sativa L.). Plant Biotechnol. J. 2021, 19, 1979–1987. [Google Scholar] [CrossRef]
- Ahmed, S.; Gao, X.; Jahan, M.A.; Adams, M.; Wu, N.; Kovinich, N. Nanoparticle-based genetic transformation of Cannabis sativa. J. Biotechnol. 2021, 326, 48–51. [Google Scholar] [CrossRef]
- Hurgobin, B.; Golicz, A.A.; Bayer, P.E.; Chan, C.K.K.; Tirnaz, S.; Dolatabadian, A.; Schiessl, S.V.; Samans, B.; Montenegro, J.D.; Parkin, I.A. Homoeologous exchange is a major cause of gene presence/absence variation in the amphidiploid Brassica napus. Plant Biotechnol. J. 2018, 16, 1265–1274. [Google Scholar] [CrossRef] [Green Version]
- NRGene. Creating the First Cannabis Pangenome. 2018. Available online: https://www.analyticalcannabis.com/news/creating-the-first-cannabis-pangenome-308027 (accessed on 1 September 2020).
- Medicinal Genomics. The Cannabis Pan-Genome Project: Advancing Cannabis Breeding. Available online: https://www.medicinalgenomics.com/cannabis-pan-genome-project-advancing-cannabis-breeding/ (accessed on 1 September 2020).
- Zhao, C.; Zhang, Y.; Du, J.; Guo, X.; Wen, W.; Gu, S.; Wang, J.; Fan, J. Crop Phenomics: Current Status and Perspectives. Front. Plant Sci. 2019, 10, 714. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Abbreviations | Definition |
|---|---|
| CBC | Cannabichromene |
| CBCA | Cannabichromenic Acid |
| CBD | Cannabidiol |
| CBDA | Acidic Precursors of Cannabidiol |
| CBG | Cannabigerol |
| CBGA | Acidic Precursors of Cannabigerolic |
| CBDRx | High Cannabidiol Content Cannabis Cultivar |
| CNV | Copy Number Variation |
| DEG | Differentially Expressed Gene |
| DLLLME | Dispersive Liquid-Liquid Microextraction |
| DMAPP | Dimethylallyl Diphosphate |
| FN | Finola Cannabis Cultivar |
| GBS | Genotype By Sequencing |
| GC-EIMS | Gas Chromatography-Electron Impact Mass Spectrometry |
| GC-FID | Gas Chromatography-Flame Ionization Detection |
| GC-MS | Gas Chromatography–Mass Spectrometry |
| GPP | Geranyl Diphosphate |
| GS | Genomic Selection |
| GWAS | Genome-Wide Association Study |
| HPLC | High Performance Liquid Chromatography |
| HRMS | High-Resolution Mass Spectrometry |
| HS-SPME | Headspace Solid-Phase Micro Extraction |
| IPP | Isopentenyl Diphosphate |
| ISO-Seq | Isoform Sequencing |
| JL | Jamaican Lion Cannabis Cultivar |
| LC-DAD | Liquid Chromatography-Diode Array Detector |
| LC-MS | Liquid Chromatography–Mass Spectrometry |
| MEP | Methylerythritol Phosphate |
| MS | Mass Spectrometry |
| NGS | Next Generation Sequencing |
| NMR | Nuclear Magnetic Resonance |
| OA | Olivetolic Acid |
| ORF | Open Reading Frames |
| PAVs | Presence/Absence Variations |
| PPFD | Photosynthetic Photon Flux Density |
| PK | Purple Kush Cannabis Cultivar |
| QTL | Quantitative Trait Loci |
| QQQ | Triple-Quadrupole Mass Spectrometry |
| qRT-PCR | Real time/Quantitative PCR—Polymerase Chain Reaction |
| RNA-Seq | RNA-Sequencing |
| ROS | Reactive Oxygen Species |
| SFE | Supercritical Fluid Extraction |
| SMRT | Single-Molecule Real-Time Sequencing |
| SNPs | Single Nucleotide Polymorphisms |
| SPE | Solid Phase Extraction |
| SPME | Solid Phase Micro Extraction |
| SVs | Structural Variants |
| THC | Tetrahydrocannabinol |
| THCA | Acidic Precursors of Tetrahydrocannabinol |
| WGCNA | Weighted Gene Co-expression Network Analysis |
| WGS | Whole Genome Shotgun Sequencing |
| Omic Technologies | Description | Reference |
|---|---|---|
| Metabolomics/GC-Cannabinoid synthase genotyping, linkage mapping and QTL analysis | Study about cannabinoids and terpenoids biosynthesis in cannabis | [38,62] |
| Metabolomics/Analytical methods available for cannabinoids analysis | Review—Cannabis metabolites | [32] |
| Metabolomics/GC-FID method | Definition of cannabis chemovars based on their terpenoid profile. The Effect of Light Spectrum on the cannabis morphology. | [46,50] |
| Metabolomics/Supercritical CO2 extraction of the cannabis inflorescence | Study of cannabinoids and terpenoids biosynthesis in cannabis | [40] |
| Metabolomics/GC-MS/LC-MS | Overview about methods for the chemical characterization of cannabis | [44] |
| Metabolomics/HRMS—LC-HRMS | Potential in the definition of cannabis chemovars of HRMS techniques | [60,61] |
| Metabolomics/SFE/SPE | Isolation of tetrahydrocannabinol from cannabis | [52] |
| Metabolomics/SPME/DLLME/LC-QQQ-MS | Physicochemical characterization of cannabis | [53] |
| Metabolomics/Analytical methods available for cannabinoids and terpenoids analysis | Review—Cannabinomics: Metabolomics applications in Cannabis. Terpenoids properties. | [20,42] |
| Metabolomics/Structural classification of phytocannabinoids | Study about phytocannabinoids | [39] |
| Metabolomics/GC-EIMS | Investigation about lipids extracted from cannabis seeds | [47] |
| Metabolomics/HPLC | Investigation about the major cannabinoids: CBD, CBG and THC. | [48,49] |
| Genomics/Illumina sequencing approach | The genome of cannabis. To assess the completeness and representivity of the ‘PK’, ‘FN’, and ‘CBDRx’ assemblies, Illumina sequences and 55 public whole-genome-sequenced samples were used | [5] |
| Genomics/Reference assemblies were annotated with mRNA sequencing (Iso-Seq) approach | Sequence and annotation of 42 cannabis genomes | [25] |
| Genomics/Draft genome sequence using PacBio single-molecule sequencing | A reference genome of wild cannabis | [27] |
| Genomics/PCR genotyping/SNP markers | Phylogenetic relationship, population genetics, and correlation with cannabis metabolites | [8] |
| Genomics/GWAS | Investigation of novel cannabis genetic variants responsible for cannabis complex traits | [68,69] |
| Genomics/whole-genome resequencing approach | A whole-genome resequencing of of 110 worldwide accessions | [64] |
| Transcriptomics/RNA-Seq approach | Study of the cannabis transcriptome. Transcriptomic applications investigated fibers cannabis development stages. Cannabis salt-responsive genes were also investigated | [24,77,79,80] |
| Transcriptomics/De novo transcriptome assembly pipeline and BLAST2GO tool | Definition of hight quality reference transcriptomes of two cannabis cultivars | [65] |
| Omic Technologies | Description | Reference |
|---|---|---|
| Metabolomics/GC, Transcriptomics/RNA-Seq e PCR | Quantification of the transcript levels of different cannabinoid synthase genes | [72,83,85] |
| Metabolomics/HPLC, Genomics/WGS and GBS | Investigation about cannabis ancestry | [15] |
| Metabolomics/SPME GC-MS and LC-MS, Transcriptomics/Rna-Seq | Investigation about trichomes changes during flower maturation in cannabis | [33] |
| Metabolomics/HPLC/GC, Transcriptomics/RNA-Seq | Study about nine cannabis cultivars having different basic characteristics | [43] |
| Metabolomics/HPLC, genomics/WGS, Transcriptomics/RNA-Seq | Study about the relation between CNVs and cannabinoids profile | [86] |
| Genomics/SNPs, Transcriptomics/RNA-sequencing pipeline based on CG technology | Investigation about sex chromosome evolution in cannabis | [67] |
| Genomics/PacBio, Transcriptomics/RNA sequencing based on Iso-Seq | Study about the cannabis sex evolution and the pathogen resistance | [65] |
| Genomics/Transcriptomics—Innovative approaches discussion, including GWAS, GS, pan-genomics | Review about cannabis genomics resources | [19] |
| Metabolomics, genomics and transcriptomics data are collected and structured, and available on a web site for researchers | “CannabisGDB”, a comprehensive multi-omics database | [91] |
| Metabolomics, Genomics and transcriptomics platforms. WGS approach in medical cannabis is discussed | Review on medicinal plants multi-omics applications | [82] |
| Metabolomics/GC, Genomics/WGS/Linkage mapping and QTL analysis | Comparation between Carmagnola and USO31 cannabis cultivars | [89] |
| Metabolomics/HPLC, Genomics/Nanopore technology/Linkage mapping and QTL analysis, Transcriptomics/RNA-Seq | Study about the CBDRx cannabis cultivar | [63] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sirangelo, T.M.; Ludlow, R.A.; Spadafora, N.D. Multi-Omics Approaches to Study Molecular Mechanisms in Cannabis sativa. Plants 2022, 11, 2182. https://doi.org/10.3390/plants11162182
Sirangelo TM, Ludlow RA, Spadafora ND. Multi-Omics Approaches to Study Molecular Mechanisms in Cannabis sativa. Plants. 2022; 11(16):2182. https://doi.org/10.3390/plants11162182
Chicago/Turabian StyleSirangelo, Tiziana M., Richard A. Ludlow, and Natasha D. Spadafora. 2022. "Multi-Omics Approaches to Study Molecular Mechanisms in Cannabis sativa" Plants 11, no. 16: 2182. https://doi.org/10.3390/plants11162182
APA StyleSirangelo, T. M., Ludlow, R. A., & Spadafora, N. D. (2022). Multi-Omics Approaches to Study Molecular Mechanisms in Cannabis sativa. Plants, 11(16), 2182. https://doi.org/10.3390/plants11162182