
Influence of Tree Species and Size on Bark Browsing by Large Wild Herbivores



Abstract
:1. Introduction
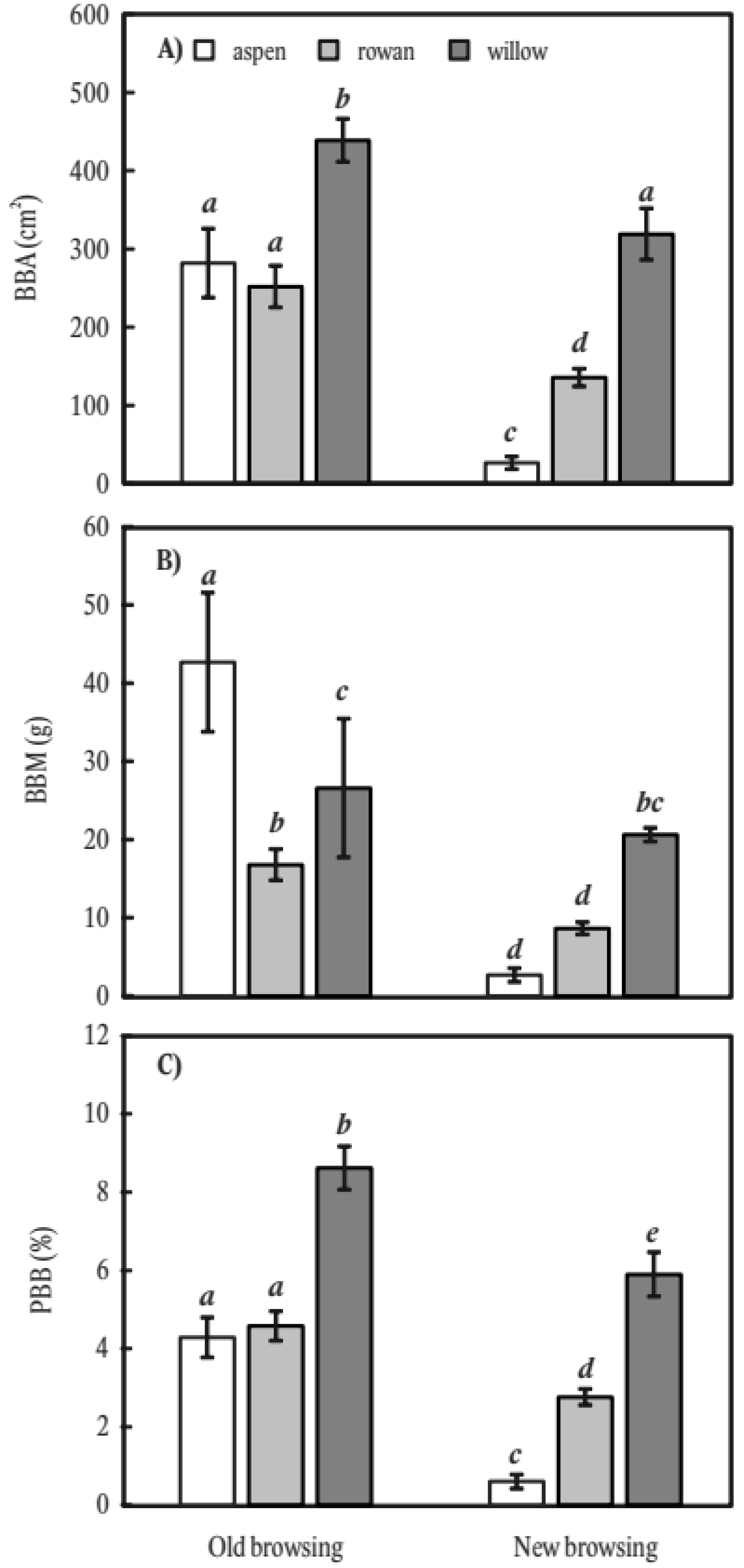
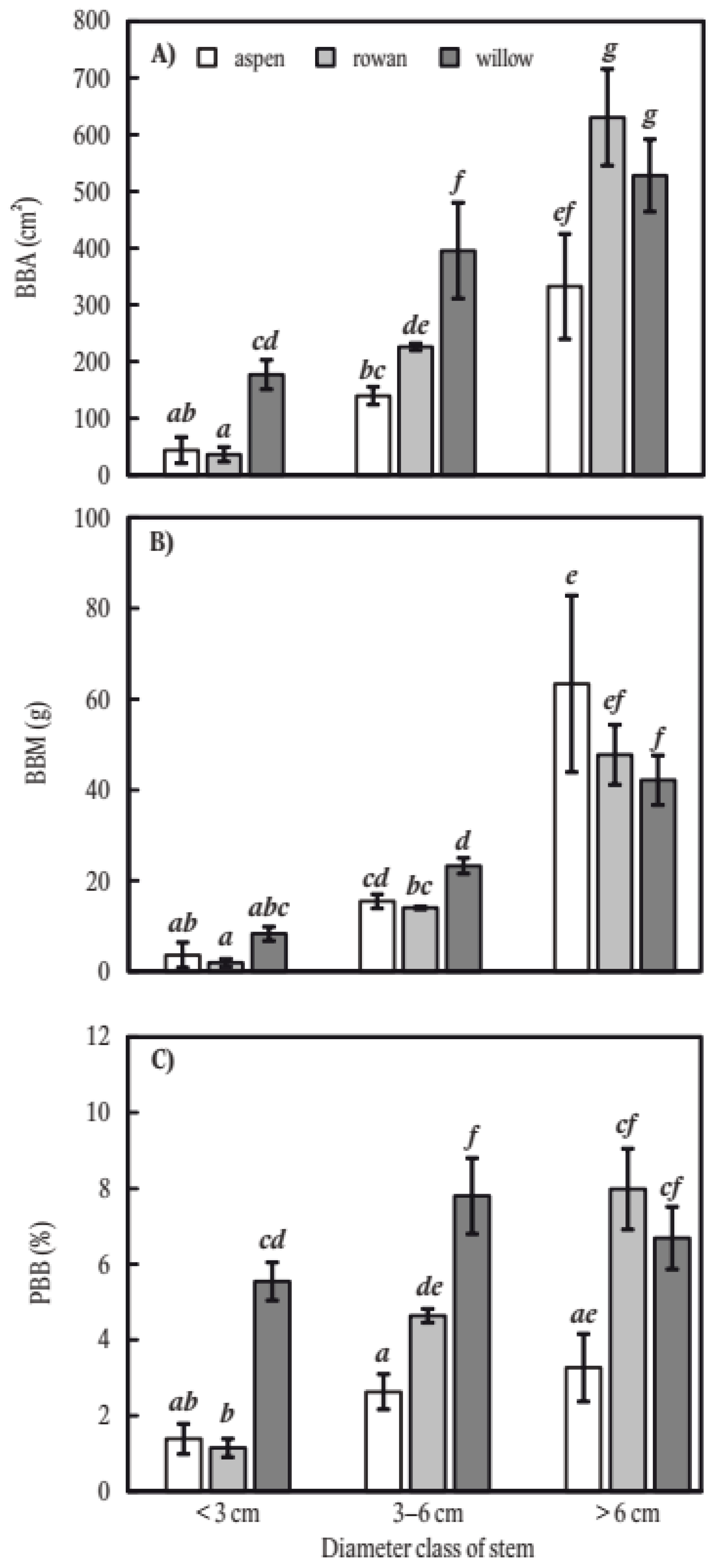
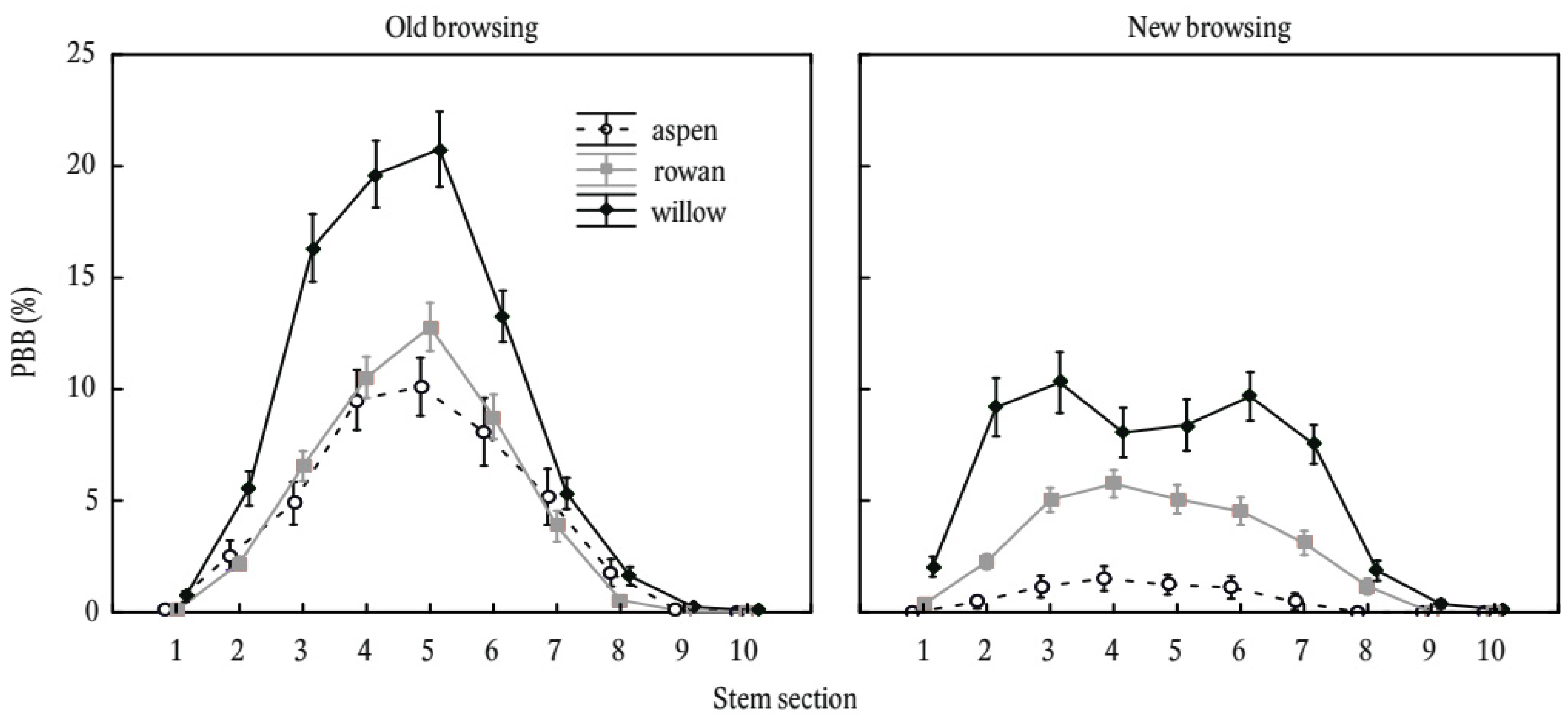
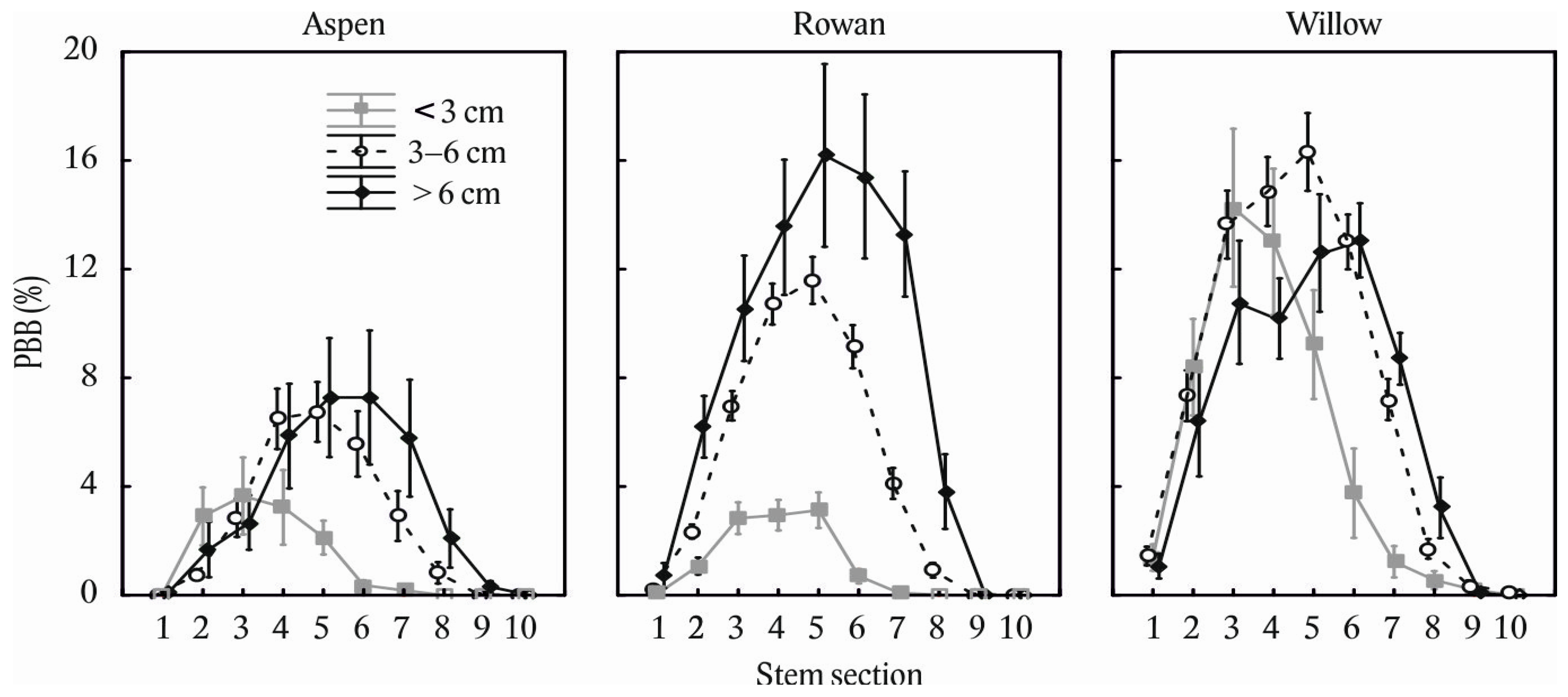
2. Results
3. Discussion
3.1. Main Factors Influencing Stem Bark Browsing
3.2. Stem Bark Browsing and Biological Control
4. Materials and Methods
4.1. Site and Plot Characteristics
4.2. Tree Measurement, Calculations, and Analyses
- r1 is a radius of the bottom end (cm);
- r2 is a radius of the top end (cm);
- ls is the length of the section (25 cm).
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lacher, T.E.; Davidson, A.D.; Fleming, T.H.; Gomez-Ruiz, E.P.; McCracken, G.F.; Owen-Smith, N.; Peres, C.A.; Vander Wall, S.B. The functional roles of mammals in ecosystems. J. Mammal 2019, 100, 942–964. [Google Scholar] [CrossRef] [Green Version]
- Smit, C.; Putman, R. Large herbivores as environmental engineers. In Ungulate Management in Europe: Problems and Practices; Putman, R., Apollonio, M., Andersen, R., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 260–283. [Google Scholar]
- Forbes, E.S.; Cushman, J.H.; Burkepile, D.E.; Young, T.P.; Klope, M.; Young, H.S. Synthesizing the effects of large, wild herbivory exclusion on ecosystem function. Funt. Ecol. 2018, 33, 1597–1610. [Google Scholar] [CrossRef] [Green Version]
- Soliveres, S.; van der Plas, F.; Manning, P.; Prati, D.; Gossner, M.M.; Renner, S.C.; Alt, F.; Arndt, H.; Baumgartner, V.; Binkenstein, J.; et al. Biodiversity at multiple tropic levels in needed for ecosystem multifunctionality. Nature 2016, 536, 456–459. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, D.; Prati, D.; Schall, P.; Ammer, C.; Fischer, M. Exclusion of large herbivores affects understorey shrub vegetation more than herb vegetation across 147 forest sites in three German regions. PLoS ONE 2019, 4, e0218741. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, J.I.; Jansen, P.A.; den Ouden, J.; Moktan, L.; Herdoiza, N.; Poorter, L. Above- and Below-ground Cascading Effects of Wild Ungulates in Temperate Forests. Ecosystems 2021, 24, 153–167. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin III, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 5, 3–35. [Google Scholar] [CrossRef]
- Levine, J.M.; Lambers, J.H. The Maintenance of Species Diversity. Nat. Educ. Know. 2010, 3, 59. [Google Scholar]
- Brockerhoff, E.G.; Barbaro, L.; Castagneyrol, B.; Forrester, D.I.; Gardiner, B.; González-Olabarria, J.R.; Lyver, P.O.; Meurisse, N.; Oxbrough, A.; Taki, A.; et al. Forest biodiversity, ecosystem functioning and the provision of ecosystem services. Biodivers. Conserv. 2017, 26, 3005–3035. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.J. Effect of reducing red deer Cervus elaphus density on browsing impact and growth of Scots pine Pinus sylvestris seedlings in semi-natural woodland in the Cairngorms, UK. Conserv. Evid. 2019, 14, 22–26. [Google Scholar]
- Ramirez, J.I.; Jansen, P.A.; den Ouden, J.; Goudzwaard, L.; Poorter, L. Long-term effects of wild ungulates on the structure, composition and succession of temperate forests. For. Ecol. Manag. 2019, 432, 478–488. [Google Scholar] [CrossRef]
- White, M.A. Long-term effects of deer browsing: Composition, structure and productivity in a northeast Minnesota old-growth forest. For. Ecol. Manag. 2012, 269, 222–228. [Google Scholar] [CrossRef]
- Kiffner, C.; Rössiger, E.; Trisl, O.; Schulz, R.; Rühe, F. Probability of recent bark stripping damage by red deer (Cervus elaphus) on Norway spruce (Picea abies) in as low mountain range in Germany—A preliminary analysis. Silva Fenn. 2008, 42, 125–134. [Google Scholar] [CrossRef]
- Arhipova, N.; Jansons, A.; Zaluma, A.; Gaitnieks, T.; Vasaitis, R. Bark stripping of Pinus contorta caused by moose and deer: Wounding patterns, discoloration of wood, and associated fungi. Can. J. For. Res. 2015, 45, 150615143947007. [Google Scholar] [CrossRef]
- Červený, J.; Hell, P.; Slamečka, J. Otto’s Encyclopedia Game Management; Otto’s Publisher: Praha, Czech Republic, 2010; p. 591. [Google Scholar]
- Valente, A.M.; Acevedo, P.; Figueiredo, A.M.; Fonseca, C.; Torres, R.T. Overabundant wild ungulate populations in Europe: Management with consideration of socio-ecological consequences. Mammal Rev. 2020, 51, 1–14. [Google Scholar] [CrossRef]
- Gebert, C.; Verheyden-Tixier, H. Variations of diet composition of Red Deer (Cervus elaphus L.) in Europe. Mammal Rev. 2001, 31, 189–201. [Google Scholar] [CrossRef]
- Spitzer, R.; Felton, A.; Landman, M.; Singh, N.J.; Widemo, F.; Cromsigt, J.P. Fifty years of European ungulate dietary studies: A synthesis. OIKOS 2020, 129, 1668–1680. [Google Scholar] [CrossRef]
- Kamler, J.; Homolka, M.; Barančeková, M.; Krojerová-Prokešová, J. Reduction of herbivory density as a tool for reduction of herbivore browsing on palatable tree species. Eur. J. For. Res. 2010, 129, 155–162. [Google Scholar] [CrossRef]
- Slanař, J.; Vacek, Z.; Vacek, S.; Bulušek, D.; Cukor, J.; Štefančík, I.; Král, J. Long-term transformation of submontane spruce-beech forests in the Jizerské hory Mts.: Dynamics of natural regeneration. Cent. Eur. For. J. 2017, 63, 212–224. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, Z.; Vacek, Z.; Vacek, S. Effect of game browsing on natural regeneration of European beech (Fagus sylvatica L.) forests in the Krušné hory Mts. (Czech Republic and Germany). Cent. Eur. For. J. 2021, 67, 166–180. [Google Scholar] [CrossRef]
- Borowski, Z.; Gil, W.; Barton, K.; Zajaczkowski, G.; Lukaszewicz, J.; Tittenbrun, A.; Radlinski, G. Density-related effect of red deer browsing on palatable and unpalatable tree species and forest regeneration dynamics. For. Ecol. Manag. 2021, 496, 119442. [Google Scholar] [CrossRef]
- Konôpka, B.; Šebeň, V.; Pajtík, J.; Shipley, L.A. Excluding Large Herbivores Reduced Norway Spruce Dominance and Supported Tree Species Richness in a Young, Naturally Regenerated Stand. Forests 2021, 12, 737. [Google Scholar] [CrossRef]
- Pajtík, J.; Konôpka, B.; Bošeľa, M.; Šebeň, V.; Kaštier, P. Modelling forage potential for red deer: A case study in post-disturbance young stands of rowan. Ann. For. Res. 2015, 58, 91–107. [Google Scholar] [CrossRef]
- Konôpka, B.; Pajtík, J.; Shipley, L.A. Intensity of deer browsing on young rowans differs between freshly-felled and standing individuals. For. Ecol. Manag. 2018, 429, 511–519. [Google Scholar] [CrossRef]
- Motta, R. Ungulate impact on rowan (Sorbus aucuparia L.) and Norway spruce (Picea abies (L.) Karst.) height structure in mountain forests in the eastern Italian Alps. For. Ecol. Manag. 2003, 181, 139–150. [Google Scholar] [CrossRef]
- Zakrisson, C.; Ericsson, G.; Edenius, L. Effects of browsing on recruitment and mortality of European aspen (Populus tremula L.). Scand. J. For. Res. 2006, 22, 324–332. [Google Scholar] [CrossRef]
- Myking, T.; Solberg, E.J.; Austrheim, G.; Speed, J.D.; Bohler, F.; Astrup, R.; Eriksen, R. Browsing of sallow (Salix caprea L.) and rowan (Sorbus aucuparia L.) in the context of life history strategies: A literature review. Eur. J. For. Res. 2013, 132, 399–409. [Google Scholar] [CrossRef]
- D’Aprile, D.; Vacchiano, G.; Meloni, F.; Garbarino, M.; Motta, R.; Ducoli, V.; Partel, P. Effects of Twenty Years of Ungulate Browsing on Forest Regeneration at Paneveggio Reserve, Italy. Forests 2020, 11, 612. [Google Scholar] [CrossRef]
- Vospernik, S. Probability of Bark Stripping Damage by Red Deer (Cervus elaphus) in Austria. Silva Fenn. 2006, 40, 589–601. [Google Scholar] [CrossRef] [Green Version]
- Ligot, G.; Gheysen, T.; Lehaire, F.; Hébert, J.; Licoppe, A.; Lejeune, P.; Brostaux, Y. Modelling recent bark stripping by red deer (Cervus elaphus) in South Belgium coniferous stands. Ann. For. Sci. 2013, 70, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Akashi, N.; Unno, A.; Uno, H. The browsing ratio as an index of impact of deer browsing on tree seedlings. Trees For. People 2022, 8, 100276. [Google Scholar] [CrossRef]
- Konôpka, B.; Pajtík, J.; Šebeň, V.; Merganičová, K. Modeling Bark Thickness and Bark Biomass on Stems of Four Broadleaved Tree Species. Plants 2022, 11, 1148. [Google Scholar] [CrossRef] [PubMed]
- Mansson, J.; Jarnemo, A. Bark-stripping on Norway spruce by red deer in Sweden: Level of damage and relation to tree characteristics. Scand. J. For. Res. 2012, 28, 117–125. [Google Scholar] [CrossRef]
- Candaele, R.; Lejeune, P.; Licoppe, A.; Malengreaux, C.; Brostaux, Y.; Morelle, K.; Latte, N. Mitigation of bark stripping on spruce: The need for red deer population control. Eur. J. For. Res. 2021, 140, 227–240. [Google Scholar] [CrossRef]
- Hahn, C.; Vospernik, S. Position, size, and spatial patterns of bark stripping wound inflicted by red deer (Cervus elaphus L.) on Norway spruce using generalized additive models in Austria. Ann. For. Sci. 2022, 79, 13. [Google Scholar] [CrossRef]
- Bergvall, U.A.; Co, M.; Bergström, R.; Sjöberg, P.J.R.; Waldebäck, M.; Turner, C. Anti-browsing effect of birch bark extract on fallow deer. Eur. J. For. Res. 2013, 132, 717–725. [Google Scholar] [CrossRef]
- Welch, D.; Staines, B.W.; Scott, D.; Catt, D.C. Bark stripping damage by red deer in Sitka spruce forest in Western Scotland II. Wound size and position. Forestry 1988, 61, 245–254. [Google Scholar] [CrossRef]
- Prien, S. Wildschäden im Wald. Ökologische Grundlagen und Integrierte Schutzmaßnahmen; Paul Parey: Berlin, Germany, 1997; 257p. [Google Scholar]
- Renaud, P.C.; Verheyden-Tixier, H.; Dumont, B. Damage to saplings by red deer (Cervus elaphus): Effect of foliage height and structure. For. Ecol. Manag. 2003, 181, 31–37. [Google Scholar] [CrossRef]
- Vacek, Z.; Cukor, J.; Linda, R.; Vacek, S.; Šimůnek, V.; Brichta, J.; Prokůpková, A. Bark stripping, the crucial factor affecting stem rot development and timber production of Norway spruce forests in Central Europe. For. Ecol. Manag. 2020, 474, 118360. [Google Scholar] [CrossRef]
- San-Miguel-Ayanz, J.; de Rigo, D.; Caudullo, G.; Houston Durant, T.; Mauri, A.; Tinner, W.; Ballian, D.; Beck, P.; Birks, H.J.B.; Eaton, E.; et al. European Atlas of Forest Tree Species; European Commission: Luxembourg, 2016; p. 200. [Google Scholar]
- Szwagrzyk, J.; Gazda, A.; Muter, E.; Pielech, R.; Szewczyk, J.; Zieba, A.; Zwijacz-Kozica, T.; Wierteloz, A.; Pachowicz, T.; Bodziarczyk, J. Effects of species and environmental factors on browsing frequency of young trees in mountain forests affected by natural disturbances. For. Ecol. Manag. 2020, 474, 118364. [Google Scholar] [CrossRef]
- Finďo, S.; Petráš, R. Ecological Principals of Forest Protection to Game Damage; National Forest Centre: Zvolen, Slovakia, 2007; p. 186. [Google Scholar]
- Mansson, J.; Roberge, J.M.; Edenius, L.; Bergström, R.; Nilsson, L.; Lidberg, M.; Komstedt, K.; Ericsson, G. Food plots as a habitat management tool: Forage productionand ungulate browsing in adjacent forest. Wildl. Biol. 2015, 21, 246–253. [Google Scholar] [CrossRef] [Green Version]
- Putman, R.J.; Staines, B.W. Supplementary winter feeding of wild red deer Cervus elaphus in Europe and North America: Justifications, feeding practice and effectiveness. Mammal Rev. 2004, 34, 258–306. [Google Scholar] [CrossRef]
- Royo, A.; Kramer, D.W.; Miller, K.V.; Nibbelink, N.P.; Stout, S.I. Spatio-temporal variation in foodscapes modifies deer browsing impact on vegetation. Landsc. Ecol. 2017, 32, 2281–2295. [Google Scholar] [CrossRef]
- Šebeň, V.; Kulla, L. Implementation Project for the Husárik Research-Demonstration Entity (Updated 1.2 Version); National Forest Centre—Forest Research Institute: Zvolen, Slovakia, 2011; p. 29. [Google Scholar]
- Hlásny, T.; Sitková, Z. Spruce Forests Decline in the Beskids; National Forest Centre: Zvolen, Slovakia, 2010; p. 182. [Google Scholar]
- Hančinský, L. Forest Types in Slovakia; Príroda: Bratislava, Slovak, 1972; p. 307. [Google Scholar]
- Kulla, L.; Sitková, Z. Reconstruction of Allochthones Spruce Stands; National Forest Centre: Zvolen, Slovakia, 2010; p. 208. [Google Scholar]
- Konôpka, J.; Kaštier, P.; Konôpka, B. Theoretical bases and practical measures to harmonise the interests of forestry and game management in Slovakia. Lesn. Časopis 2015, 61, 114–123. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Species | Characteristcs | Mean | S.D. | Minimum | Maximum | 25th Percentil | 75th Percentil |
|---|---|---|---|---|---|---|---|
| Common aspen | Tree height | 6.1 | 1.8 | 2.0 | 11.7 | 4.7 | 7.4 |
| Diameter d0 | 58.1 | 28.4 | 16.5 | 138.0 | 36.0 | 70.0 | |
| Common rowan | Tree height | 4.7 | 0.6 | 1.6 | 5.7 | 3.3 | 4.1 |
| Diameter d0 | 49.4 | 16.5 | 21.0 | 104.5 | 38.0 | 55.0 | |
| Goat willow | Tree height | 6.4 | 1.0 | 3.5 | 8.0 | 5.8 | 7.1 |
| Diameter d0 | 57.0 | 17.5 | 20.5 | 109.5 | 45.0 | 65.5 |
| Stem Section (cm) | Common Aspen | Common Rowan | Goat Willow | |||
|---|---|---|---|---|---|---|
| Old Browsing | New Browsing | Old Browsing | New Browsing | Old Browsing | New Browsing | |
| 0–25 | 4.3 (12.2) | 0 | 5.0 (9.2) | 11.6 (18.9) | 14.2 (27.9) | 27.5 (43.9) |
| 26–50 | 37.1 (38.9) | 5.7 (25.6) | 46.3 (27.9) | 42.1 (31.4) | 57.5 (49.0) | 51.7 (101.0) |
| 51–75 | 58.6 (49.9) | 10.0 (47.1) | 62.8 (58.3) | 56.2 (44.4) | 80.8 (102.0) | 55.0 (106.1) |
| 76–100 | 72.9 (80.7) | 13.1 (49.5) | 71.1 (80.2) | 57.0 (48.0) | 89.2 (113.6) | 53.3 (82.9) |
| 101–125 | 80.0 (84.5) | 12.9 (38.6) | 72.1 (97.3) | 48.9 (45.7) | 89.6 (123.6) | 56.7 (80.1) |
| 126–150 | 57.1 (101.7) | 10.0 (60.2) | 54.5 (90.7) | 47.9 (44.8) | 80.0 (91.4) | 64.2 (82.2) |
| 151–175 | 34.3 (113.9) | 4.3 (68.2) | 31.4 (74.1) | 33.9 (50.7) | 49.2 (61.3) | 57.5 (71.5) |
| 176–200 | 14.3 (90.3) | 0 | 9.9 (29.2) | 12.4 (58.2) | 19.2 (43.2) | 20.0 (51.8) |
| 201–225 | 4.3 (30) | 0 | 0.8 (13.8) | 0 | 4.2 (23.1) | 4.2 (46.3) |
| 226–250 | 0 | 0 | 0.8 (7.4) | 0 | 1.7 (18.4) | 0.8 (59.2) |
| Full stem profile | 87.1 (323.5) | 21.4 (123.4) | 77.7 (174.4) | 79.3 (320.8) | 97.5 (450.2) | 87.5 (368.2) |
| Factors vs. Indicators | BBA | BBM | PBB | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Df | F Value | p Value | Df | F Value | p Value | Df | F Value | p Value | |
| TS | 2 | 74.88 | <0.001 | 2 | 6.79 | <0.001 | 2 | 95.57 | <0.001 |
| YB | 1 | 233.70 | <0.001 | 1 | 312.76 | <0.001 | 1 | 142.31 | <0.001 |
| DC | 2 | 246.33 | <0.001 | 2 | 311.89 | <0.001 | 2 | 68.07 | <0.001 |
| SS | 9 | 121.58 | <0.001 | 9 | 90.05 | <0.001 | 9 | 99.10 | <0.001 |
| TSxYB | 2 | 37.41 | <0.001 | 2 | 114.82 | <0.001 | 2 | 12.21 | <0.001 |
| TSxDC | 4 | 17.17 | <0.001 | 4 | 14.52 | <0.001 | 4 | 15.21 | <0.001 |
| TSxSS | 18 | 6.65 | <0.001 | 18 | 4.35 | <0.001 | 18 | 7.96 | <0.001 |
| YBxDC | 2 | 63.57 | <0.001 | 2 | 118.66 | <0.001 | 2 | 26.84 | <0.001 |
| YBxSS | 9 | 43.38 | <0.001 | 9 | 44.48 | <0.001 | 9 | 26.92 | <0.001 |
| DCxSS | 18 | 19.63 | <0.001 | 18 | 23.13 | <0.001 | 18 | 10.43 | <0.001 |
| TSxYBxDC | 4 | 34.58 | <0.001 | 4 | 68.97 | <0.001 | 4 | 15.75 | <0.001 |
| TSxYBxSS | 18 | 3.29 | <0.001 | 18 | 7.6 | <0.001 | 18 | 1.86 | 0.015 |
| TSxDCxSS | 36 | 1.44 | 0.043 | 36 | 2.69 | <0.001 | 36 | 1.48 | 0.032 |
| YBxDCxSS | 18 | 13.11 | <0.001 | 18 | 18.7 | <0.001 | 18 | 6.22 | <0.001 |
| TSxYBxDCxSS | 36 | 3.99 | <0.001 | 36 | 5.29 | <0.001 | 36 | 2.51 | <0.001 |
| Factors vs. Indicators | BBA | BBM | PBB | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Df | F Value | p Value | Df | F Value | p Value | Df | F Value | p Value | |
| TS | 2 | 24.00 | <0.001 | 2 | 2.10 | 0.124 | 2 | 33.39 | <0.001 |
| YB | 1 | 74.91 | <0.001 | 1 | 96.64 | <0.001 | 1 | 46.50 | <0.001 |
| DC | 2 | 78.96 | <0.001 | 2 | 96.37 | <0.001 | 2 | 19.90 | <0.001 |
| TSxYB | 2 | 11.10 | <0.001 | 2 | 35.48 | <0.001 | 2 | 3.91 | 0.021 |
| TSxDC | 4 | 5.50 | <0.001 | 4 | 4.49 | 0.001 | 4 | 5.21 | <0.001 |
| YBxDC | 2 | 20.38 | <0.001 | 2 | 36.67 | <0.001 | 2 | 8.46 | <0.001 |
| TSxYBxDC | 4 | 11.9 | <0.001 | 4 | 21.31 | <0.001 | 4 | 5.39 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konôpka, B.; Šebeň, V.; Pajtík, J.; Shipley, L.A. Influence of Tree Species and Size on Bark Browsing by Large Wild Herbivores. Plants 2022, 11, 2925. https://doi.org/10.3390/plants11212925
Konôpka B, Šebeň V, Pajtík J, Shipley LA. Influence of Tree Species and Size on Bark Browsing by Large Wild Herbivores. Plants. 2022; 11(21):2925. https://doi.org/10.3390/plants11212925
Chicago/Turabian StyleKonôpka, Bohdan, Vladimír Šebeň, Jozef Pajtík, and Lisa A. Shipley. 2022. "Influence of Tree Species and Size on Bark Browsing by Large Wild Herbivores" Plants 11, no. 21: 2925. https://doi.org/10.3390/plants11212925