
Effects of Essential Oil Fumigation on Potato Sprouting at Room-Temperature Storage


Abstract
1. Introduction
2. Results and Discussion
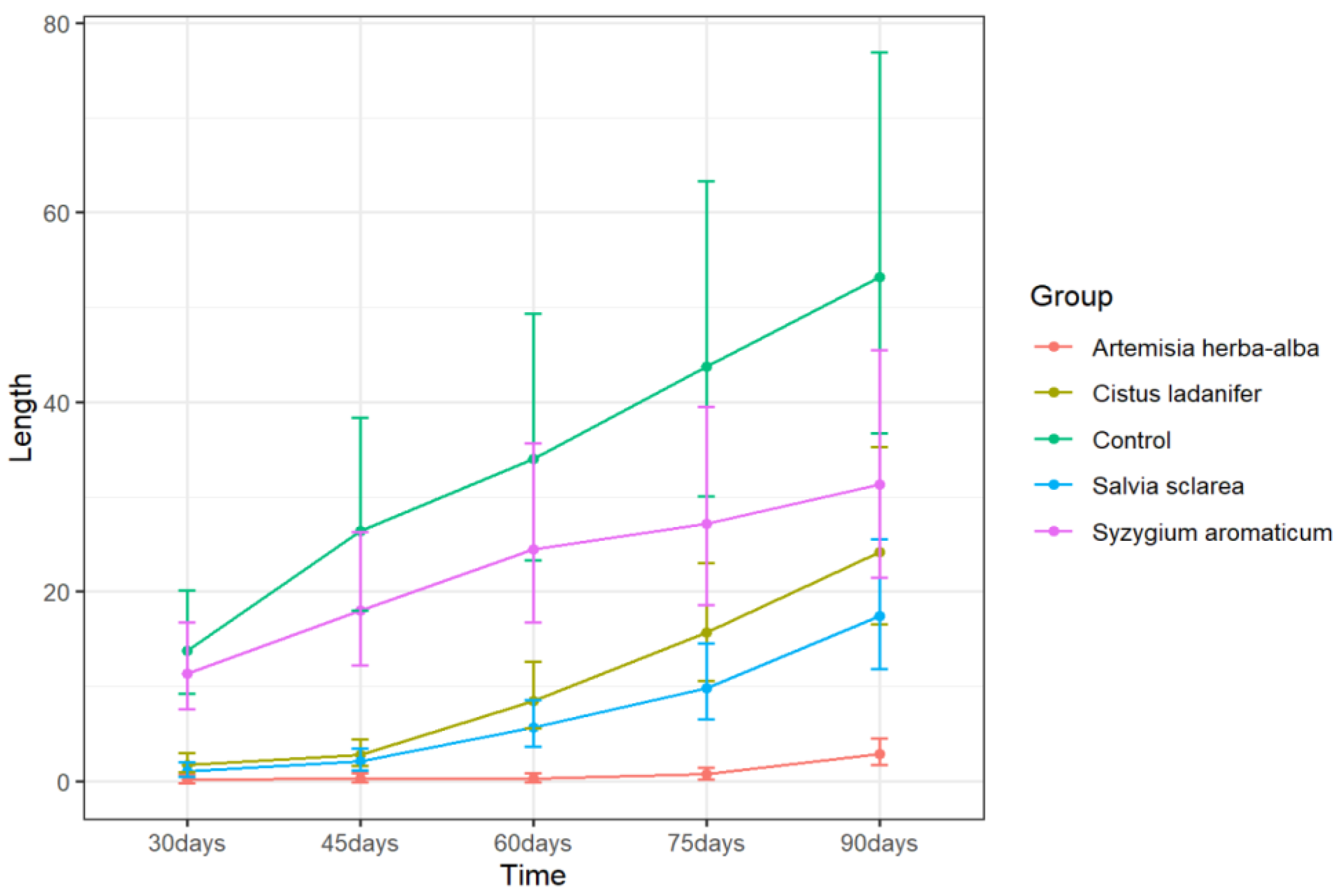
2.1. Effects of Essential Oils (EOs) on Sprout Length
2.2. Effects of Essential Oils on Number of Germinated Eyes
3. Materials and Methods
3.1. Plant Material
3.2. Experimental Materials
3.3. Gas Chromatography Mass Spectrometry Flame Ionization Detection (GC–MS–FID) Essential Oil Analysis
3.4. Experimental Design
3.5. Observations
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boivin, M.; Bourdeau, N.; Barnabé, S.; Desgagné-Penix, I. Sprout Suppressive Molecules Effective on Potato (Solanum tuberosum) Tubers during Storage: A Review. Am. J. Potato Res. 2020, 97, 451–463. [Google Scholar] [CrossRef]
- Devaux, A.; Goffart, J.-P.; Petsakos, A.; Kromann, P.; Gatto, M.; Okello, J.; Suarez, V.; Hareau, G. Global Food Security, Contributions from Sustainable Potato Agri-Food Systems. In The Potato Crop: Its Agricultural, Nutritional and Social Contribution to Humankind; Campos, H., Ortiz, O., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 3–35. ISBN 978-3-030-28683-5. [Google Scholar]
- FAO. The State of Food and Agriculture 2019; Food and Agriculture Organization of the United Nations: Rome, Italy, 2019; ISBN 978-92-5-131789-1. [Google Scholar]
- Shahbandeh, M. Potato Production Worldwide. 2019. Available online: https://www.statista.com/statistics/382174/global-potato-production/ (accessed on 15 May 2021).
- USDA NASS Press Release. Available online: https://www.nass.usda.gov/Statistics_by_State/Alaska/Publications/Current_News_Release/PT09_1.pdf (accessed on 18 October 2022).
- Burgos, G.; Zum Felde, T.; Andre, C.; Kubow, S. The Potato and Its Contribution to the Human Diet and Health. In The Potato Crop: Its Agricultural, Nutritional and Social Contribution to Humankind; Campos, H., Ortiz, O., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 37–74. ISBN 978-3-030-28683-5. [Google Scholar]
- Plant Production and Protection Division International Year of the Potato 2008—New Light on a Hidden Treasure: An End-of-Year Review; FAO: Rome, Italy, 2009; ISBN 978-92-5-106142-8.
- Paul, V.; Ezekiel, R.; Pandey, R. Sprout Suppression on Potato: Need to Look beyond CIPC for More Effective and Safer Alternatives. J. Food Sci. Technol. 2016, 53, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Sonnewald, S.; Sonnewald, U. Regulation of Potato Tuber Sprouting. Planta 2014, 239, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Suttle, J.C.; Campbell, M.A.; Olsen, N.L. Potato Tuber Dormancy and Postharvest Sprout Control. In Postharvest Ripening Physiology of Crops; CRC Press: Boca Raton, FL, USA, 2016; ISBN 978-0-429-16053-0. [Google Scholar]
- Visse-Mansiaux, M.; Soyeurt, H.; Herrera, J.M.; Torche, J.-M.; Vanderschuren, H.; Dupuis, B. Prediction of Potato Sprouting during Storage. Field Crops Res. 2022, 278, 108396. [Google Scholar] [CrossRef]
- Magdalena, G.; Dariusz, M. Losses during Storage of Potato Varieties in Relation to Weather Conditions during the Vegetation Period and Temperatures during Long-Term Storage. Am. J. Potato Res. 2018, 95, 130–138. [Google Scholar] [CrossRef]
- Sorce, C.; Lorenzi, R.; Parisi, B.; Ranalli, P. Physiological Mechanisms Involved in Potato (Solanum tuberosum) Tuber Dormancy and the Control of Sprouting by Chemical Suppressants. Acta Hortic. 2005, 684, 177–186. [Google Scholar] [CrossRef]
- Paul, V.; Ezekiel, R.; Pandey, R. Acrylamide in Processed Potato Products: Progress Made and Present Status. Acta Physiol. Plant 2016, 38, 276. [Google Scholar] [CrossRef]
- Wiltshire, J.J.J.; Cobb, A.H. A Review of the Physiology of Potato Tuber Dormancy. Ann. Appl. Biol. 1996, 129, 553–569. [Google Scholar] [CrossRef]
- Shukla, S.; Pandey, S.S.; Chandra, M.; Pandey, A.; Bharti, N.; Barnawal, D.; Chanotiya, C.S.; Tandon, S.; Darokar, M.P.; Kalra, A. Application of Essential Oils as a Natural and Alternate Method for Inhibiting and Inducing the Sprouting of Potato Tubers. Food Chem. 2019, 284, 171–179. [Google Scholar] [CrossRef]
- Authority (EFSA), E.F.S.; Arena, M.; Auteri, D.; Barmaz, S.; Bellisai, G.; Brancato, A.; Brocca, D.; Bura, L.; Byers, H.; Chiusolo, A.; et al. Peer Review of the Pesticide Risk Assessment of the Active Substance Chlorpropham. EFSA J. 2017, 15, e04903. [Google Scholar] [CrossRef]
- Kleinkopf, G.E.; Oberg, N.A.; Olsen, N.L. Sprout Inhibition in Storage: Current Status, New Chemistries and Natural Compounds. Am. J. Pot. Res. 2003, 80, 317. [Google Scholar] [CrossRef]
- Paul, V.; Ezekiel, R.; Pandey, R. Use of CIPC as a Potato Sprout Suppressant: Health and Environmental Concerns and Future Options. Qual. Assur. Saf. Crops Foods 2018, 10, 17–24. [Google Scholar] [CrossRef]
- National Potato Council Potato Statistical Yearbook. Available online: https://www.flipsnack.com/nationalpotatocouncil/2018-npc-statistical-yearbook.html (accessed on 29 April 2022).
- Briddon, A.; Stroud, G.P. Efficacy of Sprout Suppressants Used Alone, or in Combination, to Control Sprouting of Stored Potato; Sutton Bridge Crop Storage Research: Spalding, UK, 2019; p. 23. Available online: https://projectblue.blob.core.windows.net/media/Default/Potato%20knowledge%20library/S1043%202018-19%20Yr%202%20Report.pdf (accessed on 8 November 2022).
- Organic Trade Association U.S. Organic Sales Soar to New High of Nearly $62 Billion in 2020|OTA. Available online: https://ota.com/news/press-releases/21755 (accessed on 5 April 2022).
- Gómez-Castillo, D.; Cruz, E.; Iguaz, A.; Arroqui, C.; Vírseda, P. Effects of Essential Oils on Sprout Suppression and Quality of Potato Cultivars. Postharvest Biol. Technol. 2013, 82, 15–21. [Google Scholar] [CrossRef]
- Şanlı, A.; Karadogan, T. Carvone Containing Essential Oils as Sprout Suppressants in Potato (Solanum tuberosum L.) Tubers at Different Storage Temperatures. Potato Res. 2019, 62, 345–360. [Google Scholar] [CrossRef]
- Teper-Bamnolker, P.; Dudai, N.; Fischer, R.; Belausov, E.; Zemach, H.; Shoseyov, O.; Eshel, D. Mint Essential Oil Can Induce or Inhibit Potato Sprouting by Differential Alteration of Apical Meristem. Planta 2010, 232, 179–186. [Google Scholar] [CrossRef]
- El-Zaeddi, H.; Martínez-Tomé, J.; Calín-Sánchez, Á.; Burló, F.; Carbonell-Barrachina, Á.A. Volatile Composition of Essential Oils from Different Aromatic Herbs Grown in Mediterranean Regions of Spain. Foods 2016, 5, 41. [Google Scholar] [CrossRef]
- Saunders, S.; Harper, G. Integrating Alternative Sprout Suppressants for the Fresh Market; Sutton Bridge Crop Storage Research: Spalding, UK, 2019; Available online: https://projectblue.blob.core.windows.net/media/Default/Research%20Papers/Potatoes/11140043%20Interim%20Report_published%20Feb%2019.pdf (accessed on 8 November 2022).
- Visse-Manisaux, M.; Tallant, M.; Curty, F.; Schwärzel, R.; Brostaux, Y.; Dupuis, B. Storage of Processing Potato Varieties: The Post-CIPC Era. Rech. Agron. Suisse 2021, 12, 175–186. [Google Scholar] [CrossRef]
- Mohsen, H.; Ali, F. Essential Oil Composition of Artemisia herba-alba from Southern Tunisia. Molecules 2009, 14, 1585–1594. [Google Scholar] [CrossRef]
- Imelouane, B.; Bachiri, A.E.; Ankit, M.; Khedid, K.; Wathelet, J.P.; Amhamdi, H. Essential Oil Composition and Antimicrobial Activity of Artemisia herba-alba Asso Grown In Morocco. Banat. J. Biotechnol. 2011, 1, 48–55. [Google Scholar]
- El-Seedi, H.R.; Azeem, M.; Khalil, N.S.; Sakr, H.H.; Khalifa, S.A.M.; Awang, K.; Saeed, A.; Farag, M.A.; AlAjmi, M.F.; Pålsson, K.; et al. Essential Oils of Aromatic Egyptian Plants Repel Nymphs of the Tick Ixodes ricinus (Acari: Ixodidae). Exp. Appl. Acarol. 2017, 73, 139–157. [Google Scholar] [CrossRef]
- Caputo, L.; Nazzaro, F.; Souza, L.F.; Aliberti, L.; De Martino, L.; Fratianni, F.; Coppola, R.; De Feo, V. Laurus nobilis: Composition of Essential Oil and Its Biological Activities. Molecules 2017, 22, 930. [Google Scholar] [CrossRef] [PubMed]
- Fidan, H.; Stefanova, G.; Kostova, I.; Stankov, S.; Damyanova, S.; Stoyanova, A.; Zheljazkov, V.D. Chemical Composition and Antimicrobial Activity of Laurus nobilis L. Essential Oils from Bulgaria. Molecules 2019, 24, E804. [Google Scholar] [CrossRef] [PubMed]
- Ouedrhiri, W.; Balouiri, M.; Bouhdid, S.; Harki, E.H.; Moja, S.; Greche, H. Antioxidant and Antibacterial Activities of Pelargonium asperum and Ormenis mixta Essential Oils and Their Synergistic Antibacterial Effect. Environ. Sci. Pollut. Res. 2018, 25, 29860–29867. [Google Scholar] [CrossRef]
- Zheljazkov, V.D.; Callahan, A.; Cantrell, C.L. Yield and Oil Composition of 38 Basil (Ocimum basilicum L.) Accessions Grown in Mississippi. J. Agric. Food Chem. 2008, 56, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Aćimović, M.; Kiprovski, B.; Rat, M.; Sikora, V.; Popović, V.; Koren, A.; Brdar-Jokanović, M. Salvia sclarea: Chemical Composition and Biological Activity. J. Agron. Technol. Eng. Manag. (JATEM) 2018, 1, 18–28. [Google Scholar]
- Verdeguer, M.; Blázquez, M.A.; Boira, H. Chemical Composition and Herbicidal Activity of the Essential Oil from a Cistus ladanifer L. Population from Spain. Nat. Prod. Res. 2012, 26, 1602–1609. [Google Scholar] [CrossRef]
- Zheljazkov, V.D.; Micalizzi, G.; Yilma, S.; Cantrell, C.L.; Reichley, A.; Mondello, L.; Semerdjieva, I.; Radoukova, T. Melissa officinalis L. as a Sprout Suppressor in Solanum tuberosum L. and an Alternative to Synthetic Pesticides. J. Agric. Food Chem. 2022, 70, 14205–14219. [Google Scholar] [CrossRef]
- Derwich, E.; Benziane, Z.; Boukir, A. Chemical Composition and In Vitro Antibacterial Activity of the Essential Oil of Cedrus atlantica. Int. J. Agric. Biol. 2010, 12, 381–385. [Google Scholar]
- Duman, E.; Cinar, O.; Ozcan, M.M. Physico-Chemical Properties, Fatty Acid Composition, Essential Oil Composition and Heavy Metal Contents of Raw and Roasted Coffee Bean and Oils. Z. Fuer Arznei-Und. Gewuerzpflanzen 2017, 22, 154–159. [Google Scholar]
- Fernandez, X.; Lizzani-Cuvelier, L.; Loiseau, A.-M.; Périchet, C.; Delbecque, C. Volatile Constituents of Benzoin Gums: Siam and Sumatra. Part 1. Flavour Fragr. J. 2003, 18, 328–333. [Google Scholar] [CrossRef]
- Fraternale, D.; Flamini, G.; Ricci, D. Essential Oil Composition and Antimicrobial Activity of Angelica archangelica L. (Apiaceae) Roots. J. Med. Food 2014, 17, 1043–1047. [Google Scholar] [CrossRef]
- Liyanage, T.; Madhujith, T.; Wijesinghe, K.G.G. Comparative Study on Major Chemical Constituents in Volatile Oil of True Cinnamon (Cinnamomum verum Presl. Syn. C. zeylanicum Blum.) and Five Wild Cinnamon Species Grown in Sri Lanka. Trop. Agric. Res. 2017, 28, 270. [Google Scholar] [CrossRef]
- Anass, E.; El Amrani, A.; Eddine, J.J. Effect of the Parts of Plant Material (Flowers and Leaves) on Essential Oil Chemical Composition of Ormenis mixta from Morocco. J. Essent. Oil Bear. Plants 2015, 18, 398–408. [Google Scholar] [CrossRef]
- Calva-Estrada, S.J.; Utrilla-Vázquez, M.; Vallejo-Cardona, A.; Roblero-Pérez, D.B.; Lugo-Cervantes, E. Thermal Properties and Volatile Compounds Profile of Commercial Dark-Chocolates from Different Genotypes of Cocoa Beans (Theobroma cacao L.) from Latin America. Food Res. Int. 2020, 136, 109594. [Google Scholar] [CrossRef]
- Orav, A.; Raal, A.; Arak, E. Essential Oil Composition of Pimpinella anisum L. Fruits from Various European Countries. Nat. Prod. Res. 2008, 22, 227–232. [Google Scholar] [CrossRef]
- Yalçın, H.; Anık, M.; Şanda, M.A.; Çakır, A. Gas Chromatography/Mass Spectrometry Analysis of Laurus nobilis Essential Oil Composition of Northern Cyprus. J. Med. Food 2007, 10, 715–719. [Google Scholar] [CrossRef]
- Olsen, N.; Frazier, M.J.; Kleinkopf, G. Potato Sprout Suppression from Clove Oil. Available online: https://www.uidaho.edu/~/media/UIdaho-Responsive/Files/cals/programs/potatoes/Storage/potato-sprout-suppression-from-clove-oil.ashx (accessed on 8 November 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 5 April 2022).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 5 April 2022).
- Kassambara, A. Ggpubr: ‘ggplot2’ Based Publication Ready Plots 2020. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 5 April 2022).
- Kassambara, A. Rstatix: Pipe-Friendly Framework for Basic Statistical Tests. 2021. Available online: https://CRAN.R-project.org/package=rstatix (accessed on 5 April 2022).
- Lenth, R.V. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. 2022. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 5 April 2022).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models. 2022. Available online: https://CRAN.R-project.org/package=nlme (accessed on 5 April 2022).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment vs. Control | 30 Days | 45 Days | 60 Days | 75 Days | 90 Days |
|---|---|---|---|---|---|
| Cedrus atlantica | 1.000 | 1.000 | 1.000 | 0.999 | 0.999 |
| Apium graveolens | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Matricaria recutita | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Ormenis mixta | 0.002 ** | 0.812 | 0.998 | 0.996 | 0.930 |
| Anthemis nobilis | 0.705 | 0.940 | 1.000 | 1.000 | 0.991 |
| Cinnamomum zeylanicum (bark) | <0.001 *** | 0.002 ** | 0.445 | 0.472 | 0.284 |
| Cinnamomum zeylanicum (leaf) | 1.000 | 1.000 | 1.000 | 1.000 | 0.999 |
| Cistus ladanifer | <0.001 *** | <0.001 *** | 0.002 ** | 0.259 | 0.528 |
| Salvia sclarea | <0.001 *** | <0.001 *** | <0.001 *** | 0.009 ** | 0.066 |
| Syzygium aromaticum | 1.000 | 0.939 | 1.000 | 0.996 | 0.966 |
| Theobroma cacao | 0.936 | 1.000 | 1.000 | 1.000 | 1.000 |
| Coffea canephora | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Copaifera langsdorfi | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Pimenta officinalis | 0.995 | 1.000 | 1.000 | 1.000 | 0.993 |
| Amyris balsamifera | 1.000 | 1.000 | 1.000 | 1.000 | 0.997 |
| Angelica archangelica | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Pimpinella anisum | 1.000 | 1.000 | 1.000 | 0.999 | 0.991 |
| Artemisia herba-alba | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** |
| Ocimum basilicum | 0.002 ** | 0.522 | 0.999 | 0.999 | 0.989 |
| Laurus nobilis | <0.001 *** | <0.001 *** | <0.001 *** | 0.067 | 0.411 |
| Styrax tonkinensis | 0.999 | 1.000 | 1.000 | 1.000 | 1.000 |
| 30 Days | 45 Days | 60 Days | 75 Days | 90 Days | |
|---|---|---|---|---|---|
| Control | 13.71 ± 2.64 a | 26.38 ± 4.92 a | 33.98 ± 6.29 a | 43.71 ± 8.04 a | 53.20 ± 9.74 a |
| Ormenis mixta | 4.61 ± 1.00 b | 16.88 ± 3.21 ab | 23.24 ± 4.36 ab | 27.15 ± 5.06 ab | 29.99 ± 5.57 a |
| Cinnamomum zeylanicum (bark) | 3.99 ± 0.89 b | 9.46 ± 1.88 b | 15.69 ± 3.00 abc | 17.76 ± 3.37 ab | 21.44 ± 4.03 a |
| Cistus ladanifer | 1.73 ± 0.49 bc | 2.78 ± 0.68 c | 8.43 ± 1.69 bcde | 15.70 ± 3.00 ab | 24.18 ± 4.52 a |
| Salvia sclarea | 1.05 ± 0.36 cd | 2.05 ± 0.54 c | 5.67 ± 1.20 ce | 9.80 ± 1.94 bc | 17.40 ± 3.31 a |
| Artemisia herba-alba | 0.11 ± 0.20 de | 0.25 ± 0.22 d | 0.25 ± 0.22 f | 0.71 ± 0.30 d | 2.83 ± 0.69 b |
| Ocimum basilicum | 4.63 ± 1.01 b | 15.40 ± 2.94 ab | 23.44 ± 4.39 ad | 29.02 ± 5.40 ac | 33.17 ± 6.14 a |
| Laurus nobilis | 0.51 ± 0.27 ce | 1.13 ± 0.38 cd | 4.63 ± 1.01 e | 12.69 ± 2.46 ac | 22.92 ± 4.30 a |
| No. | Compound Name | Retention Time | Calculated KI | Actual KI | Identified | Area% |
|---|---|---|---|---|---|---|
| 1 | tricyclene | 5.755 | 924 | 926 | Kovat, NIST, Adams, Commercial Standard | 0.075 |
| 2 | camphene | 6.465 | 948 | 954 | Kovat, NIST, Adams, Commercial Standard | 1.294 |
| 3 | sabinene | 7.152 | 972 | 975 | Kovat, NIST, Adams, Commercial Standard | 1.952 |
| 4 | α-terpinene | 8.519 | 1013 | 1017 | Kovat, NIST, Adams, Commercial Standard | 0.101 |
| 5 | p-cymene | 8.782 | 1022 | 1024 | Kovat, NIST, Adams, Commercial Standard | 0.487 |
| 6 | eucalyptol | 9.023 | 1030 | 1031 | Kovat, NIST, Adams, Commercial Standard | 0.365 |
| 7 | γ-terpinene | 10.013 | 1059 | 1059 | Kovat, NIST, Adams, Commercial Standard | 0.232 |
| 8 | (-)-α-thujone | 12.038 | 1112 | 1102 | Kovat, NIST, Adams, Commercial Standard | 63.606 |
| 9 | β-thujone | 12.353 | 1120 | 1114 | Kovat, NIST, Adams, Commercial Standard | 8.523 |
| 10 | unknown | 12.427 | - | - | - | 0.799 |
| 11 | trans-pinocarveol | 13.263 | 1142 | 1139 | Kovat, NIST, Adams [29] | 0.395 |
| 12 | (R)-(+)-camphor | 13.434 | 1146 | 1146 | Kovat, NIST, Adams, Commercial Standard | 6.87 |
| 13 | sabina ketone | 13.938 | 1158 | 1159 | Kovat, NIST, Adams, References [29,30] | 0.283 |
| 14 | endo-borneol | 14.378 | 1168 | 1169 | Kovat, NIST, Adams, Commercial Standard | 0.337 |
| 15 | terpinen-4-ol | 14.819 | 1177 | 1177 | Kovat, NIST, Adams, Commercial Standard | 0.396 |
| 16 | myrtenal | 15.529 | 1192 | 1195 | Kovat, NIST, Adams, Commercial Standard | 0.14 |
| 17 | cuminaldehyde | 17.405 | 1239 | 1241 | Kovat, NIST, Adams, Commercial Standard | 0.314 |
| 18 | piperitone | 18.029 | 1254 | 1252 | Kovat, NIST, Adams, Commercial Standard | 0.123 |
| 19 | germacrene D | 27.485 | 1481 | 1481 | Kovat, NIST, Adams, Commercial Standard [29] | 0.133 |
| 20 | unknown | 38.184 | - | - | - | 0.706 |
| 21 | unknown | 41.588 | - | - | - | 0.689 |
| 22 | hexadecanoic acid | 45.214 | 1973 | 1959 | Kovat, NIST, Adams [31] | 12.181 |
| Essential Oil | Effective | Somewhat Effective | References | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1,8-Cineoles | Terpenin-4-ol | Geranyl Acetate | 2-Phenylethanol | Nonanol | Methyl Benzoate | Camphor | α-Pinene | α-Phellandrene | Trans-Anethole | ||
| Cedrus atlantica | x | [39] | |||||||||
| Ormenis mixta | x | x | [34,44] | ||||||||
| Cinnamomum zeylanicum(bark) | x | [43] | |||||||||
| Cinnamomum zeylanicum (leaf) | x | [43] | |||||||||
| Cistus ladanifer | x | x | [37] | ||||||||
| Salvia sclarea | x | [36] | |||||||||
| Theobroma cacao | x | [45] | |||||||||
| Coffea canephora | x | [40] | |||||||||
| Angelica archangelica | x | x | [42] | ||||||||
| Pimpinella anisum | x | [46] | |||||||||
| Ocimum basilicum | x | x | [35] | ||||||||
| Laurus nobilis | x | x | x | [32,47] | |||||||
| Styrax tonkinensis | x | [41] | |||||||||
| Treatment vs. Control | 30 Days | 45 Days | 60 Days | 75 Days | 90 Days |
|---|---|---|---|---|---|
| Cedrus atlantica | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Apium graveolens | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Matricaria recutita | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Ormenis mixta | 0.995 | 1.000 | 1.000 | 1.000 | 1.000 |
| Anthemis nobilis | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Cinnamomum zeylanicum (bark) | 0.013 * | 0.022 * | 0.001 ** | 0.075 | 0.349 |
| Cinnamomum zeylanicum (leaf) | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Cistus ladanifer | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Salvia sclarea | 0.978 | 1.000 | 1.000 | 1.000 | 1.000 |
| Syzygium aromaticum | 1.000 | 0.995 | 0.917 | 0.938 | 0.996 |
| Theobroma cacao | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Coffea canephora | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Copaifera langsdorfi | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Pimenta officinalis | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Amyris balsamifera | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Angelica archangelica | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Pimpinella anisum | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Artemisia herba-alba | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** |
| Ocimum basilicum | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Laurus nobilis | <0.001 *** | 0.007 ** | 1.000 | 1.000 | 1.000 |
| Styrax tonkinensis | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| 30 Days | 45 Days | 60 Days | 75 Days | 90 Days | |
|---|---|---|---|---|---|
| Control | 9.80 ± 0.86 a | 11.40 ± 0.86 a | 11.60 ± 0.86 a | 11.87 ± 0.86 a | 11.87 ± 0.86 a |
| Cinnamomum zeylanicum (bark) | 4.67 ± 0.86 b | 5.80 ± 0.86 b | 5.87 ± 0.86 b | 7.00 ± 0.86 bc | 7.00 ± 0.86 bc |
| Artemisia herba-alba | 0.13 ± 0.86 b | 0.40 ± 0.86 c | 0.46 ± 0.86 c | 1.53 ± 0.86 c | 4.00 ± 0.86 c |
| Laurus nobilis | 0.87 ± 0.86 b | 5.73 ± 0.86 b | 10.13 ± 0.86 ab | 10.73 ± 0.86 ab | 10.93 ± 0.86 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thoma, J.L.; Cantrell, C.L.; Zheljazkov, V.D. Effects of Essential Oil Fumigation on Potato Sprouting at Room-Temperature Storage. Plants 2022, 11, 3109. https://doi.org/10.3390/plants11223109
Thoma JL, Cantrell CL, Zheljazkov VD. Effects of Essential Oil Fumigation on Potato Sprouting at Room-Temperature Storage. Plants. 2022; 11(22):3109. https://doi.org/10.3390/plants11223109
Chicago/Turabian StyleThoma, Jena L., Charles L. Cantrell, and Valtcho D. Zheljazkov. 2022. "Effects of Essential Oil Fumigation on Potato Sprouting at Room-Temperature Storage" Plants 11, no. 22: 3109. https://doi.org/10.3390/plants11223109
APA StyleThoma, J. L., Cantrell, C. L., & Zheljazkov, V. D. (2022). Effects of Essential Oil Fumigation on Potato Sprouting at Room-Temperature Storage. Plants, 11(22), 3109. https://doi.org/10.3390/plants11223109