
The Effects of Elevated Tropospheric Ozone on Carbon Fixation and Stable Isotopic Signatures of Durum Wheat Cultivars with Different Biomass and Yield Stability

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Biomass and Yield Variations Induced by Ozone
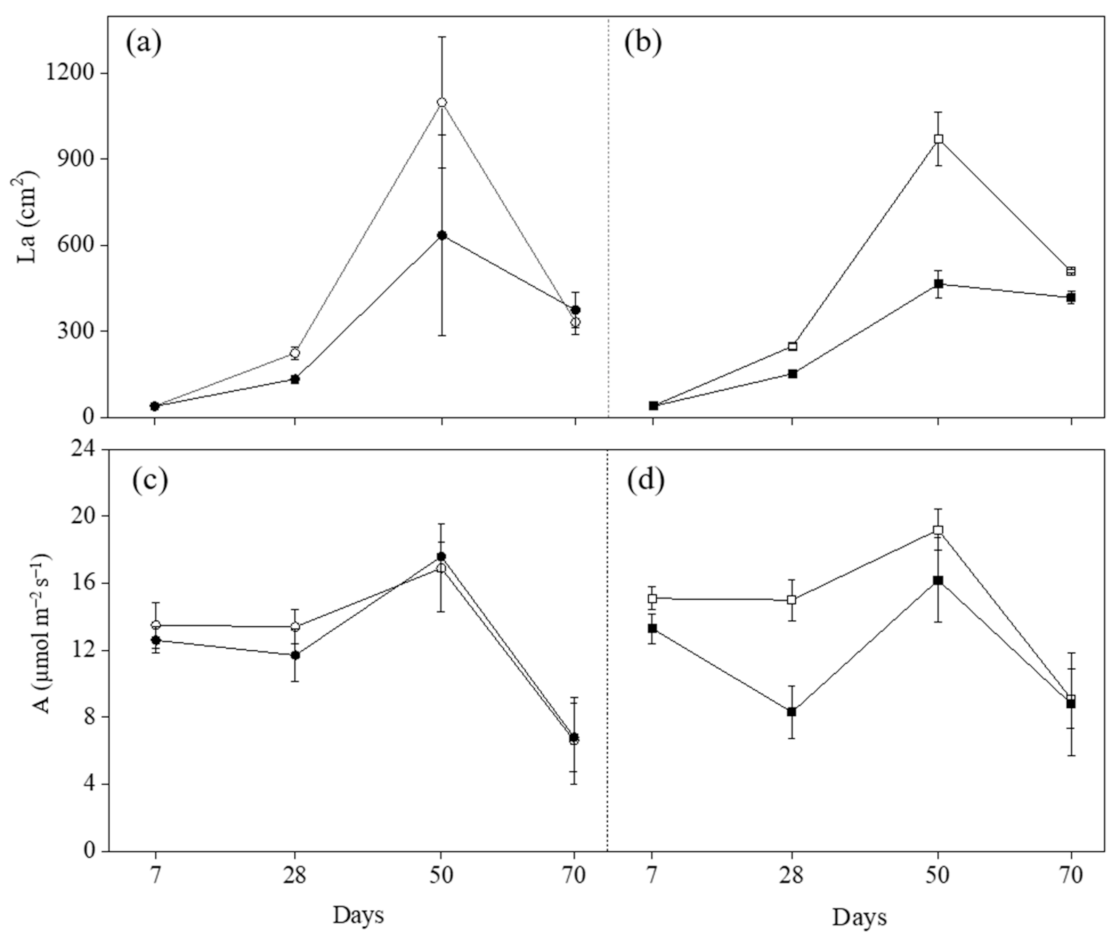
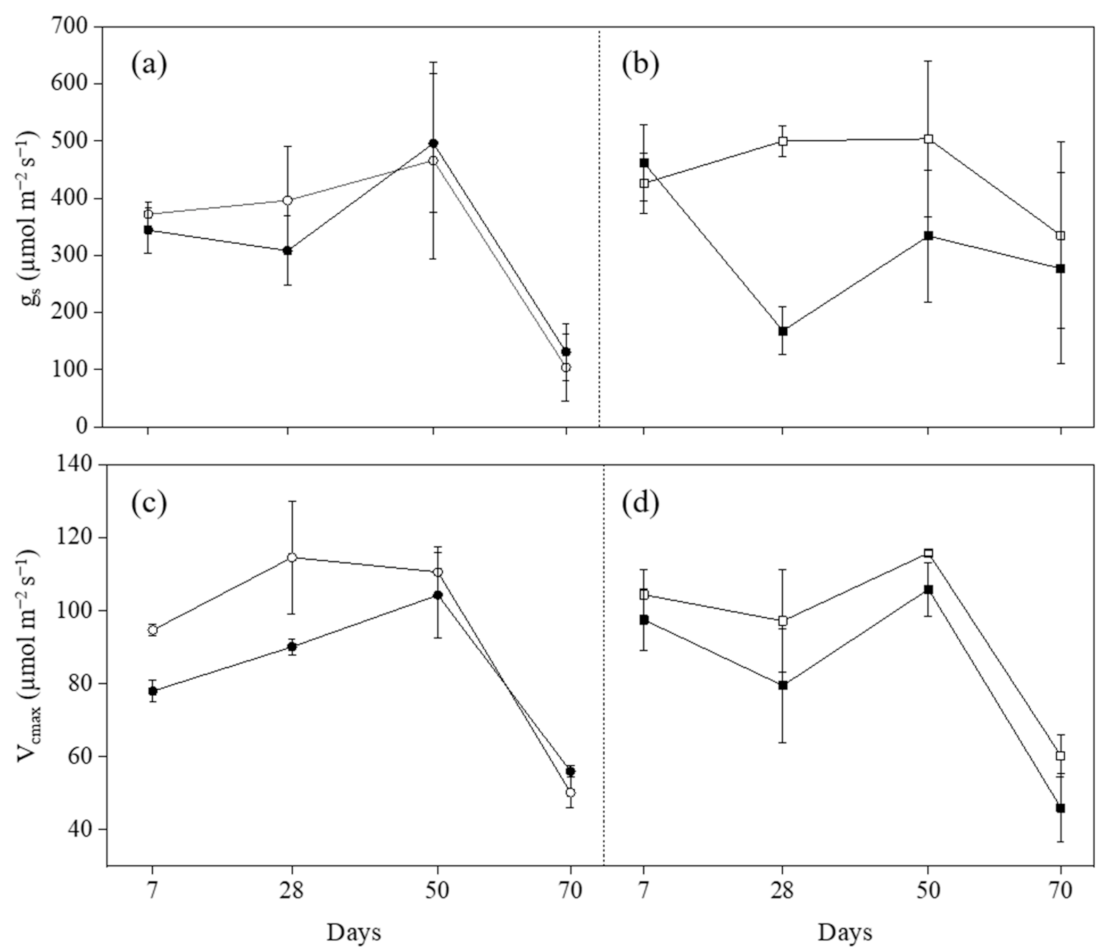
2.2. Leaf Area (La) and Ecophysiological Parameters
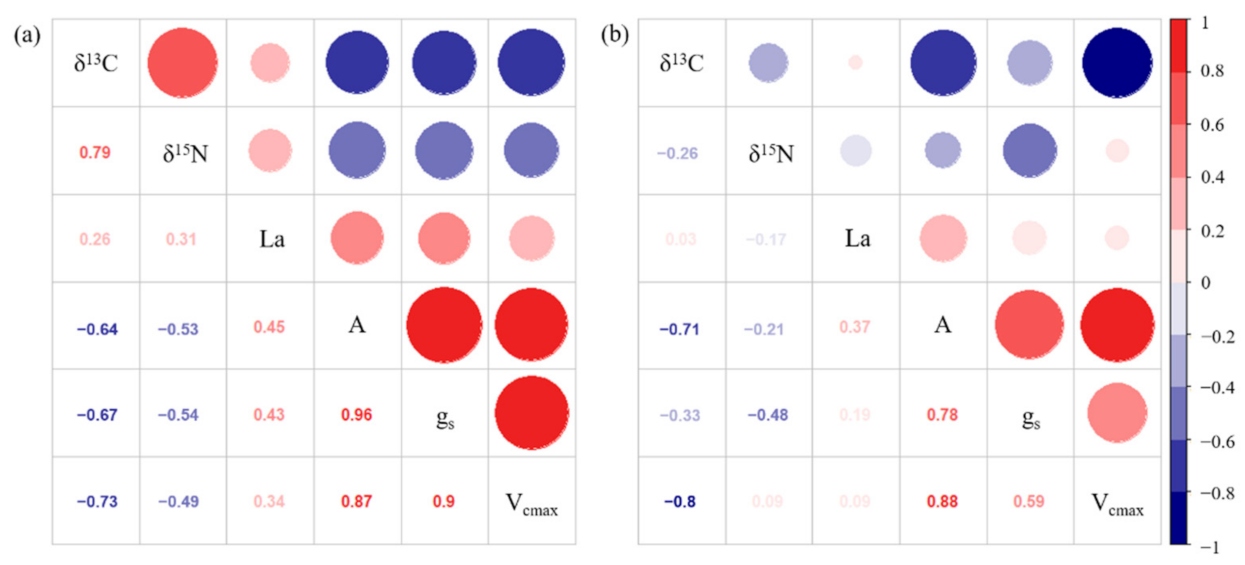
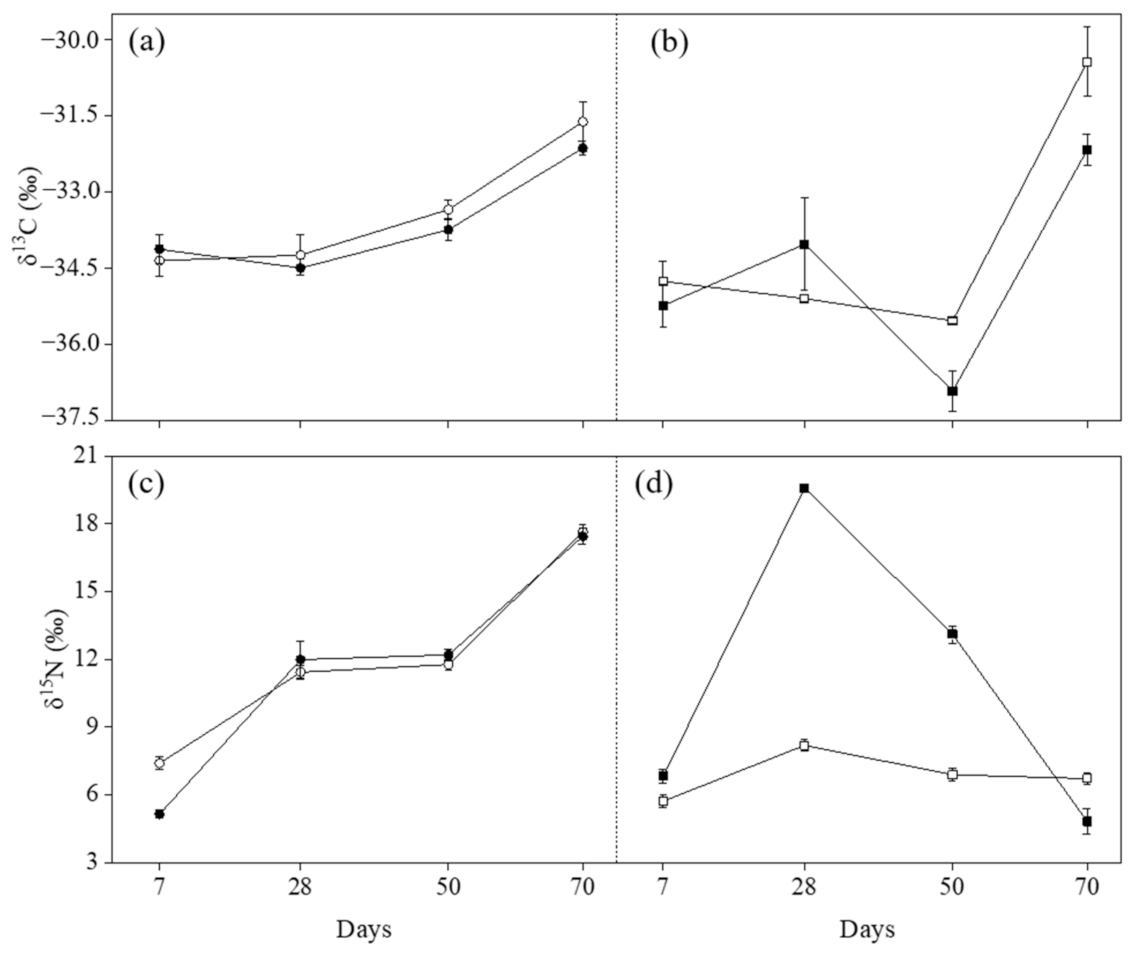
2.3. δ13C Values
2.4. δ15N Values
3. Materials and Methods
3.1. Plant Material and Ozone Exposure
3.2. Leaf Area (La) and Ecophysiological Analyses
3.3. δ13C and δ15N
3.4. Determinations of Biomass and Yield Parameters
3.5. Statistics
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Monks, P.S.; Archibald, A.T.; Colette, A.; Cooper, O.; Coyle, M.; Derwent, R.; Fowler, D.; Granier, C.; Law, K.S.; Mills, G.E.; et al. Tropospheric ozone and its precursors from the urban to the global scale from air quality to short-lived climate forcer. Atmos. Chem. Phys. 2015, 15, 8889–8973. [Google Scholar] [CrossRef] [Green Version]
- Lefohn, A.S.; Malley, C.S.; Smith, L.; Wells, B.; Hazucha, M.; Simon, H.; Naik, V.; Mills, G.; Schultz, M.G.; Paoletti, E.; et al. Tropospheric ozone assessment report: Global ozone metrics for climate change, human health, and crop/ecosystem research. Elem. Sci. Anthr. 2018, 6, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sage, R.F. Global change biology: A primer. Glob. Chang. Biol. 2020, 26, 3–30. [Google Scholar] [CrossRef] [PubMed]
- Field, C.B.; Barros, V.R.; Mach, K.J.; Mastrandrea, M.D.; Aalst, M.V.; Adger, W.N.; Arent, D.J.; Barnett, J.; Betts, R.; Bilir, T.E.; et al. International Panel on Climate Change (IPCC) Climate change 2014: Impacts, adaptation, and vulnerability. Part A: Global and sectoral aspects. In Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014; pp. 1–32. [Google Scholar]
- Yang, N.; Cotrozzi, L.; Zhang, Y.; Wang, X.; Ouyang, Z.; Zheng, F. First defense mechanisms of ornamental species suitable for urban green spaces in China exposed to ozone. Urban For. Urban Green. 2018, 36, 68–75. [Google Scholar] [CrossRef]
- Cotrozzi, L.; Pellegrini, E.; Nali, C.; Lorenzini, G. Climate change, ozone and plant life. Agrochimica 2019, 62, 181–188. [Google Scholar]
- Verstraeten, W.W.; Neu, J.L.; Williams, J.E.; Bowman, K.W.; Worden, J.R.; Boersma, K.F. Rapid increases in tropospheric ozone production and export from China. Nat. Geosci. 2015, 8, 690–695. [Google Scholar] [CrossRef]
- Ochoa-Hueso, R.; Munzi, S.; Alonso, R.; Arroniz-Crespo, M.; Avila, A.; Bermejo, V.; Bobbink, R.; Branquinho, C.; Concostrina-Zubiri, L.; Cruz, C.; et al. Ecological impacts of atmospheric pollution and interactions with climate change in terrestrial ecosystems of the Mediterranean Basin: Current research and future directions. Environ. Pollut. 2017, 227, 194–206. [Google Scholar] [CrossRef] [Green Version]
- Ridley, D.A.; Cain, M.; Methven, J.; Arnold, S.R. Sensitivity of tropospheric ozone to chemical kinetic uncertainties in air masses influenced by anthropogenic and biomass burning emissions. Geophys. Res. Lett. 2017, 44, 7472–7481. [Google Scholar] [CrossRef]
- Watson, L.; Lacressonnière, G.; Gauss, M.; Engardt, M.; Andersson, C.; Josse, B.; Marécal, V.; Nyiri, A.; Sobolowski, S.; Siour, G.; et al. Impact of emissions and +2 °C climate change upon future ozone and nitrogen dioxide over Europe. Atmos. Environ. 2016, 142, 271–285. [Google Scholar] [CrossRef]
- Sicard, P.; Anav, A.; De Marco, A.; Paoletti, E. Projected global ground-level ozone impacts on vegetation under different emission and climate scenarios. Atmos. Chem. Phys. 2017, 17, 12177–12196. [Google Scholar] [CrossRef] [Green Version]
- Ainsworth, E.A. Understanding and improving global crop response to ozone pollution. Plant. J. 2017, 90, 886–897. [Google Scholar] [CrossRef] [PubMed]
- Ahmed Bhuiyan, M.; Rashid Khan, H.U.; Zaman, K.; Hishan, S.S. Measuring the impact of global tropospheric ozone, carbon dioxide and sulfur dioxide concentrations on biodiversity loss. Environ. Res. 2018, 160, 398–411. [Google Scholar] [CrossRef] [PubMed]
- Nuvolone, D.; Petri, D.; Voller, F. The effects of ozone on human health. Environ. Sci. Pollut. Res. Int. 2018, 25, 8074–8088. [Google Scholar] [CrossRef]
- Harmens, H.; Hayes, F.; Mills, G.; Sharps, K.; Osborne, S.; Pleijel, H. Wheat yield responses to stomatal uptake of ozone: Peak vs. rising background ozone conditions. Atmos. Environ. 2018, 173, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Burkey, K.O.; Neufeld, H.S.; Souza, L.; Chappelka, A.H.; Davison, A.W. Seasonal profiles of leaf ascorbic acid content and redox state in ozone-sensitive wildflowers. Environ. Pollut. 2006, 143, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Chang-Espino, M.; González-Fernández, I.; Alonso, R.; Araus, J.L.; Bermejo-Bermejo, V. The effect of increased ozone levels on the stable carbon and nitrogen isotopic signature of wheat cultivars and landraces. Atmosphere 2021, 12, 883. [Google Scholar] [CrossRef]
- Van Dingenen, R.; Dentener, F.J.; Raes, F.; Krol, M.C.; Emberson, L.; Cofala, J. The global impact of ozone on agricultural crop yields under current and future air quality legislation. Atmos. Environ. 2009, 43, 604–618. [Google Scholar] [CrossRef]
- Pleijel, H.; Broberg, M.C.; Uddling, J.; Mills, G. Current surface ozone concentrations significantly decrease wheat growth, yield and quality. Sci. Total Environ. 2018, 613–614, 687–692. [Google Scholar] [CrossRef]
- Šramková, Z.; Gregová, E.; Šturdík, E. Chemical composition and nutritional quality of wheat grain. Acta Chim. Slovaca 2009, 2, 115–138. [Google Scholar]
- Cotrozzi, L.; Lorenzini, G.; Nali, C.; Pisuttu, C.; Pampana, S.; Pellegrini, E. Transient waterlogging events impair shoot and root physiology and reduce grain yield of durum wheat cultivars. Plants 2021, 10, 2357. [Google Scholar] [CrossRef]
- Kadkol, G.P.; Sissons, M. Durum wheat: Overview. Encycl. Food Grain 2016, 1, 117–124. [Google Scholar]
- Li, L.; Niu, Y.; Ruan, Y.; DePauw, R.; Singh, A.; Gan, Y. Agronomic advancement in tillage, crop rotation, soil health, and genetic gain in durum wheat cultivation: A 17-year Canadian story. Agronomy 2018, 8, 193. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, S.; Jha, R.K.; Cotrozzi, L.; Nali, C.; Lorenzini, G.; Ma, L. Phenol metabolism of two cultivars of durum wheat (Triticum durum Desf.) as affected by ozone and flooding stress. J. Agron. Crop Sci. 2020, 206, 338–351. [Google Scholar] [CrossRef]
- Gerosa, G.; Marzuoli, R.; Finco, A.; Monga, R.; Fusaro, I.; Faoro, F. Contrasting effects of water salinity and ozone concentration on two cultivars of durum wheat (Triticum durum Desf.) in Mediterranean conditions. Environ. Pollut. 2014, 193, 13–21. [Google Scholar] [CrossRef]
- Cotrozzi, L.; Remorini, D.; Pellegrini, E.; Guidi, L.; Nali, C.; Lorenzini, G.; Massai, R.; Landi, M. Living in a Mediterranean city in 2050: Broadleaf or evergreen ‘citizens’? Environ. Sci. Pollut. Res. Int. 2018, 25, 8161–8173. [Google Scholar] [CrossRef] [PubMed]
- Saurer, M.; Fuhrer, J.; Siegenthaler, U. Influence of ozone on the stable carbon isotope composition, δ13C, of leaves and grain of spring wheat (Triticum aestivum L.). Plant. Physiol. 1991, 97, 313–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araus, J.L.; Cabrera-Bosquet, L.; Serret, M.D.; Bort, J.; Nieto-Taladriz, M.T. Comparative perofrmance of δ13C, δ18O and δ15N for phenotyping durum wheat adaptation to a dryland environment. Funct. Plant Biol. 2013, 40, 595–608. [Google Scholar] [CrossRef]
- Spangenberg, J.E.; Schweizer, M.; Zufferey, V. Shifts in carbon and nitrogen stable isotope composition and epicuticular lipids in leaves reflect early water-stress in vineyards. Sci. Total Environ. 2020, 739, 140343. [Google Scholar] [CrossRef]
- Högberg, P. Tansley review no 95—15N natural abundance in soil-plant systems. New Phytol. 1997, 137, 179–203. [Google Scholar] [CrossRef]
- Evans, R.D. Physiological mechanisms influencing plant nitrogen isotope composition. Trends Plant Sci. 2001, 6, 121–126. [Google Scholar] [CrossRef]
- Reichenauer, T.G.; Goodman, B.A.; Kostecki, P.; Soja, G. Ozone sensitivity in Triticum durum and T. aestivum with respect to leaf injury, photosynthetic activity and free radical content. Physiol. Plant. 2010, 104, 681–686. [Google Scholar] [CrossRef]
- Rai, R.; Agrawal, M.; Agrawal, S.B. Effects of ambient O3 on wheat during reproductive development: Gas exchange, photosynthetic pigments, chlorophyll fluorescence, and carbohydrates. Photosynthetica 2011, 49, 285–294. [Google Scholar] [CrossRef]
- Feng, Z.; Jiang, L.; Calatayud, V.; Dai, L.; Paoletti, E. Intraspecific variation in sensitivity of winter wheat (Triticum aestivum L.) to ambient ozone in northern China as assessed by ethylenediurea (EDU). Environ. Sci. Pollut. Res. Int. 2018, 25, 29208–29218. [Google Scholar] [CrossRef] [PubMed]
- Latowski, D.; Grzyb, J.; Strzałka, K. The xanthophyll cycle—Molecular mechanism and physiological significance. Acta Physiol. Plant. 2004, 26, 197–212. [Google Scholar] [CrossRef]
- Pleijel, H.; Broberg, M.C.; Uddling, J. Ozone impact on wheat in Europe, Asia and North America—A comparison. Sci. Total Environ. 2019, 664, 908–914. [Google Scholar] [CrossRef]
- Fenollosa, E.; Munné-Bosch, S. Photoprotection and photo-oxidative stress markers as useful tools to unravel plant invasion success. In Advances in Plant Ecophysiology Techniques; Sánchez-Moreiras, A., Reigosa, M., Eds.; Springer: Cham, Switzerland, 2018; pp. 153–175. [Google Scholar]
- Herbinger, K.; Tausz, M.; Wonisch, A.; Soja, G.; Sorger, A.; Grill, D. Complex interactive effects of drought and ozone stress on the antioxidant defence systems of two wheat cultivars. Plant. Physiol. Biochem. 2002, 40, 691–696. [Google Scholar] [CrossRef]
- Monga, R.; Marzuoli, R.; Alonso, R.; Bermejo, V.; González-Fernández, I.; Faoro, F.; Gerosa, G. Varietal screening of ozone sensitivity in Mediterranean durum wheat (Triticum durum, Desf.). Atmos. Environ. 2015, 110, 18–26. [Google Scholar] [CrossRef]
- Hare, V.J.; Loftus, E.; Jeffrey, A.; Ramsey, C.B. Atmospheric CO2 effect on stable carbon isotope composition of terrestrial fossil archives. Nat. Commun. 2018, 9, 252. [Google Scholar] [CrossRef] [Green Version]
- Lomax, B.H.; Knight, C.A.; Lake, J.A. An experimental evaluation of the use of C3 δ13C plant tissue as a proxy for the paleoatmospheric δ13CO2 signature of air. Geochem. Geophys. Geosystems 2012, 13, Q0AI03. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Wang, X. Stable carbon isotope and long-chain alkane compositions of the major plants and sediment organic matter in the Yellow River estuarine wetlands. J. Ocean. Univ. China 2019, 18, 735–742. [Google Scholar] [CrossRef]
- Iniguez, C.; Capo-Bauca, S.; Niinemets, U.; Stoll, H.; Aguilo-Nicolau, P.; Galmes, J. Evolutionary trends in RuBisCO kinetics and their co-evolution with CO2 concentrating mechanisms. Plant. J. 2020, 101, 897–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Twohey, R.J., III; Roberts, L.M.; Studer, A.J. Leaf stable carbon isotope composition reflects transpiration efficiency in Zea mays. Plant J. 2019, 97, 475–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmann, D.; Jung, K.; Bender, J.; Gehre, M.; Schuurmann, G. Using natural isotope variations of nitrogen in plants as an early indicator of air pollution stress. J. Mass Spectrom. 1997, 32, 855–863. [Google Scholar] [CrossRef]
- Pandey, A.K.; Ghosh, A.; Agrawal, M.; Agrawal, S.B. Effect of elevated ozone and varying levels of soil nitrogen in two wheat (Triticum aestivum L.) cultivars: Growth, gas-exchange, antioxidant status, grain yield and quality. Ecotoxicol. Environ. Saf. 2018, 158, 59–68. [Google Scholar] [CrossRef]
- Alwi, N.A.; Ali, A. Dose-dependent effect of ozone fumigation on physiological characteristics, ascorbic acid content and disease development on bell pepper (Capsicum annuum L.) during storage. Food Bioprocess Technol. 2014, 8, 558–566. [Google Scholar] [CrossRef]
- Trandel, M.A.; Vigardt, A.; Walters, S.A.; Lefticariu, M.; Kinsel, M. Nitrogen isotope composition, nitrogen amount, and fruit yield of tomato plants affected by the soil-fertilizer types. ACS Omega 2018, 3, 6419–6426. [Google Scholar] [CrossRef]
- Chen, C.; Li, J.; Wang, G.; Shi, M. Accounting for the effect of temperature in clarifying the response of foliar nitrogen isotope ratios to atmospheric nitrogen deposition. Sci. Total Environ. 2017, 609, 1295–1302. [Google Scholar] [CrossRef]
- Anand, A.; Kaur, G.; Bammidi, S.; Mathur, D.; Battu, P.; Sharma, K.; Tyagi, R.; Pannu, V.; Bhanushali, D.; Limaye, N. Primer for mainstreaming Mind-Body techniques for extreme climates-insights and future directions. Medicines 2020, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Lancashire, P.D.; Bleiholder, H.; Boom, T.V.D.; Langeluddeke, P.; Stauss, R.; Weber, E.; Witzenberger, A. A uniform decimal code for growth stages of crops and weeds. Ann. Appl. Biol. 1991, 119, 561–601. [Google Scholar] [CrossRef]
- Landi, M.; Cotrozzi, L.; Pellegrini, E.; Remorini, D.; Tonelli, M.; Trivellini, A.; Nali, C.; Guidi, L.; Massai, R.; Vernieri, P.; et al. When “thirsty” means “less able to activate the signalling wave trigged by a pulse of ozone”: A case of study in two Mediterranean deciduous oak species with different drought sensitivity. Sci. Total Environ. 2019, 657, 379–390. [Google Scholar] [CrossRef]
- Davidson, J.A.; Cantrell, C.A.; Tyler, S.C.; Shetter, R.E.; Cicerone, R.J.; Calvert, J.G. Carbon kinetic isotope effect in the reaction of CH4 with HO. J. Geophys. Res. Atmos. 1987, 92, 2195–2199. [Google Scholar] [CrossRef]
- Esmeijer-Liu, A.J.; Kurschner, W.M.; Lotter, A.F.; Verhoeven, J.T.; Goslar, T. Stable carbon and nitrogen isotopes in a peat profile are influenced by early stage diagenesis and changes in atmospheric CO2 and N deposition. Water Air Soil Pollut. 2012, 223, 2007–2022. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Claudio | Mongibello | p | |||||
|---|---|---|---|---|---|---|---|
| Control | O3 | Control | O3 | Cv | O3 | Cv × O3 | |
| (df: 1) | (df: 1) | (df: 1) | |||||
| Leaf biomass (g DW plant−1) | 2.1 ± 1.3 | 1.5 ± 0.4 | 2.3 ± 1.0 | 1.5 ± 0.4 | 0.833 | <0.001 | 0.833 |
| Stem biomass (g DW plant−1) | 3.5 ± 1.2 | 2.4 ± 0.7 | 4.3 ± 1.4 | 2.5 ± 0.7 | 0.523 | <0.001 | 0.542 |
| Total aboveground biomass (g DW plant−1) | 10.11 ± 4.11 b | 7.1 ± 2.3 a | 13.3 ± 5.1 c | 8.0 ± 2.4 a | <0.001 | <0.001 | 0.015 |
| Ears plant−1 (n) | 3.7 ± 1.3 | 4.0 ± 1.3 | 4.2 ± 1.1 | 3.7 ± 1.1 | 0.888 | 0.888 | 0.577 |
| Ear weight (g DW plant−1) | 4.5 ± 1.6 b | 3.2 ± 1.2 a | 6.7 ± 2.6 c | 4.0 ± 1.3 ab | <0.001 | <0.001 | 0.014 |
| Grain weight (g DW plant−1) | 2.4 ± 1.1 a | 2.2 ± 1.0 a | 3.7 ± 0.9 b | 2.3 ± 0.6 a | 0.018 | 0.007 | 0.026 |
| Thousand grain weight (g) | 24.7 ± 3.0 a | 20.3 ± 1.4 a | 37.4 ± 3.5 b | 23.4 ± 1.4 a | <0.001 | <0.001 | 0.024 |
| Grains ear−1 (n) | 42.0 ± 9.0 c | 30.0 ± 6.1 b | 40.0 ± 7.9 c | 24.0 ± 9.0 a | 0.003 | <0.001 | 0.013 |
| Spikelets ear−1 (n) | 16.4 ± 2.3 b | 15.5 ± 1.9 b | 16.0 ± 1.8 b | 13.6 ± 2.0 a | 0.018 | <0.001 | 0.025 |
| Grains spikelet−1 (n) | 2.8 ± 0.6 | 2.0 ± 0.6 | 2.5 ± 0.6 | 1.8 ± 0.9 | 0.536 | <0.001 | 0.551 |
| df | La | A | gs | Vcmax | |
|---|---|---|---|---|---|
| Cv | 1 | 0.894 | 0.160 | 0.103 | 0.687 |
| O3 | 1 | <0.001 | 0.002 | 0.017 | <0.001 |
| Gs | 3 | <0.001 | <0.001 | <0.001 | <0.001 |
| Cv × O3 | 1 | 0.478 | 0.019 | 0.053 | 0.724 |
| Cv × Gs | 3 | 0.050 | 0.195 | 0.021 | 0.003 |
| O3 × Gs | 3 | <0.001 | 0.046 | 0.057 | 0.120 |
| Cv × O3 × Gs | 3 | 0.866 | 0.351 | 0.258 | 0.160 |
| p Values | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| La | A | gs | Vcmax | δ15N | δ13C | ||||
| CTR | 7 days | CLAU | MONG | 0.991 | 0.278 | 0.515 | 0.178 | <0.001 | 0.216 |
| 28 days | CLAU | MONG | 0.790 | 0.278 | 0.214 | 0.019 | <0.001 | 0.012 | |
| 50 days | CLAU | MONG | 0.159 | 0.122 | 0.646 | 0.466 | <0.001 | <0.001 | |
| 70 days | CLAU | MONG | 0.055 | 0.094 | 0.008 | 0.161 | <0.001 | 0.001 | |
| O3 | 7 days | CLAU | MONG | 1.000 | 0.632 | 0.160 | 0.009 | <0.001 | 0.002 |
| 28 days | CLAU | MONG | 0.833 | 0.025 | 0.097 | 0.142 | <0.001 | 0.154 | |
| 50 days | CLAU | MONG | 0.067 | 0.341 | 0.057 | 0.822 | 0.004 | <0.001 | |
| 70 days | CLAU | MONG | 0.634 | 0.177 | 0.084 | 0.165 | <0.001 | 0.907 | |
| CLAU | 7 days | CTR | O3 | 0.991 | 0.539 | 0.735 | 0.024 | <0.001 | 0.487 |
| 28 days | CTR | O3 | 0.317 | 0.249 | 0.291 | 0.001 | 0.067 | 0.431 | |
| 50 days | CTR | O3 | <0.001 | 0.632 | 0.717 | 0.378 | 0.155 | 0.226 | |
| 70 days | CTR | O3 | 0.642 | 0.891 | 0.744 | 0.416 | 0.530 | 0.116 | |
| MONG | 7 days | CTR | O3 | 0.982 | 0.223 | 0.663 | 0.335 | 0.001 | 0.147 |
| 28 days | CTR | O3 | 0.292 | <0.001 | <0.001 | 0.017 | <0.001 | 0.002 | |
| 50 days | CTR | O3 | <0.001 | 0.047 | 0.046 | 0.169 | <0.001 | <0.001 | |
| 70 days | CTR | O3 | 0.307 | 0.837 | 0.484 | 0.051 | <0.001 | <0.001 | |
| CLAU | CTR | 7 days | 28 days | 0.047 | 0.945 | 0.772 | 0.008 | <0.001 | 0.727 |
| 7 days | 50 days | <0.001 | 0.025 | 0.260 | 0.031 | <0.001 | 0.004 | ||
| 7 days | 70 days | 0.003 | <0.001 | 0.003 | <0.001 | <0.001 | <0.001 | ||
| 28 days | 50 days | <0.001 | 0.022 | 0.399 | 0.574 | 0.259 | 0.009 | ||
| 28 days | 70 days | 0.237 | <0.001 | 0.001 | <0.001 | <0.001 | <0.001 | ||
| 50 days | 70 days | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | ||
| O3 | 7 days | 28 days | 0.297 | 0.539 | 0.663 | 0.095 | <0.001 | 0.259 | |
| 7 days | 50 days | <0.001 | 0.002 | 0.073 | 0.001 | <0.001 | 0.238 | ||
| 7 days | 70 days | 0.001 | <0.001 | 0.014 | 0.004 | <0.001 | <0.001 | ||
| 28 days | 50 days | <0.001 | <0.001 | 0.029 | 0.052 | 0.483 | 0.025 | ||
| 28 days | 70 days | 0.011 | 0.002 | 0.038 | <0.001 | <0.001 | <0.001 | ||
| 50 days | 70 days | 0.006 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | ||
| MONG | CTR | 7 days | 28 days | 0.027 | 0.945 | 0.373 | 0.314 | <0.001 | 0.298 |
| 7 days | 50 days | <0.001 | 0.008 | 0.348 | 0.115 | <0.001 | 0.022 | ||
| 7 days | 70 days | <0.001 | <0.001 | 0.275 | <0.001 | 0.002 | <0.001 | ||
| 28 days | 50 days | <0.001 | 0.007 | 0.961 | 0.013 | <0.001 | 0.184 | ||
| 28 days | 70 days | 0.006 | <0.001 | 0.053 | <0.001 | <0.001 | <0.001 | ||
| 50 days | 70 days | <0.001 | <0.001 | 0.047 | <0.001 | 0.564 | <0.001 | ||
| O3 | 7 days | 28 days | 0.212 | 0.002 | 0.001 | 0.016 | <0.001 | 0.001 | |
| 7 days | 50 days | <0.001 | 0.054 | 0.128 | 0.242 | <0.001 | <0.001 | ||
| 7 days | 70 days | <0.001 | 0.004 | 0.031 | <0.001 | <0.001 | <0.001 | ||
| 28 days | 50 days | 0.001 | <0.001 | 0.051 | 0.001 | <0.001 | <0.001 | ||
| 28 days | 70 days | 0.006 | 0.732 | 0.193 | <0.001 | <0.001 | <0.001 | ||
| 50 days | 70 days | 0.596 | <0.001 | 0.492 | <0.001 | <0.001 | <0.001 | ||
| df | δ13C | δ15N | |
|---|---|---|---|
| Cv | 1 | <0.001 | <0.001 |
| O3 | 1 | 0.001 | <0.001 |
| Gs | 3 | <0.001 | <0.001 |
| Cv ×O3 | 1 | 0.090 | <0.001 |
| Cv × Gs | 3 | <0.001 | <0.001 |
| O3 × Gs | 3 | <0.001 | <0.001 |
| Cv ×O3 × Gs | 3 | 0.001 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, L.; Chen, C.; Cotrozzi, L.; Bu, C.; Luo, J.; Yao, G.; Chen, G.; Zhang, W.; Nali, C.; Lorenzini, G. The Effects of Elevated Tropospheric Ozone on Carbon Fixation and Stable Isotopic Signatures of Durum Wheat Cultivars with Different Biomass and Yield Stability. Plants 2022, 11, 3185. https://doi.org/10.3390/plants11223185
Ma L, Chen C, Cotrozzi L, Bu C, Luo J, Yao G, Chen G, Zhang W, Nali C, Lorenzini G. The Effects of Elevated Tropospheric Ozone on Carbon Fixation and Stable Isotopic Signatures of Durum Wheat Cultivars with Different Biomass and Yield Stability. Plants. 2022; 11(22):3185. https://doi.org/10.3390/plants11223185
Chicago/Turabian StyleMa, Limin, Chong Chen, Lorenzo Cotrozzi, Chengcheng Bu, Jiahong Luo, Guodong Yao, Guangyao Chen, Weiwei Zhang, Cristina Nali, and Giacomo Lorenzini. 2022. "The Effects of Elevated Tropospheric Ozone on Carbon Fixation and Stable Isotopic Signatures of Durum Wheat Cultivars with Different Biomass and Yield Stability" Plants 11, no. 22: 3185. https://doi.org/10.3390/plants11223185
APA StyleMa, L., Chen, C., Cotrozzi, L., Bu, C., Luo, J., Yao, G., Chen, G., Zhang, W., Nali, C., & Lorenzini, G. (2022). The Effects of Elevated Tropospheric Ozone on Carbon Fixation and Stable Isotopic Signatures of Durum Wheat Cultivars with Different Biomass and Yield Stability. Plants, 11(22), 3185. https://doi.org/10.3390/plants11223185