
Phytogeographic Characteristics of Montane Coniferous Forests of the Central Balkan Peninsula (SE Europe)


Abstract
:1. Introduction
2. Materials and Methods
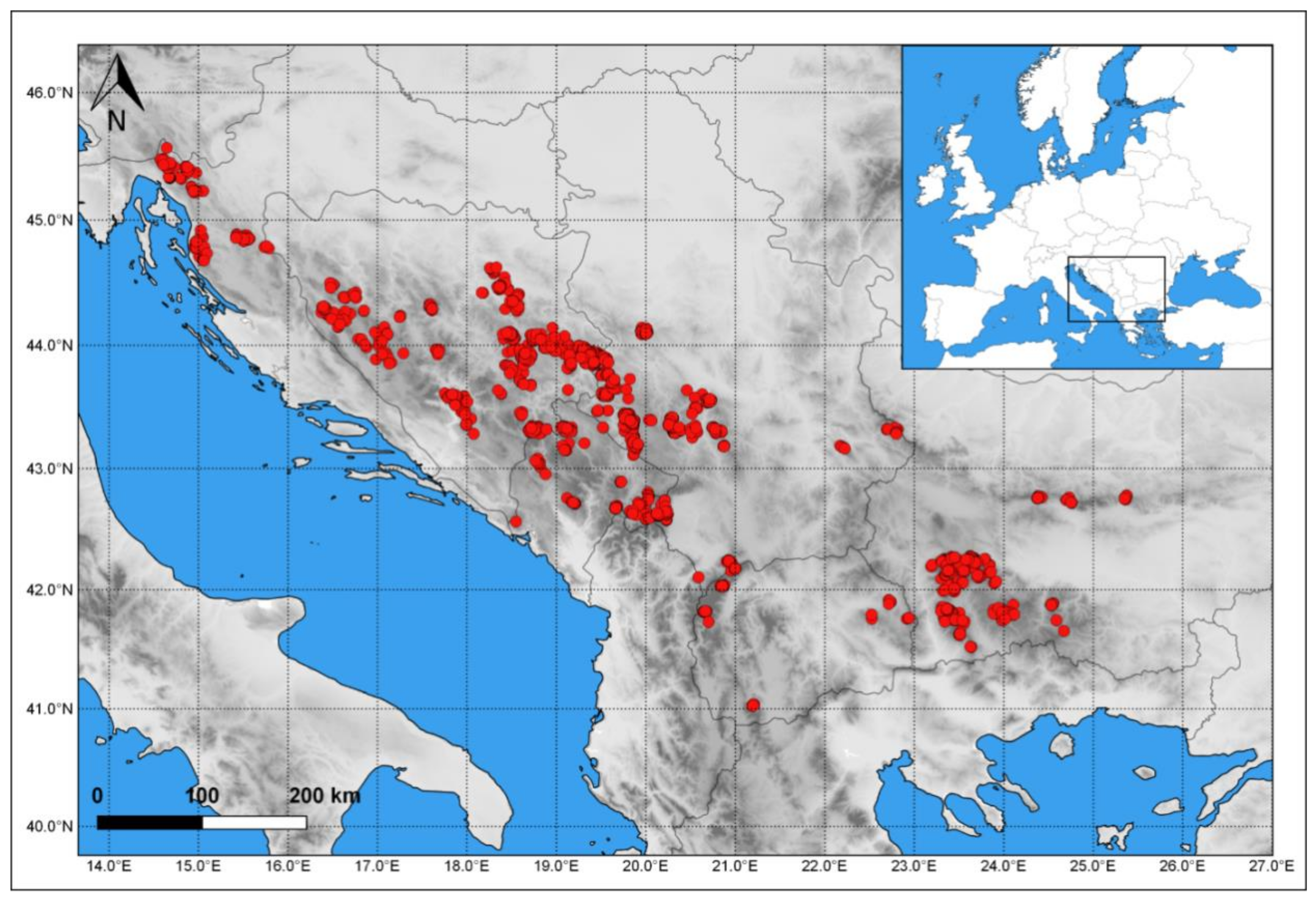
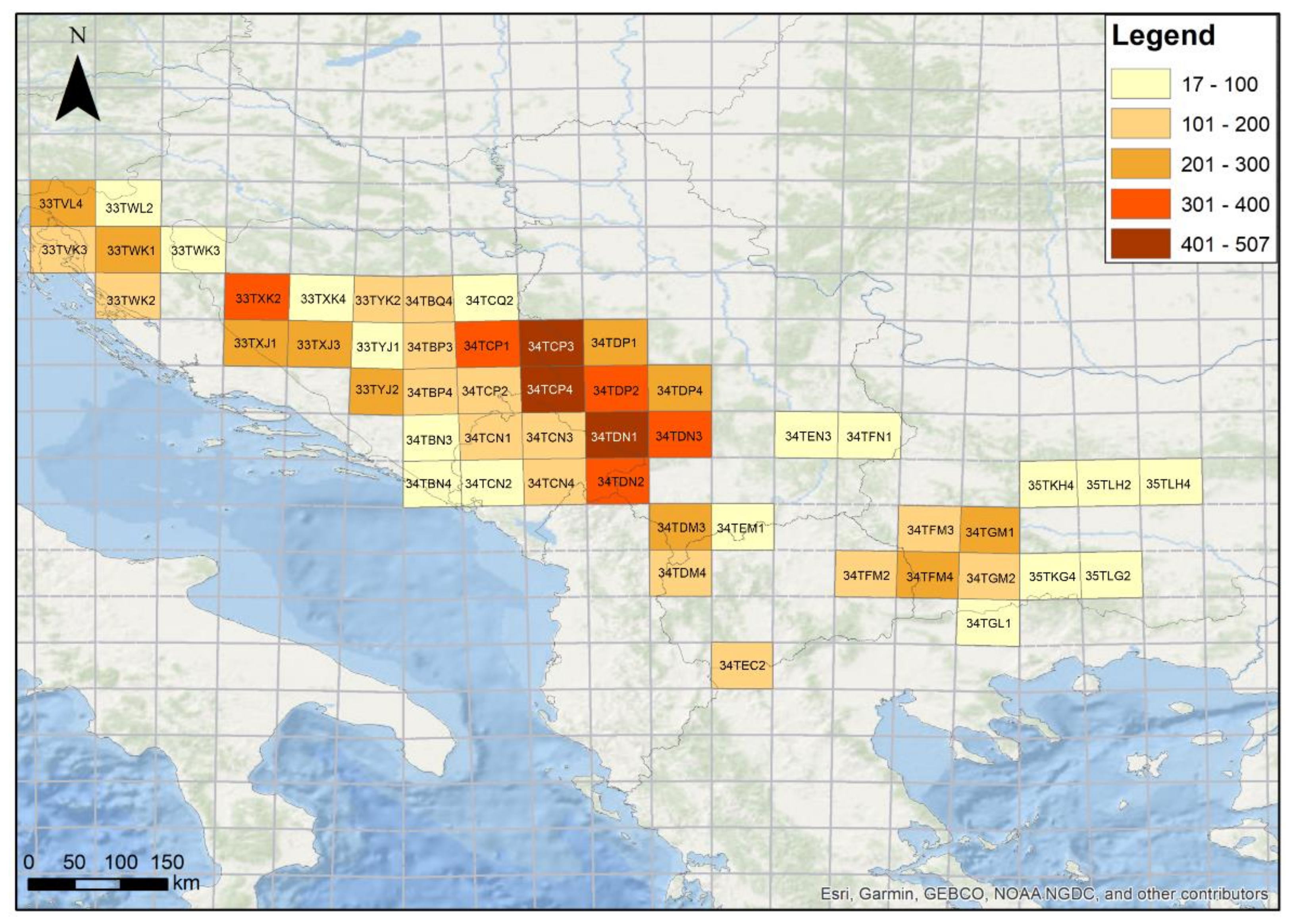
2.1. Study Area, Forest Types, and Data Gathering Principles
2.2. Statistical Analyses
3. Results and Discussion
3.1. Species Richness and Diversity
3.2. Centers of Floristic Richness
3.3. Cenotic and Spatial Diversification
3.4. Chorological Spectrum
3.5. Endemism by Vegetation Types
3.6. Life Form Spectrum
3.7. Conservation Value of Montane Coniferous Forests of the Central Balkan Peninsula
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity Hotspots for Conservation Priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Barthlott, W.; Mutke, J.; Rafiqpoor, M.D.; Kier, G.; Kreft, H. Global Centers of Vascular Plant Diversity. Nova Acta Leopold. Abh. Kais. Leopold.-Carol. Dtsch. Akad. Nat. 2005, 92, 61–83. [Google Scholar]
- Mutke, J.; Sommer, J.H.; Kreft, H.; Kier, G.; Barthlott, W. Vascular Plant Diversity in a Changing World: Global Centres and Biome-Specific Patterns. In Biodiversity Hotspots: Distribution and Protection of Conservation Priority Areas; Zachos, F.E., Habel, J.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 83–96. ISBN 978-3-642-20992-5. [Google Scholar]
- Večeřa, M.; Divíšek, J.; Lenoir, J.; Jiménez-Alfaro, B.; Biurrun, I.; Knollová, I.; Agrillo, E.; Campos, J.A.; Čarni, A.; Crespo Jiménez, G.; et al. Alpha Diversity of Vascular Plants in European Forests. J. Biogeogr. 2019, 46, 1919–1935. [Google Scholar] [CrossRef]
- Horvat, I. Šumske zajednice Jugoslavije. In Šumarska Enciklopedija; Potočić, Z., Ed.; Jugoslavenski leksikografski zavod: Zagreb, Croatia, 1963; Volume 2, pp. 560–590. [Google Scholar]
- Stevanović, V.; Jovanović, S.; Lakušić, D.; Niketić, M. Diverzitet vaskularne flore Jugoslavije sa Pregledom Vrsta od Međunarodnog Značaja. In Biodiverzitet Jugoslavije sa pregledom vrsta od međunarodnog značaja; Stevanović, V., Vasić, V., Eds.; Ecolibri & Biološki Fakultet: Beograd, Serbia, 1995; pp. 183–217. ISBN 86-7078-004-6. [Google Scholar]
- Stevanović, V. Analysis of the Central European and Mediterranean Orophytic Element on the Mountains of the W. and Central Balkan Peninsula, with Special Reference to Endemics. Bocconea 1996, 5, 77–97. [Google Scholar]
- Hewitt, G.M. Some Genetic Consequences of Ice Ages, and Their Role in Divergence and Speciation. Biol. J. Linn. Soc. 1996, 58, 247–276. [Google Scholar] [CrossRef]
- Hewitt, G.M. Genetic Consequences of Climatic Oscillations in the Quaternary. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2004, 359, 183–195, discussion 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt, T. Molecular Biogeography of Europe: Pleistocene Cycles and Postglacial Trends. Front. Zool. 2007, 4, 11. [Google Scholar] [CrossRef] [Green Version]
- Horvat, I.; Glavač, V.; Ellenberg, H. Vegetation Südosteuropas; Geobotanica selecta; Gustav Fischer Verlag: Jena, Germany, 1974; ISBN 3-437-30168-3. [Google Scholar]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; García, R.G.; et al. Vegetation of Europe: Hierarchical Floristic Classification System of Vascular Plant, Bryophyte, Lichen, and Algal Communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Bohn, U. Karte der Natürlichen Vegetation Europas Maßstab 1:2.500.000; Interaktive CD-ROM, Erläuterungstext, Legende, Karten = Map of the Natural Vegetation of Europe; Federal Agency for Nature Conservation: Bonn, Germany, 2004.
- Köhl, M.; Linser, S. Status and Trends in European Forests Characterised by the Updated Pan-European Indicators for Sustainable Forest Management. In Forest Europe, 2020: State of Europe’s Forests 2020; Ministerial Conference on the Protection of Forests in Europe-Forest Europe; Liaison Unit Bratislava: Bratislava, Slovakia, 2020; pp. 28–214. [Google Scholar]
- Janković, M.M. Fitoekologija sa Osnovama Fitocenologije i Pregledom Tipova Vegetacije na Zemlji, 6th ed.; Naučna knjiga: Beograd, Serbia, 1990; ISBN 86-23-23039-6. [Google Scholar]
- Em, H. Na južnoj granici areala smrče. Prilozi Makedonske Akademije na Naukite i Umetnostite 1986, V, 10–27. [Google Scholar]
- Council Directive 92/43/EEC of 21 May 1992 on the Conservation of Natural Habitats and of Wild Fauna and Flora. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A31992L0043 (accessed on 24 October 2022).
- Stevanović, V.; Jovanović, S.; Lakušić, D. Diverzitet vegetacije Jugoslavije. In Biodiverzitet Jugoslavije sa Pregledom Vrsta od Međunarodnog Značaja; Stevanović, V., Vasić, V., Eds.; Ecolibri & Biološki Fakultet: Beograd; pp. 219–241. 1995; ISBN 86-7078-004-6. [Google Scholar]
- Lakušić, D. Odnos specijskog i ekosistemskog diverziteta. In Biodiverzitet na Početku Novog Milenijuma, Zbornik Radova sa Naučnog Skupa Održanog 24. Novembra 2005; Anđelković, M., Ed.; Naučni skupovi knj. CXI, Odeljenje hemijskih i bioloških nauka, knj. 2; SANU: Beograd, Serbia, 2005; pp. 75–104. [Google Scholar]
- Brković, D.L.; Tomović, G.M.; Niketić, M.S.; Lakušić, D.V. Diversity Analysis of Serpentine and Non-Serpentine Flora–Or, Is Serpentinite Inhabited by a Smaller Number of Species Compared to Different Rock Types? Biologia 2015, 70, 61–74. [Google Scholar] [CrossRef]
- Bulić, Z.; Lakušić, D.; Stevanović, V. Comparative Analysis of the Vascular Floras of the Morača and Cijevna Canyons (Montenegro). Arch. Biol. Sci. 2008, 60, 485–492. [Google Scholar] [CrossRef]
- Georghiou, K.; Delipetrou, P. Patterns and Traits of the Endemic Plants of Greece. Bot. J. Linn. Soc. 2010, 162, 130–153. [Google Scholar] [CrossRef] [Green Version]
- Djordjević, V.; Tsiftsis, S.; LakuŠić, D.; Stevanović, V. Niche Analysis of Orchids of Serpentine and Non-Serpentine Areas: Implications for Conservation. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2014, 150, 710–719. [Google Scholar] [CrossRef]
- Djordjević, V.; Tsiftsis, S.; Lakušić, D.; Jovanović, S.; Stevanović, V. Factors Affecting the Distribution and Abundance of Orchids in Grasslands and Herbaceous Wetlands. Syst. Biodivers. 2016, 14, 355–370. [Google Scholar] [CrossRef]
- Djordjević, V.; Tsiftsis, S.; Lakušić, D.; Jovanović, S.; Jakovljević, K.; Stevanović, V. Patterns of Distribution, Abundance and Composition of Forest Terrestrial Orchids. Biodivers. Conserv. 2020, 29, 4111–4134. [Google Scholar] [CrossRef]
- Jakovljević, K.; Lakušić, D.; Vukojičić, S.; Tomović, G.; Šinžar-Sekulić, J.; Stevanović, V. Richness and Diversity of Pontic Flora on Serpentine of Serbia. Cent. Eur. J. Biol. 2011, 6, 260–274. [Google Scholar] [CrossRef]
- Lakušić, D. Visokoplaninska Flora Kopaonika-Ekološko Fitogeografska Studija-Highmountain Flora of mt. Kopaonik-Ecological-Phytogeographical Study. Magisterium, Biološki Fakultet; Univerzitet u Beogradu: Beograd, Serbia, 1993. [Google Scholar]
- Lakušić, D. Phytogeographical Characteristics of the High-Mountain Flora of Mt Kopaonik. Bocconea 1997, 5, 445–449. [Google Scholar]
- Lakušić, D.; Niketić, M.; Stevanović, V. Floristička raznovrsnost rezervata “Kanjon Lazareve reke i Malinik”. Ekologija 1996, 31, 49–59. [Google Scholar]
- Lubarda, B.; Stupar, V.; Milanović, Đ.; Stevanović, V. Chorological Characterization and Distribution of the Balkan Endemic Vascular Flora in Bosnia and Herzegovina. Bot. Serbica 2014, 38, 167–184. [Google Scholar]
- Matevski, V.; Petkovski, S.; Andonov, S.; Melovski, L.; Krstić, S. Country Study for Biodiversity of the Republic of Macedonia: (First National Report); Ministry of Environment and Physical Planning: Skopje, North Macedonia, 2003; ISBN 978-9989-110-15-3. [Google Scholar]
- Petrova, A.; Vladimirov, V. Balkan Endemics in the Bulgarian Flora. Phytol. Balcan. 2010, 16, 293–311. [Google Scholar]
- Ranđelović, V. Flora i vegetacija Vlasinske visoravni. Ph.D. Thesis, Biološki fakultet, Univerzitet u Beogradu, Beograd, Serbia, 2002. [Google Scholar]
- Stevanović, V.; Jovanović, S.; Lakušić, D.; Niketić, M. Characteristics of the Flora of Serbia and Its Phytogeographical Division. In The Red Data Book of the Flora of SERBIA; Extinct and Critically Endangered Taxa; Stevanović, V., Ed.; Ministry of Environment of the Republic of Serbia, Faculty of Biology, University of Belgrade, Instution for Protection of Nature of the Republic of Serbia: Belgrade, Serbia, 1999; Volume 1, pp. 393–399. ISBN 86-7078-012-7. [Google Scholar]
- Stevanović, V.; Kit, T.; Petrova, A. Diversity and Centres of Endemism in the Balkan Flora. In Proceedings of the Plenary Lecture in Book of Abstracts Third International Balkan Botanical Congress “Plant Resources in the Creation of New Values”, Sarajevo, Bosnia and Herzegovina, 18–24 May 2003; Redžić, S., Djug, S., Eds.; Faculty of Science, University of Sarajevo: Sarajevo, Bosnia and Herzegovina, 2003; pp. 13–14. [Google Scholar]
- Stevanović, V.; Tan, K.; Petrova, A. Mapping the Endemic Flora of the Balkans-A Progress Report. Bocconea 2007, 21, 131–137. [Google Scholar]
- Stevanović, V.; Vukojičić, S.; Šinžar-Sekulić, J.; Lazarević, M.; Tomović, G.; Tan, K. Distribution and Diversity of Arctic-Alpine Species in the Balkans. Plant Syst. Evol. 2009, 283, 219–235. [Google Scholar] [CrossRef]
- Tan, K.; Stevanović, V.; Strid, A. Distribution and Centres of Diversity for Endemic Geophytic Monocots in the Balkans. Bocconea 2007, 21, 139–146. [Google Scholar]
- Tomović, G. Fitogeografska Pripadnost, Fistribucija i Centri Diverziteta Balkanske Endemične Flore u Srbiji. Ph.D. Thesis, Biološki fakultet, Univerzitet u Beogradu: Beograd, Serbia, 2007. [Google Scholar]
- Tomović, G.; Niketić, M.; Lakušić, D.; Ranđelović, V.; Stevanović, V. Balkan Endemic Plants in Central Serbia and Kosovo Regions: Distribution Patterns, Ecological Characteristics, and Centres of Diversity. Bot. J. Linn. Soc. 2014, 176, 173–202. [Google Scholar] [CrossRef] [Green Version]
- Trigas, P.; Iatrou, G.; Karetsos, G. Species Diversity, Endemism and Conservation of the Family Caryophyllaceae in Greece. Biodivers. Conserv. 2007, 16, 357–376. [Google Scholar] [CrossRef]
- Velchev, V. Floral and Plant Biodiversity on Calcareous Terrains in Bulgaria. Phytol. Balc. 1998, 4, 81–92. [Google Scholar]
- Vukojičić, S.; Jakovljević, K.; Matevski, V.; Randjelović, V.; Niketić, M.; Lakušić, D. Distribution, Diversity and Conservation of Boreo-Montane Plant Species in the Central Part of the Balkan Peninsula and the Southern Part of the Pannonian Plain. Folia Geobot. 2014, 49, 487–505. [Google Scholar] [CrossRef]
- Vuksanović, S.; Tomović, G.; Niketić, M.; Stevanović, V. Balkan Endemic Vascular Plants of Montenegro—Critical Inventory with Chorological and Life-Form Analyses. Will 2016, 46, 387–397. [Google Scholar] [CrossRef] [Green Version]
- Zlatković, B. Flora i Fitogeografska Pripadnost Doline Reke Pčinje u Jugoistočnoj Srbiji. Ph.D. Thesis, Biološki fakultet, Univerzitet u Beogradu, Beograd, Serbia, 2011. [Google Scholar]
- Zlatković, B.; Nikolić, L.; Ranđelović, V.; Ranđelović, N.; Stevanović, V. Comparative Analyses of the Vascular Flora of the Pčinja River Gorges in Serbia and Macedonia. Arch. Biol. Sci. 2011, 63, 1157–1166. [Google Scholar] [CrossRef]
- Kryštufek, B.; Reed, J.M. Pattern and Process in Balkan Biodiversity—An Overview. In Balkan Biodiversity: Pattern and Process in the European Hotspot; Griffiths, H.I., Kryštufek, B., Reed, J.M., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 1–8. ISBN 978-1-4020-2854-0. [Google Scholar]
- Reed, J.M.; Kryštufek, B.; Eastwood, W.J. The Physical Geography of The Balkans and Nomenclature of Place Names. In Balkan Biodiversity; Springer: Dordrecht, The Netherlands, 2004; pp. 9–22. [Google Scholar]
- Jax, K. Ecological Units: Definitions and Application. Q. Rev. Biol. 2015, 81, 237–258. [Google Scholar] [CrossRef]
- Vassilev, K.; Pedashenko, H.; Alexandrova, A.; Tashev, A.; Ganeva, A.; Gavrilova, A.; Gradevska, A.; Assenov, A.; Vitkova, A.; Grigorov, B.; et al. Balkan Vegetation Database: Historical Background, Current Status and Future Perspectives. Phytocoenologia 2016, 46, 89–95. [Google Scholar] [CrossRef]
- Vassilev, K.; Pedashenko, H.; Alexandrova, A.; Tashev, A.; Ganeva, A.; Gavrilova, A.; Macanović, A.; Assenov, A.; Vitkova, A.; Genova, B.; et al. Balkan Vegetation Database (BVD)–Updated Information and Current Status. Veg. Classif. Surv. 2020, 1, 151–153. [Google Scholar] [CrossRef]
- Lampinen, R. Universal Transverse Mercator (UTM) and Military Grid Reference System (MGRS). Available online: https://www.luomus.fi/en/utm-mgrs-atlas-florae-europaeae (accessed on 10 September 2022).
- Meusel, H.; Jäger, E.; Weinert, E. Vergleichende Chorologie der Zentraleuropäischen Flora; Text und Kartenband; Gustav Fischer Verlag: Jena, Germany, 1965; Volume 1. [Google Scholar]
- Meusel, H.; Jäger, E.; Rauschert, S.; Weinert, E. Vergleichende Chorologie der Zentraleuropäischen Flora; Text und Kartenband; Gustav Fischer Verlag: Jena, Germany, 1978; Volume 2. [Google Scholar]
- Meusel, H.; Jäger, E.J. Vergleichende Chorologie der Zentraleuropäischen Flora; Text und Kartenband; Gustav Fischer Verlag: Jena, Germany; Stuttgart, Germany; New York, NY, USA, 1992; Volume 3, ISBN 3-334-00411-2. [Google Scholar]
- Stevanović, V. Floristička podela teritorije Srbije sa pregledom viših horiona i odgovarajućih flornih elemenata. In Flora Srbije; Sarić, M.R., Ed.; SANU, Odeljenje prirodno-matematičkih nauka: Beograd, Serbia, 1992; Volume 1, pp. 47–56. [Google Scholar]
- Raunkiaer, C. The Life Forms of Plants and Statistical Plant Geography; Clarendon Press: Oxford, UK, 1934. [Google Scholar]
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; John Wiley & Sons: New York, NY, USA, 1974; ISBN 0-471-62290-7. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- ter Braak, C.J.F.; Šmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination, Version 5.0; Microcomputer Power: Ithaca, NY, USA, 2012. [Google Scholar]
- Španiel, S.; Rešetnik, I. Plant Phylogeography of the Balkan Peninsula: Spatiotemporal Patterns and Processes. Plant Syst. Evol. 2022, 308, 1–31. [Google Scholar] [CrossRef]
- Mutke, J.; Kreft, H.; Kier, G.; Barthlott, W. European Plant Diversity in the Global Context. In Atlas of Biodiversity Risk; Pensoft Publishers: Sofia, Bulgaria, 2010; pp. 1–5. ISBN 978-954-642-446-4. [Google Scholar]
- Esseen, P.-A.; Ehnström, B.; Ericson, L.; Sjöberg, K. Boreal Forests—The Focal Habitats of Fennoscandia. In Ecological Principles of Nature Conservation: Application in Temperate and Boreal Environments; Conservation Ecology Series: Principles, Practices and Management; Hansson, L., Ed.; Springer US: Boston, MA, USA, 1992; pp. 252–325. ISBN 978-1-4615-3524-9. [Google Scholar]
- Otýpková, Z.; Chytrý, M.; Tichý, L.; Pechanec, V.; Jongepier, J.; Hájek, O. Floristic Diversity Patterns in the White Carpathians Biosphere Reserve, Czech Republic. Biologia 2011, 66, 266–274. [Google Scholar] [CrossRef]
- Wilson, J.B.; Peet, R.K.; Dengler, J.; Pärtel, M. Plant Species Richness: The World Records. J. Veg. Sci. 2012, 23, 796–802. [Google Scholar] [CrossRef]
- Ewald, J. The Calcareous Riddle: Why Are There so Many Calciphilous Species in the Central European Flora? Folia Geobot. 2003, 38, 357–366. [Google Scholar] [CrossRef]
- Ujházyová, M.; Ujházy, K.; Chytrý, M.; Willner, W.; Čiliak, M.; Máliš, F.; Slezák, M. Diversity of Beech Forest Vegetation in the Eastern Alps, Bohemian Massif and the Western Carpathians. Preslia 2016, 88, 435–457. [Google Scholar]
- Ewald, J. Plant Species Richness in Mountain Forests of the Bavarian Alps. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2008, 142, 594–603. [Google Scholar] [CrossRef]
- Pärtel, M. Local Plant Diversity Patterns and Evolutionary History at the Regional Scale. Ecology 2002, 83, 2361–2366. [Google Scholar] [CrossRef]
- Zelený, D.; Li, C.-F.; Chytrý, M. Pattern of Local Plant Species Richness along a Gradient of Landscape Topographical Heterogeneity: Result of Spatial Mass Effect or Environmental Shift? Ecography 2010, 33, 578–589. [Google Scholar] [CrossRef]
- Sandel, B.; Arge, L.; Dalsgaard, B.; Davies, R.G.; Gaston, K.J.; Sutherland, W.J.; Svenning, J.-C. The Influence of Late Quaternary Climate-Change Velocity on Species Endemism. Science 2011, 334, 660–664. [Google Scholar] [CrossRef]
- Peñas, J.; Pérez-García, F.J.; Mota, J.F. Patterns of Endemic Plants and Biogeography of the Baetic High Mountains (South Spain). Acta Bot. Gall. 2005, 152, 347–360. [Google Scholar] [CrossRef] [Green Version]
- WWF; IUCN. Centres of Plant Diversity. A Guide and Strategy for Their Conservation; Europe, Africa, South West Asia and the Middle East; Davis, S.D., Heywood, V.H., Hamilton, A.C., Eds.; IUCN Publications Unit: Cambridge, UK, 1994; Volume 1, ISBN 978-2-8317-0197-4. [Google Scholar]
- Favarger, C. Endemism in the Montane Floras of Europe. In Taxonomy, Phytogeography and Evolution; Valentine, D.H., Ed.; Academic Press: London, UK, 1972; pp. 191–204. ISBN 0-12-710250-7. [Google Scholar]
- Tzedakis, P.C.; Lawson, I.T.; Frogley, M.R.; Hewitt, G.M.; Preece, R.C. Buffered Tree Population Changes in a Quaternary Refugium: Evolutionary Implications. Science 2002, 297, 2044–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Essl, F.; Staudinger, M.; Stöhr, O.; Schratt-Ehrendorfer, L.; Rabitsch, W.; Niklfeld, H. Distribution Patterns, Range Size and Niche Breadth of Austrian Endemic Plants. Biol. Conserv. 2009, 142, 2547–2558. [Google Scholar] [CrossRef]
- Dominguez Lozano, F.; Galicia Herbada, D.; Moreno Rivero, L.; Moreno Saiz, J.; Sainz Ollero, H. Areas of High Floristic Endemism in Iberia and the Balearic Islands: An Approach to Biodiversity Conservation Using Narrow Endemics. Belg. J. Entomol. 2000, 2, 171–185. [Google Scholar]
- Bognar, A.; Faivre, S.; Pavelić, J. Glaciation traces on the North Velebit. Hrvat. Geogr. Glas. 1991, 53, 27–38. [Google Scholar]
- Milivojević, M.; Menković, L.; Ćalić, J. Pleistocene Glacial Relief of the Central Part of Mt. Prokletije (Albanian Alps). Quat. Int. 2008, 190, 112–122. [Google Scholar] [CrossRef]
- Stefanović, V. Fitocenologija sa Pregledom šumskih Fitocenoza Jugoslavije, 2nd ed.; Svjetlost: Sarajevo, Bosnia and Herzegovina, 1986. [Google Scholar]
- Alegro, A. Vegetacija Hrvatske; Interna skripta, Botanički zavod PMF-a: Zagreb, Croatia, 2000. [Google Scholar]
- Prirodna Potencijalna Vegetacija Jugoslavije (Komentar Karte M 1: 1,000,000); Jovanović, B.; Jovanović, R.; Zupančič, M. (Eds.) Naučno veće vegetacijske karte Jugoslavije: Ljubljana, Slovenia, 1986. [Google Scholar]
- Čolić, D. Antropogena degradacija jedne mešovite reliktne zajednice sa Pančićevom omorikom (Picea Omorika Pančić). Zb. Rad. Biol. Inst. Srb. 1964, 7, 1–32. [Google Scholar]
- Janković, M.M. Razmatranja o uzajamnim odnosima molike (Pinus Peuce) i munike (Pinus Heldreichii), kao i o njihovim ekološkim osobinama, posebno u odnosu na geološku podlogu. Glas. Bot. Zavoda i Bašte Univ. u Beogr. 1960, 1, 141–180. [Google Scholar]
- Košanin, N. Četinari Južne Srbije. Glas. Skopskog Naučnog Društva 1925, 1, 247–261. [Google Scholar]
- Fukarek, P. Prilog poznavanju dendrogeografskih i fitocenoloških odnosa planina sjeverozapadne Crne Gore. Rad. Od. Privred.-Teh. Nauka Naučno Društvo SR Bosne Hercegovine 1963, 22, 113–166. [Google Scholar]
- Fukarek, P. Zajednice endemne munike na planini Prenju u Hercegovini. Acta Bot. Croat. 1966, 25, 61–83. [Google Scholar]
- Janković, M.M. Prilog poznavanju munikovih šuma (Pinetum Heldreichii) na Metohijskim Prokletijama. Arh. Biol. Nauka 1958, 10, 51–69. [Google Scholar]
- Janković, M.M. Peucedano-Pinetum heldreichii M.Jank., nova asocijacija subendemičnog balkanskog bora Pinus heldreichii na Orjenu. Glas. Bot. Zavoda Bašte Univ. Beogr. 1962, 2, 203–206. [Google Scholar]
- Janković, M.M. Vegetacija SR Srbije; Istorija i opšte karakteristike. In Vegetacija SR Srbije; SANU: Beograd, Serbia, 1984; Volume 1, Opšti deo; pp. 1–189. [Google Scholar]
- Zupančič, M. Smrekovi gozdovi Evrope in Balkanskega polotoka, I. Biol. Vestn. 1980, 28, 137–158. [Google Scholar]
- Zupančič, M. Smrekovi gozdovi Evrope in Balkanskega polotoka, II. Biol. Vestn. 1982, 30, 171–188. [Google Scholar]
- Zupančič, M. Illyrische und Balkanische Arten in den Subalpinen Fichtengesellschaften der zentralen Balkanhalbinsel. Sauteria-Schr. Syst. Bot. Flor. Geobot. 1988, 4, 33–42. [Google Scholar]
- Zupančič, M. Smrekovi gozdovi Evrope in Balkanskega polotoka, III. Biol. Vestn. 1990, 38, 5–22. [Google Scholar]
- Diklić, N. Životne forme biljnih vrsta i biološki spektar Flore SR Srbije. In Vegetacija SR Srbije; SANU: Beograd, Serbia, 1984; Volume 1, Opšti deo; pp. 291–316. [Google Scholar]
- Tatić, B.; Tomić, Z. Šume crnog i belog bora. In Vegetacija Srbije; Škorić, D.M., Ed.; SANU: Beograd, Serbia, 2006; Volume 2, Šumske zajednice (2); pp. 127–154. ISBN 86-7025-428-X. [Google Scholar]
- Dinić, A.; Tatić, B. Šume Pančićeve omorike. In Vegetacija Srbije; Škorić, D.M., Ed.; SANU: Beograd, Serbia, 2006; Volume 2, Šumske zajednice (2); pp. 213–244. ISBN 86-7025-428-X. [Google Scholar]
- Turrill, W.B. The Plant-Life of the Balkan Peninsula, a Phytogeographical Study; Oxford Memoirs on Plant Geography; Clarendon Press: Oxford, UK, 1929. [Google Scholar]
- European Commission Directorate-General for Environment Forests. Available online: https://ec.europa.eu/environment/forests/index_en.htm (accessed on 10 October 2022).
- Magri, D. Patterns of Post-Glacial Spread and the Extent of Glacial Refugia of European Beech (Fagus Sylvatica). J. Biogeogr. 2008, 35, 450–463. [Google Scholar] [CrossRef]
- Tzedakis, P.C.; Emerson, B.C.; Hewitt, G.M. Cryptic or Mystic? Glacial Tree Refugia in Northern Europe. Trends Ecol. Evol. 2013, 28, 696–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez-Alfaro, B.; Chytrý, M.; Mucina, L.; Grace, J.B.; Rejmánek, M. Disentangling Vegetation Diversity from Climate–Energy and Habitat Heterogeneity for Explaining Animal Geographic Patterns. Ecol. Evol. 2016, 6, 1515–1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, A.; Lunt, D.H. Refugia within Refugia: Patterns of Phylogeographic Concordance in the Iberian Peninsula. In Phylogeography of Southern European Refugia; Weiss, S., Ferrand, N., Eds.; Springer: Dordrecht, Germany, 2007; pp. 155–188. ISBN 978-94-007-0752-8. [Google Scholar] [CrossRef]
- Surina, B.; Schönswetter, P.; Schneeweiss, G.M. Quaternary Range Dynamics of Ecologically Divergent Species (Edraianthus Serpyllifolius and E. Tenuifolius, Campanulaceae) within the Balkan Refugium. J. Biogeogr. 2011, 38, 1381–1393. [Google Scholar] [CrossRef]
- Council of Europe Convention on the Conservation of European Wildlife and Natural Habitats Resolution No. 6 (1998) Listing the Species Requiring Specific Habitat Conservation Measures (Adopted by the Standing Committee on 4 December 1998). Available online: https://rm.coe.int/1680746afc (accessed on 8 November 2022).
- CITES Convention on International Trade in Endangered Species of Wild Fauna and Flora, Appendices I, II and III. Available online: https://cites.org/sites/default/files/eng/app/2022/E-Appendices-2022-06-22.pdf (accessed on 9 November 2022).
- European Commission Directorate-General for Environment EU Biodiversity Strategy for 2030. Available online: https://environment.ec.europa.eu/strategy/biodiversity-strategy-2030_en (accessed on 10 October 2022).
- European Commission Directorate-General for Environment New EU Forest Strategy for 2030. Available online: https://environment.ec.europa.eu/strategy/forest-strategy_en (accessed on 10 October 2022).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level | Forest Types | Codes | Estimated Area of Occupancy (in ha) |
|---|---|---|---|
| I | Dark spruce forest types (Vaccinio-Piceetea) | Vaccinio-Piceetea | 1,059,824 |
| II | Norway spruce forests (Vaccinio-Piceion) | Vaccinio-Piceion | 635,389 |
| III | Norway spruce forests (Vaccinio-Piceion) in Dinarides | Vacc_Pic_Din | |
| III | Norway spruce forests (Vaccinio-Piceion) in Scardo-Pindic mountains | Vacc_Pic_ScPind | |
| III | Norway spruce forests (Vaccinio-Piceion) in Rhodope mountains | Vacc_Pic_Rhod | |
| III | Norway spruce forests (Vaccinio-Piceion) in Balkan mountains | Vacc_Pic_Balk | |
| II | Scots pine forests (Pinion sylvestris) | Pinion silvestris | 318,880 |
| III | Scots pine forests (Pinion sylvestris) in Dinarides | Pin_syl_Din | |
| III | Scots pine forests (Pinion sylvestris) in Rhodope mountains | Pin_syl_Rhod | |
| III | Scots pine forests (Pinion sylvestris) in Balkan mountains | Pin_syl_Balk | |
| II | Serbian spruce forest (Piceion omorikae) | Piceion omorikae | 6697 |
| III | Serbian spruce forest (Piceion omorikae) in Dinarides | Pic_omor_Din | |
| II | Macedonian pine forests (Pinion peucis) | Pinion peucis | 98,858 |
| III | Macedonian pine forests (Pinion peucis) in Dinarides | Pin_peuc_Din | |
| III | Macedonian pine forests (Pinion peucis) in Scardo-Pindic mountains | Pin_peuc_ScPind | |
| III | Macedonian pine forests (Pinion peucis) in Rhodope mountains | Pin_peuc_Rhod | |
| III | Macedonian pine forests (Pinion peucis) in Balkan mountains | Pin_peuc_Balk | |
| I | Light pine forest types (Erico-Pinetea) | Erico-Pinetea | 149,730 |
| II | Bosnian pine forests (Pinion heldreichii) | Pinion heldreichii | 133,334 |
| III | Bosnian pine forests (Pinion heldreichii) in Dinarides | Pin_heldr_Din | |
| III | Bosnian pine forests (Pinion heldreichii) in Scardo-Pindic mountains | Pin_heldr_ScPind | |
| III | Bosnian pine forests (Pinion heldreichii) in Rhodope mountains | Pin_heldr_Rhod | |
| II | Ultramafic black pine forests (Orno-Ericion) | Orno-Ericion | 16,395 |
| III | Ultramafic black pine forests (Orno-Ericion) in Dinarides | Orn_Eric_Din |
| Level I | No. Relevés | Relevé Area (m2) | S | Logs/Loga | S End | % End |
|---|---|---|---|---|---|---|
| Vaccinio-Piceeetea | 1499 | 1,601,977 | 1212 | 0.497 | 188 | 16 |
| Erico-Pinetea | 431 | 545,314 | 861 | 0.512 | 162 | 19 |
| Level II | No. Relevés | Relevé area (m2) | S | logS/logA | S End | % End |
| Vaccinio-Piceion | 944 | 768,955 | 905 | 0.502 | 115 | 13% |
| Piceion omorikae | 59 | 24,854 | 246 | 0.544 | 21 | 9% |
| Pinion sylvestris | 348 | 463,215 | 807 | 0.513 | 108 | 13% |
| Pinion peucis | 148 | 344,954 | 282 | 0.442 | 50 | 18% |
| Orno-Ericion | 219 | 239,633 | 531 | 0.507 | 69 | 13% |
| Pinion heldreichii | 212 | 305,681 | 579 | 0.504 | 114 | 20% |
| Level III | No. Relevés | Relevé area (m2) | S | logS/logA | S End | % End |
| Vacc Pic Din | 758 | 617,444 | 841 | 0.505 | 89 | 11 |
| Vacc Pic ScPind | 26 | 21,179 | 121 | 0.481 | 14 | 12 |
| Vacc Pic Rhod | 124 | 101,007 | 194 | 0.457 | 19 | 10 |
| Vacc Pic Balk | 36 | 29,325 | 177 | 0.503 | 13 | 7 |
| Pic omor Din | 59 | 24,854 | 246 | 0.544 | 21 | 9 |
| Pin syl Din | 261 | 347,411 | 686 | 0.512 | 76 | 11 |
| Pin syl Rhod | 82 | 109,148 | 320 | 0.497 | 42 | 13 |
| Pin syl Balk | 5 | 6655 | 62 | 0.469 | 2 | 3 |
| Pin peuc Din | 21 | 48,946 | 114 | 0.439 | 13 | 11 |
| Pin peuc ScPind | 11 | 25,638 | 146 | 0.491 | 28 | 19 |
| Pin peuc Rhod | 95 | 221,423 | 129 | 0.395 | 15 | 12 |
| Pin peuc Balk | 21 | 48,946 | 79 | 0.405 | 11 | 14 |
| Orn Eric Din | 219 | 239,633 | 531 | 0.507 | 69 | 13 |
| Pin heldr Din | 114 | 164,376 | 433 | 0.505 | 77 | 18 |
| Pin heldr ScPind | 15 | 21,628 | 140 | 0.495 | 20 | 14 |
| Pin heldr Rhod | 83 | 119,677 | 236 | 0.467 | 39 | 17 |
| Spruce forests (Vaccinio-Piceion) | Aconitum lycoctonum ssp. vulparia, Adenostyles glabra, Aquilegia nigricans, Asperula taurina, Caltha palustris, Cardamine trifolia, Cardamine waldsteinii, Chaerophyllum aromaticum, Chaerophyllum hirsutum, Circaea lutetiana, Cirsium waldsteinii, Clematis alpina, Coeloglossum viride, Dryopteris dilatata, Dryopteris expansa, Equisetum sylvaticum, Festuca altissima, Homogyne sylvestris, Hypericum umbellatum, Lamium orvala, Lathraea squamaria, Listera cordata, Lysimachia nemorum, Melampyrum barbatum, Myosotis scorpioides, Omphalodes verna, Petasites albus, Petasites hybridus, Phyteuma spicatum ssp. coeruleum, Poa hybrida, Ranunculus aconitifolius, Ranunculus acris, Ranunculus carinthiacus, Ranunculus platanifolius, Ranunculus serbicus, Rumex alpinus, Scopolia carniolica, Senecio papposus ssp. papposus, Silene alba, Soldanella dimoniei, Stachys dinarica, Symphytum tuberosum agg., Thelypteris limbosperma, Tilia platyphyllos, Trollius europaeus, Vicia oroboides etc. |
| Serbian spruce forests (Piceion omorikae) | Epipogium aphyllum, Equisetum hyemale, Euphorbia glareosa, Hierochloe australis, Petasites kablikianus and Saxifraga tridactylites |
| Scots pine forests (Pinion sylvestris) | Avenula planiculmis, Bupleurum apiculatum, Bupleurum praealtum, Campanula phrygia, Centaurea bracteata, Cerastium fontanum ssp. vulgare, Chrysopogon gryllus, Cirsium grecescui, Coronilla coronata, Coronilla vaginalis, Chamaecytisus polytrichus, Cytisus jankae, Euphorbia serpentini, Festuca bosniaca ssp. pirinensis, Festuca spadicea, Genista carinalis, Inula hirta, Laburnum anagyroides, Lathyrus alpestris, Linum serbicum, Narcissus radiiflorus, Orchis morio, Pedicularis comosa, Poa stiriaca, Potentilla regisborisii, Potentilla rupestris, Primula elatior, Quercus robur, Ranunculus pseudomontanus, Sanguisorba officinalis, Saxifraga blavii, Scleranthus perennis ssp. dichotomus, Stachys betonica, Stellaria palustris, Thlaspi ochroleucum, Veronica vindobonensis, Vicia cassubica etc. |
| Macedonian pine forests (Pinion peucis) | Aconitum burnatii ssp. pentheri, Campanula lanata, Crataegus orientalis, Crocus atticus, Festuca balcanica, Gentiana punctata, Geum reptans, Helleborus cyclophyllus, Jasione orbiculata, Knautia magnifica, Luzula spicata, Pedicularis friderici-augusti, Rhododendron ferrugineum, Senecio abrotanifolius ssp. carpathicus, Soldanella carpatica, Stachys tymphaea, Tozzia alpina, Viola orphanidis etc. |
| Ultramafic pine forests (Orno-Ericion) | Acinos hungaricus, Aristolochia pallida, Artemisia alba, Asplenium adulterinum, Astragalus onobrychis, Bupleurum ranunculoides, Calamagrostis villosa, Calamintha menthifolia, Capsella bursa-pastoris, Cardamine amara, Cardamine plumieri, Centaurea epapposa, Centaurea reichenbachii, Centaurea scabiosa ssp. spinulosa, Chamaecytisus austriacus, Chamaecytisus ciliatus, Chamaecytisus supinus, Cota tinctoria, Cytisus haeufelii, Dianthus sylvestris, Doronicum grandiflorum, Euphorbia glabriflora, Euphorbia gregersenii, Euphorbia montenegrina, Euphorbia virgata, Festuca rupicola, Fumana procumbens, Galium flavescens, Haplophyllum boissieranum, Isatis praecox, Lathyrus bauhinii, Linaria rubioides, Medicago falcata, Medicago prostrata, Ornithogalum umbellatum, Pedicularis brachyodonta, Peucedanum carvifolia, Phleum hirsutum, Phleum montanum, Phleum phleoides, Platanthera chlorantha, Poa chaixii, Podospermum laciniatum, Scleranthus perennis, Silene paradoxa, Stachys recta ssp. baldaccii, Thesium divaricatum, Thymus glabrescens, Tragopogon balcanicus, Viola beckiana etc. |
| Bosnian pine forests (Pinion heldreichii) | Alchemilla glaucescens, Amphoricarpos neumayeri, Anthyllis vulneraria ssp. alpestris, Arabis sudetica, Aster alpinus, Bornmuellera dieckii, Campanula hercegovina, Carlina biebersteinii, Centaurea achtarovii, Crepis dinarica, Draba lasiocarpa, Edraianthus tenuifolius, Festuca penzesii, Genista subcapitata, Gentiana clusii, Helleborus purpurascens, Iberis sempervirens, Leucanthemum chloroticum, Moltkia petraea, Poa macedonica, Poa media, Potentilla clusiana, Potentilla crantzii, Rhinanthus wagneri, Scabiosa graminifolia, Seseli kochii, Silene fabarioides, Thesium auriculatum etc. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ilić, T.; Kuzmanović, N.; Vukojičić, S.; Lakušić, D. Phytogeographic Characteristics of Montane Coniferous Forests of the Central Balkan Peninsula (SE Europe). Plants 2022, 11, 3194. https://doi.org/10.3390/plants11233194
Ilić T, Kuzmanović N, Vukojičić S, Lakušić D. Phytogeographic Characteristics of Montane Coniferous Forests of the Central Balkan Peninsula (SE Europe). Plants. 2022; 11(23):3194. https://doi.org/10.3390/plants11233194
Chicago/Turabian StyleIlić, Tijana, Nevena Kuzmanović, Snežana Vukojičić, and Dmitar Lakušić. 2022. "Phytogeographic Characteristics of Montane Coniferous Forests of the Central Balkan Peninsula (SE Europe)" Plants 11, no. 23: 3194. https://doi.org/10.3390/plants11233194
APA StyleIlić, T., Kuzmanović, N., Vukojičić, S., & Lakušić, D. (2022). Phytogeographic Characteristics of Montane Coniferous Forests of the Central Balkan Peninsula (SE Europe). Plants, 11(23), 3194. https://doi.org/10.3390/plants11233194